ク ロ コ ブ タ ケ か ら の 血 管 新 生 阻 害 活 性 物 質 の 探 索
~ 血 管 内 皮 細 胞 増 殖 阻 害 活 性 を 指 標 と し て ~
S e a r c h f o r A n t i a n g i o g e n i c C o n s t i t u e n t s f r o m H y p o x y l o n t r u n c a t u m
~ E v a l u a t i o n o f A n t i p r o l i f e r a t i v e A c t i v i t i e s a g a i n s t Va s c u l a r E n d o t h e l i a l C e l l s ~
平 成 2 3 年 度 入 学
深 井 み ゆ き ( F u k a i , M i y u k i )
指 導 教 員
小 山 清 隆
目 次
P a g e 序 論
第 一 章 キ ノ コ 1
第 二 章 細 胞 増 殖
第 一 節 C D K と サ イ ク リ ン 5
第 二 節 細 胞 周 期 9
第 三 章 血 管 新 生 と が ん 1 5 第 四 章 V E G F と V E G F 受 容 体
第 一 節 V E G F 1 9
第 二 節 V E G F 受 容 体 2 1 第 五 章 分 子 標 的 治 療 薬 2 5
本 論
研 究 目 的 2 9
第 一 章 ク ロ コ ブ タ ケ ( H y p o x y l o n t r u n c a t u m ) 子 実 体 か ら の
H U V E C 増 殖 阻 害 化 合 物 の 探 索 3 1
第 一 節 H U V E C 増 殖 阻 害 3 5 第 二 節 ク ロ コ ブ タ ケ 子 実 体 抽 出 エ キ ス の
H U V E C 増 殖 阻 害 活 性 3 6
第 三 節 ク ロ コ ブ タ ケ 子 実 体 C H C l
3エ キ ス か ら の
H U V E C 増 殖 阻 害 化 合 物 の 探 索 3 7
第 四 節 ク ロ コ ブ タ ケ 子 実 体 M e O H エ キ ス か ら の
構 造 類 縁 体 の 探 索 4 3
第 五 節 H y p o x y l o n o l 類 の 構 造 解 析 4 8
第 二 章 血 管 新 生 阻 害 作 用 の 検 討 9 4
第 一 節 細 胞 選 択 性 9 5
第 二 節 細 胞 毒 性 試 験 9 7
第 三 節 H U V E C 遊 走 阻 害 試 験 9 8 第 四 節 H U V E C 管 腔 形 成 阻 害 試 験 1 0 0 第 三 章 標 的 分 子 の 探 索 1 0 2 第 一 節 K D R リ ン 酸 化 阻 害 試 験 1 0 3 第 二 節 c D N A マ イ ク ロ ア レ イ 解 析 を 用 い た
標 的 分 子 の 探 索 1 0 4
第 三 節 リ ア ル タ イ ム R T- P C R 1 0 9
結 語 111
実 験 の 部 11 7
第 一 章 ク ロ コ ブ タ ケ ( H y p o x y l o n t r u n c a t u m ) 子 実 体 か ら の
H U V E C 増 殖 阻 害 化 合 物 の 探 索
第 一 節 H U V E C 増 殖 阻 害 試 験 1 2 2 第 二 節 ク ロ コ ブ タ ケ 子 実 体 抽 出 エ キ ス の
H U V E C 増 殖 阻 害 活 性 1 2 4
第 三 節 ク ロ コ ブ タ ケ 子 実 体 C H C l
3エ キ ス か ら の
H U V E C 増 殖 阻 害 化 合 物 の 探 索 1 2 5
第 四 節 ク ロ コ ブ タ ケ 子 実 体 M e O H エ キ ス か ら の
構 造 類 縁 体 の 探 索 1 3 2
第 五 節 ス ペ ク ト ル デ ー タ 1 3 9
第 二 章 血 管 新 生 阻 害 作 用 の 検 討
第 一 節 細 胞 選 択 性 1 4 6
第 二 節 細 胞 毒 性 試 験 1 5 0 第 三 節 H U V E C 遊 走 阻 害 試 験 1 5 2 第 四 節 H U V E C 管 腔 形 成 阻 害 試 験 1 5 4 第 三 章 標 的 分 子 の 探 索
第 一 節 K D R リ ン 酸 化 阻 害 試 験 1 5 5 第 二 節 c D N A マ イ ク ロ ア レ イ 解 析 を 用 い た
標 的 分 子 の 探 索
1 5 7 第 三 節 リ ア ル タ イ ム R T- P C R 1 6 0
参 考 文 献 1 6 2
謝 辞 1 6 4
付 表
略 語 一 覧
A L K a n a p l a s t i c l y m p h o m a k i n a s e A M L a c u t e m y e l o g e n o u s l e u k e m i a B A D B c l 2 a n t a g o n i s t o f c e l l d e a t h B c r - A b l b c r - a b l f u s i o n p r o t e i n
B R A F v - r a f m u r i n e s a r c o m a v i r a l o n c o g e n e h o m o l o g B 1 C C R 4 C - C c h e m o k i n e r e c e p t o r t y p e 4
C D c l u s t e r o f d i f f e r e n t i a t i o n C H C l
3c h l o r o f o r m
C M L c h r o n i c m y e l o g e n o u s l e u k e m i a C O S Y c o r r e l a t e d s p e c t r o s c o p y
C O X c y c l o o x y g e n a s e
C T L A - 4 c y t o t o x i c T- l y m p h o c y t e a n t i g e n 4 D M S O d i m e t h y l s u l f o x i d e
D N M T D N A m e t h y l t r a n s f e r a s e
E G F R e p i d e r m a l g r o w t h f a c t o r r e c e p t o r E I e l e c t r o n i o n i z a t i o n
E l k 1 e t s - l i k e g e n e p r o d u c t
E M L 4 e c h i n o d e r m m i c r o t u b u l e - a s s o c i a t e d p r o t e i n - l i k e 4 E R K e x t r a c e l l u l a r s i g n a l - r e g u l a t e d k i n a s e
e x t . e x t r a c t
F B S f e t a l b o v i n e s e r u m f . c . f i n a l c o n c e n t r a t i o n F l k - 1 f e t a l l e v e r k i n a s e 1
F l t - 1 f m s - l i k e t y r o s i n e k i n a s e 1
F l t - 4 f m s - l i k e t y r o s i n e k i n a s e 4
F r. F r a c t i o n
F y n p r o t o - o n c o g e n e t y r o s i n e - p r o t e i n k i n a s e F y n G I S T g a s t r o i n t e s t i n a l s t r o m a l t u m o r
G r b 2 g r o w t h f a c t o r r e c e p t o r - b o u n d p r o t e i n 2 H C l h y d r o g e n c h l o r i d e
H E R 2 h u m a n E G F R - r e l a t e d 2 H D A C h i s t o n e d e a c e t y l a s e
H h h e d g e h o g
H M B C h e t e r o n u c l e a r m u l t i p l e b o n d c o n n e c t i v i t y H M Q C h e t e r o n u c l e a r m u l t i p l e q u a n t u m c o h e r e n c e H R h i g h r e s o l u t i o n
H U A E C h u m a n u m b i l i c a l a r t e r y e n d o t h e l i a l c e l l s H U V E C h u m a n u m b i l i c a l v e i n e n d o t h e l i a l c e l l s i n s o l . i n s o l u t i o n
J A K J a n u s k i n a s e
K D R k i n a s e d o m a i n r e c e p t o r
L c k l y m p h o c y t e c e l l - s p e c i f i c p r o t e i n - t y r o s i n e k i n a s e M A P m i t o g e n - a c t i v a t e d p r o t e i n k i n a s e
M C L m a s t c e l l l e u k e m i a M E K M A P K / E R K k i n a s e
M E M m i n i m u m e s s e n t i a l m e d i u m M e O H m e t h a n o l
m p m e l t i n g p o i n t
m T O R m a m m a l i a n t a r g e t o f r a p a m y c i n
M T T m e t h y l t h i a z o l y l t e t r a z o r i u m N D n o t d e t e r m i n a t i o n
N E T n e u r o e p i t h e l i a l c e l l - t r a n s f o r m i n g g e n e p r o t e i n N H D F n o r m a l h u m a n d e r m a l f i b r o b l a s t
n - h e x a n e n o r m a l - h e x a n e O D o p t i c a l d e n s i t y O D S o c t a d e c y l s i l y l
P B S p h o s p h a t e b u f f e r e d s a l i n e
P D G F R p l a t l e t d e r i v e d g r o w t h f a c t o r r e c e p t o r
P h + A L L P h i l a d e l p h i a - p o s i t i v e s u b t y p e o f a c u t e l y m p h o b l a s t i c l e u k e m i a
R a f r a f g e n e p r o d u c t
R A N K L r e c e p t o r a c t i v a t o r o f n u c l e a r f a c t o r - к B l i g a n d R a s r a s o n c o g e n e p r o d u c t
R B r e t i n o b l a s t o m a - a s s o c i a t e d p r o t e i n r p m r o t a t i o n p e r m i n u t e
R t r e t e n t i o n t i m e
S E G A s u b e p e n d y m a l g i a n t c e l l a s t r o c y t o m a S h b S H 2 d o m a i n - c o n t a i n i n g a d a p t e r p r o t e i n B S H 2 S r c h o m o l o g y 2 d o m a i n
s o l . s o l u t i o n
T S A D T c e l l - s p e c i f i c a d a p t e r p r o t e i n
W S T- 8 2 - ( 2 - m e t h o x y - 4 - n i t r o p h e n y l ) - 3 - ( 4 - n i t r o p h e n y l ) - 5 -
( 2 , 4 - d i s u l f o p h e n y l - 2 H - t e t r a z o l i u m , m o n o s o d i u m s a l t
5 - F U 5 - f l u o r o p y r i m i d i n e - 2 , 4 ( 1 H , 3 H ) - d i o n e ( f l u o r o u r a c i l )
Figure 一 覧
Page
Fig. 1. Classification of Mycota 4
Fig. 2. The Mammalian CDK Family 6
Fig. 3. Mammalian Proteins with A Cyclin-box Domain 8
Fig. 4. Overview of Cell Cycle 9
Fig. 5. G1 Phase of Cell Cycle 11
Fig. 6. G1 and S Phases of Cell Cycle 13
Fig. 7. G2 and M Phases of Cell Cycle 14
Fig. 8. Anatomy of Blood Vessel 17
Fig. 9. Process of Vasculogenesis 17
Fig. 10 . Process of Physiological and Pathological Angiogenesis 18
Fig. 11 . Comparison of Structures of the VEGF Family 20
Fig. 12 . Interactions of VEGF Family with Their Receptors 21
Fig. 13 . VEGF-A/VEGFR-2 Signaling in Vascular Endothelial Cell Proliferation and Migration 23
Fig. 14 . VEGF-A/VEGFR-2 Signaling in Vascular Endothelial Cell Survival 24
Fig. 15 . Isolated Compounds from H . truncatum 33
Fig. 16 . Benzo[ j ]fluoranthene Derivatives from Natural Resource 34
Fig. 17 . Isolation Procedure of H. truncatum CHCl
3ext. Fr. D 40
Fig. 18 . Isolation Procedure of H. truncatum CHCl
3ext. Fr. B 41
Fig. 19 . Isolated Compounds from H. truncatum CHCl
3ext. 42
Fig. 20 . Isolation Procedure of H. truncatum MeOH ext. Fr. D 45
Fig. 21 . Isolation Procedure of H. truncatum MeOH ext. Fr. E 46
Fig. 22 . Isolated Compounds from H. truncatum MeOH ext. 47 Fig. 23 .
1H-
1H COSY and HMBC Correlations of HT-1 in acetone- d
650 Fig. 24 . ORTEP Drawing of HT-1 Obtained by X-ray Analysis 50
Fig. 25 . Structure of Hypoxylonol C ( 23) 51
Fig. 26 .
1H-
1H COSY and HMBC Correlations of HT-2 in acetone- d
654
Fig. 27 . ORTEP Drawing of HT-2 Obtained by X-ray Analysis 54
Fig. 28 . Structure of Hypoxylonol F ( 24) 55
Fig. 29 . ORTEP Drawing of HT-3 Obtained by X-ray Analysis 57
Fig. 30 . Structure of Hypoxylonol B ( 25) 58
Fig. 31 . CD spectra of Hypoxylonol D( 26 ), E ( 27) and F ( 24) 60
Fig. 32 . Structure of Hypoxylonol D( 26 ) 61
Fig. 33 . ORTEP Drawing of HT-5 Obtained by X-ray Analysis 63
Fig. 34 . Structure of Hypoxylonol E ( 27) 64
Fig. 35 . CD spectra of Hypoxylonol A ( 28), B ( 25) and C ( 23) 66
Fig. 36 . Structure of Hypoxylonol A ( 28) 67
Fig. 37 .
1H-
1H COSY, HMBC and NOESY Correlations of HT-7
in acetone- d
669
Fig. 38 . CD spectra of Hypoxylonol E ( 27), F( 24) and G ( 29) 69
Fig. 39 . NOESY Correlations of HT-7 in acetone- d
670
Fig. 40 . Structure of Hypoxylonol G( 29 ) 71
Fig. 41 .
1H-
1H COSY and HMBC Correlations of HT-8 in CDCl
373
Fig . 42. ORTEP Drawing of HT-8 Obtained by X-ray Analysis 73
Fig. 43 . Structure of Truncatone( 7 ) 74
Fig. 44 .
1H-
1H COSY and HMBC Correlations of HT-9 in CD
3OD 76
Fig. 45 .
1H-NMR Spectra of HT-9 and Methylated HT-9 in CD
3OD 77
Fig. 46 . CD spectra of Hypoxylonol B ( 25), C ( 23) and H( 30 ) 78
Fig. 47 . Long-range HMBC and NOESY Correlations of HT-9
in CD
3OD 78
Fig. 48 . ORTEP Drawing of 4- O -Methyl hypoxylonol H
Obtained by X-ray Analysis 79
Fig. 49 . Structure of Hypoxylonol H( 30 ) 81
Fig. 50 .
1H-
1H COSY, HMBC and Long-range HMBC Correlations
of HT-10 CD
3OD 83
Fig. 51 . CD spectra of Hypoxylonol E ( 27), F( 24) and I ( 31) 83
Fig. 52 . ORTEP Drawing of Me-HT-10 Obtained by X-ray Analysis 84
Fig. 53 . Structure of Hypoxylonol I( 31) 86
Fig. 54 .
1H-
1H COSY and HMBC Correlations of HT-11 in CD
3OD 88 Fig. 55 . CD spectra of Hypoxylonol B ( 25), C ( 23) and J ( 32) 88
Fig. 56 . Structure of Hypoxylonol J( 32) 89
Fig. 57 .
1H-
1H COSY and HMBC Correlations of HT-12 in CD
3OD 91 Fig. 58 . CD spectra of Hypoxylonol B ( 25), C ( 23) and K ( 33) 91
Fig. 59 . ORTEP Drawing of HT-12 Obtained by X-ray Analysis 92
Fig. 60 . Structure of Hypoxylonol K( 33 ) 93
Fig. 61 . Cytotoxicity of Hypoxylonol C 97
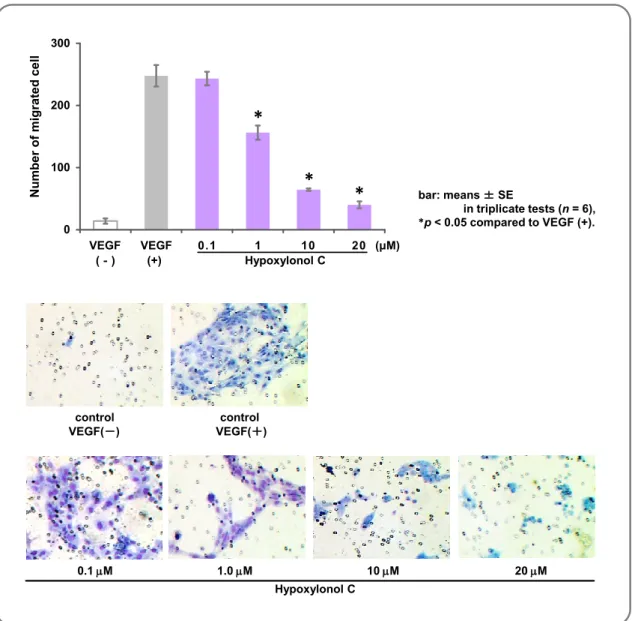
Fig. 62 . Inhibitory Effect of Hypoxylonol C for Migration
of HUVEC 99
Fig. 63 . Inhibitory Effect of Hypoxylonol C for Tubule Formation
of HUVEC 101
Fig. 64 . Pathway of Cell Cycle 106
Fig. 65 . Pathway of Adherens Junction 107
Fig. 66 . Pathway of Focal Adhesion 108
Fig. 67 . Effect of Hypoxylonol C on mRNA Expression
of Cyclins and CDKs 110
Fig. 68 . Effect of Hypoxylonol C on mRNA Expression
of Adhesion Molecules 110
Fig. 69 . HUVEC Growth Inhibition Assay Method 122
Fig. 70 . Isolation Procedure of the Compounds
from H. truncatum CHCl
3ext. Fr. D 126
Fig. 71 . Isolation Procedure of the Compounds
from H. truncatum CHCl
3ext. Fr. B 127
Fig. 72 . Isolation Procedure of the Compounds
from H. truncatum MeOH ext. Fr. D 133
Fig. 73 . Isolation Procedure of the Compounds
from H. truncatum MeOH ext. Fr. E 134
Fig. 74 . HUAEC Growth Inhibition Assay Method 146
Fig. 75 . HeLa Growth Inhibition Assay Meth od 148
Fig. 76 . NHDF Growth Inhibition Assay Method 149
Fig. 77 . Cytotoxicity Assay Method 151
Fig. 78 . HUVEC Migration Assay Method 152
Fig. 79 . HUVEC Tubule Formation Assay Method 154
Fig. 80 . KDR Kinase Inhibitory Assay Method 156
Fig. 81 . Method of RNA Extraction 158
Fig. 82 . Method of Real-time RT-PCR 161
Table 一 覧
Page
Table 1 . Molecular-targeted Drugs 27
Table 2. Effect of the Compounds from H. truncatum CHCl
3ext.
on HUVEC Prolifelation Assay 42
Table 3. Effect of the Compounds from H. truncatum MeOH ext.
on HUVEC Prolifelation Assay 47
Table 4.
1H- and
13C-NMR Spectral Data of Hypoxylonol C ( 23)
in acetone- d
651
Table 5.
1H- and
13C-NMR Spectral Data of Hypoxylonol F ( 24)
in acetone- d
655
Table 6.
1H- and
13C-NMR Spectral Data of Hypoxylonol B ( 25)
in acetone- d
658
Table 7.
1H- and
13C-NMR Spectral Data of Hypoxylonol D ( 26)
in acetone- d
661
Table 8.
1H- and
13C-NMR Spectral Data of Hypoxylonol E ( 27)
in acetone- d
664
Table 9.
1H- and
13C-NMR Spectral Data of Hypoxylonol A ( 28)
in acetone- d
667
Table 10.
1H- and
13C-NMR Spectral Data of Hypoxylonol G ( 29)
in acetone- d
671
Table 11.
1H- and
13C-NMR Spectral Data of Truncatone ( 7)
in CDCl
374
Table 12.
1H- and
13C-NMR Spectral Data of Hypoxylonol H ( 30)
in CD
3OD 80
Table 13.
1H- and
13C-NMR Spectral Data of Hypoxylonol I ( 31)
in CD
3OD 85
Table 14.
1H- and
13C-NMR Spectral Data of Hypoxylonol J ( 32)
in CD
3OD 89
Table 15.
1H- and
13C-NMR Spectral Data of Hypoxylonol K ( 33)
in CD
3OD 93
Table 16. Effect of the Compounds from H. truncatum on HUAEC,
HeLa cells and NHDF Growth inhibitory Assay 96
Table 17.Inhibitory Effect of Hypoxylonol C for Migration of HUVEC 99 Table 18.Inhibitory Effect of Hypoxylonol C for KDR Phosphorylation 103
Table 19. The Pathways including up-regulated genes 105
Table 20. The Pathways including down-regulated genes 105
序 論
第 一 章 キ ノ コ
現 在 , 地 球 上 に は 約 1 5 0 万 種 の 生 物 が 存 在 す る こ と が 確 認 さ れ て お り , 生 物 界 は 細 胞 構 造 や エ ネ ル ギ ー の 摂 取 法 に よ り 生 産 者 , 消 費 者 , 還 元 者 に 大 別 さ れ る . 生 産 者 で あ る 植 物 は , 太 陽 エ ネ ル ギ ー を 光 合 成 に よ り 二 酸 化 炭 素 を 固 定 し て 有 機 物 を 作 り 出 し , 消 費 者 で あ る 動 物 は 植 物 が 生 産 し た 有 機 物 や 他 の 動 物 を 食 す る . さ ら に , 植 物 や 動 物 は , 還 元 者 で あ る 菌 類 に よ っ て 分 解 さ れ て 再 び 無 機 物 へ と 還 元 さ れ , こ れ ら 三 生 物 群 が 共 同 生 活 を 営 む こ と に よ っ て 永 遠 の 生 命 と 生 活 が 保 証 さ れ て い る . 菌 類 の 生 態 系 に お け る 役 割 と し て , 生 物 の 死 骸 を 分 解 す る 分 解 者 と し て の 働 き が 強 調 さ れ る . し か し , 実 際 に 菌 類 の 生 態 を 観 察 す る と , 分 解 者 と し て の 働 き は も ち ろ ん の こ と , 植 物 病 原 菌 に 代 表 さ れ る 寄 生 者 や 線 虫 捕 食 菌 の よ う な 捕 食 者 , あ る い は , 樹 木 や 藻 類 , 昆 虫 類 と 相 利 共 生 す る 共 生 者 と い っ た 多 芸 多 才 な 生 態 を 観 察 で き る . 陸 生 菌 類 の 2 大 系 統 と い え る 担 子 菌 類 と 子 嚢 菌 類 は 様 々 な 陸 上 生 物 の 寄 生 者 ま た は 共 生 者 で あ り , 強 力 な 分 解 者 で も あ る . 担 子 菌 亜 門 は 約 3 万 種 か ら な り , 菌 類 の 3 分 の 1 を 占 め る 大 き な グ ル ー プ で あ る . 担 子 菌 類 は 海 水 や 淡 水 中 か ら も 報 告 さ れ る が , ほ と ん ど が 陸 生 で あ る こ と か ら 陸 上 で 多 様 化 し た 系 統 で あ る と 考 え ら れ て い る . 子 嚢 菌 亜 門
は 5 6 7 4 属 , 約 6 万 4 千 種 か ら な る 菌 類 で 最 も 大 き な グ ル ー プ で , 真
核 生 物 の 中 で も 多 様 性 に 富 む 生 物 群 の 一 つ で あ る . 子 嚢 菌 類 は 陸 上 お
よ び 水 中 の い ず れ に も 広 く 分 布 し て い る . 地 球 上 の 極 限 的 な 場 所 , 例
え ば 南 極 大 陸 の 凍 結 し た 岩 の 内 部 , 深 海 の 木 材 や 堆 積 物 な ど か ら 子 嚢
菌 類 が 分 離 さ れ た 報 告 も あ る .ま た ,子 嚢 菌 の 種 の 約 4 0 % が 藻 類 と の
共 生 体 で あ る 地 衣 を 形 成 し ,地 球 の 陸 上 表 面 の 約 8 % を 被 覆 し て い る .
「 キ ノ コ 」 と 呼 ば れ る 大 形 の 生 殖 器 , つ ま り は 子 実 体 を 形 成 す る 菌 は 一 部 で あ り , そ の 大 部 分 が 担 子 菌 亜 門 に , そ れ 以 外 は 子 嚢 菌 亜 門 に 属
す る ( F i g . 1 ). こ の 俗 に キ ノ コ と 呼 ば れ る 菌 類 は シ ダ 植 物 や 種 子 植
物 と 時 代 を 同 じ く し て 地 球 上 に 出 現 し , 長 い 進 化 の 歴 史 を 背 景 に , 生 活 体 は 単 純 で あ る が 極 め て 複 雑 多 彩 な 子 実 体 を も つ よ う に な っ た .
1 , 2)昔 か ら 一 部 の キ ノ コ 類 は , 食 用 だ け で な く 薬 用 と し て も 利 用 さ れ て き た . 中 国 最 古 の 薬 物 書 『 神 農 本 草 経 』 の 上 品 に は 「 赤 芝 」,「 黒 芝 」,
「 青 芝 」, 「 白 芝 」, 「 黄 芝 」 ,「 紫 芝 」 の 六 芝 お よ び 「 茯 苓 」 が ,中 品 に は「 猪 苓 」 お よ び 「 桑 耳 」 が , 下 品 に は 「 雷 丸 」 が 収 載 さ れ て い る . 六 芝 の う ち 主 な も の は 紫 芝 と 赤 芝 で , こ れ ら は 霊 芝 と 称 す る マ ン ネ ン タ ケ G a n o d e r m a l u c i d u m ( L e y s s . e x . F r. ) K a r s t ま た は そ の 近 縁 種 の 子 実 体 で あ る . 茯 苓 と 猪 苓 は 「 桂 枝 茯 苓 丸 」, 「 猪 苓 湯 」 , 「 五 苓 散 」 な ど の 漢 方 処 方 に 使 用 さ れ る . 桑 耳 は 『 薬 性 論 』 に お い て は 桑 檽 ( そ う じ ) , 桑 黄 ( そ う お う ) と も 呼 ば れ , そ れ ぞ れ キ ク ラ ゲ の 類 , サ ル ノ コ シ カ ケ の 類 を さ す . 梁 の 『 本 草 経 集 注 』 に は 「 鬼 蓋 ( き が い )」 と 「 馬 勃 ( ば ぼ つ )」が あ げ ら れ て い る .鬼 蓋 は ヒ ト ヨ タ ケ 科 の ヒ ト ヨ タ ケ C o p r i n u s a t r a m e n t a r i u s ( B u l l . ) F r. の 子 実 体 で 小 児 の 諸 癇 な ど に 用 い ら れ る . 馬 勃 は ホ コ リ タ ケ 科 の オ ニ フ ス ベ L a s i o s p h a e r a f e n z l i i R e i c h . な ど の 子 実 体 で , 止 血 , 抗 炎 症 薬 と し て 用 い ら れ る . 唐 代 の 『 新 修 本 草 』 に は「 槐 耳 」が 収 載 さ れ て い る が ,こ れ は エ ン ジ ュ( 中 国 名:槐 ) S o p h o r a j a p o n i c a L . に つ く キ ク ラ ゲ A u r i c u l a r i a a u r i c u l a ( L . e x H o o k . )
U n d e r w . で , 止 血 薬 と し て 用 い ら れ , ま た 食 用 に も し て い る .『 酉 陽
雑 爼 ( ゆ う よ う ざ っ そ )』 に は 「 猢 猻 眼 ( こ そ ん が ん )」 の 名 が み ら れ
る が , こ れ は 桑 黄 と 同 類 と さ れ て い る . 日 本 で も 「 梅 寄 生 」 と 称 す る
生 薬 に 抗 が ん 作 用 が あ る と し て 民 間 で 用 い ら れ て い る が , こ れ も 同 様
の も の で あ る . 「 雷 丸 」は サ ル ノ コ シ カ ケ 科 の ラ イ ガ ン キ ン P o l y p o r u s m y l i t t a e C o o k . e t M a s s . の 菌 核 で , 条 虫 駆 除 薬 と す る .『 本 草 拾 遺 』 に は ,「 土 菌 」 と 「 鬼 筆 」 が 収 載 さ れ て い る . 土 菌 の う ち の 1 種 「 仙 人 帽 」 は , ス ッ ポ ン タ ケ 科 の キ ヌ ガ サ タ ケ D i c t y o p h o r a i n d u s i a t a
F i s c h . で ,血 病 に 主 効 が あ る と さ れ て い る .鬼 筆 は ス ッ ポ ン タ ケ 科 の
キ ツ ネ ノ タ イ マ ツ P h a l l u s r u b i c u n d u s ( B o s c . )F r. の 子 実 体 な ど で , で き も の に 外 用 す る . 宋 代 の 『 図 経 本 草 』 の 「 杉 菌 」 は 心 痛 を 治 す キ ノ コ で , お そ ら く モ エ ギ タ ケ 科 の ス ギ タ ケ P h o l i o t a s q u a r r o s a ( F r. )
Q u é l . の 子 実 体 で あ る と い う 説 も あ る が 確 か で は な い . 『 証 類 本 草 』に
は 「 蟬 花 ( 現 在 で は 金 蟬 花 )」 と 称 す る 変 わ っ た 真 菌 類 が 登 場 す る . こ れ は セ ミ 科 の 幼 虫 に 寄 生 し た バ ッ カ ク キ ン 科 の セ ミ タ ケ C o r d y c e p s s o b o l i f e r a ( H i l l . ) B e r k . e t B r o o m e の 菌 核 ( 寄 生 虫 体 ) お よ び 子 座 で , 小 児 の 驚 癇 , 夜 啼 き な ど に 用 い て い る . 元 代 の 『 日 用 本 草 』 に は
「 香 蕈 」 や 「 天 花 蕈 」 な ど が 収 載 さ れ て い る . 香 蕈 は キ シ メ ジ 科 の シ イ タ ケ L e n t i n u s e d o d e s ( B e r k . ) S i n g . で , 食 用 キ ノ コ と し て 最 も よ く 使 わ れ る も の で あ る が , 薬 と し て は 胃 腸 障 害 , 貧 血 症 な ど に 用 い ら れ て い る .天 花 蕈 は キ シ メ ジ 科 の ヒ ラ タ ケ P l e u r o t u s o s t r e a t u s ( J a c q .
e x F r. ) Q u é l . で , こ れ も 食 用 キ ノ コ で あ る . 明 代 の 『 本 草 綱 目 』 に
は 食 用 キ ノ コ と し て 名 高 い キ シ メ ジ 科 の マ ツ タ ケ Tr i c h o l o m a m a t s u t a k e ( S . I t o e t I m a i ) S i n g . も ,「 松 蕈 」 の 名 で 収 載 さ れ ,「 小 便 が 濁 っ て 禁 ぜ ざ る を 治 す 」 と そ の 効 を 誌 し て い る . 清 代 の 『 本 草 從 新 』 に は ,「 冬 虫 夏 草 」 が 収 載 さ れ て い る .
3 , 4)近 年 で は キ ノ コ 類 が 制 が ん 剤 と し て 注 目 さ れ て い る . 現 在 , 抗 悪 性
腫 瘍 剤 と し て ク レ ス チ ン
®, レ ン チ ナ ン
®, シ ゾ フ ィ ラ ン ( ソ ニ フ ィ ラ
ン
®) の 3 剤 が 臨 床 に お い て 使 用 さ れ て い る . こ れ ら は そ れ ぞ れ カ ワ
ラ タ ケ C o r i o l u s v e r s i c o l o r ( L , : F r. ) Q u é l . の 培 養 菌 糸 体 , シ イ タ ケ L e n t i n u l a e d o d e s ( B e r k . ) S i n g . , ス エ ヒ ロ タ ケ S c h i z o p h y l l u m c o m m u n e F r. : F r. か ら 得 ら れ る 多 糖 ま た は タ ン パ ク 質 ‐ 多 糖 複 合 体 と の 混 合 物 で あ り , い ず れ も 他 の 化 学 療 法 や 放 射 線 療 法 と 組 み 合 わ せ て 使 わ れ て い る .
キ ノ コ が 産 生 す る 二 次 代 謝 産 物 に は 他 の 菌 類 に は み ら れ な い 特 徴 的 な 構 造 を も つ も の が あ る . 担 子 菌 は 高 等 植 物 に 似 て シ キ ミ 酸 経 路 が 発 達 し て お り , し ば し ば テ ル フ ェ ニ ル キ ノ ン が 得 ら れ る . そ の 他 , メ バ ロ ン 酸 経 路 由 来 の テ ル ペ ン 類 も 多 く , 中 で も 酢 酸 - マ ロ ン 酸 経 路 の ポ リ ア セ チ レ ン は 担 子 菌 に 特 有 で あ る . ア ミ ノ 酸 由 来 の 呈 味 性 の 強 い 殺 蝿 物 質 イ ボ テ ン 酸 , 幻 覚 性 の あ る イ ン ド ー ル 化 合 物 , 致 死 性 毒 の ヘ テ ロ 環 状 ペ プ チ ド な ど も キ ノ コ 由 来 の 独 特 な 二 次 代 謝 産 物 で あ る .
4)し か し , キ ノ コ の 二 次 代 謝 産 物 や , 菌 類 の 分 類 に つ い て , 系 統 だ っ た 研 究 は ま だ ま だ 充 分 で あ る と は 言 い 難 い . 自 然 環 境 の 保 護 が 叫 ば れ る 昨 今 に お い て , 自 然 界 の 秩 序 を 正 し く 理 解 す る た め に 菌 類 研 究 の 展 開 は 必 要 不 可 欠 で あ る .
F i g . 1 . C l a s s i f i c a t i o n o f M y c o t a
1)第 二 章 細 胞 増 殖
細 胞 の 成 長 と は , 細 胞 の 質 量 の 大 部 分 を 占 め る タ ン パ ク 質 や 膜 , 細 胞 小 器 官 や そ の 他 の 構 成 要 素 を 合 成 す る こ と に よ り 細 胞 の 大 き さ を 増 加 さ せ る 過 程 で あ る . 細 胞 は あ る 程 度 の 大 き さ に な る と , 成 長 を 止 め る か 分 裂 を す る . 分 裂 か ら 次 の 分 裂 ま で を 細 胞 周 期 ( c e l l c y c l e ) と 呼 び , 細 胞 分 裂 は , 核 が 分 裂 す る 有 糸 分 裂 ( m i t o s i s ) と 細 胞 質 が 二 つ に 分 か れ る 細 胞 質 分 裂 ( c y t o k i n e s i s ) に 分 け ら れ る . 多 細 胞 生 物 に お い て は , 細 胞 の 成 長 と 分 裂 が ど の よ う に し て 連 携 し て い る の か , い ま だ 未 知 な 部 分 が 多 い . 細 胞 , 組 織 そ し て 生 体 の 大 き さ を 制 御 す る 分 子 機 構 は , 生 物 学 の 中 で も 興 味 深 く 神 秘 的 な 課 題 の 一 つ で あ る .
第 一 節 C D K と サ イ ク リ ン
細 胞 周 期 の 制 御 機 構 を 構 成 す る 中 心 的 な 因 子 は , サ イ ク リ ン 依 存 性 キ ナ ー ゼ ( c y c l i n - d e p e n d e n t k i n a s e ; C D K ) で あ る . 細 胞 周 期 の 進 行 に 従 い , こ の キ ナ ー ゼ 活 性 は 急 激 に 変 化 し , そ れ に 従 っ て 細 胞 周 期 の 進 行 を 制 御 す る タ ン パ ク 質 の リ ン 酸 化 状 態 と そ れ に よ る 活 性 化 状 態 に 変 化 が も た ら さ れ る . C D K タ ン パ ク 質 の 細 胞 内 濃 度 は 細 胞 周 期 を 通 じ て 一 定 で あ る の で , C D K の 活 性 の 変 動 は ,原 則 的 に サ イ ク リ ン( c y c l i n ) と し て 知 ら れ る 制 御 サ ブ ユ ニ ッ ト の 量 の 変 動 に 依 存 し て い る .
C D K ( c y c l i n - d e p e n d e n t k i n a s e )
C D K は セ リ ン / ス レ オ ニ ン キ ナ ー ゼ の 一 フ ァ ミ リ ー で ,定 義 上 す べ
て の C D K は そ の 酵 素 活 性 に 調 節 サ ブ ユ ニ ッ ト で あ る サ イ ク リ ン の 結
合 を 必 要 と す る .C D K の 機 能 は ,細 胞 周 期 の 各 過 程 に お い て 細 胞 内 の 数 多 く の タ ン パ ク 質 の 特 異 的 な ア ミ ノ 酸 配 列 上 の セ リ ン も し く は ス レ オ ニ ン 残 基 を リ ン 酸 化 す る こ と で あ る .
5)1 9 8 7 年 に N u r s e ら に よ り ヒ ト の C D K 1 が , 1 9 9 1 年 に Ts a i ・H a r l o w ・ M e y e r s o n ,あ る い は E l l e d g e ら に よ り C D K 2 が そ れ ぞ れ 同 定 さ れ て か ら , こ れ ま で に C D K 11 ま で が 同 定 さ れ て い る ( F i g . 2 ). こ の う ち の
C D K 1 , 2 , 4 お よ び 6 の わ ず か 4 種 が 細 胞 周 期 の 制 御 に 直 接 関 わ っ て
い る . 細 胞 周 期 の 各 過 程 は , C D K 1 と 2 と い う 2 つ の C D K に よ っ て 制 御 さ れ て お り , 主 に M 期 と S 期 に そ れ ぞ れ 作 用 す る . さ ら に , 細 胞 外 因 子 に 応 答 し て 細 胞 周 期 へ の 進 行 を 調 節 す る 2 つ の 重 要 な C D K 4 と 6 が 存 在 す る .そ の 他 に P C TA I R E や P F TA I R E ,C C R K , C H E D , C R K 7 と い っ た 高 度 に 関 連 す る 分 子 も C D K フ ァ ミ リ ー の 一 員 と 称 さ れ る が , そ れ ぞ れ の 活 性 化 調 節 サ ブ ユ ニ ッ ト は い ま だ 同 定 さ れ て い な い .
6 , 7)CDK1 297 aa
CDK2 298 aa
CDK3 305 aa
CDK5 292 aa
PCTAIRE3 502 aa PCTAIRE1 496 aa PCTAIRE2 523 aa PFTAIRE1 451 aa PFTAIRE2 435 aa
CDK4 303 aa
CDK6 326 aa
CDK10 360 aa
PITSLRE 795 aa
CCRK 346 aa
CDK9 372 aa
CHED 1512 aa
CRK7 1490 aa
CDK7 346 aa
CDK8 464 aa
CDK11 502 aa
Ser/Thr protein kinase Nuclear localization Coiled-coil domains
F i g . 2 . T h e M a m m a l i a n C D K F a m i l y
7)サ イ ク リ ン
サ イ ク リ ン は C D K フ ァ ミ リ ー に 属 す る 分 子 と 結 合 し 活 性 化 さ せ る と い う 特 徴 を 示 す . ほ と ん ど の サ イ ク リ ン は 細 胞 周 期 を 通 じ て そ の 細 胞 内 濃 度 を 劇 的 に 変 化 さ せ , そ れ に よ っ て C D K 活 性 が 周 期 的 に 変 化 し , 細 胞 周 期 制 御 の 中 心 的 役 割 を 担 っ て い る . サ イ ク リ ン の 細 胞 内 濃 度 は 主 に サ イ ク リ ン 遺 伝 子 発 現 と サ イ ク リ ン タ ン パ ク 質 の 分 解 に よ っ て 調 節 さ れ て い る .
現 在 ま で に , 「 c y c l i n - b o x 」 と 呼 ば れ る 1 5 0 の ア ミ ノ 酸 残 基 か ら な る 保 存 さ れ た ス ト レ ッ チ を も つ 2 9 の サ イ ク リ ン 関 連 分 子 が 同 定 さ れ て い る ( F i g . 3 ). こ の ド メ イ ン は 5 つ の へ リ ッ ク ス 領 域 か ら な り , こ れ に よ っ て C D K な ど の パ ー ト ナ ー タ ン パ ク 質 と 結 合 す る .
細 胞 周 期 に 関 わ る サ イ ク リ ン は , 主 に 細 胞 周 期 に お け る 発 現 の 時 期 と そ の 機 能 に よ っ て G 1 期 サ イ ク リ ン ( サ イ ク リ ン D ), G 1 / S 期 サ イ ク リ ン ( サ イ ク リ ン E ), S 期 サ イ ク リ ン ( サ イ ク リ ン A ) お よ び M 期 サ イ ク リ ン ( サ イ ク リ ン B ) の 4 つ の ク ラ ス に 分 類 さ れ る .
サ イ ク リ ン A に は , 別 々 の 遺 伝 子 に コ ー ド さ れ た サ イ ク リ ン A 1 と
サ イ ク リ ン A 2 が 存 在 す る .サ イ ク リ ン A 1 は 生 殖 細 胞 お よ び 初 期 胚 の
細 胞 で あ り , サ イ ク リ ン A 2 は 発 生 初 期 お よ び 成 体 で 発 現 す る が , 細
胞 周 期 に お け る 機 能 に 関 し て 2 つ の サ ブ タ イ プ 間 で ほ と ん ど 差 は な い .
サ イ ク リ ン B に は サ イ ク リ ン B 1 と サ イ ク リ ン B 2 , サ イ ク リ ン B 3 が
存 在 す る も の の , サ イ ク リ ン B 3 は 分 裂 に 関 し て 明 確 な 機 能 を も っ て
い な い . 細 胞 周 期 に お い て , サ イ ク リ ン B 1 の 方 が よ り 重 要 だ と 考 え
ら れ , よ く 研 究 さ れ て い る .
5)Cyclin F 786 aa Cyclin O 316 aa Cyclin A1 465 aa Cyclin A2 432 aa Cyclin B3 1395 aa Cyclin B1 433 aa Cyclin B2 398 aa Cyclin D3 292 aa Cyclin D1 295 aa Cyclin D2 289 aa Cyclin E1 410 aa Cyclin E2 404 aa Cyclin J 372 aa Cyclin C 302 aa Cyclin H 323 aa Cyclin K 355 aa Cyclin T1 726 aa Cyclin T2 730 aa Cyclin L1 526 aa Cyclin L2 424 aa Cyclin M3 707 aa Cyclin M1 586 aa Cyclin M2 854 aa Cyclin M4 727 aa Cables 1 633 aa Cables 2 478 aa Cyclin I 377 aa Cyclin G1 295 aa Cyclin G2 344 aa
N-terminal cyclin box C-terminal cyclin box F-box Nuclear localization signal Proline-rich
F i g . 3 . M a m m a l i a n P r o t e i n s w i t h A C y c l i n - b o x D o m a i n
7)第 二 節 細 胞 周 期
「 細 胞 ( c e l l u l a )」 と い う 名 称 が 1 7 世 紀 半 ば に 付 与 さ れ て 以 来 , 約 2 0 0 年 を 経 て 「 細 胞 は 細 胞 に 由 来 す る ( O m n i s c e l l u l a e c e l l u l a )」 と い う 概 念 が 提 唱 さ れ た . さ ら に 約 1 0 0 年 を 要 し て は じ め て , こ の 「 細 胞 に 由 来 す る 」過 程 が G 1 期( G a p 1 p h a s e )- S 期( S y n t h e s i s p h a s e )
- G 2 期 ( G a p 2 p h a s e ) - M 期 ( M i t o t i c p h a s e ) と い う 4 つ の 位 相 と し て 定 義 さ れ た . し か し 細 胞 周 期 と い う 用 語 が い つ か ら 使 用 さ れ る よ う に な っ た の か は 定 か で は な い .
細 胞 周 期 の う ち , 有 糸 分 裂 の 時 期 を M 期 と い い , M 期 と M 期 の 間 の 長 い 期 間 を 間 期 ( i n t e r p h a s e ) と い う . 間 期 は さ ら に G 1 , S , G 2 期 に 分 け ら れ る . S 期 は 核 の D N A の 複 製 が お こ る 時 期 で あ り , M 期 か ら S 期 の 始 ま り ま で の 期 間 が G 1 期 , つ ま り 最 初 の ギ ャ ッ プ 期 で あ る . S 期 を 完 了 す る と , 細 胞 は 二 番 目 の ギ ャ ッ プ 期 で あ る G 2 期 に 入 る . こ の 時 期 に 細 胞 分 裂 の 最 終 段 階 に 到 達 し , タ ン パ ク 質 の 合 成 が 増 加 す る ( F i g . 4 ).
6)cyclin E
CDK2 cyclin A CDK2
cyclin A CDK1
cyclin B CDK1
S
G
2G
1G
0M
cyclin D CDK4/6
F i g . 4 . O v e r v i e w o f C e l l C y c l e
多 細 胞 生 物 で は 新 た な 細 胞 は 必 要 と さ れ る と き に の み 作 り 出 さ れ る よ う , 組 織 特 異 的 な 遺 伝 子 の プ ロ グ ラ ミ ン グ と , 他 の 細 胞 が 分 泌 す る 分 裂 促 進 因 子 ( m i t o g e n ; マ イ ト ジ ェ ン ) と 呼 ば れ る 細 胞 外 タ ン パ ク 質 か ら 受 け 取 る シ グ ナ ル に よ っ て , そ の 増 殖 が 制 御 さ れ て い る . 培 養 細 胞 か ら 分 裂 促 進 因 子 を 除 去 し た と き な ど に は , 細 胞 は 細 胞 周 期 か ら 完 全 に 脱 出 し て G 0 期 と 呼 ば れ る 特 別 な 休 止 状 態 に 入 る .
細 胞 外 か ら の シ グ ナ ル で あ る 分 裂 促 進 因 子 に よ っ て , 細 胞 周 期 の 初 期 の 現 象 を 引 き 起 こ す サ イ ク リ ン D - C D K 4 / 6 の 活 性 化 が 起 こ る .分 裂 促 進 因 子 は , R a s - M A P キ ナ ー ゼ 経 路 の 活 性 化 を 介 し て サ イ ク リ ン D 遺 伝 子 の 発 現 を 誘 導 す る . 休 止 期 の 細 胞 に わ ず か に 残 る サ イ ク リ ン
D - C D K 4 / 6 は , グ リ コ ー ゲ ン シ ン タ ー ゼ キ ナ ー ゼ 3 ( g l y c o g e n
s y n t h a s e k i n a s e 3;G S K 3 )に よ っ て 阻 害 的 リ ン 酸 化 を 受 け て い る が , 分 裂 促 進 因 子 の 刺 激 は サ イ ク リ ン D の 脱 リ ン 酸 化 を 引 き 起 こ す .さ ら に ,活 性 を も つ サ イ ク リ ン D - C D K 4 / 6 複 合 体 の 形 成 に は 補 助 因 子 が 必 要 で あ り , p 2 7 や p 2 1 な ど の C i p / K i p フ ァ ミ リ ー タ ン パ ク 質 が 両 サ ブ ユ ニ ッ ト と 相 互 作 用 す る こ と で こ れ ら の 結 合 が 強 く な る . 安 定 化 し た サ イ ク リ ン D - C D K 4 / 6 複 合 体 は 標 的 で あ る R B タ ン パ ク 質 が 存 在 す る 核 に 蓄 積 す る .
R B タ ン パ ク 質 に は p R B ,p 1 0 7 ,p 1 3 0 の 三 つ の 関 連 タ ン パ ク 質 が 存
在 し ,そ れ ぞ れ が E 2 F と 呼 ば れ る 特 定 の タ ン パ ク 質 複 合 体 フ ァ ミ リ ー
と 結 合 す る . E 2 F 複 合 体 は , E 2 F フ ァ ミ リ ー タ ン パ ク 質 の サ ブ ユ ニ ッ
ト と , D P フ ァ ミ リ ー タ ン パ ク 質 の サ ブ ユ ニ ッ ト 一 つ ず つ か ら 成 る ヘ
テ ロ 二 量 体 で あ り , G 1 / S 期 の 標 的 遺 伝 子 の プ ロ モ ー タ ー に 存 在 す る
特 定 の D N A 配 列 に 結 合 す る . E 2 F タ ン パ ク 質 は , そ の 機 能 に よ っ て
活 性 化 E 2 F と 抑 制 性 E 2 F の 2 種 類 に 分 け ら れ る .休 止 期 の 細 胞 で は ,
活 性 化 E 2 F は リ ン 酸 化 さ れ た R B タ ン パ ク 質 , p R B に よ っ て 抑 制 さ れ て お り , 一 方 , G 1 / S 期 遺 伝 子 の プ ロ モ ー タ ー は 抑 制 性 E 2 F と p 1 0 7 も し く は p 1 3 0 と の 複 合 体 に よ っ て 抑 制 さ れ て い る .R B フ ァ ミ リ ー タ ン パ ク 質 は ,活 性 化 し た サ イ ク リ ン D - C D K 4 / 6 複 合 体 依 存 的 に リ ン 酸 化 さ れ て E 2 F か ら 解 離 す る . D N A 上 の 抑 制 性 E 2 F は , p R B と 結 合 し て い な い 活 性 化 E 2 F に 置 き 換 わ り , G 1 / S 期 遺 伝 子 の 発 現 が 活 性 化 さ れ , こ れ に よ っ て 細 胞 周 期 の ス タ ー ト が 切 ら れ る ( F i g . 5 ).
5)S
G 2 G 1
G 0 M
DP E2F
p107
cyclin D CDK4/6
発現 増殖刺激 DP
E2F
cyclin D CDK4/6
P
脱リン酸化 cyclin D
CDK4/6 p27
DP E2F
pRB
pRB
P P
DP E2F
P P
p107
E2F DP
抑制性
活性化
p27
F i g . 5 . G 1 P h a s e o f C e l l C y c l e
G 1 / S 期 遺 伝 子 産 物 の 中 で 決 定 的 に 重 要 な の は ,G 1 / S 期 サ イ ク リ ン の サ イ ク リ ン E と S 期 サ イ ク リ ン の サ イ ク リ ン A の 二 つ で あ る .サ イ ク リ ン E は C D K 2 と 複 合 体 を 形 成 す る が , 細 胞 周 期 に 入 る 際 の
C D K 2 の 活 性 は p 2 7 に よ っ て 部 分 的 に 抑 制 さ れ て い る . サ イ ク リ ン D
- C D K 4 / 6 複 合 体 の 量 が 増 加 す る こ と で p 2 7 の 流 通 量 が 減 少 し , さ
ら に p 2 7 が タ ン パ ク 質 分 解 を 受 け 不 活 性 化 さ れ る こ と で サ イ ク リ ン E
- C D K 2 の 活 性 化 が 促 進 さ れ る .G 1 期 後 期 に な る と ,ホ ス フ ァ タ ー ゼ
C d c 2 5 A に よ る C D K 2 の 阻 害 的 リ ン 酸 化 の 除 去 が 起 こ り , サ イ ク リ ン
E - C D K 2 複 合 体 の 活 性 は さ ら に 上 昇 す る . サ イ ク リ ン D - C D K 4 / 6
は 部 分 的 な R B タ ン パ ク 質 の リ ン 酸 化 と E 2 F の 活 性 化 し か 行 わ な い .
E 2 F の 完 全 な 活 性 化 は , サ イ ク リ ン E - C D K 2 複 合 体 の 活 性 が G 1 期
後 期 に 上 昇 し , R B タ ン パ ク 質 が 完 全 に リ ン 酸 化 さ れ た と き に 起 こ る
( 正 の フ ィ ー ド バ ッ ク ). そ し て S 期 初 期 に な る と サ イ ク リ ン E -
C D K 2 が サ イ ク リ ン E の T h r 3 8 0 を リ ン 酸 化 す る な ど し て , ユ ビ キ チ
ン 化 を 経 て サ イ ク リ ン E が 分 解 さ れ る .
サ イ ク リ ン A は C D K 2 と 複 合 体 を 形 成 す る が ,サ イ ク リ ン A - C D K 2 の 活 性 化 は D N A の 複 製 を 開 始 す る た め に 必 要 で あ る . サ イ ク リ ン E
- C D K 2 と 同 様 に , サ イ ク リ ン A - C D K 2 複 合 体 は p 2 7 に よ っ て 部 分
的 に 抑 制 さ れ る の で , 完 全 な 活 性 化 に は p 2 7 の 除 去 が 必 要 で あ る . ま
た , サ イ ク リ ン A - C D K 2 の 活 性 化 は C d c 2 5 A に よ る 脱 リ ン 酸 化 に も
依 存 し て い る . サ イ ク リ ン A - C D K 2 は 活 性 化 さ れ る と , 複 製 開 始 点
の p r e - r e p l i c a t i o n c o m p l e x を リ ン 酸 化 し , D N A 合 成 が 開 始 さ れ る( F i g .
6 ).
5)S
G 2 G 1
G 0 M
DP E2F
p107 E2F DP
DP E2F
pRB
pRB
P P
E2F DP
P P
p107
DP E2F
抑制性
活性化 cyclin E
CDK2 p27
P p27
cyclin E CDK2 Cdc25A
CDK2 cyclin E
CDK2 P 自己
リン酸化 cyclin ACDK2 p27
P
p27 Cdc25A
cyclin A CDK2
Cdc6 Cdt1 Dbf4Cdc7
ORC
ORC
ORC DNApol
DNA合成 Cdc6
+P
F i g . 6 . G 1 a n d S P h a s e s o f C e l l C y c l e
サ イ ク リ ン A は ま た , サ イ ク リ ン B と と も に M 期 開 始 に お け る 主 要 か つ 必 須 な 制 御 因 子 で も あ る . サ イ ク リ ン A は M 期 前 期 に は そ の パ ー ト ナ ー を C D K 1 に 変 え , 活 性 化 し た 状 態 で 核 内 に 存 在 す る . 染 色 体 の 凝 縮 な ど と い っ た M 期 最 初 の 核 内 で の 事 象 は ,お そ ら く サ イ ク リ
ン A - C D K 1 複 合 体 に よ っ て 開 始 さ れ る .
サ イ ク リ ン B は G 2 期 に 発 現 量 が 上 昇 し , 細 胞 質 に 存 在 す る . サ イ ク リ ン B - C D K 1 複 合 体 の 活 性 化 は M 期 初 期 の 初 め に 細 胞 質 で 始 ま り , と り わ け 中 心 体 で 顕 著 で あ る . M 期 初 期 に 蓄 積 し た 不 活 性 化 状 態 の サ イ ク リ ン B - C D K 1 は , C d c 2 5 フ ァ ミ リ ー ホ ス フ ァ タ ー ゼ に よ っ て
C D K 1 の 阻 害 的 リ ン 酸 化 が 除 か れ 活 性 化 さ れ る .サ イ ク リ ン A - C D K 1
は C d c 2 5 な ど を リ ン 酸 化 し , サ イ ク リ ン B - C D K 1 を 活 性 化 す る フ ィ
ー ド バ ッ ク を 手 助 け す る . M 期 前 期 の 後 半 に は , ほ と ん ど の サ イ ク リ
ン B - C D K 1 は 突 然 核 内 に 移 行 し , 核 膜 崩 壊 を 誘 導 す る . そ の 後 間 も な く 核 膜 は 消 失 し , サ イ ク リ ン B - C D K 1 は 細 胞 全 体 に 分 布 さ れ る . 一 部 は 細 胞 質 に 残 り ,中 心 体 の 分 離 や ゴ ル ジ 体 の 再 構 築 な ど M 期 に 起 こ る 他 の 事 象 を 推 し 進 め る . さ ら に , 染 色 体 凝 縮 の 完 了 も 誘 導 す る
( F i g . 7 ).
5)S
G 2 G 1
G 0 M
cyclin A CDK1
cyclin B
cyclin B CDK1
P P P
cyclin B CDK1
P Cdc25 核膜崩壊
CDK1
中心体の分離 ゴルジ体再構築
F i g . 7 . G 2 a n d M P h a s e s o f C e l l C y c l e
第 三 章 血 管 新 生 と が ん
血 管 は , 全 身 に 分 布 し 血 液 の 灌 流 を 可 能 に す る 生 体 に と っ て 基 本 的 な 組 織 の 一 つ で あ る . 血 管 は , 内 腔 を 覆 う 一 層 の 血 管 内 皮 細 胞 と 血 管 基 底 膜 , そ の 周 囲 を 取 り 囲 む 壁 細 胞 ( 動 静 脈 に お い て は 平 滑 筋 細 胞 , 毛 細 血 管 に お い て は 周 皮 細 胞 ( ペ リ サ イ ト )) に よ り 構 成 さ れ て い る
( F i g . 8 ).
生 体 内 で 血 管 が 形 成 さ れ る 現 象 は , 脈 管 形 成 ( v a s c u l o g e n e s i s ) と 血 管 新 生 ( a n g i o g e n e s i s ) の 二 つ に 大 別 さ れ る . 脈 管 形 成 は , 胎 生 期 に お い て 胎 児 外 の 卵 黄 嚢 に 血 島 と 呼 ば れ る 細 胞 集 団 が 形 成 さ れ る こ と に 始 ま る . 血 島 の 内 側 の 細 胞 は 血 球 に 分 化 し , 外 側 付 近 の 細 胞 は 内 皮 細 胞 に 分 化 し て 原 始 血 管 叢 を 形 成 す る ( F i g . 9 ). 一 方 , 血 管 新 生 と は 既 存 の 血 管 か ら 新 た な 血 管 ネ ッ ト ワ ー ク が 形 成 さ れ る 現 象 の こ と で あ る . こ の 血 管 新 生 の 過 程 は , 脈 管 形 成 の 終 了 後 , 臓 器 中 に 血 管 が 張 り 巡 ら さ れ る 際 に 観 察 さ れ る が , 成 熟 固 体 の 生 理 的 , 病 的 血 管 新 生 で も 同 様 の 機 序 が 観 察 さ れ る .
8)血 管 新 生 の 生 物 学 的 な 基 礎 研 究 は , 固 形 腫 瘍 の 発 育 が 腫 瘍 血 管 新 生 に 依 存 し て い る と の 仮 説 が さ れ 開 始 さ れ た .近 年 , 血 管 の 分 子 生 物 学 の め ざ ま し い 発 展 と と も に , 血 管 新 生 は が ん の 増 殖 と 遠 隔 転 移 に 深 く 関 与 し て い る こ と , ま た , 血 管 新 生 に は 多 く の 因 子 が 関 与 す る こ と が 明 ら か に な っ た .
8 , 9)血 管 新 生 は , ① ペ リ サ イ ト の 離 解 , ② 細 胞 外 マ ト リ ッ ク ス の 消 化 ,
③ 内 皮 細 胞 の 遊 走 と 増 殖 , ④ 内 皮 細 胞 に よ る 管 腔 形 成 , ⑤ ペ リ サ イ ト
に よ る 血 管 の 成 熟 化 の プ ロ セ ス か ら な る . 通 常 で は , ペ リ サ イ ト か ら
分 泌 さ れ る A n g - 1 ( a n g i o p o i e t i n - 1 ) が 内 皮 細 胞 上 の 受 容 体 T I E 2
( t y r o s i n e k i n a s e w i t h I g a n d E G F h o m o l o g y d o m a i n ) に 結 合 し , 内 皮 細 胞 か ら P D G F ( p l a t e l e t - d e r i v e d g r o w t h f a c t o r ; 血 小 板 由 来 増 殖 因 子 ) が 産 生 さ れ る . こ の P D G F の 刺 激 に よ り , ペ リ サ イ ト が 内 皮 細 胞 を 取 り 囲 む よ う に 集 積 す る と と も に , 細 胞 外 マ ト リ ッ ク ス を 介 し て 血 管 構 造 を 安 定 化 し て い る .し か し ,低 酸 素 や 低 栄 養 状 態 に な る と , 内 皮 細 胞 や 周 囲 の 間 葉 系 細 胞 か ら A n g - 1 の ア ン タ ゴ ニ ス ト の A n g - 2
( a n g i o p i e t i n - 2 ) の 産 生 が 高 ま る . A n g - 2 が T I E 2 と 結 合 し , 一 過 性
に T I E 2 を 不 活 性 化 さ せ て 下 流 の シ グ ナ ル 伝 達 を 競 合 的 に 阻 害 す る と ,
ペ リ サ イ ト は 内 皮 細 胞 と の 接 着 を 失 い , 離 解 す る . ペ リ サ イ ト が 離 解 し た 部 分 の 内 皮 細 胞 は , 血 管 新 生 促 進 因 子 の 刺 激 を 受 け や す く な る . 血 管 新 生 促 進 因 子 の 中 で も 特 に , V E G F ( v a s c u l a r e n d o t h e l i a l g r o w t h
f a c t o r ; 血 管 内 皮 増 殖 因 子 ) が 重 要 で あ る . 内 皮 細 胞 同 士 は V E - カ ド
ヘ リ ン と い う 細 胞 間 接 着 因 子 に よ り 接 着 し て い る が , V E G F の 刺 激 に
よ り V E - カ ド ヘ リ ン が リ ン 酸 化 さ れ る こ と で 内 皮 細 胞 同 士 の 接 着 が 弱
ま る . さ ら に , V E G F の 刺 激 を 受 け た 内 皮 細 胞 は 種 々 の プ ロ テ ア ー ゼ を 産 生 し , 基 底 膜 や 周 囲 の 細 胞 外 マ ト リ ッ ク ス を 消 化 す る . 内 皮 細 胞 は 接 着 因 子 で あ る イ ン テ グ リ ン を 介 し て 細 胞 外 マ ト リ ッ ク ス に 接 着 し , 接 着 方 向 に 向 か っ て 遊 走 す る . 基 底 膜 を 破 り , 間 質 へ と 浸 潤 し た 内 皮 細 胞 は , 遊 走 と 増 殖 に よ っ て そ の 細 胞 数 を 増 加 し て い く . こ の 際 , 隣 接 し た 内 皮 細 胞 同 士 は V E - カ ド ヘ リ ン に よ り 接 着 す る . こ の よ う に し て 形 成 さ れ た 未 熟 な 血 管 は ,ペ リ サ イ ト が 接 着 す る こ と で 成 熟 化 す る .
A n g - 1 が T I E 2 に 結 合 し , 内 皮 細 胞 が 刺 激 さ れ て P D G F の 産 生 を 促 進
す る こ と で , ペ リ サ イ ト は 内 皮 細 胞 に 続 い て 遊 走 , 増 殖 し , 新 生 血 管
の 周 囲 を 取 り 囲 む . ペ リ サ イ ト か ら 産 生 さ れ た T G F - ( t r a n s f o r m i n g
g r o w t h f a c t o r ) に よ り 内 皮 細 胞 の 遊 走 と 増 殖 は 停 止 し , 新 た な 基 底
膜 が 形 成 さ れ , 成 熟 し た 血 管 が 構 築 さ れ る ( F i g . 1 0 - a ).
腫 瘍 血 管 新 生 は 正 常 な 血 管 新 生 と 異 な る 部 分 が あ る . が ん 細 胞 は , は じ め は 自 身 に よ り 1 - 2 m m
3ま で 成 長 す る が , そ れ 以 上 の 大 き さ に な る と が ん 細 胞 の 周 囲 で は 低 酸 素 状 態 と な り ,二 酸 化 炭 素 ,一 酸 化 窒 素 ,
C O X - 2 な ど の 上 昇 が 起 こ る .こ の う ち 特 に 低 酸 素 状 態 を 引 き 金 と し て ,
V E G F を は じ め と す る 様 々 な 血 管 新 生 促 進 因 子 が 分 泌 さ れ , が ん 細 胞
へ 向 か っ て 血 管 の 新 生 が 誘 導 さ れ る . 腫 瘍 血 管 新 生 で は , A n g - 2 の 分 泌 が 優 勢 で あ る た め ペ リ サ イ ト が 欠 如 あ る い は 減 少 し て い る と 考 え ら れ て お り , が ん 細 胞 か ら 分 泌 さ れ る 持 続 的 な V E G F の 刺 激 に よ り , 未 成 熟 な 新 生 血 管 が 構 築 さ れ る プ ロ セ ス を 繰 り 返 す . そ の 中 で , 脆 弱 , 不 規 則 , 無 秩 序 , 透 過 性 亢 進 な ど の 特 徴 を も つ 腫 瘍 血 管 が 形 成 さ れ て い く . こ う し て 新 た な 血 管 を 形 成 し た が ん 細 胞 は , 栄 養 素 や 酸 素 を 取 り 込 み , 二 酸 化 炭 素 や 乳 酸 な ど の 老 廃 物 を 排 泄 し て , 加 速 度 的 に 成 長 す る と と も に , 新 た な 組 織 へ と 転 移 す る ( F i g . 4 - b ).
8 )F i g . 8 . A n a t o m y o f B l o o d Ve s s e l
6 )F i g . 9 . P r o c e s s o f Va s c u l o g e n e s i s
4 )a ) P h y s i o l o g i c a l c o n d i t i o n b ) P a t h o l o g i c a l c o n d i t i o n
F i g . 1 0 . P r o c e s s o f P h y s i o l o g i c a l a n d P a t h o l o g i c a l A n g i o g e n e s i s
第 四 章 V E G F と V E G F 受 容 体
血 管 新 生 は 一 般 に , 促 進 因 子 と 抑 制 因 子 の バ ラ ン ス に よ っ て 制 御 さ れ る . 実 際 に , 様 々 な 腫 瘍 に お い て 血 管 新 生 促 進 因 子 の 発 現 亢 進 と 抑 制 因 子 の 発 現 低 下 が 確 認 さ れ て い る .促 進 因 子 の 一 つ で あ る V E G F は そ の 受 容 体 , V E G F R ( V E G F r e c e p t o r ; V E G F 受 容 体 ) を 介 し て , 血 管 新 生 に お い て 重 要 な 役 割 を 担 っ て い る .
1 0)第 一 節 V E G F
V E G F は , 血 管 内 皮 細 胞 特 異 的 な 増 殖 促 進 作 用 と 血 管 透 過 性 亢 進 作
用 を も つ 因 子 と し て 単 離 さ れ た 分 子 量 2 3 k D の サ ブ ユ ニ ッ ト が ジ ス ル フ ィ ド 結 合 に よ り ホ モ ダ イ マ ー を 形 成 し た 糖 タ ン パ ク 質 で あ る . V E G F フ ァ ミ リ ー と し て V E G F - A , P I G F ( p l a c e n t a g r o w t h f a c t o r ; 胎 盤 由 来 増 殖 因 子 ), V E G F - B , V E G F - C , V E G F - D , パ ラ ポ ッ ク ス ウ イ ル ス 由 来 の V E G F - E , ヘ ビ 毒 由 来 の V E G F - F が 知 ら れ て い る . ア ミ ノ 酸 配 列 の 相 同 性 は 3 0 - 5 0 % 程 度 で あ る が , 8 つ の シ ス テ イ ン 残 基 が 保 存 さ れ て い る と い う 特 徴 を も つ . ヒ ト に は 2 3 対 の 染 色 体 が あ り ,
ヒ ト V E G F 遺 伝 子 は 7 番 染 色 体 上 に 存 在 す る . 遺 伝 子 D N A は 8 つ の
エ ク ソ ン か ら な り , D N A を 鋳 型 と し て R N A が 合 成 さ れ る が , 転 写 の 際 の ス プ ラ イ シ ン グ の 違 い に よ り 大 き さ の 異 な る m R N A が 産 生 さ れ
る .
7 , 8)V E G F - A は 単 に V E G F と も 呼 ば れ , 血 管 新 生 に 最 も 関 わ り が
深 い と い わ れ る . V E G F - A は ス プ ラ イ シ ン グ の 違 い に よ り , ヒ ト ゲ ノ
ム に お い て 9 つ の ア イ ソ フ ォ ー ム が 確 認 さ れ て お り , 中 で も
V E G F - A
1 6 5は 最 も 豊 富 に 存 在 す る ( F i g . 11 ).
1 1 , 1 2)V E G F は ほ と ん ど の 臓 器 を は じ め , 上 皮 系 細 胞 , 繊 維 芽 細 胞 , 平 滑 筋 細 胞 , 心 筋 細 胞 な ど 様 々 な 細 胞 や , 多 く の 固 形 腫 瘍 と そ れ に 由 来 す る 細 胞 株 に お い て 発 現 し て い る . ま た , 低 酸 素 状 態 に 反 応 し て 転 写 が 誘 導 さ れ る と い う 大 き な 特 徴 が あ り , V E G F の 転 写 開 始 点 の 上 流 に は H I F - 1 ( h y p o x i a - i n d u c i b l e f a c t o r - 1 ; 低 酸 素 誘 導 因 子 ) が 結 合 す る 配 列 が 存 在 す る .
8)F i g . 11 . C o m p a r i s o n o f S t r u c t u r e s o f t h e V E G F F a m i l y
1 1 , 1 2 )第 二 節 V E G F 受 容 体
V E G F R に は , V E G F R - 1 ( F l t - 1 ), V E G F R - 2 ( K D R / F l k - 1 ), V E G F R - 3
( F l t - 4 ) の 3 種 が 存 在 す る . こ れ ら は , 細 胞 外 ド メ イ ン , 膜 貫 通 ド メ
イ ン , チ ロ シ ン キ ナ ー ゼ ド メ イ ン を も つ 典 型 的 な 受 容 体 型 チ ロ シ ン キ ナ ー ゼ で あ る . V E G F R - 1 お よ び 2 は , 細 胞 外 ド メ イ ン 内 に 7 つ の I g 様 構 造 を も ち , キ ナ ー ゼ ド メ イ ン 内 に は 約 7 0 の ア ミ ノ 酸 の キ ナ ー ゼ 挿 入 領 域 を も つ . V E G F R - 3 も 類 似 の 構 造 を も つ が , 5 番 目 の I g 様 ド メ イ ン が 開 裂 し , ジ ス ル フ ィ ド 結 合 で 架 橋 さ れ て い る .
9 , 1 1)V E G F R は そ の リ ガ ン ド で あ る V E G F お よ び P I G F が 結 合 す る こ と
で 活 性 化 す る が ,V E G F - A は V E G F R - 1 お よ び 2 と 結 合 す る .V E G F - C お よ び D は V E G F R - 2 お よ び 3 と 結 合 す る が ,P I G F ,V E G F - B お よ び V E G F - E は そ れ ぞ れ V E G F R - 1 , 2 に 特 異 的 に 結 合 す る ( F i g . 1 2 ).
S SS S
VEGFR-1
(Flt-1) VEGFR-1
(Flt-1) VEGFR-2 (KDR/Flk-1)
VEGFR-2
(KDR/Flk-1) VEGFR-3 (Flt-4) VEGF-A
VEGF-B PIGF
VEGF-A VEGF-C VEGF-D VEGF-E VEGF-A
VEGF-B PIGF
VEGF-A VEGF-C VEGF-D VEGF-E
VEGF-C VEGF-D
Monocytes
Macropharges Vascular endothelium Lymphatic endothelium
F i g . 1 2 . I n t e r a c t i o n s o f V E G F F a m i l y w i t h T h e i r R e c e p t o r s
1 3 )こ の う ち , 血 管 新 生 に 最 も 関 わ り が 深 い の は 血 管 内 皮 細 胞 に 発 現 し て い る V E G F R - 2 で あ り , 2 番 目 と 3 番 目 の I g 様 ド メ イ ン を 介 し て
V E G F - A と 結 合 し , 受 容 体 の 二 量 体 化 が 起 こ る . 受 容 体 が 相 互 に リ ン
酸 化 し あ え る 距 離 ま で 近 づ き , キ ナ ー ゼ ド メ イ ン が 活 性 化 さ れ る と , 受 容 体 の 自 己 リ ン 酸 化 が 起 こ り 細 胞 内 へ と シ グ ナ ル が 伝 達 さ れ る . V E G F R - 2 の 主 な 自 己 リ ン 酸 化 部 位 に は 9 5 1 ,1 0 5 4 ,1 0 5 9 ,11 7 5 ,1 2 1 4 番 目 の チ ロ シ ン 残 基 ( Y 9 5 1 , Y 1 0 5 4 , Y 1 0 5 9 , Y 11 7 5 , Y 1 2 1 4 ) が あ る . リ ン 酸 化 さ れ た Y 11 7 5( p Y 11 7 5 ) は P L C ( p h o s p h o l i p a s e C - ; ホ ス ホ リ パ ー ゼ C ) 経 路 に よ り 血 管 内 皮 細 胞 増 殖 の シ グ ナ ル 伝 達 を 行 う . P L C は そ の S H 2 ド メ イ ン を 介 し て Y 11 7 5 と 結 合 し , P I P 2
( p h o s p h a t i d y l i n o s i t o l 4 , 5 - b i s p h o s p h a t e; ホ ス フ ァ チ ジ ル イ ノ シ ト ー ル 4 , 5 ‐ 二 リ ン 酸 ) を D A G ( d i a c y l g l y c e r o l ; ジ ア シ ル グ リ セ ロ ー ル ) と I P 3 ( i n o s i t o l 1 , 4 , 5 - t r i s p h o s p h a t e ; イ ノ シ ト ー ル 1 , 4 , 5 ‐ 三 リ ン 酸 ) に 加 水 分 解 す る . D A G は P K C ( p h o s p h o k i n a s e C ; プ ロ テ イ ン キ ナ ー ゼ C ) を , P K C は R a s を 介 す る こ と な く R a f を 活 性 化 し , M A P K ( m i t o g e n - a c t i v a t e d p r o t e i n k i n a s e ; 分 裂 促 進 因 子 活 性 化 タ ン パ ク 質 キ ナ ー ゼ )カ ス ケ ー ド で あ る R a f - M E K - E R K 経 路 を 介 し て シ グ ナ ル を 核 内 へ と 伝 え る . 活 性 化 さ れ た E R K は 核 内 へ 移 行 し , 標 的 で あ る 転 写 因 子 E l k 1 を 活 性 化 し て 遺 伝 子 転 写 を 行 い , 細 胞 増 殖 の シ グ ナ ル を 伝 達 す る . p Y 11 7 5 は さ ら に , ア ダ プ タ ー 分 子 の S h b に も 結 合 す る . S h b を 介 し て P I 3 K( p h o s p h a t i d y l i n o s i t o l - 3 k i n a s e; ホ ス フ ァ チ ジ ル イ ノ シ ト ー ル 3 - キ ナ ー ゼ ) が リ ン 酸 化 さ れ る と , 細 胞 の 遊 走 を 調 節 す る 接 着 斑 の 会 合 が 誘 導 さ れ る . p Y 1 2 1 4 は G r b 2 の S H 2 ド メ イ ン を 介 し て G r b 2 に 結 合 す る . Y 1 2 1 4 の リ ン 酸 化 は C d c 4 2 や
p 3 8 M A P K の 活 性 化 に 必 要 で あ り , ア ク チ ン の 再 構 成 を 引 き 起 こ す .
p Y 9 5 1 は T S A d ( T- c e l l - s p e c i f i c a d a p t o r p r o t e i n ) と 結 合 す る . Y 9 5 1 を 介 し た V E G F R - 2 と T S A d の 結 合 も ま た , ア ク チ ン の 再 構 成 や 細 胞 の 遊 走 に 必 要 不 可 欠 で あ る ( F i g . 1 3 ).
PIP2 DAG
1175 PLC-
P
+ IP3
Ca2+
release
PKC
Raf
MEK
ERK Gene transcription Elk1
Cell proliferation
PIP2
1175 TSAd 951
P
PIP3 PI3K
Src
PShb FAK
Paxillin Rac
Rho
Focal adhesion turn over
Actin reorganization
Cell migration 1214
Grb2P
Cdc42 MAPKp38
FAK
Actin reorganization VEGFR-2
VEGF-A
VEGFR-2 VEGF-A
F i g . 1 3 . V E G F - A / V E G F R - 2 S i g n a l i n g i n Va s c u l a r E n d o t h e l i a l C e l l P r o l i f e r a t i o n a n d M i g r a t i o n
1 3 )V E G F R は V E - カ ド ヘ リ ン , - カ テ ニ ン , P I 3 K と 複 合 体 を 形 成 す る
こ と で , 内 皮 細 胞 の 生 存 に も 関 与 す る . P I 3 K の 活 性 化 に よ り P I P 3
( p h o s p h a t i d y l - i n o s i t o l 3 , 4 , 5 - t r i s p h o s p h a t e ; ホ ス フ ァ チ ジ ル イ ノ
シ ト ー ル 3 , 4 , 5 - 三 リ ン 酸 ) が 増 加 す る と A k t が 活 性 化 さ れ る . A k t の
活 性 化 は B A D や カ ス パ ー ゼ 9 に 作 用 し , ア ポ ト ー シ ス 促 進 経 路 を 阻
害 す る こ と で 細 胞 の 生 存 を 引 き 起 こ す ( F i g . 1 4 ).
1 3)PIP2 VE-cadherin PIP3
PI3K
-catenin
Akt
Caspase 9
Cell survival BAD
VEGFR-2 VEGF-A