九州大学学術情報リポジトリ
Kyushu University Institutional Repository
口腔扁平上皮癌におけるprotease-activated
receptor 1の発現と機能に関する研究 : ΔNp63を介 した上皮間葉転換との関連
服部, 多市
http://hdl.handle.net/2324/4110460
出版情報:九州大学, 2020, 博士(歯学), 課程博士 バージョン:
権利関係:
⼝腔扁平上⽪癌における protease-activated receptor 1 の 発現と機能に関する研究
〜ΔNp63 を介した上⽪間葉転換との関連〜
A study on the expression and functions of protease-activated receptor in oral squamous cell carcinoma: the association with
epithelial-mesenchymal transition mediated by ΔNp63 2020 年 9 ⽉
九州⼤学⼤学院⻭学府
⼝腔顎顔⾯病態学講座 顎顔⾯腫瘍制御学分野
服部 多市 指導教員
九州⼤学⼤学院⻭学研究院
⼝腔顎顔⾯病態学講座 顎顔⾯腫瘍制御学分野
中村 誠司 教授
本研究の⼀部は以下の学術雑誌に掲載された。
Elevated expression of protease-activated receptor 1 via ΔNp63 down-
regulation contributes to nodal metastasis in oral squamous cell carcinoma
Taichi Hattori, Shintaro Kawano, Shoichi Tanaka, Ryota Matsubara, Taiki Sakamoto, Yuma Hashiguchi, Naoki Kaneko, Yurie Mikami, Masahiko Morioka, Yasuyuki Maruse, Ryoji Kitamura, Eiki Hamada,
Megumi Hiwatashi, Kazunari Oobu, Tamotsu Kiyoshima, and Seiji Nakamura
Accepted to International Journal of Oral and Maxillofacial Surgery
略 語 ⼀ 覧
bHLH: basic helix-loop-helix
cDNA: complementary deoxyribonucleic acid(相補的デオキシリボ核酸)
CI: confidence interval(信頼区間)
CK: cytokeratin(サイトケラチン)
DAB: 3,3ʼ-diaminobenzidine(3,3ʼ-ジアミノベンジジン)
DEPC: diethyl pyrocarbonate(ジエチルピロカーボネート)
DMEM: Dulbeccoʼs modified Eagleʼs medium DNA: deoxyribonucleic acid(デオキシリボ核酸)
dNTP: deoxyribonucleotide triphosphate(デオキシリボヌクレオチド三リン酸)
EGFR: epidermal growth factor receptor
EMT: epithelial-mesenchymal transition(上⽪-間葉転換)
ERK: extracellular regulated kinase
FBS: fetal bovine serum(ウシ胎児⾎清)
GAPDH: glyceraldehyde-3-phosphate dehydrogenase H2O2: hydrogen peroxide(過酸化⽔素⽔)
HE: hematoxylin and eosin(ヘマトキシリン−エオジン)
HRP: horseradish peroxidase(⻄洋わさびペルオキシダーゼ)
Ig: immunoglobulin(免疫グロブリン)
KLK: kallikrein-related peptidase MMP: matrix metalloproteinase
M-MuLV : Moloney Murine Leukemia Virus
mRNA: messenger ribonucleic acid(伝令リボ核酸)
NF-κB: nuclear factor-kappa B N.S.: not significant(有意差なし)
OR: odds ratio(オッズ⽐)
OSCC: oral squamous cell carcinoma(⼝腔扁平上⽪癌)
PAR: protease-activated receptor
PBS: phosphate-buffered saline(リン酸緩衝⾷塩⽔)
PCR: polymerase chain reaction(ポリメラーゼ連鎖反応)
PFA: paraformaldehyde(パラホルムアルデヒド)
PGE2: prostaglandin E2 (プロスタグランジン E2)
P/S: penicillin/streptomycin(ペニシリン/ストレプトマイシン)
RNA: ribonucleic acid(リボ核酸)
RT-PCR: reverse transcription polymerase chain reaction(逆転写ポリメラーゼ 連鎖反応)
siRNA: small interfering RNA
TAD: transactivation domain(転写活性領域)
UICC: Union for International Cancer Control(国際対がん連合)
ZEB: zinc-finger E-box binding homeobox
⽬ 次
要旨 6 緒⾔ 9 研究1. OSCC⽣検標本におけるPAR1の免疫組織化学的検討
材料および⽅法 14 結果 19 1-1. OSCCにおけるPAR1の発現と臨床病理組織学的所⾒との関連
1-2. OSCCの頸部リンパ節転移の要因となる臨床病学的因⼦の検索 1-3. OSCC⽣検標本におけるPAR1の発現と予後との関連 1-4. 浸潤先端部におけるPAR1、thrombin、ΔNp63の発現 研究2. OSCC細胞株におけるPAR1の発現と機能に関する研究
材料および⽅法 29
結果 35 2-1. OSCC 細胞株における PAR1、thrombin およびΔNp63 の発現
2-2. ΔNp63 ノックダウンが PAR1 および thrombin の発現に与える影響 2-3. PAR1 ノックダウンによる EMT 関連遺伝⼦の発現変動
2-4. PAR1 ノックダウンが遊⾛能および浸潤能に与える影響
考察 42 謝辞 46 参考⽂献 47
要 旨
近年、癌の浸潤・転移に上⽪-間葉転換(epithelial-mesenchymal transition: EMT)
が深く関与することが明らかとなってきた。これまでに我々は、⼝腔扁平上⽪癌
(oral squamous cell carcinoma: OSCC)の浸潤先端部でΔNp63 の発現が減弱する ことにより EMT が誘導されることを報告した。またその際に、浸潤先端部の癌 細胞で thrombin 受容体である protease-activated receptor(PAR)1 が強く発現 していることを⾒出した。さらに、OSCC 細胞でΔNp63 をノックダウンする と、PAR1 の発現が増強していたことから、癌細胞での EMT 誘導に PAR1 シグ ナルが関与していることが⽰唆された。しかしながら、EMT における PAR1 の 役割やΔNp63 との関わりについての詳細は不明であった。そこで本研究では、
OSCC ⽣検標本における PAR1 の局在を検索し、その発現様式と臨床病理学的 所⾒や予後との関連を明らかにするとともに、ΔNp63 を介した EMT における PAR1 の機能について解析を⾏った。
1. OSCC ⽣検標本における PAR1 の免疫組織化学的検討
OSCC ⽣検標本 116 例における PAR1 の発現を免疫組織化学的に検索した。
まず腫瘍細胞に着⽬して PAR1 の発現を観察すると、発現が全く認められない 症例から、ほとんどすべての細胞で発現しているものまで様々であった。また、
⼀部の症例では癌胞巣を取り囲む間質細胞にも PAR1 の発現を認めた。これら の PAR1 の発現様式から全症例を、Group A: 腫瘍細胞および間質細胞がともに 陰性、Group B: 腫瘍細胞が陰性かつ間質細胞が陽性、Group C: 腫瘍細胞およ
び間質細胞がともに陽性の 3 群に分類した。そこで、PAR1 の発現様式と臨床病 理学的所⾒との関連について検討したところ、Group C は、Group A および B と⽐較して組織学的悪性度の⾼い症例が多く、頸部リンパ節転移の発⽣頻度が 有意に⾼かった。また、頸部リンパ節転移の発⽣要因となり得る因⼦についてロ ジスティック回帰分析による多変量解析を⾏ったところ、PAR1 の発現様式にの み統計学的有意差を認め、Group C では Group A と⽐較して頸部リンパ節転移 の発⽣リスクが有意に⾼かった。疾患特異的累積 5 年⽣存率は Group A が 97.8%、
Group B が 93.3%、Group C が 79.8%であった。さらに、浸潤先端部の腫瘍細 胞における thrombin とΔNp63 の発現を検索したところ、Group A では thrombin の発現は認められなかったが、ΔNp63 は強く発現していた。⼀⽅、
Group C では thrombin の発現は強かったものの、ΔNp63 の発現強度は逆に減 弱しており、PAR1 の発現は ΔNp63 の発現と逆相関していた。
2. OSCC 細胞株における PAR1 の発現と機能に関する研究
本研究では、5 種類の OSCC 細胞株(低転移株: HSC-2、HSC-3、SQUU-A、
SAS、⾼転移株: SQUU-B)とヒト正常⾓化上⽪細胞株(HaCaT)を⽤いた。ま た、SQUU-B 細胞にΔNp63βを過剰発現させた SQUU-BO 細胞と empty vector を導⼊した SQUU-BC 細胞も使⽤した。まずΔNp63、PAR1 および thrombin の発現を RT-PCR 法にて検索したところ、SQUU-B 細胞ではΔNp63 の発現は 低く、PAR1 と thrombin の発現は⾼かった。⼀⽅、低転移株と SQUU-BO 細胞 ではΔNp63 の発現は⾼いものの PAR1 と thrombin の発現は低かった。そこで、
SQUU-A 細胞を⽤いてΔNp63 をノックダウンすると、PAR1 と thrombin の発 現が増強した。さらに、SQUU-B 細胞で PAR1 をノックダウンしたところ、上
⽪系マーカーである E-cadherin、cytokeratin(CK)5、CK14 の発現量増加、間 葉系マーカー(vimentin、N-cadherin、fibronectin)および EMT 関連因⼦(ZEB1、
ZEB2、snail、slug、twist)の発現量減少を認め、遊⾛能と浸潤能が著明に抑制 された。
以上の結果から、ΔNp63 の発現減弱により、PAR1 シグナルが活性化するこ とで EMT が誘導され、OSCC の運動能が亢進するものと考えられた。
緒 ⾔
⼝腔扁平上⽪癌(oral squamous cell carcinoma: OSCC)は⼝腔粘膜に由来す る悪性腫瘍で、⼝腔癌全体の約 90%以上を占めている(1-3)。OSCC に対する 治療法は⼿術が第⼀選択であり、切除断端陽性や頸部リンパ節多発転移などの 術後再発ハイリスク症例に対しては、⼿術施⾏後に放射線化学療法を⾏うこと が⼀般的である(4-6)。最近では、最も根治が困難な遠隔転移症例に対し、抗 epidermal growth factor receptor(EGFR)抗体である cetuximab や抗 PD-1 抗 体の nivolumab などの分⼦標的薬が臨床応⽤されているが、治療効果が得られ ない場合も多く、⽣存率は⼤きくは改善されていないのが現状である(7-9)。こ のように、OSCC の治療を困難にしている最⼤の要因は転移であり、頸部リン パ節転移や遠隔転移の制御は⽣存率の向上に⼤いに貢献するものと考えられる (10)。⼀般に転移は、原発巣で増殖した癌細胞が粘膜下層にある⾎管やリンパ管 に浸潤することにより⽣じる。その後、癌細胞が標的臓器に定着後、細胞増殖を 開始し転移が成⽴する(11-15)。これらのプロセスには、さまざまな遺伝⼦や 分⼦機構が関与していると考えられているが、その詳細なメカニズムの解明に は⾄っていない。
近年、癌の転移に上⽪‒間葉転換(epithelial-mesenchymal transition:EMT)
が関与していることが明らかになってきた (16-18)。EMT は、1982 年に Hay が提唱した概念で、胎児の発⽣期や創傷治癒の過程で観察され、上⽪細胞が形態 的および機能的に間葉系細胞様に変化する現象である (19.20)。EMT が誘導さ
れた上⽪細胞は、上⽪系マーカーの発現量減少、間葉系マーカーの発現量増加、
細胞極性や細胞接着の喪失、遊⾛能の亢進および増殖能の低下を⽰す(21.22)。
すなわち、EMT が誘導された癌細胞は、細胞間の接着が喪失することにより原 発巣から離脱し、遊⾛能が亢進することで標的臓器への転移を容易にしている。
この概念が提唱されてから、癌細胞の EMT に関与する遺伝⼦が徐々に明らかに なってきており、zinc-finger 型転写因⼦である snail、slug、zinc-finger E-box binding homeobox (ZEB)1、ZEB2 や basic helix-loop-helix(bHLH)型転写 因⼦である twist などが知られている(17.23.24)。これらの転写産物は、上⽪
の細胞間接着分⼦である E-cadherin などの発現を抑制することで、細胞間接着 を減弱させるとともに、vimentin や N-cadherin などの間葉系マーカーの発現を 増強させ、強⼒に EMT を誘導するとされている(24-26)。このように、個々 の EMT 関連遺伝⼦については解明されつつあるが、癌が浸潤する際にこれらの 発現がどのように制御されているかは未だ不明な点が多い。
そこで我々は、癌細胞のEMTを制御する分⼦の1つとしてp53のホモログで ある転写因⼦のp63に着⽬した。p63はプロモーター領域の違いにより、N末端 側に転写活性領域(transactivation domain: TAD)を有するTAp63とTADが⽋
損したΔNp63の2つのアイソフォームに分けられる(27-30)。p53およびTAp63 は、細胞の⽼化やDNA障害に伴うアポトーシス亢進および細胞増殖の抑制に関 与するのに対し、ΔNp63はp53およびTAp63の転写領域に結合し、標的遺伝⼦
の発現をドミナントネガティブに抑制する(31.32)。また、C末端側での選択的 スプライシングにより、それぞれ3つのバリアント(α,β,γ)が⽣成され、そ れぞれ異なる機能を有する。Jaimeらは、EMTの誘導に関してΔNp63αとΔ
Np63βは抑制的に働き、ΔNp63γは促進的に機能すると報告している(33)。
これらの知⾒は、OSCCが浸潤・転移する過程でΔNp63がoncoproteinとして機 能し、EMTの誘導に何らかの役割を担っている可能性を⽰している(34-38)。
そのためわれわれは、特にΔNp63に焦点を絞り、OSCCでの発現と機能につい て検討を⾏った。
先⾏研究において、OSCC⽣検標本を⽤いΔNp63の発現を免疫組織学的に検 索したところ、ほとんど全てのOSCC細胞でΔNp63の発現を認めた。興味深い ことに、⼀部の症例において腫瘍中⼼部から浸潤先端部に向かうにつれてΔ Np63の発現強度が減弱しており、それらの症例では頸部リンパ節転移の発⽣頻 度が有意に⾼かった(39.40)。このことから、ΔNp63の発現減弱は転移と何 らかの関連があると考えられたため、OSCC細胞株を⽤いてさらに機能解析を
⾏った。その結果、ΔNp63のノックダウンによりセリンプロテアーゼの1つで あるkallikrein-related peptidase(KLK)6とその受容体であるprotease-activated receptor(PAR)2の発現が減弱するとともに、そのサブタイプの1つである PAR1の発現が増強されることによりEMTが誘導されていることを⾒出した
(41)。これらの結果より、ΔNp63の発現減弱を介したEMT誘導には、発現 が増強していたPAR1シグナルが重要であることが⽰唆された。
PARは、Gタンパク共役7回膜貫通型受容体の1つで、⾎管内⽪細胞、上⽪細 胞、⾎⼩板などで発現していることが知られている。PARの細胞外領域のN末 端ペプチド配列がプロテアーゼにより特定部位で切断され、切断末端に受容体 活性化領域が露出し、その領域がtethered ligandとして受容体の別部位と結合 するとPARが活性され、細胞内へシグナルが伝達される(42-46)。PARは、現
在までにPAR1、PAR2、PAR3、PAR4の4種のサブタイプがクローニングされ ており、特にPAR1はthrombin、matrix metalloproteinase(MMP)およびprotein Cにより活性化されることが明らかとなっている(47.48)。正常組織における PAR1の活性化はさまざな⽣理的機能に関与しており、例えば⾎⼩板ではPAR1 の活性化により⾎⼩板の凝集反応を起こすことが知られている(47)。また、
胃粘膜上⽪細胞においては、ムチンやprostaglandin E2(PGE2)の分泌に関わっ ていることが報告されている(49)。また最近の研究によれば、PAR1は乳癌、
卵巣癌、膵臓癌、前⽴腺癌、肺癌などのさまざまな悪性腫瘍でも発現しており、
PAR1⾼発現症例では転移の発⽣率が⾼く、予後が不良であることが知られて いる(50-56)。また、⾼浸潤性乳癌細胞株では、低浸潤性細胞株と⽐較し、PAR1 の発現が⾼いことが報告されている(50)。以上より、PAR1は癌の浸潤・転移 機構において何らかの重要な機能を果たしていると考えられるが、EMTにおけ るPAR1の役割やΔNp63との関わりについての詳細は不明なままである。
そこで本研究では、OSCC⽣検標本におけるPAR1の局在を検索し、その発現 様式と臨床病理学的所⾒や予後との関連を明らかにするとともに、ΔNp63を介 したEMTにおけるPAR1の機能について解析を⾏った。
研究1. OSCC⽣検標本におけるPAR1の免疫組織化学的検討
材料および⽅法
1-1. 対象患者
2011 年 1 ⽉から 2016 年 12 ⽉に九州⼤学病院顎⼝腔外科を受診し、病理組織 学的に OSCC と診断され、⼿術による根治的治療を⾏った 116 名(男性 70 名、
⼥性 46 名、平均年齢 65.9±12.0 歳〈30〜91 歳〉)を対象とした(表 1)。
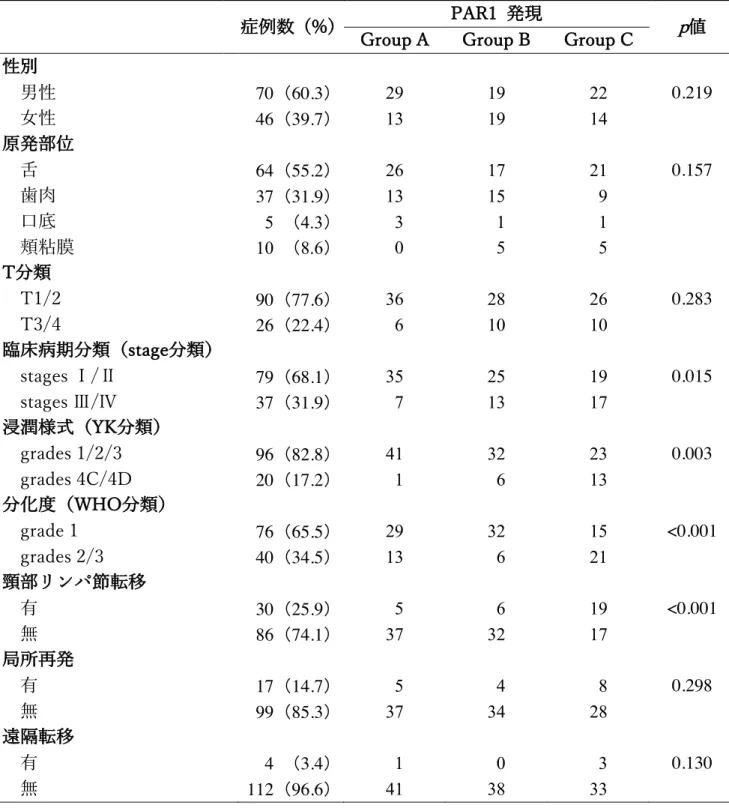
表 1. OSCC 患者の内訳
症例数 (%)
性別
男性 70 (60.3)
⼥性 46 (39.7)
原発部位
⾆ 64 (55.2)
⻭⾁ 37 (31.9)
⼝底 5 (4.3)
頬粘膜 10 (8.6)
T分類
T1/2 90 (77.6)
T3/4 26 (22.4)
臨床病期(stage分類)
Ⅰ/Ⅱ 79 (68.1)
Ⅲ/Ⅳ 37 (31.9)
組織学的悪性度
分化度(grade分類)
grade 1 76 (65.5)
grades 2/3 40 (34.5)
浸潤様式(YK分類)
grades 1/2/3 96 (82.8)
grades 4C/4D 20 (17.2)
1-2. OSCCのステージングと⽣検組織の病理組織学的診断
本研究では、国際対がん連合(Union for International Cancer Control: UICC)
の「TNM分類 第8版」をもとにstage分類を⾏った(57)。
OSCC の組織学的悪性度分類として、grade 分類(WHO 分類)と⼭本・⼩浜 の分類(YK 分類)を⽤いた。grade 分類により、腫瘍細胞の分化度に応じて grade 1(⾼分化型)、grade 2(中分化型)、および grade 3(低分化型)に分けた。
また、YK 分類により、腫瘍宿主境界部における浸潤様式に基づいて grade 1、
grade 2、grade 3、grade 4C、および grade 4D に分類した(58)。
YK 分類の基準を以下に⽰す。
grade 1: 腫瘍と宿主の境界が明瞭である。
grade 2: 境界線にやや乱れがある。
grade 3: 境界線は不明瞭で⼤⼩の癌胞巣が散在している。
grade 4C: 境界線は不明瞭で⼩さな癌胞巣が索状に浸潤している。
grade 4D: 境界線は不明瞭で癌は胞巣を作らず、び漫性に浸潤している。
1-3. 免疫組織化学的染⾊法
OSCC 患者より採取した⽣検および切除標本は、直ちに 4%パラホルムアルデ ヒド(paraformaldehyde: PFA)に 24〜48 時間浸漬固定し、パラフィン包埋を
⾏った。回転式ミクロトーム RM2145(Leica Mycrosystems, Tokyo, Japan)に て 4 μm の切⽚を作製し、ヘマトキシリン−エオジン(hematoxylin and eosin:
HE)染⾊および免疫組織化学的染⾊に⽤いた。作製したパラフィン切⽚をキシ
レンに 20 分間、さらに 100%、95%、85%、75%エタノールの順にそれぞれ各 5 分間浸漬させ、脱パラフィン処理および⽔和処理を⾏った。切⽚を精製⽔にて
⽔和したのちに、Target Retrieval Solution(pH 9.0, Dako, Glostrup, Denmark)
を⽤いて抗原賦活化処理(121℃、5 分間)を⾏った。30 分間室温で放冷したの ち、切⽚をリン酸緩衝⽣理⾷塩⽔(phosphate buffered saline: PBS; 137 mM NaCl、
2.7 mM KCl、8.1 mM Na2PO4、1.5 mM KH2PO〈Nacalai Tesque, Kyoto, Japan〉)4
にて洗浄した。この後、内因性ペルオキシダーゼ除去のため、3%過酸化⽔素⽔
を室温で 15 分間反応させた。再度、PBS で洗浄したのち、抗体の⾮特異的吸着 を防ぐために Blocking One(Nacalai Tesque)を 30 分間反応させた。⼀次抗体
(表 2)は 4℃で⼀晩反応させた。PBS にて洗浄後、⼆次抗体として horseradish peroxidase(HRP)標識抗マウス immunoglobulin(Ig)G ポリクロール抗体(ヒ ストファイン MAX-PO; Nichirei Bioscience, Tokyo, Japan)を⽤い、室温で 1 時 間反応させた。PBS にて洗浄後、ジアミノベンジジン(3,3ʼ-diaminobenzidine:
DAB)(Nichirei Bioscience)にて可視化し、ヘマトキシリン(Mayerʼs hemalum solution: Millipore, Burlington, Massachusetts)を⽤いて対⽐染⾊を⾏った。そ の後、切⽚を 75%、85%、95%、100%エタノールに各 5 分間、キシレンに 10 分 間 浸 漬 さ せ 、 脱 ⽔ お よ び 透 徹 処 理 を ⾏ い 、 Malinol mounting medium
(MutoPure Chemicals, Tokyo, Japan)を⽤いて封⼊した。陰性対照として、⼀
次抗体の代わりに PBS を⽤いた。観察には、顕微鏡⽤デジタルカメラ(DP80;
OLYMPUS, Tokyo, Japan)を使⽤した。また、切⽚中の浸潤先端部より 0.5 mm2 の範囲を無作為に 3 か所選択して、腫瘍細胞および間質細胞の PAR1 の発現様 式をもとに全症例を以下の 3 群に分類した。Group A : 腫瘍細胞および間質細
胞がともに陰性、Group B : 腫瘍細胞が陰性かつ間質細胞が陽性、Group C : 腫 瘍細胞および間質細胞がともに陽性。PAR1 の発現様式と臨床病理組織学的所⾒
との関連について統計学的に解析した。PAR1 の発現様式と予後との関連性につ いて検討するために、Kaplan-Meier 法を⽤いて⽣存曲線を作成し、疾患特異的 累積⽣存率を算出した。さらに、浸潤先端部の腫瘍細胞におけるΔNp63 発現減 弱の有無と PAR1 の発現様式との関連について統計学的に分析した。
表 2. 免疫組織化学的染⾊に使⽤した⼀次抗体
抗体(clone 名、製造会社、所在地) アイソタイプ 希釈倍率
抗ヒト PAR1 マウスモノクローナル抗体
(sc-13503, Santa Cruz Biotechnology,Dallas, Texas) IgG 1 : 100 抗ヒトΔNp63 マウスモノクローナル抗体
(ab172731, Abcam, Cambridge, UK) IgG 1 : 100 抗ヒト thrombin マウスモノクローナル抗体
(ab17199, Abcam) IgG 1 : 100
1-4. 統計学的解析
OSCC ⽣検組織における PAR1 の発現と病理組織学的所⾒との関連および浸 潤先端部におけるΔNp63 の発現減弱の有無と PAR1 発現様式との関連につい ては、カイ⼆乗検定を⽤いて統計処理を⾏った。さらに、頸部リンパ節転移に関 連する因⼦を検索するため、ロジスティック回帰分析による多変量解析を⾏っ た。⽣存曲線は Kaplan-Meier 法を⽤いて解析を⾏い、統計処理には log-rank 検 定を⽤いた。なお、統計ソフトとして JMP software version 13(SAS Institute, Cary, North Carolina)を使⽤し、p<0.05 の場合を統計学的に有意差ありとした。
結 果
研究1. OSCC⽣検標本におけるPAR1免疫組織化学的検討
1-1. OSCCにおけるPAR1の発現と臨床病理組織学的所見との関連
OSCC ⽣検標本における PAR1 の発現を検索するため、免疫組織化学的染⾊
を⾏った。まず腫瘍細胞に着⽬して PAR1 の発現を観察すると、発現が全く認 められない症例から、ほとんどすべての腫瘍細胞で発現しているものまで様々 であった。興味深いことに、⼀部の症例では、癌胞巣を取り囲む間質細胞にも PAR1 の発現を認めた。また、腫瘍細胞および間質細胞では、腫瘍中⼼部から浸 潤先端部にかけて PAR1 の発現が強くなっていた(図 1)。そのため、浸潤先端 部の腫瘍細胞と間質細胞における PAR1 の発現様式から全症例を 3 群に分類し て、臨床病理組織学的所⾒との関連を検討した。その結果、Group C は Group A および B と⽐較して、stage Ⅲ/Ⅳなどの局所進展例が多く、浸潤様式が grade 4C/4D や分化度が低い症例、すなわち組織学的悪性度の⾼い症例が多かった。
また、Group C では頸部リンパ節転移の発⽣頻度が有意に⾼かった。しかし、
性別、部位、T 分類、局所再発の有無および遠隔転移の有無に関して、3 群間に 統計学的有意差は認められなかった(表 3)。
0sODH1OrAmepluBA,BcoA.BA,.BlAB1OmepluAB:KPcabaLS1OrFRCAB:KPca1OrPRCFbaP1OrRAB:KPcabaLS1OrRcaDbaDCJcoEEHJ1OrFGIJC5 HEPAR1 Group A
Group C Group B
AB
IJ EF HEPAR1
CD
GH
KL co
表 3. OSCC ⽣検標本における PAR1 発現と臨床病理組織学的所⾒との関連
カイ⼆乗検定
症例数 (%) PAR1 発現
p値 Group A Group B Group C
性別
男性 70 (60.3) 29 19 22 0.219
⼥性 46 (39.7) 13 19 14
原発部位
⾆ 64 (55.2) 26 17 21 0.157
⻭⾁ 37 (31.9) 13 15 9
⼝底 5 (4.3) 3 1 1 頬粘膜 10 (8.6) 0 5 5
T分類
T1/2 90 (77.6) 36 28 26 0.283 T3/4 26 (22.4) 6 10 10
臨床病期分類(stage分類)
stages Ⅰ/Ⅱ 79 (68.1) 35 25 19 0.015 stages Ⅲ/Ⅳ 37 (31.9) 7 13 17
浸潤様式(YK分類)
grades 1/2/3 96 (82.8) 41 32 23 0.003 grades 4C/4D 20 (17.2) 1 6 13
分化度(WHO分類)
grade 1 76 (65.5) 29 32 15 <0.001 grades 2/3 40 (34.5) 13 6 21
頸部リンパ節転移
有 30 (25.9) 5 6 19 <0.001
無 86 (74.1) 37 32 17
局所再発
有 17 (14.7) 5 4 8 0.298
無 99 (85.3) 37 34 28
遠隔転移
有 4 (3.4) 1 0 3 0.130
無 112 (96.6) 41 38 33
1-2. OSCCの頸部リンパ節転移の要因となる臨床病理学的因子の検索
1-1 の結果より、PAR1 の発現は病期分類、浸潤様式、分化度、頸部リンパ節 の発⽣頻度と有意に相関していた。そこで、頸部リンパ節転移の要因となり得る これらの臨床病理学的因⼦が、相互に影響を及ぼし合っていないかを検討する ために、ロジスティック回帰分析による多変量解析を⾏った。その結果、PAR1 の発現様式にのみ統計学的有意差を認め、Group C では Group A と⽐較して頸 部リンパ節転移の発⽣リスクが有意に⾼かった(オッズ⽐: 5.56)(表 4)。
表4. OSCCの頸部リンパ節転移の要因となる臨床病理学組織学的因⼦の解析
OR: odds ratio(オッズ⽐)95% CI: 95% confidence interval(95%信頼区間)
頸部リンパ節
転移 単変量解析 多変量解析
有 無 OR(95% CI) p値 OR(95% CI) p値
性別
男性 17 53 Reference Reference
⼥性 13 33 1.23(0.53-2.85) 0.633 1.46(0.56-3.89) 0.444 T 分類
T1/2 23 67 Reference Reference
T3/4 7 19 1.07(0.38-2.80) 0.889 0.96(0.30-2.88) 0.948 浸潤様式(YK 分類)
grades 1/2/3 19 77 Reference Reference
grades 4C/4D 11 9 4.95(1.80-14.01) 0.002 2.58(0.75-9.06) 0.132 分化度(WHO 分類)
grade 1 14 62 Reference Reference
grades 2/3 16 24 2.95(1.26-7.06) 0.013 1.50(0.51-4.26) 0.456 PAR1 発現
Group A 5 37 Reference Reference
Group B 6 32 1.39(0.38-5.23) 0.615 1.20(0.30-4.87) 0.790 Group C 19 17 8.27(2.64-25.87) <0.001 5.56(1.70-20.56) 0.004
1-3. OSCC生検標本におけるPAR1の発現と予後との関連
次に、OSCC ⽣検標本における PAR1 の発現と予後との関連を検討するため、
Kaplan-Meier 法を⽤いて⽣存分析を⾏った。その結果、腫瘍細胞における PAR1 陽性群は、陰性群と⽐較し疾患特異的 5 年累積⽣存率が有意に低かった(陽性 群: 79.8%、陰性群: 95.4%)。⼀⽅、間質細胞における PAR1 陽性群と陰性群と の間では有意差は認められなかった(陽性群: 86.5%、陰性群: 97.6%)。PAR1 の発現様式別では、Group C の⽣存率が最も低かったが、有意差は認められな かった(Group A: 97.6%、Group B: 93.3%、Group C: 79.8%)(図 2)。
図2. 疾患特異的5年累積⽣存率
(a)腫瘍細胞におけるPAR1陽性群は陰性群と⽐較して、有意に予後不良である。(b)
間質細胞におけるPAR1陽性群と陰性群との間に有意差は認められない。(c)Group C はGroup A、Group Bと⽐較し、予後不良である。なお、統計処理にはlog-rank検定を⽤
いている。*p < 0.05。
1-4. 浸潤先端部におけるPAR1、thrombin、ΔNp63の発現
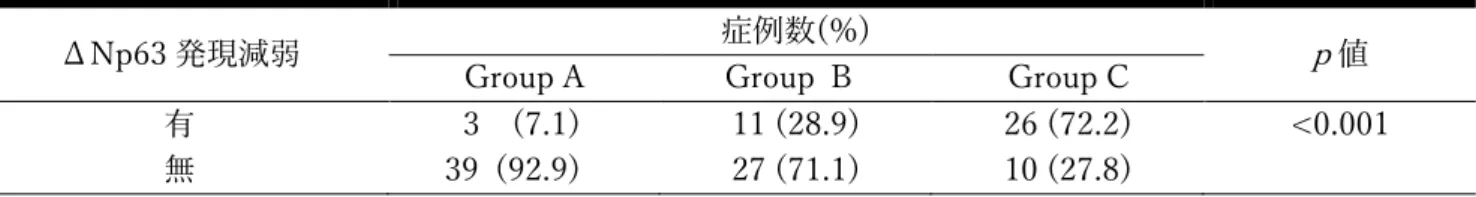
PAR1 は thrombin により活性化されることが知られているため、OSCC にお ける thrombin を免疫組織学的に検索した。また、ΔNp63 についても同様に検 索を⾏った。Group A では、thrombin は腫瘍細胞で発現を認めなかったのに対 して、ΔNp63 は強く発現していた。⼀⽅、Group C では、thrombin は PAR1 の発現様式と同様に腫瘍細胞で強く発現していたが、ΔNp63 の発現強度は逆に 減弱していた(図 3)。さらに、ΔNp63 の発現減弱は、Group A で 3 例(7.1%)、
Group B で 11 例(28.9%)、および Group C で 26 例(72.2%)に認められ、
腫瘍細胞における PAR1 の発現は、ΔNp63 の発現減弱と有意に関連していた
(p <0.001)(表 5)。
図 3. OSCC ⽣検標本の浸潤先端部における PAR1、ΔNp63 および thrombin の発現
(免疫組織化学的染⾊法)
(A, E)HE 染⾊。(B, F)PAR1。(C, G)ΔNp63。(D, H)thrombin。Group A で は、thrombin は腫瘍細胞に発現を認めないが、ΔNp63 の発現は強く発現している。
Group C では thrombin は腫瘍細胞で強く発現しているが、ΔNp63 の発現強度は減弱 している。scale bars: 50 μm。
表 5. OSCC 浸潤先端部における ΔNp63 の発現減弱の有無と PAR1 発現様式との関連
カイ⼆乗検定
ΔNp63 発現減弱 症例数(%)
p値 Group A Group B Group C
有 3 (7.1) 11 (28.9) 26 (72.2) <0.001 無 39 (92.9) 27 (71.1) 10 (27.8)
研究2. OSCC細胞株におけるPAR1の発現と機能に関する
研究
材料および⽅法
2-1. 細胞培養
本研究では、OSCC 細胞株である HSC-2(⾼分化型⼝底癌由来)、HSC-3(低 分化型⾆癌由来)、SAS(低分化型⾆癌由来)、SQUU-A(⾼分化型⾆癌由来:
低転移株)、SQUU-B(⾼分化型⾆癌由来:⾼転移株)ならびに HaCaT(ヒト 正常⾓化上⽪由来)を⽤いた。また、SQUU-B 細胞に ΔNp63β を強制発現さ せた SQUU-BO 細胞および empty vector を導⼊した SQUU-BC 細胞も使⽤し た (52)。培地には、Dulbeccoʼs modified Eagleʼs medium(DMEM)/F12(Life Technologies, Carlsbad, California)に 10%ウシ胎児⾎清(fetal bovine serum:
FBS; Life Technologies)、100 units/ml のペニシリン/ストレプトマイシン
(penicillin/streptomycin: P/S; Nacalai Tesque)を添加したものを⽤いて、37℃、
5% CO2存在下で細胞培養を⾏った。
2-2. RNA(ribonucleic acid)の抽出および相補的DNA(complementary deoxyribonucleic acid: cDNA)の合成
RNA の抽出では、まず培養⽫上の細胞に TRIzol®(Thermo Fisher Scientific, Waltham, Massachusetts)を加え、セルスクレーパーにて細胞を破砕した。その 後、これらに 0.2 mL の chloroform(Nacalai Tesque)を加えて撹拌し、4℃、
14,000 rpm で 15 分間遠⼼分離を⾏った後、RNA を含む⽔層を採取し、70%エ タ ノ ー ル と 混 和 し た 。 RNeasy® Lipid Tissue Mini Kit ( QIAGEN, Hilden, Germany)を⽤い、混和した溶液を RNeasy スピンカラムに⼊れ、20℃、10,000
rpm で 30 秒間遠⼼分離した。その後、Buffer RW1(20%エタノール、グアニジ ン塩含有)を 0.7 mL 添加し、20℃、10,000 rpm で 15 秒間遠⼼分離をかけた。
同じ作業を再度⾏った後、Buffer RPE(80%エタノール、塩化ナトリウム含有)
を添加し、20℃、10,000 rpm で 15 秒間遠⼼分離を⾏なった。RNeasy スピンカ ラムを新しい 1.5 mL コレクションチューブに置き換え、RNase free water 30 μL を直接スピンカラムメンブレンに添加し、20℃、10,000 rpm で 1 分間遠⼼
分離を⾏い、RNA を抽出した。その後、吸光度計 NANO DROP 1000(Thermo Fisher Scientific)にて total RNA の濃度を測定した。
cDNA の合成には、diethylpyrocarbonate(DEPC)処理⽔に約 2.0 μg の total RNA、35 units/μm RNase inhibitor(Nacalai Tesque)を 1.0 μl、100 mM Tris- HCl(pH8.8)、500 mM KCl および 0.8% Nonidet P40 を含む 10×Taq DNA polymerase buffer(Bio Basic, Markham, Canada)を 2.0 μl、25 mM MgCl2(Bio Basic)を 4.0 μl、2.0 mM deoxyribonucleotide triphosphate(dNTP)mix(Toyobo, Osaka, Japan)を 2.0 μl、50 μM random hexamers(Invitrogen, Carlsbad, California)を 1.0 μl、50 units/μl Moloney Murine Leukemia Virus (M-MuLV)
Reverse Transcriptase(Applied Biosystems, Foster City, California)を 1.0 μl 加 えて、合計 20.0 μl とし、42℃で 15 分間インキュベートした。その後、99℃で 5 分間加温して酵素を失活させ、5℃にて 5 分間冷却し、これを messenger RNA
(mRNA)の発現解析に⽤いた。
2-3. reverse transcription(RT)‒polymerase chain reaction(PCR)および real-time PCR による mRNA の発現解析
RT-PCR は滅菌⽔に template DNA を 50 ng、DreamTaq buffer を 1.0 μl、
Dream Taq polymerase(Thermo scientific)を 0.05 μl、2.0 mM dNTPmix を 1.0 μl、20 μM センスおよびアンチセンスプライマーをそれぞれ 0.5 μl 加えて 全反応量を 9.55 μl とした。反応条件は、熱変性は 94℃で 1 サイクル⽬が 3 分 間、2 サイクル⽬以降は 30 秒間で⾏い、アニーリングは 30 秒間、伸⻑反応は 72℃で 15 秒間とした。
real-time PCR は Brilliant Ⅲ Ultra-Fast SYBR Green QPCR Master Mix
(Agilent Technologies, Santa Clara, California)を⽤いて⾏った。滅菌⽔に Master Mix を 10 μl、template DNA を 9 ng、20 μM センスおよびアンチセン スプライマーをそれぞれ 0.5 μl 加え、全反応量を 20 μl とした。反応条件は、
熱変性は 94℃で 1 サイクル⽬が 10 分間、2 サイクル⽬以降は 30 秒間で⾏い、
アニーリングは 30 秒間、伸⻑反応は 72℃で 15 秒間とし、全 40 サイクルの増 幅を⾏った。また、mRNA 発現量を定量化するためにハウスキーピング遺伝⼦
である glyceraldehyde-3-phosphate dehydrogenase(GAPDH)を⽤いて補正し、
ΔΔCt 法により相対的発現量を算出した。なお、得られた PCR 産物を 2%アガ ロースゲル(Nacalai Tesque)上で電気泳動を⾏い、エチジウムブロマイド溶液
(AMRESCO, Solon, Ohio)を⽤いて染⾊し、紫外線により可視化した。今回解 析を⾏った遺伝⼦の各プライマー配列、PCR 産物の断⽚サイズおよびアニーリ ング温度(表 6)を⽰す。上記操作を独⽴して 3 回⾏い、統計処理を⾏った。
表6. RT-PCR のプライマーとPCR 産物の断⽚サイズ
mRNA
PCR 産物 のサイズ
(bp)
プライマー塩基配列 アニーリング
温度 (℃)
∆Np63 117 forward TGCCCAGACTCAATTTAGTGAG reverse TGCGCGTGGTCTGTGTTATA 60
PAR1 240 forward TCCGGATAT TTGACCAGCTC reverse AGACCCAAACTGCCAATCAC 60
thrombin 115 forward AGAGCCCCTTTAACAACCGC reverse AGCTGCCCATGAATAGCACTG 60
E-cadherin 196 forward TGCTCTTCCAGGAACCTCTG reverse AGGGAAACTCTCTCGGTCCA 60
cytokeratin 5 207 forward GGTTGATGCACTGATGGATG reverse TACCAGGACTCGGCTTCTGT 60
cytokeratin 14 135 forward ATCCTGCTGGACGTGAAGAC reverse GGAGGAGGTCACATCTCTGG 60
vimentin 196 forward TGCCCTTAAAGGAACCAATG reverse CTCAATG TCAAGGGCCATCT 60
N-cadherin 163 forward TGAAGGAGTCAGCAGAAGTTGA reverse TCAGACCTGATCCTGACAAGC 60
fibronectin 170 forward GCAAGCCCATAGCTGAGAAG reverse GTCCTGATCGTTGCATCTATTT C 60
ZEB1 118 forward AAGACATGTGACGCAGTCTGG reverse TGGCTTCTCTCCACTGTGAATTC 60
ZEB2 211 forward AGCGGAAACAAGGATTTC reverse GGTCTTTTTCCTGTGTGTTCG 60
snail 231 forward GCAGGACTCTAATCCAGAGTTTACC reverse CCTTTCCCACTGTCCTCATC 60
slug 224 forward CACATACAGTGATTATTTCCCCG reverse GTCCTTGGAGGAGGTGTCAG 60
twist 201 forward GGAGTCCGCAGTCTTACGAG reverse TCTGGAGGACCTGGTAGAGG 60
GAPDH 104 forward ATCAGCAATGCCTCCTGCAC reverse ATGGCATGGACTGTGGTCAT 60
2-4. PAR1 small interfering RNA (siRNA)の導⼊
PAR1 を⾼発現している SQUU-B 細胞に PAR1 siRNA を導⼊し、機能解析を
⾏った。なお、導⼊には Dhama FECT 1 Transfection Reagent(Dharmacon, Lafayette, Colorado)を使⽤した。まず、6 ⽳プレートに 2.5×104 /well の SQUU- B 細胞を播種し、37℃、5%CO2存在下で 24 時間培養後、FBS 含有の DMEM/F- 12 に培地を交換し、10 nM の PAR1 siRNA(DhamaFECT)を導⼊した。37℃、
5% CO2存在下で培養した後、導⼊後 24 時間後の mRNA を含む total RNA を 抽出し、発現解析を⾏った。なお siPAR1 は、ON-TARGETplus Human F2R (2149) siRNA - SMARTpool(DhamaFECT)を、陰性対象には ON-TARGETplus Non-targeting siRNA(DhamaFECT)を導⼊した。
2-5. wound healing assay
PAR1 が OSCC 細胞の遊⾛能に与える影響を検討するため、wound healing assay を⾏った。まず、24 ⽳プレートに 2.5×104 /well の PAR1 ノックダウン細 胞(siPAR1)およびコントロール細胞(siCtrl)を播種し、37℃、5% CO2存在 下でコンフルエントになるまで培養した。コンフルエントの状態で 200 μl ⽤滅 菌ピペットチップ(Greiner Bio-One, Kremsmünster, Austria)の先端を⽤いて 間隙形成を⾏った。また、細胞増殖による影響を回避するため、10 μg/ml の mitomycin C を間隙形成後より 2 時間作⽤させた。その後培地を交換し、間隙 形成 24 時間後の遊⾛状態を倒⽴顕微鏡(対物レンズ ×4, CKX41 NB-31PHP;
Olympus, Tokyo, Japan)にて観察した。
2-6. MatrigelTM invasion assay
PAR1 のノックダウンが OSCC 細胞の浸潤能に与える影響を検討するため、
Falcon Cell Culture Insert(BD Biosciences, Franklin Lakes, New Jersey)、
BiocoatTM MatrigelTM invasion chamber(BD Biosciences)および 24 ⽳プレート を⽤いて、MatrigelTM invasion assay を⾏った。まず、PAR1 ノックダウン細胞
(siPAR1)およびコントロール細胞(siCtrl)を 37℃、5% CO2存在下で 70%コ ンフルエントまで培養した後、無⾎清の DMEM/F12 に培地交換し、37℃、5%
CO2 存在下でさらに 6 時間培養した。底⾯に細胞の移動が可能な 8 μm のポア が空いたインサート内へ、無⾎清培地で懸濁した 2.5×104 /well の各々の細胞を 播種し、プレート内へは⾎清添加培地を⼊れることで細胞の遊⾛を促した。これ を同条件下に 22 時間培養後、インサート下⾯を 100%メタノール(Junsei, Tokyo, Japan)にて固定し、HE 染⾊した後に、0.5 mm2の範囲を無作為に 3 か所選択 して染⾊された細胞数の平均値を算出した。また、別のインサートには浸潤細胞 のみインサート下⾯に移動するように、MatrigelTMをコーティングし、同様の⽅
法で、浸潤細胞数の平均値を算出した。さらに、平均浸潤細胞数を平均遊⾛細胞 数で除して百分率で表したものを浸潤細胞率(%)として解析した。
2-7. 統計学的解析
real-time PCR 法、wound healing assay、invasion assay の各群間の⽐較に Mann-Whitney U検定を⽤いて統計処理を⾏った。なお、統計ソフトとして JMP software version 13(SAS Institute)を使⽤し、p<0.05 の場合を統計学的に有意 差ありとした。
結 果
研究2. OSCC細胞株におけるPAR1の発現と機能に関する研究 2-1. OSCC 細胞株におけるPAR1、thrombin およびΔNp63 の発現
研究 1 より、OSCC 浸潤先端部で PAR1 および thrombin の発現は強く、Δ Np63 の発現は減弱していた。そこで OSCC 細胞においても同様の傾向がある かを RT-PCR 法にて検索した。
⾼転移株 SQUU-B 細胞では、ΔNp63 の発現は低く、PAR1 と thrombin の発 現は⾼かった。⼀⽅、低転移株や SQUU-BO 細胞では、ΔNp63 の発現が⾼い ものの、PAR 1 と thrombin の発現は低かった。このように、浸潤先端部の発現 様式と同様に、OSCC 細胞株においても PAR1 および thrombin の発現は、Δ Np63 の発現と逆相関していた(図 4)。
図4. OSCC 細胞株ならびにヒト正常⾓化上⽪細胞におけるΔNp63、PAR1および thrombinの発現(RT-PCR法)
⾼転移株 SQUU-B 細胞では、ΔNp63 の発現は低く、PAR1 と thrombin の 発現は⾼い。⼀⽅、低転移株や SQUU-BO 細胞では、ΔNp63 の発現が⾼いも のの、PAR 1 と thrombin の発現は低い。
-
3 2 6
1 1
2-2. ΔNp63ノックダウンがPAR1およびthrombinの発現に与える影響
2-1 の結果より、ΔNp63 の発現は PAR1 および thrombin の発現と逆相関し ていたことから、転写因⼦であるΔNp63 が PAR1 と thrombin の制御している かを検討した。SQUU-A 細胞にΔNp63 siRNA を導⼊し、real-time PCR 法にて PAR1 と thrombin の発現を検索したところ、ΔNp63 のノックダウンにより PAR1 および thrombin の発現が有意に亢進していた(図 5)。
図5. ΔNp63ノックダウンによるPAR1およびthrombinの発現(real-time PCR法)
SQUU-A細胞を使⽤し、ΔNp63をノックダウンするとPAR1、thrombinの発現は有意 に増強している。グラフは独⽴して⾏った3回のデータの平均および標準偏差を⽰す。
なお、統計処理にはMann-Whitney U 検定を⽤いている(*p<0.05)。
siCtrl: scrambled siRNA導⼊SQUU-A細胞。
siΔNp63: ΔNp63 siRNA 導⼊ SQUU-A 細胞。
1 2
4 1
1 2 1 2
3 20.
3
2-3. PAR1 ノックダウンによる EMT 関連遺伝⼦の発現変動
2-2 の結果より、ΔNp63 ノックダウンにより PAR1 の発現が増強することを
⽰したが、PAR1 と EMT との関連性については明らかになっていない。そこで SQUU-B 細胞に PAR1 siRNA を導⼊し、上⽪系マーカー、間葉系マーカーおよ び EMT 関連遺伝⼦の発現変動について real-time PCR 法にて検索した。その結 果、PAR1 のノックダウンにより、上⽪系マーカーである E-cadherin、CK5、
CK14 の発現量は増加したが、間葉系マーカー(vimentin、N-cadherin、
fibronectin)と EMT 関連遺伝⼦(ZEB1、ZEB2、snail、slug、twist)の発現量 は減少した(図 6)。
図6. PAR1ノックダウンによるEMT関連遺伝⼦の発現変動
SQUU-B細胞にPAR1 siRNAを導⼊すると、上⽪系マーカー(E-cadherin、CK5、CK14)
の発現は有意に増強しており、間葉系マーカー(vimentin、N-cadherin、fibronectin)
およびEMT関連遺伝⼦(ZEB1、ZEB2、snail、slug、twist)の発現が有意に減弱してい る。グラフは独⽴して⾏った3回のデータの平均および標準偏差を⽰しており、対照群 の相対的発現量に対する統計学的有意差を⽰す。なお、統計処理にはMann-Whitney U 検定を⽤いている(*p<0.05)。
siCtrl: emptyベクターを導⼊したSQUU-B細胞。
siPAR1: PAR1 siRNAを導⼊したSQUU-B細胞。
ECEC
- . 20 4 8
20 4 8
A8 4 8 581 42 8
08 6 B8
EC
8 8- . 8 8- . 8 8- . 8 8- .
8 8- .
8 8- .
8 8- .
8 8- .
8 8- . 8 8- . 8 8- . 8 8- .
EC EC ECECECECEC EC EC
2-4. PAR1 ノックダウンが遊⾛能および浸潤能に与える影響
2-3 の結果より、PAR1 の発現増強が EMT の誘導に関与していることが⽰唆 されたため、次に PAR1 の発現変動が OSCC 細胞の運動能に与える影響につい て検討した。PAR1 の発現が最も⾼かった SQUU-B 細胞に PAR1 siRNA を導⼊
し、wound healing assay にて解析を⾏った。間隙形成から 24 時間後に間隙を計 測したところ、コントロール群と⽐較して PAR1 をノックダウンした細胞では 遊⾛能が有意に抑制されていた(図 7)。また、MatrigelTM invasion assay にお いても、PAR1 のノックダウンにより SQUU-B 細胞の浸潤能が有意に抑制され た(図 8)。
図 7. PAR1 ノックダウンが OSCC 細胞の遊⾛能に与える影響
(a)顕微鏡像。(b)間隙縮⼩率。PAR1 siRNA 導⼊群の遊⾛能は、対照群と⽐較して 有意に低下している。グラフは、独⽴して⾏った 3 回のデータの平均値及び標準偏差を
⽰す。なお、統計処理には Mann-Whitney U検定を⽤いている(*p<0.05)。scale bars:
50 μm。
図 8. PAR1 ノックダウンが OSCC 細胞の浸潤能に与える影響
(a)浸潤細胞の顕微鏡像(HE 染⾊)。(b)浸潤細胞率。PAR1 siRNA 導⼊群の浸潤 能は、対照群と⽐較して有意に低下している。グラフは、独⽴して⾏った 3 回のデータ の平均値及び標準偏差を⽰す。なお、統計処理には Mann-Whitney U 検定を⽤いてい る(*p<0.05)。scale bars: 100 μm。
42610
考 察
これまでに我々は、OSCCの浸潤先端部でΔNp63の発現が減弱するとEMTが 誘導され、癌細胞の浸潤能が亢進することを報告してきた(40)。またその際に、
浸潤先端部の癌細胞においてthrombin受容体であるPAR1が強く発現しているこ とを⾒出した(41)。これらのことから、PAR1シグナルがEMTの誘導に関与し ていることが⽰唆されたが、EMTにおけるPAR1の役割については不明であった。
そこで本研究では、OSCCにおけるPAR1の発現と機能について研究を⾏った。
まず最初に、OSCC⽣検標本におけるPAR1の発現を免疫組織学的⼿法により 検索した。PAR1は腫瘍細胞と腫瘍間質のそれぞれにおいて発現が認められたた め、その発現様式から3群に分類して臨床病理学的所⾒との関連を検討した。そ の結果、腫瘍細胞および腫瘍間質の両⽅にPAR1の発現を認めた症例では、組織 学的悪性度の⾼い症例が多く、頸部リンパ節転移の発⽣頻度が有意に⾼いこと が明らかとなった。従来より、OSCCの頸部リンパ節転移は分化度や浸潤様式と 強く相関することが知られており、PAR1の発現がこれらの因⼦による影響を強 く受けている可能性がある。そのため、ロジスティック回帰分析による多変量解 析を⾏ったところ、浸潤様式や分化度とは独⽴して、PAR1の発現様式が頸部リ ンパ節転移の唯⼀の予測因⼦となり得ることが明らかとなった。特に注⽬すべ きは、Group Bにおける頸部リンパ節転移の発⽣リスクはGroup Aの1.20倍であ るが、Group Cでは、5.56倍となっていたことである。このことは、腫瘍細胞に おけるPAR1の発現が頸部リンパ節転移に強く影響を及ぼすことを⽰している。
Fanらは、OSCCにおけるPAR1陽性群は、陰性群と⽐較し、頸部リンパ節転移の 発⽣頻度が⾼かったことを報告している(61)。また本研究においても、RT-PCR 法によりOSCC細胞株におけるPAR1の発現を検索したところ、低転移株に⽐べ、
⾼転移株においてPAR1の発現量が多いことを⽰した。これらの知⾒は、腫瘍細 胞におけるPAR1の発現が頸部リンパ節転移に強く影響を及ぼすことを⽀持す る研究結果であると考えられる。
研究1において、我々はOSCCにおけるPAR1の発現を検索したが、興味深いこ とにPAR1陽性症例では全ての症例において間質細胞にPAR1の発現を認めたも のの、腫瘍細胞のみにPAR1の発現を認めた症例は1例もなかった。このことか ら、PAR1を発現している間質細胞が腫瘍細胞のPAR1の発現に関与している可 能性が考えられた。最近の研究によると、腫瘍微⼩環境において間質細胞が成⻑
因⼦や炎症性サイトカインを分泌することで、癌の浸潤・転移に影響を及ぼすこ とが報告されている(62-66)。このような腫瘍微⼩環境における腫瘍細胞と間 質細胞のクロストークにPAR1シグナルが関与していることが徐々に明らかに なってきた。Wangらは、前⽴腺癌において、PAR1の活性化因⼦を腫瘍細胞が 分泌することで、間質細胞内のPAR1シグナルが活性化し、それによって間質か ら分泌されたサイトカインにより癌の増殖能が亢進することを報告している
(67)。本研究においても、浸潤先端部の腫瘍細胞と間質細胞でPAR1の活性化 因⼦であるthrombinが強く発現していたことからも、腫瘍細胞と間質細胞のク ロストークにPAR1シグナルが関与していることが⽰唆された。
PAR1の発現とOSCC患者の予後との関連では、統計学的有意差を認めなかっ たものの腫瘍細胞および間質細胞にPAR1の発現を認めるGroup Cにおいて最も
⽣存率が低かった。これは、最も重要な予後因⼦である頸部リンパ節転移が Group Cにおいて⾼頻度に⾒られたことが⼤きく影響しているものと考えられ る。また、腫瘍細胞におけるPAR1陽性症例は、有意に⽣存率が低かったが、間 質細胞におけるPAR1発現の陽性症例と陰性症例との間に有意差を認めなかっ たことから、腫瘍細胞におけるPAR1の発現がOSCC患者の予後により⼤きな影
響を及ぼすものと考えられた。Fanらは、OSCC患者74例でPAR1の発現を検討 したところ、PAR1陽性群において有意に⽣存率が低下していたことを報告して いる。このことから、PAR1の発現様式は頸部リンパ節転移の予測因⼦としてだ けでなく、OSCC患者の予後を予測するバイオマーカーとしても有⽤である可能 性が⽰唆された。
本研究では、OSCCにおけるPAR1の機能についても検討を⾏った。まず、
OSCC細胞を⽤いてΔNp63をノックダウンしたところ、PAR1およびthrombin の発現が増強した。さらに、PAR1の発現とEMTとの関連を検討するために、
PAR1の発現が最も⾼かったSQUU-B細胞でPAR1のノックダウンを⾏った。そ の結果、SQUU-B細胞が⽰していた間葉系細胞様の形質が失われ、上⽪として の細胞特性が回復し、遊⾛能と浸潤能が著明に抑制されていたことが明らかと なった。Yangらは、ヒト乳癌細胞株であるMCF-7細胞にPAR1を強制発現させる と、上⽪系マーカーの発現量減少、間葉系マーカーの発現量増加、遊⾛能・浸潤 能の亢進が認められ、EMTが誘導されたことを報告している(68)。これらの 知⾒は、PAR1シグナルがEMTの誘導に深く関与している可能性を⽰している。
以上の結果より、OSCC浸潤先端部におけるΔNp63の発現減弱により、腫瘍 細胞におけるPAR1の発現が増強され、腫瘍間質や腫瘍細胞⾃⾝が分泌した thrombinによりPAR1シグナルが活性化し、EMTが誘導されると考えられた。そ の結果、癌細胞の遊⾛・浸潤能が亢進することで、転移がより⾼頻度に⽣じるも のと推察された。
本研究では、PAR1シグナルによりEMTが誘導されることが明らかとなった が、その⼀⽅でPAR1シグナルがEMTを誘導する分⼦メカニズムについては解 明されていない。そのためには、PAR1が活性化された後の細胞内シグナル経路 を解析する必要がある。Zhongらは、⾼転移乳癌細胞でPAR1シグナルが活性化
されると、nuclear factor-kappa B(NF-κB)シグナル伝達系を介してE-cadherin の発現が減弱し、EMTが誘導されることを報告している(69)。また、Cisowski らは、肺癌細胞でのPAR1シグナルの活性化により、extracellular regulated kinase
(ERK)1/2経路が活性化されたことを報告している(70)。このように、PAR1 が活性化された後の細胞内シグナル伝達系については、徐々に明らかになって きているものの、依然として不明な点が多い。今後の研究によりPAR1の細胞内 伝達経路が明らかにされた暁には、癌の浸潤・転移に関わる重要なシグナル経路 を阻害するinhibitorを開発することで、新たな癌治療薬の創成の⼀助となると考 えられる。
謝 辞
稿を終えるにあたり、このような研究の機会を与えて頂きましたとともに終 始御懇篤なる御指導を賜りました九州⼤学⼤学院⻭学研究院⼝腔顎顔⾯病態学 講座顎顔⾯腫瘍制御学分野 中村誠司 教授に深甚なる謝意を表します。
また、直接御指導、御校閲を頂きました九州⼤学⼤学院⻭学研究院⼝腔顎顔⾯病 態学講座顎顔⾯腫瘍制御学分野 川野真太郎 講師に深謝致します。また、常に研 究の協⼒ならびに励ましの⾔葉を頂きました九州⼤学⼤学院⻭学研究院⼝腔顎 顔⾯病態学講座顎顔⾯腫瘍制御学分野の教室員の皆様に深く感謝致します。
参 考 ⽂ 献
1. Carvalho AL, Nishimoto IN, Califano JA, Kowalski LP. Trends in incidence and prognosis for head and neck cancer in the United States: a site-specific analysis of the SEER database. Int J Cancer. 2005; 114: 806-816.
2. Warnakulasuriya S. Global epidemiology of oral and oropharyngeal cancer.
Oral Oncol. 2009; 45: 309-316.
3. Gupta B, Johnson NW, Kumar N. Global epidemiology of head and neck cancers: a continuing challenge. Oncology. 2016; 91(1): 13-23.
4. Cooper JS, Pajak TF, Forastiere AA, Jacobs J, Campbell BH, Saxman SB, et al. Postoperative concurrent radiotherapy and chemotherapy for high-risk squamous-cell carcinoma of the head and neck. N Engl J Med. 2004; 350:
1937-1944.
5. Pignon JP, Bourhis J, Domenge C, Designe L. Chemotherapy added to locoregional treatment for head and neck squamous-cell carcinoma: three meta-analyses of updated individual data. MACH-NC Collaborative Group.
Meta-Analysis of Chemotherapy on Head Neck Cancer. Lancet. 2000; 355:
949-955.
6. Bernier J, Domenge C, Ozsahin M, Matuszewska K, Lefebvre JL, Greiner RH, et al. Postoperative irradiation with or without concomitant
chemotherapy for locally advanced head and neck cancer. N Engl J Med.
2004; 350: 1945-1952.
7. Funk GF, Karnell LH, Robinson RA, Zen WK, Trask DK, and Hoffman HT.
Presentation, treatment and outcome of oral cavity cancer: A national cancer data base report. Head Neck. 2002; 24: 165-180.
8. Sihver W, Pietzsch J, et al. Radiolabeled cetuximab conjugates for EGFR targeted cancer diagnostics and therapy. Pharmaceuticals. 2014; 7: 311-338,.
9. Herbst RS, Langer CJ. Epidermal growth factor receptors as a target for cancer treatment: the emerging role of IMC-C225 in the treatment of lung and head and neck cancers. Semin Oncol. 2002. 29: 27-36.
10. Woodhouse EC, Chuaqui RF, Liotta LA. General mechanisms of metastasis.
Cancer. 1997; 80: 1529-1537.
11. Fidler IJ. The pathogenesis of cancer metastasis: the 'seed and soil' hypothesis revisited. Nat Rev Cancer. 2003; 3: 453-458.
12. Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000; 100: 57-70.
13. Stacker SA, Achen MG, Jussila L, Baldwin ME, Alitalo K.
Lymphangiogenesis and cancer metastasis. Nat Rev Cancer. 2002; 2: 573- 2583.
14. Hanahan D, Folkman J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell. 1996; 86: 353-364.
15. Sahai E. Illuminating the metastatic process. Nat Rev Cancer. 2007; 7: 737- 749.
16. Thiery JP. Epithelial-mesenchymal transitions in tumour progression. Nat Rev Cancer. 2002; 2: 442-454.
17. Yang J, Weinberg RA. Epithelial-mesenchymal transition: at the crossroads of development and tumor metastasis. Dev Cell. 2008; 14: 818-829.
18. Hugo H, Ackland ML, Blick T, Lawrence MG, Clements JA, Williams ED, et al. Epithelial-mesenchymal and mesenchymal-epithelial transitions in
carcinoma progression. J Cellular Physiol. 2007; 213: 374-383.
19. Greenburg G, Hay ED. Epithelia suspended in collagen gels can lose polarity and express characteristics of migrating mesenchymal cells. J Cell Biol. 1982;
95: 333-339.
20. Hay ED. An overview of epithelio-mesenchymal transformation. Acta Anatomica. 1995; 154: 8-20.
21. Kalluri R, Weinberg RA. The basics of epithelial-mesenchymal transition. J Clin Invest. 2009; 119: 1420-1428.
22. Zeisberg M, Neilson EG. Biomarkers for epithelial-mesenchymal transitions.
J Clin Invest. 2009; 119: 1429-1437.
23. Christiansen JJ, Rajasekaran AK. Reassessing epithelial to mesenchymal transition as a prerequisite for carcinoma invasion and metastasis. Cancer Res. 2006; 66: 8319-8326.
24. Peinado H, Olmeda D, Cano A. Snail, Zeb and bHLH factors in tumour progression: an alliance against the epithelial phenotype? Nat Rev Cancer.
2007; 7: 415-428.
25. Wu SY, Yang YP, McClay DR. Twist is an essential regulator of the skeletogenic gene regulatory network in the sea urchin embryo. Dev Biol.
2008; 319: 406-415.
26. Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009; 139: 871-890.
27. Ikawa S, Nakagawara A, Ikawa Y. p53 family genes: structural comparison, expression and mutation. Cell Death Differ. 1999; 6: 1154-1161.
28. Augustin M, Bamberger C, Paul D, Schmale H. Cloning and chromosomal mapping of the human p53-related KET gene to chromosome 3q27 and its murine homolog Ket to mouse chromosome 16. Mammalian Genome : Official Journal of the International Mammalian Genome Society. 1998; 9:
899-902.
29. Trink B, Okami K, Wu L, Sriuranpong V, Jen J, Sidransky D. A new human p53 homologue. Nat Med. 1998; 4: 747-748.
30. Osada M, Ohba M, Kawahara C, Ishioka C, Kanamaru R, Katoh I, et al.
Cloning and functional analysis of human p51, which structurally and functionally resembles p53. Nat Med. 1998; 4: 839-843.
31. Jost CA, Marin MC, Kaelin WG, Jr. p73 is a simian [correction of human]
p53-related protein that can induce apoptosis. Nature. 1997; 389: 191-194.
32. Hagiwara K, McMenamin MG, Miura K, Harris CC. Mutational analysis of the p63/p73L/p51/p40/CUSP/KET gene in human cancer cell lines using intronic primers. Cancer Res. 1999; 59: 4165-4169.
33. Lindsay J, McDade SS, Pickard A, McCloskey KD, McCance DJ. Role of DeltaNp63gamma in epithelial to mesenchymal transition. J Biol Chem.
2011; 286: 3915-3924.
34. Bourdon JC, Fernandes K, Murray-Zmijewski F, Liu G, Diot A, Xirodimas DP, et al. p53 isoforms can regulate p53 transcriptional activity. Genes Dev.
2005; 19: 2122-2137.
35. Ratovitski EA, Patturajan M, Hibi K, Trink B, Yamaguchi K, Sidransky D.
p53 associates with and targets Delta Np63 into a protein degradation pathway. Proc Natl Acad Sci. 2001; 98: 1817-1822.
36. Wu G, Nomoto S, Hoque MO, Dracheva T, Osada M, Lee CC, et al.
DeltaNp63alpha and TAp63alpha regulate transcription of genes with distinct biological functions in cancer and development. Cancer Res. 2003;
63: 2351-2357.
37. Hibi K, Trink B, Patturajan M, Westra WH, Caballero OL, Hill DE, et al.
AIS is an oncogene amplified in squamous cell carcinoma. Proc Natl Acad Sci. 2000; 97: 5462-5467.
38. Crook T, Nicholls JM, Brooks L, O'Nions J, Allday MJ. High level expression of deltaN-p63: a mechanism for the inactivation of p53 in undifferentiated nasopharyngeal carcinoma (NPC)? Oncogene. 2000; 19: 3439-3444.
39. Matsubara R, Kawano S, Kiyosue T, Goto Y, Hirano M, Jinno T, et al.
Increased DeltaNp63 expression is predictive of malignant transformation in
oral epithelial dysplasia and poor prognosis in oral squamous cell carcinoma.
Int J Clin Oncol. 2011; 39: 1391-1399.
40. Goto Y, Kawano S, Matsubara R, Kiyosue T, Hirano M, Jinno T, et al.
Possible involvement of DeltaNp63 downregulation in the invasion and metastasis of oral squamous cell carcinoma via induction of a mesenchymal phenotype. Clin Exp Metastasis. 2014; 31: 293-306.
41. Kaneko N, Kawano S, Yasuda K, Hashiguchi Y, Sakamoto T, Matsubara R, et al. Differential roles of kallikrein-related peptidase 6 in malignant
transformation and ΔNp63β-mediated epithelial-mesenchymal transition of oral squamous cell carcinoma. Oral Oncol. 2017; 75: 148‒157.
42. Vu TK, Hung DT, Wheaton VI, Coughlin SR. Molecular cloning of a functional thrombin receptor reveals a novel proteolytic mechanism of receptor activation. Cell. 1991; 64: 1057-1068.
43. Nystedt S, Emilsson K, Wahlestedt C, Sundelin J. Molecular cloning of a potential proteinase activated receptor. Proc Natl Acad Sci. 1994; 91: 9208- 9212.
44. Kahn ML, Zheng YW, Huang W, Bigornia V, Zeng D, Moff S, et al. A dual thrombin receptor system for platelet activation. Nature. 1998; 394: 690- 694.
45. Coughlin SR. Thrombin signalling and protease-activated receptors. Nature.
2000; 407(6801): 258-264.
46. Ramachandran R, Noorbakhsh F, Defea K, Hollenberg MD.
Targeting proteinase-activated receptors: therapeutic potential and challenges. Nat Rev Drug Discov. 2012; 11(1): 69-86.
47. Ludeman MJ, Kataoka H, Srinivasan Y, Esmon NL, Esmon CT, Coughlin SR. PAR1 cleavage and signaling in response to activated protein
C and thrombin. J Biol Chem. 2005; 280(13): 13122-13128.
48. Pei D. Matrix metalloproteinases target protease-activated receptors on the tumor cell surface. Cancer Cell. 2005; 7(3): 207-208.
49. Toyoda N, Gabazza EC, Inoue H, Araki K, Nakashima S, Oka S, et al.
Expression and cytoprotective effect of protease-activated receptor 1 in gastric epithelial cells. Scand J Gastroenterol. 2003; 38(3): 253-259.
50. Boire A, Covic L, Agarwal A, Jacques S, Sherifi S, Kuliopulos A. PAR1 is a matrix metalloprotease-1 receptor that promotes invasion and tumorigenesis of breast cancer cells. Cell. 2005; 120(3): 303-313.