Kobe University Repository : Thesis
学位論文題目
Title
キシロオリゴ糖の微生物生産と関与する酵素群の精製および特性解析
氏名
Author
袁, 清珠
専攻分野
Degree
博士(農学)
学位授与の日付
Date of Degree
2008-09-25
資源タイプ
Resource Type
Thesis or Dissertation / 学位論文
報告番号
Report Number
甲4420
権利
Rights
JaLCDOI
URL
http://www.lib.kobe-u.ac.jp/handle_kernel/D1004420
※当コンテンツは神戸大学の学術成果です。無断複製・不正使用等を禁じます。著作権法で認められている範囲内で、適切にご利用ください。PDF issue: 2019-03-09
博 士 論 文
キシロオリゴ糖の微生物生産と関与する
酵素群の精製および特性解析
平成
20
年
8
月
神戸大学大学院自然科学研究科
哀 清 珠
目 次
序 論
1第 1 章
Bacillus pumilus X-6-19 お よ び そ の 変 異 株 に よ る 長 鎖キ シ ロ オ リ ゴ 糖 の 生 産 3 第 1 節 序 3 第 2 節 キ シ ロ オ リ ゴ 糖 高 生 産 の た め の 培 養 条 件 の 検 討 3 第 3 節 紫 外 線 照 射 に よ る キ シ ロ オ リ ゴ 糖 高 生 産 性 変 異 株 の 取 得 11 第 4 節 N-methyl-N-nitro-N-nitorosoguanidine( NTG) に よ る キ シ ロ オ リ ゴ 糖 高 生 産 性 変 異 株 の 取 得 13 第 5 節 キ シ ロ オ リ ゴ 糖 高 生 産 性 株 の 選 択 15 第 6 節 要 約 と 考 察 15
第 2 章
B. pumilus X-6-19 strain U-3 株 の 生 産 す る キ シ ラ ナ ー ゼ の精 製 と 特 性 解 析 17 第 1 節 序 17 第 2 節 酵 素 活 性 測 定 法 17 第 3 節 酵 素 の 生 産 条 件 の 検 討 18 第 4 節 酵 素 の 精 製 20 第 5 節 精 製 酵 素 の 特 性 24 第 6 節 要 約 と 考 察 32
第 3 章
B. pumilus X-6-19 strain U-3 株 の 生 産 す る β -キ シ ロ シ ダ ー ゼ の精 製 と 特 性 解 析 34 第 1 節 序 34 第 2 節 酵 素 活 性 測 定 法 34 第 3 節 酵 素 の 生 産 条 件 の 検 討 34 第 4 節 酵 素 の 精 製 36 第 5 節 精 製 酵 素 の 特 性 44 第 6 節 キ シ ラ ナ ー ゼ と β D-グ ル コ ー ス の
第 7 節 要 約 と 考 察 51
第 4 章 キ シ ラ ナ ー ゼ に よ る キ シ ロ オ リ ゴ 糖 の 生 産
52 第 1 節 序 52 第 2 節 酵 素 反 応 の 条 件 52 第 3 節 キ シ ロ オ リ ゴ 糖 の HPLC に よ る 分 析 52 第 4 節 要 約 と 考 察 54総 括
55要 旨
58参 考 文 献
60謝 辞
63序論 バイオ マス とは、 燃料 や種々 の工 業製品 の原 料とし て使 用され る動 物,植物,微生物 由来の生物資源の総称である。なかでも、植物由来セルロース系バイオマスは地球上 に最も多く存在する植物繊維性物質であるが、その殆どが利用されることなく、廃棄 物となっている。バイオマスは再生産可能な有機資源であり、燃焼による有害ガスの 排出量が少なく、また貯蔵性に優れている事などから近年、エネルギー対策や地球環 境汚染改善の観点から注目されるようになった。これらの未利用バイオマスは、微生 物などを用いて低分子化することにより、有効に利用されることが期待される 1)。 キシランはセルロース系バイオマス資源の一つであり、キシロースが β-1,4 結合で 重合した主鎖にアラビノフラノースやグルクロン酸などの側鎖が結合した構造を持つ 多糖類である。これまでに、キシランを有効利用するために、キシランを分解する微 生物酵素を用いた研究が多数行われてきた2-4)。 キシラン分解酵素には、キシラナーゼ(endo-1,4-β-xylanase; EC.3.2.1.8)と β-キシロ シダーゼ(exo-1,4-β-xylosidase; EC.3.2.1.37)があるが、キシラナーゼは、キシラン主 鎖のβ-1,4-D-キシロシド結合を分子の内部からアトランダムに加水分解してキシロオ リゴ糖を生成するのに対して(endo 型)、β-キシロシダーゼは、キシランの非還元性 末端部に作用し、キシロースを生成する(exo 型)5)。 キシランを分解して得られるキシロオリゴ糖は、特定の腸内細菌に対して増殖を促 進する作用があり、整腸作用を有する特定保健用食品として認定された乳酸菌飲料や チョコレートなどに利用される有用な糖類である6-8)。また、甘味料、保湿剤としての 食品添加剤、家畜の飼料などの用途もある。さらに、医薬品やサニタリー製品の分野 において乳化剤や皮膚の保湿剤として使用されている。 一般に、特定保健用食品に用いられるオリゴ糖類は、そのほとんどが腸内悪玉菌で ある大腸菌や腸内腐敗発酵菌の数を減少させ、相対的に腸内善玉菌であるビフィズス 菌の数を増加させるなどの整腸作用を持つ。なかでもキシロオリゴ糖は他のオリゴ糖 と比較して胃酸に対する耐性が強く、低分子化されることなく腸内に到達するため、 他のオリゴ糖と比較して整腸効果が高いとされている 9,10)。キシロオリゴ糖を摂取す ることにより得られるビフィズス菌の選択的増殖促進効果は、キシロオリゴ糖の鎖長 が長いほど優れていると言われ、特に3 量体以上 10 量体程度までのキシロオリゴ糖が この選択的増殖性に有効である 11)。 従来、キシロオリゴ糖は、微生物酵素によるキシランの加水分解(2-4)のほか、キシラ ナーゼと硫酸 11)、アンモニア水 12)、加熱 13) による処理とを組み合わせる方法などに よって製造されてきた。しかし、現状において市販されているキシロオリゴ糖は 2 量
体や 3 量体を主成分とするものがほとんどであり、より鎖長が長いキシロオリゴ糖(長 鎖キシロオリゴ糖)の開発が望まれている。 そこで本研究では、バイオマス資源(キシラン)の有効利用と長鎖キシロオリゴ糖 の有用性の観点から、キシランを原料として、長鎖キシロオリゴ糖を生産する微生物 を用いて、本菌が生産する長鎖キシロオリゴ糖を効率的かつ大量に得ることを目的と して研究を進めた。 微生物をキシロオリゴ糖の製造に利用する場合、従来は上述のように微生物の生産 するキシラナーゼを主として用い、キシランからキシロオリゴ糖を生産していた。も し、微生物の培養そのもの(growing cells)を利用して、キシランからキシロオリゴ糖 が生産できれば、キシラナーゼを使用するより有利な点が多い。まず、キシラナーゼ を精製する必要がないことである。キシラナーゼは一般に、細胞外酵素であるから細 胞内酵素に比べて精製は容易であるが、精製酵素を得るためにはかなりの時間と労力 を要する。また、優れた野生株や変異株が得られれば、キシラン以外の安価な基質を 炭素源およびエネルギー源として増殖させ、同じ培地に共存させたキシランから効率 的にオリゴ糖を生産することが可能となる。 本研究では、こうした点をふまえ、まず、微生物培養液中にキシロオリゴ糖を生産・ 蓄積する株の培養条件の検討を行った。得られた菌株を変異処理し、効率的にキシロ オリゴ糖を培養液中に蓄積する変異株を得た。同変異株の生産するキシラナーゼを精 製し、精製したキシラナーゼを用いたキシロオリゴ糖の製造法も検討した。 本論文は、序論、第1∼4 章および総括より構成される。第 1 章では Bacillus pumilus X-6-19 によるキシロオリゴ糖高生産のための培養条件の検討と、キシロオリゴ糖高生 産性変異株の取得について述べる。第 2 章では前章で得られたキシロオリゴ糖高生産 性変異株 B. pumilus X-6-19 strain U-3 株が生産するキシラナーゼの精製と特性を述べ る。第 3 章では B. pumilus X-6-19 strain U-3 が生産する β-キシロシダーゼの精製と特性 を述べる。第4 章ではキシラナーゼによるキシロオリゴ糖の生産について述べる。さ らに、以上の結果を総括において総合的に考察する。
第 1 章 Bacillus pumilus X-6-19 およびその変異株による長鎖キシロオリゴ糖の生産
第1節 序 本章では、Bacillus pumilus X-6-19 を培養した時、培養液中に目的とする長鎖キシロ オリゴ糖を多量に生産するような培養条件を検討することを目的とした。菌株の分離 に用いた培地(Table 1-1)は、唯一の炭素源、窒素源としてキシランを含むが、本菌 が他の炭素化合物を炭素源、窒素源として利用できれば、キシランからキシロオリゴ 糖の生産効率が上昇する可能性がある。また、本章では、本菌を変異処理し、長鎖キ シロオリゴ糖を多量に生産・蓄積する変異株の取得を試みた。 第2節 キシロオリゴ糖高生産のための培養条件の検討 【方法】 (1)培地の調製
Table 1-1 に実験に用いる培地の組成を示す。A 液と B 液は別々に調製した。但し、A 液中のキシラン(Sigma、St. Louis、USA)は、キシランを除く他の成分を調製して各 試験管に分注した後、当該試験管に固体の状態で加えた。オートクレーブ(121℃、13 分)にて殺菌して冷却した後、A、B 液を無菌的に混合して培地を作製した。固体培 地は A 液に 1.5 g の寒天を添加して調製した。
Table 1-1. Medium for culture
Solution A
Oat spelt xylan 1.0 g
Yeast extract 0.05 g NH4Cl 0.2 g KH2PO4 0.4 g NaCl 0.2 g (Agar) 1.5 g H2O 86 mL Solution B MgSO4・7H2O 0.1 g CaCl2・2H2O 0.02 g FeSO4・7H2O 5.0 g MnSO4・4-5H2O 7.7 g H2O 14 mL pH 7.0 (2)還元糖の定量法 1.試薬の調製14) ・Somogyi 試薬 酒石酸カリウムナトリウム 12 g と炭酸ナトリウム 24 g を 250 mL の水に溶解した。 4.0 g の硫酸銅五水和物を約 40 mL の水に溶かしたものを撹拌しながら加えた。次に、 16 g の炭酸水素ナトリウムを加えて溶解した。180 g の硫酸ナトリウムを 500 mL の水 に溶解した後、煮沸して溶存酸素を除き、放冷した。次に両液を合わせ、水を加えて 1 L とした。37℃で 2∼3 日間放置した後、沈殿を吸引濾過し、濾液を褐色共栓瓶に入 れ、暗所にて保管した。 ・Nelson 試薬 モリブデン酸六アンモニウム四水和物25 g を 450 mL の水に溶解し、濃硫酸 21 mL を徐々に加えた。次に、ヒ酸二ナトリウム七水和物 3 g を 25 mL の水に溶解したもの
を加えて、37℃で 24∼48 時間放置した後、褐色共栓瓶に入れ、暗所にて保管した。 2.還元糖の定量方法 Somogyi-Nelson 法 14)を 用 い た 。 本 菌 株 を 培 養 し て 得 ら れ た 培 養 上 清 0.2 mL に Somogyi 試薬 0.2 mL を加えて 100℃で 10 分間加熱した後、氷中にて 3 分間冷却した。 冷却後、Nelson 試薬 0.2 mL を加え、さらに水 2 mL を加えて混合した。30 分間放置し た後、 日立 U-2000 分光光度計(Hitachi、Tokyo、Japan)を用いて 500 nm における 吸光度を測定した。標準曲線は D-キシロース(Nacalai Tesque、Kyoto、Japan)を用い て作成した。 (3)還元糖の組成分析 1.培養液の調製 前培養は、Table 1-1 に示す液体培地 7 mL を用いて、30℃、210 rpm の条件で 24 時 間振盪培養した。得られた培養液14 mL(試験管 2 本分)を、同培地 56 mL を含む 500 mL の振盪フラスコに加え、30℃、130 rpm の条件で振盪培養した。得られた培養液を 遠心分離(20,000×g、10 分、4℃)し、培養上清を以下の精製操作に用いた。 2.80%(v/v)エタノールによる可溶性画分の抽出 (1)で得られた100 mL の培養上清(500 mL コルベン 2 本分)を 100 mL ナス型 フラスコに入れ、エバポレーターにて減圧乾固した。200 mL の 80% (v/v) エタノール を数回に分けて加え、70℃湯浴中で攪拌しながら、可溶性画分を抽出した。抽出した 可溶性画分を再び減圧乾固した後、10 mL の脱塩水に溶解し、フラクション 1 とした。 3.カーボンカラムクロマトグラフィー
フラクション1 を、1N 塩酸で洗浄した活性炭素(Wako Pure Chemicals、Osaka、Japan) を充填したカラム(2.0×15 cm)に供した。カラムを 150 mL の水で洗浄した後、0∼80% (v/v)エタノールを用いてリニアグラジエント法により、流速 60 mL/h で吸着した還 元糖を溶出した。各フラクション(5 mL/tube)における還元糖の定量と、組成分析を 行った。還元糖の組成分析は下記に示すTLC により行った。高濃度な還元糖画分を集 め、フラクション2 とした。 4.ゲル濾過クロマトグラフィー15) フラクション2 を減圧乾固して 15 mL の水に溶解した後、10% (v/v) エタノールで 平衡化した Bio Gel P-2 Extra Fine(Bio-Rad、Richmond、USA)を充填したカラム(3.2×62
cm)に供した。500 mL の 10% (v/v) エタノールを用いて、流速 20 mL/hr で還元糖を 溶出した。各フラクション(2 mL/tube)における還元糖の定量と、TLC による組成分 析を行った。 5.TLC による還元糖の組成分析 a.試薬の調製 ・展開溶媒 エタノール-n-ブタノール-水(5:5:3、v/v)を用いた。 ・発色試薬 2-クロム酸カリウム 3.4g を水 20 mL に溶解し、濃硫酸 10 mL を加えて調製した。 b.還元糖の組成分析 TLC アルミニウムシート(シリカゲル 60、Merck、Darmstadt、Germany)に培養上 清をキャピラリーで10 回スポットし、上記の展開溶媒を用いて 2 回展開した。展開後、 十分に乾燥し、発色試薬をプレートに噴霧した。プレートを 100∼110℃で 5 分間加熱 し、発色したスポットのRf値を求めた。マーカーとして D-キシロース、キシロオリゴ 糖混合物を用いた。 (4)キシロオリゴ糖の生産に与える種々炭素源の影響 Table 1-1 に示す液体培地 7.0mL に、種々の炭素源を添加した培地を調製した。用い た炭素源(終濃度 1% (w/v))は、D-グルコース、スクロース、ラクトース一水和物、 クエン酸三ナトリウム二水和物、クエン酸二ナトリウム六水和物、酢酸ナトリウム三 水和物、フマル酸、マレイン酸、グリセロース、DL-リンゴ酸の計 10 種であった。そ れぞれ調製した培地において本菌を振盪培養(30℃、210 rpm)し、24 時間ごとに 0.5 mL ずつサンプリングした。遠心分離(20,000xg、10 分、4℃)後、上清中の還元糖の 定量と組成分析を行った。 (5)キシロオリゴ糖の生産に与える酢酸ナトリウム三水和物の影響 Table 1-1 に示す液体培地 7 mL に、終濃度が 0、0.2、0.5、0.8、1、3、5% (w/v)とな るように酢酸ナトリウム三水和物を添加し、それぞれの培地で本菌を振盪培養(30℃、 210 rpm)した。24 時間ごとに 0.5 mL ずつサンプリングし、遠心分離(20,000xg、10 分、4℃)後、上清中の還元糖の定量と TLC による組成分析を行った。 (6)キシロオリゴ糖の生産に与える種々窒素源の影響
1% (w/v) 酢酸ナトリウム三水和物を添加した 1% (w/v) キシラン培地 7 mL に終濃 度が 0.2% (w/v)となるように、硫安、NH4NO3、Urea、NaNO3、NaNO2、NH4Cl を添 加し、それぞれの培地で本菌を振盪培養(30℃、210rpm)した。24 時間ごとに 0.5 mL ずつサンプリングし、遠心分離(20,000xg、10 分、4℃)後、上清中の還元糖の定量 と組成分析を行った。 【結果】 (1)キシロオリゴ糖の組成分析 本菌がキシランを加水分解して蓄積するキシロオリゴ糖の、Bio Gel P-2 EF を用い たゲル濾過クロマトグラフィーの結果と、それぞれの画分(ピーク)の還元糖量のピ ークを示すフラクションを TLC に供した時の結果を Fig. 1-1 に示す。オーセンティッ クなキシロオリゴ糖のRf値から推測して、ピークⅠを示すフラクションはキシロース、 ピークⅡはキシロビオース、ピークⅢはキシロトリオース、ピークⅣはキシロテトラ オース、ピークⅤはキシロペンタオース以上の高分子オリゴ糖の混合物であることが 分かった。
Fig. 1-1. Separation of xylooligosaccharides on Bio Gel P-2 (A) and TLC of the separated xylooligosaccharides (B).
(A) The cultural fluid (100 mL) was dried on an evaporator and then the residues were extracted with ethanol. The extracts were then passed through activated charcoal. The fractions containing xylooligosaccharides were chromatographed on Bio Gel P-2.
(B) The separated xylooligosaccharides were detected on a TLC plates. See the text for details.
M: marker; X1: xylose; X2: xylobiose; X3: xylotriose; X4: xylotetraose; X5: xylopentaose.
(2)キシロオリゴ糖の生産に与える種々炭素源の影響
酢酸ナトリウム三水和物の添加により、培養開始24 時間後において、本菌はコント ロールの約 1.5 倍の還元糖を培地中に生産した(Fig. 1-2)。培養液中のキシロオリゴ 糖の組成は、炭素源を加えない培地で培養した場合の組成とほぼ同じであった。コハ ク酸二ナトリウム六水和物とグリセロールを添加した場合は、コントロールとほぼ同
様の還元糖を生産した。マレイン酸、グルコース、ラクトース一水和物を添加した場 合には、培地中に還元糖をほとんど生産しなかった。以上の結果から、以後の実験に は、1% (w/v) 酢酸ナトリウム三水和物を培地中に添加することにした。
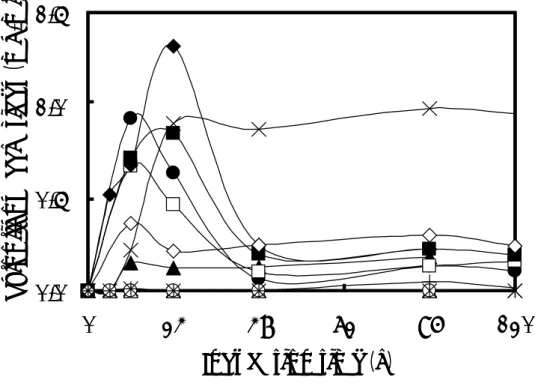
Fig. 1-2. Effects of carbon sources on the production of reducing sugars by B. pumilus X-6-19.
The strain was incubated at 30℃ in 1% (w/v) xylan medium, to which were added D-glucose (○), sucrose (▲), lactose (△), trisodium citrate trihydrate (■), disodium succinate hexahydrate (□), sodium acetate trihydrate (◆), fumaric acid (◇), maleic acid (*), glycerol (×), or DL-malic acid (+). Control (●).
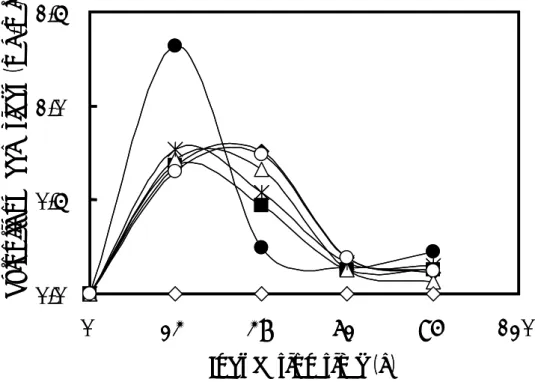
(2)キシロオリゴ糖の生産に与える酢酸ナトリウム三水和物の濃度の影響 添加する酢酸ナトリウム三水和物の濃度が高くなるにつれて、培地中に蓄積する還 元糖量が最大値を示す培養時間は遅くなった(Fig. 1-3)。1% (w/v) 酢酸ナトリウム三 水和物を添加した場合と比較して、いずれの濃度においてキシロオリゴ糖の生産量は 増加しなかった。
0.0
0.5
1.0
1.5
0
24
48
72
96
120
Incubation time(h)
Xy
looligosaccharid
es
(mg
/m
l)
0.0
0.5
1.0
1.5
0
24
48
72
96
120
Incubation time(h)
Xy
looligosaccharid
es
(mg
/m
l)
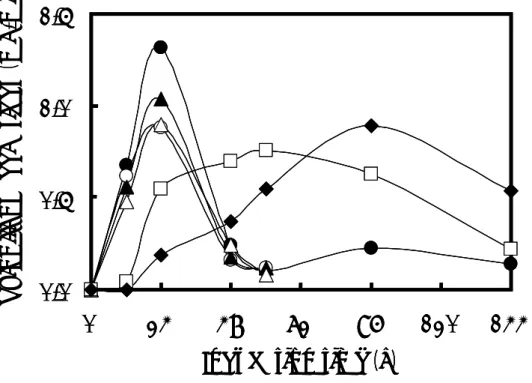
Fig. 1-3. Effects of the concentrations of sodium acetate trihydrate on the production of reducing sugars B. pumilus X-6-19.
B. pumilus X-6-19 was incubated at 30℃ in 1% (w/v) xylan medium with sodium acetate trihydrate at various concentrations. Symbols: ○, 0.2%; ▲, 0.5%; △, 0.8%; ●, 1%; □, 3%; and ◆, 5%. (3)キシロオリゴ糖の生産に与える種々窒素源の影響 添加した窒素源は全て、コントロール(窒素源として NH4Cl を使用)と比較して還 元糖の生産を抑制した(Fig. 1-4)。特に、NaNO2を添加すると、還元糖の生産は完全 に阻害された。
0.0
0.5
1.0
1.5
0
24
48
72
96
120
144
Incubation time(h)
X
ylool
igosaccharid
es
(mg
/m
l)
0.0
0.5
1.0
1.5
0
24
48
72
96
120
144
Incubation time(h)
0.0
0.5
1.0
1.5
0
24
48
72
96
120
144
Incubation time(h)
X
ylool
igosaccharid
es
(mg
/m
l)
Fig. 1-4. Effects of nitrogen sources on the production of reducing sugars by B. pumilus X-6-19.
The strain was incubated at 30℃ in 1% (w/v) xylan medium containing 1% (w/v) sodium acetate trihydrate, to which were added NH4Cl (○), NH4NO3 (△), urea (■), (NH4)2SO4
(◆), NaNO2 (◇), or NaNO3 (*). Control (●).
第3節 紫外線照射によるキシロオリゴ糖高生産性変異株の取得 【方法】 (1)生存率曲線の作成 最も効率的に変異株が出現するのは、生存率が0.1∼1%となるように変異処理した 場合であるとされる16)。そこで、本菌に紫外線照射による変異処理を行った時の、紫 外線照射時間と生存率の関係を示す生存率曲線を作成した。 本菌を Table 1-1 に示す液体培地 7 mL に 1 白金耳植菌し、30℃で、24 時間振盪培養 (210 rpm)した。菌体懸濁液を、滅菌した 0.8% (w/v) NaCl 溶液で希釈した後、希釈 液 1 mL を 9 mL の滅菌した 0.8% (w/v) NaCl 溶液を含むシャーレに加え、静かに攪拌 した。シャーレは紫外線ランプの真下に置き、常にランプからシャーレまでの距離が 30 cm に保たれるようにした。また紫外線を安定化するため、使用する 30 分前からラ ンプをつけておいた。紫外線照射開始後、5 分おきに菌懸濁液を 100 µl ずつサンプリ
0.0
0.5
1.0
1.5
0
24
48
72
96
120
Incubation time(h)
X
ylooli
gosacch
arides
(mg/
m
l)
0.0
0.5
1.0
1.5
0
24
48
72
96
120
Incubation time(h)
X
ylooli
gosacch
arides
(mg/
m
l)
0.1
1
10
100
0
5
10
15
20
Time(min)
Sur
vi
va
l ra
te
(%
)
0.1
1
10
100
0
5
10
15
20
Time(min)
Sur
vi
va
l ra
te
(%
)
ングし、Table 1-1 に示す平板(固体)培地に塗抹した。光による変異株の復帰を防ぐ ため、塗抹後は素早く平板をインキュベーターに入れた。コロニーが出現するまで 30℃で培養し、紫外線照射時間とコロニー数により生存率曲線を作成した。 (2)長鎖キシロオリゴ糖高生産性変異株の分離 (1)に示す方法により 15 分間変異処理した菌懸濁液 100 µl を、0.25% (w/v) D-グ ルコース、0.25% (w/v) 酢酸ナトリウム三水和物を含む 0.25% (w/v) キシラン平板培 地に塗抹し、30℃で培養した。D-グルコース存在下でもキシランに対して強い分解能 力を示すハロー形成能の高い菌株を選択し、Table 1-1 に示す液体培地 7 mL に 1 白金 耳植菌し、30℃で振盪培養(210rpm)した。培養開始後 6、12、18、24 時間目にサン プリングし、遠心分離(20,000xg、10 分、4℃)後、上清中に含まれる還元糖の定量 と組成分析を行った。 【結果】 (1)生存率曲線の作成 Fig. 1-5 に示すように、本菌は紫外線照射時間が 15 分の時、生存率が 1.0%になる ことが分かった。この結果より、以後の実験におけるUV 照射時間を 15 分と決定した。Fig. 1-5. Survival curve of B. pumilus X-6-19 irradiated with UV. Symbols: ●, survival rate (%).
B. pumilus X-6-19 株の菌体懸濁液 1 mL を 9 mL の滅菌した 0.8%(w/v)NaCl 溶液入 りのシャーレに加え、30 cm 距離の紫外線ランプの真下で照射した。照射開始後の 0、 5、10、15 分に菌懸濁液を 100 µl ずつサンプリングして、Table 1-1 に示す平板培地(固 体)に塗抹し、30℃でコロニーが出現するまで培養した。 (2)長鎖キシロオリゴ糖高生産性変異株の分離 pumilus X-6-19 を紫外線照射することにより、突然変異株 1,241 株を取得した。こ れらの変異株のうち、ハロー形成能の高い菌株を 51 株得ることが出来た。得られた菌 株を液体培養した結果、キシロオリゴ糖の蓄積量が親株に比べて、2 倍に増加した変 異株を 3 株得ることが出来た。 第 4 節 N-methyl-N-nitro-N-nitrosoguanidine(NTG)によるキシロオリゴ糖高生産性変 異株の取得 【方法】 (1)生存率曲線の作成 本菌を Table 1-1 に示す液体培地 7 mL に 1 白金耳植菌し、30℃で、24 時間振盪培養 した。菌懸濁液を1 mL ずつ 6 本の 1.5 mL マイクロチューブに分注した。それぞれの チューブを遠心分離(20,000xg、10 分、4℃)し、50 mM リン酸ナトリウムカリウム 緩衝液(pH7.0)(buffer A)1 mL で洗浄後、再び遠心分離し、0.5% (w/v) NTG 溶液 0.5 mL を加えてそれぞれ 0、10、30、60、90、120 分間変異処理した。変異処理後、遠心 分離し、沈殿した菌体を直ちに buffer A 1 mL で洗浄した。再び遠心分離した後、1 mL の肉汁培地を加えて、30℃に設定したウォーターバスで 1 時間培養した。培養後、得 られた菌体をbuffer A で 2 回洗浄し、同 buffer で希釈した後、希釈液 100 µl を Table 1-2 に示す肉汁平板培地に塗抹した。コロニーが出現するまで 30℃で培養し、NTG 処理 時間とコロニー数により生存率曲線を作成した。
Table 1-2. Nutrient medium. Meat extract 10.0 g Polypenpton 10.0 g NaCl 5.0 g
H2O 1,000 mL
(2)長鎖キシロオリゴ糖高生産性変異株の分離 30、60、120 分間それぞれ NTG 処理した菌懸濁液 100 µl を、0.25%(w/v) D-グル コース、 0.25% (w/v) 酢酸ナトリウム三水和物を含む 0.25% (w/v) キシラン平板固 体培地に塗抹し、コロニーが出現するまで30℃で培養した。グルコース存在下でもキ シランに対して高い分解能力を示すハロー形成能の高い菌株を選択した後、Table 1-1 に示す液体培地7 mL に 1 白金耳植菌し、30℃で振盪培養(210 rpm)した。培養開始 6、12、18、24 時間後それぞれサンプリングし、遠心分離(20,000xg、10 分、4℃)後、 上清中に含まれる還元糖の定量と組成分析を行った。 【結果】 (1)生存率曲線の作成 Fig. 1-6 に示すように、本菌は NTG 処理時間が 120 分の時、生存率が約 10%になる ことが分かった。NTG は強力な突然変異誘起性を持つため、高い生存率でも高効率的 に変異体を得ることが出来るとされているので8)、以後の実験における NTG 処理時間 は 30、60、120 分間の 3 通りに決定した。
Fig.1-6. Survival curve of B. pumilus X-6-19 irradiated with NTG. Symbols: ●, survival rate (%).
1
10
100
0
30
60
90
120
150
180
Time(min)
Surv
iv
al
rate
(%
)
1
10
100
0
30
60
90
120
150
180
Time(min)
Surv
iv
al
rate
(%
)
(2)キシロオリゴ糖高生産性変異株の分離 B. pumilus X-6-19 の NTG 処理により、突然変異株 2,929 株を取得した。これらの変 異株のうち、ハロー形成能の高い菌株を31 株得ることが出来た。得られた菌株 31 株 を液体培養し、培養液中に蓄積する還元糖と組成分析を行った結果、還元糖の蓄積量 が親株に比べて、1.3∼1.7 倍上昇した変異株を 7 株得ることが出来た。 第 5 節 キシロオリゴ糖高生産性株の選択 【方法】 第3 節と第 4 節で得られた長鎖キシロオリゴ糖高生産性変異株 10 株を、1% (w/v) 酢 酸ナトリウム三水和物および D-グルコースを含む 1% (w/v) キシラン培地 7 mL で液 体培養し、培養開始12 時間ごとに 0.50 mL ずつサンプリングした。遠心分離(20,000xg、 10 分、4℃)後、上清中の還元糖の定量と TLC を用いた組成分析を行った。試験した 10 株の中から、培養液中に長鎖キシロオリゴ糖を最も多く蓄積する菌株を選択した。 【結果】 試験した 10 株の中で培養液中に長鎖キシロオリゴ糖を最も多く蓄積する菌株を 1 株選択し、U-3 株とした。本変異株は紫外線照射により得られた変異株であった。、培 養開始 36 時間後、培養液中のグルコースをほぼ完全に利用し、長鎖キシロオリゴ糖を 蓄積することがわかった。この時の培養液中の還元糖量(31 mg/mL)は、親株と比較 して 3.2 倍に増加した。培養液中のキシロオリゴ糖の組成は親株の培養液中の組成と ほぼ同じであった。以上の結果から、本変異株を以下の実験に用いた。 第 6 節 要約と考察 【要約】 B. pumilus X-6-19 株は、培養液中にキシロトリオース(X3)以上の長鎖オリゴ糖を 含むオリゴ糖を蓄積することを見いだした。B. pumilus X-6-19 が、長鎖キシロオリゴ 糖をより効率的に生産するための培養条件の検討を行った結果、1% (w/v) 酢酸ナトリ ウム三水和物を添加することにより、コントロールと比較して 1.5 倍量の長鎖キシロ オリゴ糖を生産することがわかった。さらに長鎖キシロオリゴ糖の生産量を上げるた め、紫外線照射および、NTG 処理によりキシロオリゴ糖高生産性変異株の取得を試み た。その結果、得られた変異株 4,170 株中、長鎖キシロオリゴ糖を親株の 3.2 倍量生 産するキシロオリゴ糖高生産性変異株 U-3 株を取得することが出来た。
【考察】 B. pumilus X-6-19 のキシロオリゴ糖蓄積に与える種々の炭素源の影響を調べた際、 D-グルコースやラクトース一水和物を添加した時、キシロオリゴ糖の生産が抑制され た。これはグルコース効果により、キシラン分解酵素系の発現が抑制されたためであ ると考えられる。しかし、変異株 U-3 株は親株と比較して培養開始後早い時期にグル コースを完全に利用しており、グルコース存在下でもキシラン分解酵素が発現するよ うな変異が遺伝子内部で生じたものと思われる。 また、クエン酸三ナトリウム二水和物やコハク酸二ナトリウム六水和物を添加した 培地で B. pumilus X-6-19 を培養した場合、著しく菌の生育が良かったが、培養液中に 蓄積するキシロオリゴ糖量は増加しなかった。また種々の窒素源を添加した場合にお いても、菌の生育は良好であったが、培地中に蓄積するキシロオリゴ糖量は同様に増 加しなかった。これらの結果から、菌の生育が良好しすぎると、一度生産されたキシ ロオリゴ糖が本菌の炭素源及びエネルギー源として利用されるため、培養液中にキシ ロオリゴ糖が蓄積しないと考察した。
第 2 章 B. pumilus X-6-19 strain U-3 株の生産するキシラナーゼの精製と特性解析 第 1 節 序 前章において、親株である B. pumilus X-6-19 と比較して、3.2 倍量の長鎖キシロオ リゴ糖を生産・蓄積する変異株U-3 株を取得した。目的とするキシロオリゴ糖の生産 効率をさらに向上させるためには、一度生産されたキシロオリゴ糖がさらに、本生産 菌の炭素源およびエネルギー源として利用されないことが重要である。 一方、キシランからキシロオリゴ糖の生成に関与すると予想されるキシラン分解酵 素系を得て、それらの特性を明らかにした後、キシロオリゴ糖製造に利用できれば、 この問題に対する一つの解決策になると考えられる。 そこで本章では、エンド型キシラン分解酵素であるキシラナーゼを精製し、その特 性を明らかにすることを目的とした。また、次章では、エクソ型キシラン分解酵素で あるβ-キシロシダーゼの精製と特性解析について述べる。 第 2 節 酵素活性測定法 【方法】 (1)キシラナーゼ活性測定法 1.0% (w/v)キシランを含む 50 mM リン酸ナトリウムカリウム緩衝液(pH 7.0)(buffer A)0.5 mL に酵素溶液 0.25 mL を添加し、30℃において反応させた。反応時間は 1 分 および 11 分とした。反応終了後、直ちに沸騰水浴中で 10 分間煮沸して酵素反応を停 止させた。酵素反応液を遠心分離(20,00×g、10 分、4℃)し、得られた上清 0.2 mL に、第 1 章、第 2 節に示す Somogyi 試薬 0.2 mL を添加して、沸騰水浴中で 10 分間加 熱した。加熱後、氷水中で3 分間冷却し、Nelson 試薬 0.2 mL を添加して十分に撹拌し た。次に、水2 mL を加えて撹拌し、30 分間放置した後、500 nm における吸光度を測 定した。11 分間反応における吸光度と 1 分間反応における吸光度の差から、生成した 還元糖を算出した。 本酵素 1 unit は、1 分間に 1 µmol のD-キシロースに相当する還元糖を生成する酵素 量と定義した。また、比活性はタンパク質1 mg 当たりの unit とした。 (2) タンパク質の定量法 タンパク質は Lowry 法 16)により定量した。標準タンパク質としてウシ血清アルブ
第 3 節 酵素の生産条件の検討
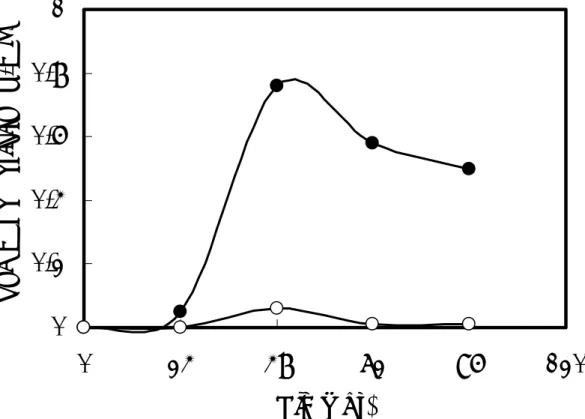
B. pumilus X-6-19 strain U-3 株を Table 1-1 に示す液体培地 7 mL を用いて 30℃、210 rpm の条件下で 24 時間振盪培養した。この培養液 14 mL(試験管 2 本分)を、同培地 500 mL を含む 5 L 三角フラスコに加え、30℃で振盪培養(105 rpm)した。 培養開始 24、48、72、96、120 時間後に培養液をそれぞれ 50 mL 採取した。同培養 液を遠心分離(20,000xg、10 分、4℃)し、培養上清と菌体に分離した。湿菌体 1 g あ たり10 mL の 20 mM Tris-HCl 緩衝液(pH 8.0)を加え、超音波破砕機(INSONATOR 201M、 KUBOTA、Tokyo)により菌体を破砕した(180W、5 分、2 回)。遠心分離(20,000xg、 10 分、4℃)した後、得られた上清を細胞抽出液とした。培養上清中と細胞抽出液中 のキシラナーゼ活性を測定した。 【結果】 培養上清中のキシラナーゼ活性は、培養開始48 時間後において、最大値を示したが、 細胞抽出液中のキシラナーゼ活性は、培養開始48 時間後にわずかに見出されのみであ った(Fig. 2-1)。以上の結果から、培養開始 48 時間後の培養上清中にキシラナーゼが 最も蓄積されることがわかった。
0
0.2
0.4
0.6
0.8
1
0
24
48
72
96
120
Time(hr)
X
yl
an
ase acti
vity (
U
/m
L
)
Fig. 2-1. Production of xylanase at 30℃ by strain U-3.
第 4 節 酵素の精製 【方法】 (1)粗酵素液の調製 本菌の培養は、前節で述べた方法により行った。本培養液を遠心分離(20,000×g、 10 分、4℃)し、得られた上清を酵素精製に用いた(フラクション 1、1,260 mL)。 (2)キシラナーゼの精製 1.硫安分画 フラクション1 に粉末状に磨砕した硫安を 35%飽和になるように攪拌しながら加え た。30 分間攪拌した後、上清を回収した。得られた上清にさらに、硫安を 70%飽和に なるように攪拌しながら加え、30 分間攪拌した後、遠心分離(20,000×g、10 分、4℃) により沈殿を回収した。沈殿を20 mM Tris-HCl 緩衝液(pH 8.0)に溶解した後、50 mM リン酸ナトリウムカリウム緩衝液(pH 6.0)(buffer B)1 L に対して 2 回透析を行った。 透析液(40 mL)をフラクション 2 とした。 2.CM52 cellulose カラムクロマトグラフィー
フラクション2 を buffer B で平衡化した CM52 cellulose(Whatman Chemical Separation、 Clifton、USA)を充填したカラム(2.2×10 cm)にアップライした。カラムを buffer B 120 mL で洗浄した後、0∼0.30 M NaCl を含む buffer B 600 mL を用いてリニアグラジエン ト法により、流速 40 mL/h で酵素を溶出した(Fig. 2-2)。各フラクション(5.0 mL/tube) の酵素活性およびタンパク質量を測定した。高キシラナーゼ活性画分を集めて、buffer B で 2 回透析を行った。透析した溶液をフラクション 3(39 mL)とした。 3. CM-Toyopearl 650S カラムクロマトグラフィー
フラクション3 を buffer B で平衡化した CM-Toyopearl 650S(Tohso Corporation、 Tokyo)を充填したカラム(2.2×10 cm)にアップライした。カラムを 120 mL の buffer B で洗浄した後、0∼0.30 M NaCl を含む buffer B 600 mL を用いたリニアグラジエント 法により、流速40 mL/h で酵素を溶出した(Fig. 2-3)。各フラクション(5.0 mL/tube) の酵素活性およびタンパク質量を測定した。native PAGE で単一なバンドを示したフラ クションを集めてフラクション 4(20 mL)とした。 (3) 電気泳動 1.ポリアクリルアミドゲル電気泳動(native PAGE)
Reisfeld らの方法 18)により、7.5% (w/v) ポリアクリルアミドゲル(pH 4.0)および
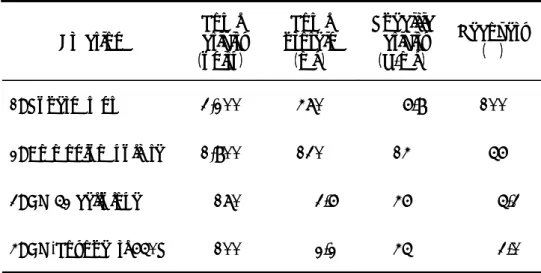
β-アラニン-酢酸(pH 4.5)の泳動用緩衝液を用いて、2.5 mA/tube の条件下で 3 時間泳 動した。タンパク質の染色は、0.25% (w/v) Coomassie Brilliant Blue R-250/エタノール -酢酸-水(9:2:9)染色液を用いて 1 時間行った。エタノール-酢酸-水(25:8:65) 溶液でゲルの脱色を3 時間行った後、エタノール-酢酸-水(10:15:175)溶液中に保 存した。 2.SDS-PAGE Weber および Osborn の方法 19)により、7.5% (w/v) ポリアクリルアミドゲルおよび 0.10% (w/v) SDS-0.10 M リン酸ナトリウム(pH 7.2)の泳動緩衝液を用いて、6.0 mA/tube の条件下で 3 時間泳動した。タンパク質の染色および脱色は、1 で述べた方法により 行った。 【結果】 本酵素の CM52 cellulose カラムクロマトグラフィー、CM-Toyopearl 650S カラムクロ マトグラフィーの結果をそれぞれ、Fig. 2-2、Fig. 2-3 に示す。 各精製段階における酵素活性、タンパク質量、比活性、回収率を Table 2-1 に示す。 キシラナーゼの最終調製酵素標品は培養上清と比較して、比活性45 units/mg となり、 7 倍に精製された。また、その回収率は 3.0%であった。
最終調整酵素のNative PAGE および SDS-PAGE を行ったところ、いずれも単一なタ ンパク質のバンドを示した(Fig. 2-4)。
0.0
2.0
4.0
6.0
8.0
10
30
50
70
90
Fraction numbers(5ml/tube)
Enz
ym
e
acti
vi
ty
(U/m
l)
0.0
1.0
2.0
3.0
Protein
(mg/m
l)
0.3
0.2
0.1
0
NaCl
(M
)
0.0
2.0
4.0
6.0
8.0
10
30
50
70
90
Fraction numbers(5ml/tube)
Enz
ym
e
acti
vi
ty
(U/m
l)
0.0
1.0
2.0
3.0
Protein
(mg/m
l)
0.3
0.2
0.1
0
NaCl
(M
)
Fig. 2-2. Column chromatography of xylanase on CM52 cellulose.
The enzyme solution(Fraction 2, 40 mL)was applied to a column(2.2×10 cm)of CM52 cellulose equilibrated with buffer B. The column was washed with buffer B of 120 mL and then proteins were eluted with a linear gradient of 0 to 0.30 M NaCl in 600 mL of buffer B. Fractions of 5.0 mL were collected at flow rate of 40 mL/h. Protein concentrations and xylanase activities were assayed. Symbols: ■, enzyme activity; ○, protein concentration; and ---, NaCl.
Fig. 2-3. Column chromatography of xylanase on CM-Toyopearl 650S.
The enzyme solution(Fraction 3, 39 mL)was applied to a column(2.2×10 cm)of CM-Toyopearl 650S equilibrated with buffer B. The column was washed with buffer B of 120 mL and then proteins were eluted with a linear gradient of 0 to 0.30 M NaCl in 600 mL of buffer B. Fractions of 5 mL were collected at flow rate of 40 mL/h. Protein concentrations and xylanase activities were assayed. Symbols: ■ , enzyme activity; ○ , protein concentration; and ---, NaCl.
Table 2-1. Summary of xylanase purification.
3.0 45 2.2 100 4: CM-Toyopearl 650S 5.3 46 3.6 170 3: CM 52 cellulose 56 14 130 1,800 2: Ammonium sulfate 100 6.8 470 3,200 1: Supernatant Recovery (%) Specific activity (U/mg) Total protein (mg) Total activity (units) Fraction 3.0 45 2.2 100 4: CM-Toyopearl 650S 5.3 46 3.6 170 3: CM 52 cellulose 56 14 130 1,800 2: Ammonium sulfate 100 6.8 470 3,200 1: Supernatant Recovery (%) Specific activity (U/mg) Total protein (mg) Total activity (units) Fraction
0
1.0
2.0
3.0
4.0
5.0
6.0
0
10
20
30
40
50
Fraction number(5ml/tube)
Enz
yma
tic
ac
ti
vi
ty
(U/
ml
)
0
0.1
0.2
0.3
Pr
otein
(mg/
m
l)
NaCl
(M
)
0
1.0
2.0
3.0
4.0
5.0
6.0
0
10
20
30
40
50
Fraction number(5ml/tube)
Enz
yma
tic
ac
ti
vi
ty
(U/
ml
)
0
0.1
0.2
0.3
Pr
otein
(mg/
m
l)
NaCl
(M
)
Fig. 2-4. Native PAGE(A)and SDS-PAGE(B)of xylanase.
(A)Native PAGE. The purified enzyme(5.0 µg)was run on a 7.5%(w/v)gel of pH 4.0 at 2.5 mA/tube for 3.0 h in a running buffer(pH 4.5)of β-alanine-acetic acid.
(B)SDS-PAGE. The purified enzyme(5.0 µg)denatured with SDS was run on a 7.5% (w/v)gel containing 0.10%(w/v)SDS at 6.0 mA/tube for 3.0 h in a running buffer(pH 7.2) of 0.10%(w/v)SDS-0.10 M sodium phosphate. The gels were stained with 0.25%(w/v) Coomassie Brilliant Blue R-250 in a solvent of ethanol-acetic acid-water (9:2:9).
M, markers:α-lactalbumin (molecular weight 14,400); soybean trypsin inhibitor (20,100); carbonic anhydrase (30,000); ovalbumin (43,000); bovine serum albumin (67,000); phosphorylase b (94,000). 第 5 節 精製酵素の特性 【方法】 (1)精製酵素の分子的性質 (+) M 94,000 (A) (B) (-) 67,000 43,000 30,000 20,100 14,400 26,000 (-) (+)
酵素のnative分子量はゲルろ過法を用いて行った。すなわち、フラクション 4(10 mL)をアミコンウルトラ-15(Nihon Millipore Ltd.、Yonezawa、Japan)を用いて1 mL まで遠心濃縮(4,500xg、4℃)した。濃縮した本酵素液を0.2 M NaClを含む50 mMリン 酸ナトリウムカリウム緩衝液(pH7.0)234 mLで平衡化したToyopearl HW-55 SF(Tosoh Corporation、Tokyo)を充填したカラム(2.0×92 cm)にアップライした。同溶液を用 いて、酵素を流速20 mL/hで溶出した。各フラクション(2.0 mL/tube)の酵素活性およ びタンパク質を測定し、ピークを示すフラクションを決定した。検量線作成に用いた 分子量マーカーとして、calibration proteins gel chromatography kit(Boehringer Mannheim、 Mannheim、Germany)を用いた。
サブユニットの分子量は SDS-PAGE により測定した(本章第 4 節参照)。分子量マ ーカーとして、low molecular weight [LMW] calibration kit(Amarsham Pharmacia Biotech、 Peapack、USA)を使用した。 (2)精製酵素の一般的性質 1.酵素活性に与える温度の影響 20∼80℃に設定した各温度で酵素活性を測定した。各酵素反応において、本酵素を それぞれ 1.3 µg 使用した。 2.熱安定性 本酵素をそれぞれ20∼80℃の各温度で 10 分間処理した後、残存する酵素活性を 30℃ で測定した。各酵素反応において、本酵素をそれぞれ 1.3 µg 使用した。 3.酵素活性に与える pH の影響 1 %(w/v)キシランを含む 100 mM 酢酸-酢酸ナトリウム緩衝液(pH 4.0∼5.5)、100 mM リン酸ナトリウムカリウム緩衝液(pH 5.0∼8.0)、100 mM Tris-HCl 緩衝液(pH 7.0 ∼9.5)および 100 mM 炭酸ナトリウム-炭酸水素ナトリウム緩衝液(pH 9.0∼11.0)を 用いて 30℃で酵素反応を行った。各酵素反応において、本酵素をそれぞれ 1.3 µg 使用 した。 4.pH 安定性 本酵素 1.3 µg を 100 mM 酢酸-酢酸ナトリウム緩衝液(pH 4.0∼6.0)、100 mM リン酸 ナトリウムカリウム緩衝液(pH 5.0∼8.0)、100 mM Tris-HCl 緩衝液(pH 7.0∼9.5)お よび 100 mM 炭酸ナトリウム-炭酸水素ナトリウム緩衝液(pH 9.0∼11.0)で、4℃、24 時間透析した後、残存する酵素活性を30℃で測定した。
(3)NH2-末端アミノ酸配列 Matsudaira20)の 方 法 に よ り 精 製 酵 素 を エ レ ク ト ロ ブ ロ ッ テ ィ ン グ し た 後 、 島 津 PPSQ-10 プロテインシークエンサー(Shimadzu、Kyoto)を用いて、NH2-末端アミノ 酸配列を分析した。 (4)酵素活性に与える各種試薬の影響 キシラナーゼ活性に与える金属イオン、SH 基修飾試薬、キレート試薬および変性 剤の影響を調べた。本酵素(2 µg)溶液 0.5 mL に、終濃度が 1.0 mM になるように各 種化合物を添加し,0.55 mL とした。氷中で 10 分間処理した後、残存する酵素活性を 30℃で測定した。 (5)酵素の動力学的性質 キシランの終濃度が6.7∼33 mg/mL となるように、50 mM リン酸ナトリウムカリウ ム緩衝液(pH 7.0)で調整した溶液に、キシラナーゼの最終調整酵素標品 0.9 µg を加 え、30℃で振盪(140 rpm)しながら酵素反応を行った。反応液中に生産する還元糖を 測定した。Lineweaver-Burk プロット法により Km、Vmax 値を算出した。 【結果】 (1)分子的性質 ゲル濾過法により、本酵素の Native 分子量は 24,000 であった。また、本酵素のサブ ユニットの分子量は26,000 であった。 (2)酵素の一般的性質 1.酵素活性に与える温度の影響 本酵素は、55℃において最も高い活性を示した。また 80℃においては、ほぼ活性を 示さなかった(Fig. 2-6)。
0
20
40
60
80
100
120
0
20
40
60
80
100
Temperature(℃)
R
e
la
ti
ve act
iv
it
y(
%
)
Fig. 2-6. Effects of temperatures on the enzyme activity.
Xylanase was assayed in 50 mM sodium-potassium phosphate buffer(pH 7.0)containing 1% (w/v) xylan at various temperatures.
0
20
40
60
80
100
120
20
40
60
80
100
Temperature(℃)
R
e
s
id
u
a
l a
c
ti
v
ity
(%
)
(2) 熱安定性 本酵素は、45℃以下の熱処理において 95%以上の残存活性を示した。また 60℃以 上の熱処理では完全に活性を失った(Fig. 2-7)。Fig. 2-7. Thermostability of xylanase.
Xylanase was incubated at various temperatures for 10 min, and then residual activities were assayed.
0
20
40
60
80
100
120
3
5
7
9
11
13
pH
R
ela
ti
ve
a
ct
iv
it
y(%)
(3) 酵素活性に与える pH の影響 本酵素はそれぞれpH 6.5 で最も高い活性を示した(Fig. 2-8).Fig. 2-8. Effects of pHs on enzyme activity.
Xylanase was assayed in 100 mM buffer of various pHs containing 1%(w/v)xylan. The buffers used were: ●, acetic acid-sodium acetate; ○, sodium-potassium phosphate; ▲, Tris-HCl; and △, Na2CO3-NaHCO3.
0
20
40
60
80
100
120
3
6
9
12
pH
R
e
la
ti
v
i a
c
tiv
it
y
(%
)
(4) pH 安定性 本酵素は、pH 6.0∼9.0 において 4℃、24 時間処理しても 80%以上の残存活性を維 持した(Fig. 2-9)。Fig. 2-9. Effects of pHs on the stability of xylanase.
Xylanase was incubated in 100 mM buffers of various pHs at 4℃ for 24 h, and then residual activities were assayed. The buffers used were: ●, acetic acid-sodium acetate; ○, sodium-potassium phosphate; ▲, Tris-HCl;, and △, Na2CO3-NaHCO3.
(3)NH2-末端アミノ酸配列 NH2-末端アミノ酸配列を分析した結果、本酵素の NH2-末端アミノ酸配列は RTITNNEMG であった。 (4)酵素活性に与える各種試薬の影響 キシラナーゼ活性に与える各種試薬の影響をTable 2-2 に示す。試験した試薬の中で は、Hg2+、Ag2+が本酵素を著しく阻害した。一方、SH 試薬である PCMB、DTNB、
CH2ICOOH や、キレート剤である EDTA および変性剤である SDS はほとんど本酵素を 阻害しなかった。
Table 2-2. Effect of chemical regents on xylanase activity Compound1 Residual activity(%)
None 100 FeSO4・7H2O 60 FeCl3・6H2O 84 K3Fe(CN)6 48 HgCl2 20 MgSO4・7H2O 84 CuSO4・5H2O 65 AgNO3 29 MnSO4・4-5H2O 78 CoCl2・6H2O 87 NiSO4・6H2O 81 ZnSO4・7H2O 84 CH2ICOOH 94 PCMB2 69 DTNB3 68 α,α’-Dipyridyl 82 N-Ethylmaleimide 88 Tiron 84 EDTA 88 o-Phenanthroline 90 NaN3 92 SDS 86
1The incubation was performed at the concentration of 1 mM compound. 2PCMB, p-chloromercuribenzoic acid.
(5)動力学的性質
本酵素の Km 値と Vmax がそれぞれ 14 mg/mL と 22 µg/min であった。 第 6 節 要約と考察
【要約】
B. pumilus X-6-19 strain U-3 株は主として、キシラナーゼを培養上清中に生成し、細 胞中には蓄積しなかった。本酵素を、硫安分画、CM52 cellulose、CM-Toyopearl 650S を用いたカラムクロマトグラフィーにより、電気泳動的に単一に精製した。本酵素は、 分子量 26,000 のモノマーであった。本酵素の至適 pH は 6.0 であり、55℃において最 大活性を示した。また、pH 6.0∼9.0、45℃まで安定であった。本酵素活性は Hg2+、Ag2+ により著しく阻害されたが、SH 試薬やキレート剤では阻害されなかった。本酵素の Km、Vmax はそれぞれ、14 mg/mL、22 µg/min であった。 【考察】 本酵素は PAGE において、塩基性緩衝液(pH 8.3)を用いた場合にはゲル中を移動 しなかったが、酸性緩衝液(pH 4.5)を用いた場合にはゲル中を陰極側へ移動した。 これは本酵素の等電点がpH 8.3 よりも高いことを示しており、本酵素が塩基性タンパ ク質であると考えられる。 本菌の生産するキシラナーゼは、中性付近において最大活性を示し、分子量は 26,000 であった。既報のキシラナーゼにおいても、分子量 20,000 前後のものが多数あり、本 キ シ ラ ナ ー ゼ に お い て も 既 報 の キ シ ラ ナ ー ゼ と 類 似 し た 分 子 量 の 比 較 的 小 さ な 酵 素 であることが分かった。 Wong ら21)は、真菌、細菌由来キシラナーゼの分子量や等電点など物理化学特性に 基づき、キシラナーゼを分子量 30,000 以下で弱塩基性キシラナーゼと分子量 30,000 以上で弱酸性キシラナーゼに分類した。また、Henrissat22)は、疎水性クラスター分析 によって、キシラナーゼを F/10 および G/11 の二つのファミリーに分類した。それぞ れには、高分子量キシラナーゼと低分子量キシラナーゼが属する。一般に、高分子量 キシラナーゼは、キシランに対する加水分解速度が速く、生産物に低分子キシロオリ ゴ糖が多い。一方、低分子キシラナーゼは、基質に対する加水分解速度は遅いが、加 水分解生産物に長鎖キシロオリゴ糖が多い。B. pumilus X-6-19 U-3 株が生産するキシ ラナーゼは、G/11 の低分子量キシラナーゼファミリーに分類され、長鎖キシロオリゴ 糖を生産しやすいタイプであると考察した。
由来キシラナーゼに比べて耐熱性を持っており、それらの菌株由来キシラナーゼより 高 い 温 度 で 処 理 し て も 活 性 を 維 持 し て い た 。 ま た 、 本 菌 由 来 の キ シ ラ ナ ー ゼ は 、B. pumilus IPO 株由来キシラナーゼより広い pH 範囲で活性を示した24)。したがって、両
菌株の生産するキシラナーゼの9 残基までの NH2-アミノ酸配列が一致したにもかかわ らず、両酵素の内部アミノ酸配列は異なると考察した。
第 3 章 B. pumilus X-6-19 U-3 株の生産する β-キシロシダーゼの精製と特性解析 第 1 節 序
B. pumilus X-6-19 strain U-3 株は、キシランの分解に関与する酵素として、キシラナ ーゼだけでなく、β-キシロシダーゼも生産する。β-キシロシダーゼはキシランやキシ ロオリゴ糖を非還元末端から D-キシロース単位で加水分解する酵素であるから、長鎖 キシロオリゴ糖の生産においては、不都合な酵素といえる。 そこで本章では、β-キシロシダーゼを精製し、酵素特性を明らかにするとともに、 同酵素の生産特性を解析することにより、キシロオリゴ糖の生産・蓄積に与える同酵 素の生合成の影響等を明らかにすることを目的とした。 第2節 酵素活性測定法 (1)β-キシロシダーゼ活性測定法 2 mM p-ニトロフェニル-β-D-キシロピラノシド(Nacalai Tesque,)溶液 0.2 mL に酵素 液 0.2 mL を加え、30℃で 10 分間反応させた。1M 炭酸ナトリウム溶液を 1.6 mL 加え て反応を停止させた後、405 nm における吸光度を測定することによって、遊離した p-ニトロフェノールを定量した。 本酵素 1 unit は、1 分間に 1 µmol の p-ニトロフェノールを生成する酵素量と定義し た。また、比活性はタンパク質 1 mg 当たりの unit とした。 (2)タンパク質の定量法 第 2 章第 2 節に示す方法によった。 第 3 節 酵素の生産条件の検討 【方法】
B. pumilus X-6-19 strain U-3 株を Table 1-1 に示す液体培地 7 mL を用いて 30℃、140 rpm の条件下で 24 時間振盪培養した。この培養液を、同培地 70 mL を含む 500 mL 振 盪フラスコに加え、30℃、105 rpm で振盪培養した。このような培地を含むフラスコ を 8 本用意し、それぞれ、培養開始 0、24、44、50、59、65、72、96 時間後に培養液 を回収した。培養液を遠心分離(20,000xg、10 分、4℃)した後、培養上清と休止菌 体を得た。湿菌体 1 g 当たり 10 mL の 10% (v/v) グリセロールを含む 20 mM リン酸ナ
トリウムカリウム緩衝液(pH7.0)(buffer C)を加えて、菌体を超音波破砕した(180W、 5 分、2 回)。遠心分離(20,000xg、10 分、4℃)した後、得られた上清を細胞抽出液 とした。培養上清と細胞抽出液中の β-キシロシダーゼ活性を測定した。 【結果】 細胞抽出液中の β-キシロシダーゼ活性は、培養開始 65 時間後まで増加し続け,その 以後急激に減少した(Fig.3-1)。培養上清中の β-キシロシダーゼ活性は、細胞抽出液 中の活性に比べて、わずかであった。以上の結果から、β-キシロシダーゼは、培養開 始 65 時間後の菌体中に最も多量に蓄積されることが分かった。本酵素を精製する場合 には、培養開始60 時間後の休止菌体を使用することにした。
Fig. 3-1. Production of β-xylosidase by strain U-3.
Symbols: ● , β-xylosidase activity (cell extract); ○ , β-xylosidase activity (culture supernatant).
0
1
2
3
4
0
24
48
72
96
120
Time(hr)
β
-X
y
lo
s
id
a
s
e
a
c
tiv
ity
(U
/m
L
)
第 4 節 酵素の精製 【方法】 (1)粗酵素液の調製 本菌の培養および細胞抽出液の調製は、前節に述べた方法によって行った。培養液 2 L から得られた休止菌体 18g を使用し、細胞抽出液 185 mL を得た(フラクション 1)。 (2)β-キシロシダーゼの精製 1.除核酸 フラクション1に 20% (w/v) 硫酸ストレプトマイシンを終濃度が 1% (w/v) にな るように加え、30 分間氷水で冷却しながら攪拌した。これを遠心分離(20,000xg、10 分、4℃)し、上清を回収した(フラクション 2、190 mL )。 2.硫安分画 フラクション2 に、粉末状の硫安を 45%飽和になるように氷中で攪拌しながら添加 した。30 分間攪拌した後、遠心分離(20,000xg、10 分、4℃)により上清を回収した。 得られた上清に硫安を80%飽和になるように氷中で攪拌しながら加え、30 分間攪拌し た後、遠心分離(20,000xg、10 分、4℃)により沈殿を回収した。沈殿を buffer C に溶 解した後、buffer C に対して 3 回透析を行った。透析内液をフラクション 3(40 mL) とした。 3.DE52 cellulose カラムクロマトグラフィー
フラクション3 を buffer C で平衡化した DE52 cellulose(Whatman Chemical
Separation)を充填したカラム(1.8×24 cm)にアップライした。同カラムを 300 mL の buffer C で洗浄した後、0∼0.5 M NaCl を含む buffer C 600 mL を用いてリニアグラジエ ント法により、流速50 mL/h で酵素を溶出した(Fig. 3-2)。各フラクション(6.0 mL/tube) の酵素活性およびタンパク質を定量した。高β-キシロシダーゼ活性画分を集めて、 buffer C に対して 2 回透析を行った。透析した溶液をフラクション 4(42 mL)とした。 4.DEAE-Toyopearl 650S カラムクロマトグラフィー
フラクション4 を buffer C で平衡化した DEAE-Toyopearl 650S(Tosoh Corporation) を充填したカラム(2.0×16 cm)にアップライした。同カラムを 250 mL の buffer C で洗 浄した後、0∼0.5 M NaCl を含む buffer C 500 mL を用いてリニアグラジエント法によ り、流速 50 mL/h で酵素を溶出し、各フラクション(3.0 mL/tube)の酵素活性および
タンパク質を定量した。酵素活性を示す2 つのピークが見られ、それぞれピークⅠと ピークⅡとした(Fig. 3-3)。ピークⅠとピークⅡの高活性画分を集めて、終濃度とし て 1.0 M になるように硫安を加え、それぞれフラクション 5-1(18 mL)とフラクショ ン 5-2(12 mL)とした。
5.Phenyl-Toyopearl 650M カラムクロマトグラフィー
硫安 1.0 M を含む buffer C(buffer D)で Phenyl-Toyopearl 650M(Tosoh Corporation) を充填したカラム(1.6×13 cm)を平衡化し、フラクション 5-1 とフラクション 5-2 を 別々にアップライした。同カラムを75 mL の buffer D で洗浄した後、1.0∼0 M 硫安を 含む buffer C 250 mL を用いてリニアグラジエント法により、流速 40 mL/h で酵素を溶 出した。各フラクション(3.0 mL/tube)の酵素活性およびタンパク質を定量し、native PAGE で単一のバンドを示したフラクションを集めてそれぞれ、フラクション 6-1(12 mL)とフラクション 6-2(21 mL)とした。 (3)電気泳動
native PAGE は Davis25)の方法に従って行った。また、SDS-PAGE は第 2 章第 4 節に 示す方法により行った。
【結果】
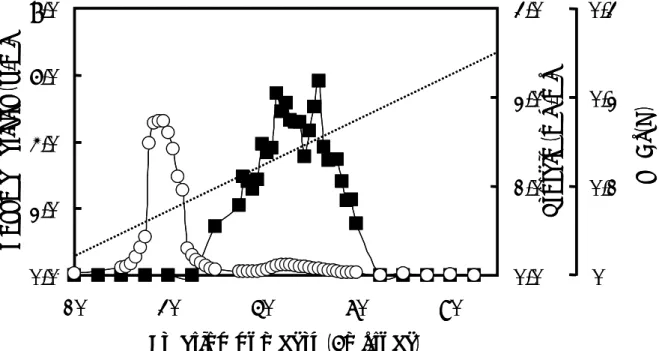
本酵素の DE52 cellulose、DEAE-Toyopearl 650S、Phenyl-Toyopearl 650M カラムクロ マトグラフィーの結果をそれぞれ、Fig.3-2、Fig.3-3、Fig.3-4a、Fig.3-4b に示す。本酵 素は DEAE-Toyopearl 650S カラムクロマトグラフィーにおいて、2 種類のキシロシダ ーゼに分離され、それぞれを β-シロシダーゼ I、II と称することとした。 各精製段階における酵素活性、タンパク質量、比活性、回収率を Table 3-1 に示す。 キシラナーゼの最終調製酵素標品は細胞抽出液と比較して、β-キシロシダーゼ I およ び β-キシロシダーゼ II はそれぞれ、比活性 10 および 8.5 units/mg となり、7.1 および 6.1 倍に精製された。また、その回収率は両者を合わせて 9.8%であった。 β-キシロシダーゼ I および β-キシロシダーゼ II の最終調製酵素は、native PAGE およ び SDS-PAGE においていずれも単一なタンパク質のバンドを示した(Fig. 3-5)。
Fig. 3-2. Column chromatography of β-xylosidase on DE52 cellulose.
The enzyme solution(Fraction 3, 40 mL)was applied to a column(1.8×24 cm)of DE52 cellulose equilibrated with buffer C. The column was washed with buffer C of 300 mL and then proteins were eluted with a linear gradient of 0 to 0.50 M NaCl in 600 mL of buffer C. Fractions of 6.0 mL were collected at flow rate of 50 mL/h. Protein concentrations and β-xylosidase activities were assayed. Symbols: ● , enzyme activity; ○ , protein concentration; and …, NaCl.
0
7
14
1
21
41
61
81
Fraction number(6mL/tube)
Enzy
me act
iv
it
y(
U/mL)
0
0.6
1.2
1.8
2.4
3
Protein(mg/mL)
0.5
0.4
0.3
0.2
0.1
0
Na
Cl
(M
)
0.4
0.1
0 2 4 6 8 1 21 41 61 81 101
Fraction number(3mL/tube)
E
n
zy
m
e
a
c
tiv
ity
(U
/m
L
)
0 0.3 0.6 0.9 1.2 1.5Pr
ot
ei
n(
m
g
/m
L)
0.5 0.3 0.1 0.0Na
Cl
(M
)
0.4 0.2 ピークⅠ ピークⅡFig. 3-3. Column chromatography of β-xylosidase on DEAE-Toyopearl 650S.
The enzyme solution(Fraction 4, 42 mL)was applied to a column(2.0×16 cm)of DEAE Toyopearl 650S equilibrated with buffer C. The column was washed with buffer C of 250 mL and then proteins were eluted with a linear gradient of 0 to 0.50 M NaCl in 500 mL of buffer C. Fractions of 3.0 mL were collected at flow rate of 50 mL/h. Protein concentrations and β-xylosidase activities were assayed. Symbols: ● , enzyme activity; ○ , protein concentration; and …, NaCl.