熊本大学学位論文
転写因子 CooA のガス感知機構および転写活性化機構の解明
2004 山 下 沢
S t u d i e s on t h e Gas S e n s i n g and A c t i v a t i o n Mechanisms o f a T r a n s c r i p t i o n a l F a c t o r CooA
2004
Taku Y a m a s h i t a
第1章 緒論
人間は進化の過程において多くの物事に興味を持つようになり、それを探求する手段 として手を使い道具を用い、いくつもの謎を解き明かしてきた。昔は手段も限られてお り探求可能なことも多くはなかったと思われるが、文明の進歩に伴って手段も増え、そ れにより解明できる事象も増えていき、生命科学の分野において、人間の探求心はつい に生命を一つのものとしてとらえるのではなく、細分化していくつもの個体が集まった 集合体として考え、構成成分であるタンパク質に関する研究へと発展していった。タン パク質は、20 種類のアミノ酸の組み合わせで構成された一次構造が、
α
ヘリックスやβ
シートなどの二次構造を形成し、さらにそれらがフォールディングされて三次構造を 形成したもので、分子量は大小様々である。タンパク質を単離して機能や構造を解明出 来るようになったのはここ数十年程度であるが、その間にも相次いで多くのタンパク質 が発見され、その数は今では優に一万を超えている。生物種にも多種があり生体内の器 官も多様なものが存在するにも関わらず、報告されているタンパク質の形状には構造上 に多くの類似性が存在し、機能が多岐にわたっているにもかかわらず構造上の特徴から いくつかのグループに分類されて言及されることもしばしばである1)。では、なぜ宿主 が異なるタンパク質において類似する構造やアミノ酸配列を持つ現象が起きているのか。そこで、この構造上の共通する特徴をもとに機能を類推する試みがここ数年で現れてき た。その試みにおいては「機能は構造が元になっており、構造はアミノ酸配列が決定す る」という観点から、アミノ酸配列を調べることにより、そのタンパク質の持つ機能が わかると考えたのである。現在においてすべてが該当するとは言えないものの2, 3)、ア ミノ酸配列の近いもの同士、つまり相同性の高いタンパク質間ではその機能も近いと考 えられ、新規タンパク質が発見された際の機能解明へのアプローチの指針となっている
4,5)。これが今現在のタンパク質の研究においてもっとも盛んな分野の一つであるバイ オインフォマティクスである。さらに、アミノ酸配列をもとに検討するという視点から 現在の流れに基づいてこの観点を発展させたものが、ここ最近よく耳にするようになっ たヒトゲノムの解明に関する研究である。昨年にヒトゲノムの全塩基配列が決定され、
今後更なる遺伝子レベルでの研究分野における発展が望まれるが、ヒトゲノムの研究の
意図にはタンパク質の機能解明も含まれており、遺伝子がわかればアミノ酸配列がわか り、それをもとに配列が類似するタンパク質の構造や機能と比較することで、どの遺伝 子配列がどの機能に関与しているかを同定することが出来るというものである。これに より今まで遺伝的要因という名の下に解明されていなかった疾病なども、どの遺伝子が 原因であるかを見極めることができるようになると思われる。しかしながらそれらの正 確な要因決定には、タンパク質の性質もさることながらその量的あるいは質的な面も関 与してくるため、さらに転写レベルでの話が必要になってくるのではあるが、それにつ いてはさらに後で述べることにする。
では、現在のタンパク質研究者達の仕事とは何であろうか。ヒトゲノムが解明された ことで、生体内で発現しているタンパク質はどのようなものが存在するのかは明らかで あり、改めてタンパク質について探求する必要がないのではないかとも考えられる。し かしながら話はそんなに易くはない。例えば、遺伝子から判明するタンパク質のアミノ 酸配列に類似のものが存在しない場合はどうであろうか。遺伝子からわかったアミノ酸 配列に相同性のあるものが無ければその機能や構造すら推測できず、疾病への関与の有 無を問うなど不可能である。また相同性があるからと言って機能が類似しているとは必 ずしも言えず、さらに、実際に遺伝子配列からアミノ酸配列が分かっていても、そのタ ンパク質が生体内においてどの程度発現しているかは生体における様々な要因にもよる ため、疾病の直接的な原因究明には繋がらない。しかしながら、構造や機能が分かって いれば生体内でどのような機能分子が存在しどのような制御機構に関与しているかなど、
ある程度の推測が可能なわけであり、その点においては構造や機能を比較可能なタンパ ク質がより多く存在することが望ましく、多くの研究者が構造決定や機能解明に力を注 いでいる理由の一つでもあり、構造が判明し始めたここ数十年程度で、わずかな期間に 多くのタンパク質の構造が明らかとされてきた背景にもなっている。
以上の流れを総合して考えると以下の結論に至る。タンパク質の構造や機能を研究す る際には、アミノ酸配列から想定される構造や機能をもとにアプローチすることが必然 である。またタンパク質を研究するのであれば、ヒトゲノム解明のこの時代に相応しく、
研究対象は遺伝子レベルでタンパク質の発現を制御している転写因子を扱うことが、遺 伝的な要因とされる疾病に関与する研究ともなる。そこで本研究は転写制御因子を対象
とすることにした。
では転写因子にはどのようなものが存在し、どのような構造的特徴を有しているので あろうか。先にも述べたように、タンパク質の発現は多くの場合転写レベルで制御され ているため、転写因子の制御機構を理解できれば遺伝子の情報に基づくゲノム創薬が可 能になると考えられる。転写因子の研究は RNA ポリメラーゼの発見を追うように 1970 年代から急速に進展し、今現在までに発見されている転写因子の数はゆうに 1000 を越え、その数は現在でも増え続けている。転写因子は大きく分けて二つのカテ ゴリーに分類することが出来る。一つは RNA ポリメラーゼを含む基本転写因子、もう 一つは塩基配列特異的に DNA に結合して転写制御で中心的な役割を果たす転写制御因 子である。転写制御因子には Zn フィンガーやロイシンジッパー、塩基性-ヘリックス・
ループ・ヘリックスなどのいくつかの構造モチーフが存在するが、その中の一つである ヘリックス・ターン・ヘリックス(以下 HTH)構造は約 20 個のアミノ酸からなり、
ターンにはグリシンとその前後に疎水性残基が、また二つのヘリックスの中央部にはい くつかの疎水性残基が存在するという共通の特徴を持つ。二つのヘリックスのうち、一 つのヘリックスは認識ヘリックスと呼ばれ、転写制御因子として重要な DNA との相互 作用に関与し、DNA らせんの主溝に入り、突き出したアミノ酸側鎖を介して特定の DNA 塩基対と相互作用する。原核生物の HTH 構造を持つ転写制御因子はすべて、対 称的な同型二量体を形成することで DNA と結合し、それぞれの認識ヘリックスは対称 的な塩基配列、つまり回文構造と結合する。このことは、1980 年代に発表された CRP (cAMP 受容体タンパク質) と DNA との X 線結晶構造などを代表とした二量体を形成 する転写制御因子と DNA の複合体の立体構造解析により解明されつつあり6, 7)、原核 生物における転写因子で HTH 構造を有するとされるタンパク質と DNA の結合様式を 類推することが可能となった。さらに HTH モチーフに類似するものについて述べるな らば、真核生物においてはショウジョウバエの形態形成に関与するホメオ遺伝子が存在 する8)。HTH に類似する二本のヘリックスにさらに一本ヘリックスが追加された構造 上の特徴を持ち、二量体ではなく単量体で安定な構造をとり、その結果、認識配列も回 文構造とはならない。また oct-19) や pit-1 などが保有する POU ドメイン、さらに HNF-3
γ
10)などに含まれる winged helix も HTH モチーフ類似とされる構造であり、ターンの長さが異なったりヘリックスだけでなく
β
シートも保有するなど、構造上で 多少は異なるものの、DNA への結合には二本のヘリックスとその間を結ぶターンが関 与するという点では共通であり、HTH 構造を持つものとして分類されている。HTH 構造をもつこれらの転写因子だけでなく、構造が明らかとなった転写因子のほとんどが ここ 10 年程に報告されており、その数は今後ますます増えていくと考えられることか ら、より多くの複合体の構造情報を得ることが、来るべき新しい時代にむけて求められ ていると考えられる。転写制御によるタンパク質発現については上記に述べたような転写制御因子が非常に 大きく寄与していると考えられる。しかしながら、生体内においては転写レベルでのタ ンパク質発現制御に加え、外部からの刺激に対応するべく様々なシグナル伝達系を介し た制御機構が存在する。ここでは特にガス分子の及ぼす影響について記述したい。生命 がこの地球上に誕生してから非常に長い年月が流れているが、その生命が誕生する以前 から地球上に存在していたものがガス分子であり、ガス分子を構成する窒素や酸素、炭 素、水素などの原子によって有機体が構成され、生命の誕生が起こったと考えられてい る。言い換えれば、人間はおろか生命が誕生する遙か以前からガス分子は存在している わけであり、生体の構成および機能においてガス分子が大きく関与しているであろうこ とは言うまでもない。ガス分子は非常に分子量が小さく、酸素や窒素などの二原子分子 や二酸化炭素などの三原子分子が存在するものの、構造は極めてシンプルでありほとん ど違いがないと言っても過言ではない。生理活性を示す際にはその単純な構造をどのよ うに生体が認識し活性化へと導かれていくのか、ガス分子と生体との関係については解 明されていない点が多く、研究を行っているグループも少なくない。ガス分子と生体と の関係という点で、生体内のタンパク質との結合で考えれば一番身近なものはヘモグロ ビンとミオグロビンではないだろうか。ヘモグロビンは
αβ
二つのサブユニットがそれ ぞれ二つずつ集まった四量体を形成して存在し、酸素を運搬する役割を担っている11)。 またミオグロビンはヘモグロビンとは異なり、運搬ではなく酸素貯蔵に関与しているタ ンパク質であり、その構造はタンパク質において初期の段階から明らかとされたもので、ヘモグロビンに比べ構造は単純で分子量が小さく安定性も高いことから、今日でも変異 導入や合成ポルフィリン錯体の導入など様々な研究分野で利用されることが多い12-15)。
このようにガス分子と結合するものにはヘムタンパク質が多く、酸素以外のガス分子と 結合するものとして、一酸化窒素の運搬を行うニトロフォリン16)なども存在する。さ らに、ガス分子を感知することで様々な機能を発現するものが存在し、近年発見された ものについて言及すれば、一酸化窒素と結合することで GTP を cGMP へと変化させる 可溶性グアニル酸シクラーゼ (sGC)17, 18)や、酸素分子の濃度を感知し窒素固定遺伝子 発現を制御しているヒスチジンキナーゼ (FixL)19, 20)、酸素分子を認識するホスホジエス テラーゼ (Dos および AxPDEA1)21-23)、さらには酸素の走化性制御系におけるトランス デューサー(HemAT) などがあり、これらのシグナル伝達機構については、ガス分子の 認識機構とそれに引き続いて起こる酵素反応の活性化機構が研究対象として非常に注目 されている。
ヘムタンパク質は生体内において重要な役割を果たしており、上記に示したガス分子 に関与するもの以外にも、電子伝達系に関与するシトクロム類、酵素の働きを示すカタ ラーゼやペルオキシダーゼなども存在し、その機能は多岐に渡っている。また近年では 病原性細菌類が生体内で生きていく際に必要とする鉄分の供給源もヘムタンパク質であ ることが指摘されており、必要不可欠な存在であると言える。これらヘムタンパク質の 活性中心には、いずれも鉄-プロトポルフィリン錯体、すなわちヘムが補欠分子として 含まれており、鉄原子の酸化還元により電子の受け渡しで活性を示す場合には、特にヘ ムを取り囲むタンパク質部分の構造およびアミノ酸残基の性質により種々の活性が発現 あるいは制御されている。一般に補欠分子を含まないタンパク質についてはアミノ酸変 異による検討によって活性を調べることが主な研究となりがちであるが、ヘムタンパク 質を用いることによって、補欠分子との相互作用などの要因が絡んではくるものの、分 光学的手法を用いることで他のヘムを含まないタンパク質の存在を無視可能なほど特異 的に検出できるという利点がある。さらに、活性中心であるヘムに着目することが容易 な測定環境が整っていれば、活性に関与する変化のみを特異的に検出できることから、
ヘムタンパク質を研究対象とすることで、天然に存在するタンパク質部分によるヘムの 活性制御機構の解明と、得られた結果に基づくタンパク質工学的手法によるモデルタン パク質合成の試みなど、種々の機能を発現するタンパク質分子の自由な設計、合成、機 能変換が可能である。
以上の点を総合し、先に述べた転写制御因子としての重要性とそれを踏まえた上での 転写活性化機構の解明、また生体にとって必要不可欠であるガス分子の認識、さらには ヘムタンパク質としての分子設計の発展性などに着目し、一酸化炭素により転写活性を 制御するヘムタンパク質である CooA を研究対象として選択した。CooA は、元来紅 色非硫黄光合成細菌
Rhodospirillum rubrum
中に含まれており、この菌が光合成細菌 にもかかわらず光がない環境においても一酸化炭素存在下では生育可能であることから、一酸化炭素を感知しエネルギーへと変換するタンパク質が存在することが示唆され、そ の制御を担っているのが CooA であることが 1990 年頃に明らかとなった26-31)。今日 では CooA は一酸化炭素を感知することによって活性化され、一酸化炭素の酸化など に関与するタンパク質の転写制御因子として働くことが明らかとなっており、この機能 が光合成細菌が光のない環境でも生き抜くための手段の一つとして持つようになったも のであると考えられている。では一酸化炭素を感知し転写を制御可能な構造とはいかな るものであろうか。また転写活性化機構はどのような仕組みであろうか。
CooA の構造的な特徴を述べる前に他のタンパク質の例を見ていただきたい。ガス分 子を感知し転写を制御するヘムタンパク質は非常に希少であり、その研究価値は非常に 高いと考えられる。今現在までに明らかとされているものでは、近年哺乳類中で発見さ れ生体時計の機能に関係するタンパク質とアミノ酸配列の相同性が高い NPAS232)が存 在する。NPAS2 は CooA と同様に一酸化炭素を感知して転写を活性化するが、こちら は異型二量体を形成することからその活性化機構は CooA とは異なるものであると推 測される。また酸素を感知して転写を制御しているものとして HIF1
α
33-35)があり、酸 素濃度の高低に応じて血球増殖や血管新生などの適応反応を転写調節により行っている とされるが、今日では酸素濃度を感知しているのは共存する別の酵素によることがわかっ ており、正確にはガス感知転写因子とは言えない。いずれのタンパク質も構造は未だわ かっておらず、またアミノ酸配列の特徴も異なることから CooA と比較検討すること は出来ない。発見当初 CooA は、ヘムタンパク質ではないものの cAMP によって DNA 結合性が制御されている CRP 7, 36-39)と、アミノ酸配列において相同性が非常に高 く DNA 結合性も確認されたことから27, 40-43)、CRP と同様にエフェクター結合ドメイ ンと HTH 構造を有す DNA 結合ドメインおよび二つのドメインをつなぐ一本の長いヘリックスによって構成されていると推測され、精製過程や質量分析器による測定からや はり CRP と同じく同型二量体で存在していることが明らかとなった (Figure 1A) 44, 45)。 以降、紫外可視吸収スペクトルなどの各種測定から、一つのプロトヘムを含み、酸化型、
還元型、一酸化炭素結合型のいずれにおいてもヘム鉄は6配位低スピン状態にあること、
部位特異的変異を用いた研究から、酸化型では 75 位のシステインと 2 位のプロリンが、
還元型では 77 位のヒスチジンと2位のプロリンが配位し、6配位構造をとっているヘ ム鉄の二つの軸配位子のうち、一方が鉄の酸化還元に伴い置換されていることが明らか
となった46-49)。なお、このプロリンの還元型における軸配位は部位特異的変異では解
明できず、2000 年に発表された還元型の X 線結晶構造解析によって判明したもので50)、 今日では酸化型でもこのプロリンの配位は変わらないとされている。またこの還元型の 立体構造から、やはり CRP と同様に二つのドメインがヘリックスによってつながった 構造 (Figure 1B) を持ち同型二量体として存在すること、部位特異的変異による結果か らわかった軸配位子が正しいことが明らかとなった。さらに CooA の軸配位構造が二 つのサブユニット間で軸配位を供給しあうという極めて特殊な構造をとっていることも 明らかとなった。この明らかとされた構造は、一酸化炭素が結合した形、つまり一酸化 炭素結合型ではないため、実際の転写活性型の構造は未だ明らかとはされていない。し かしながら、構造が非常に類似しているとされる CRP については転写活性型と DNA との複合体の立体構造が明らかとされており、CooA の構造を比較することによってあ る程度の推測は可能である。
よって著者は CooA を用いることによって、ガス分子という小分子がいかにして転写 活性化という複雑な構造変化を伴う機構を制御しているのか、またガス分子の生体内に おける役割において、運搬や貯蔵を行うものと転写を制御するなどのシグナルを感知す るものの間に何らかの違いは見出せるのか。さらには、未だ構造が明らかとされていな い転写活性型である一酸化炭素結合型の構造に関する何らかの知見が得られることを期 待し、本研究に着手した。以下、各章節にわけて段階を追って詳述する。
Figure 1. Structural differences between cAMP-bound CRP (left) and ferrous CooA (right). (A)
α -helices and β -sheets are colored in red and cyan, respectively. (B) Effectors, effector
domains, C-helices, and DNA-binding domains are colored in red, orange, green, and
blue, respectively.
第2章 CooA 遺伝子の全合成および発現系の構築
第1節 序論
タンパク質を研究するには、目的タンパク質をそのままタンパク質として、あるいは 発現可能である菌株を入手する必要がある。本研究は分光学的な手法を中心として進め られるが、常に必要量のタンパク質を任意に獲得するためには菌株に目的タンパク質を 発現させる必要がある。そこで獲得方法として二つの道を考えた。一つは本来の光合成 細菌株を手に入れて単離抽出する方法、もう一方は CooA を遺伝子として獲得し大腸 菌株に組み込み単離精製する方法である。本研究では遺伝子クローニング操作を習得し かつ変異体の調製などを容易にするなど将来的な観点に立ち後者を選択した。外来性の タンパク質を大腸菌などの宿主に組み込んで大量発現させる方法は、近年の遺伝子工学 においては、外来性のタンパク質のみならずアミノ酸配列を変化させた変異体タンパク 質に対しても行われるのが一般的であり、過去においてもヘムタンパク質の分野だけで も多くの例が存在し、1980 年代の半ばまでに、ミオグロビン51)、ヘモグロビン52)、シ トクロム c ペルオキシダーゼ53) さらにはシトクロム c 54) と次々と大腸菌での発現系が 確立され、その後、さらに複雑な構造を持つシトクロム P450 55)や西洋ワサビペルオキ シダーゼ56)、シトクロム酸化酵素57)などまでが可能となってきた。しかしながら遺伝 子組み換えにより異種生物に存在するタンパク質を大腸菌で大量発現させる場合には、
両者でコドンの使用頻度が異なる点が問題となってくる。しかし逆に考えれば、使用頻 度を大腸菌に合わせることによって異種由来タンパク質の遺伝子も大腸菌での発現に最 適化することができることになり、より多くの目的タンパク質の発現を望める。以上の 点を検討し、CooA 遺伝子を本来の光合成細菌由来の塩基配列によらず、発現するタン パク質のアミノ酸配列をもとに大腸菌での発現に最適化したものとして設計することに した。
第2節 CooA 遺伝子の獲得
まず、より大量に発現させるために、アミノ酸配列を変えないようにしながら大腸菌 で使用頻度の高いコドン58)に置き換えることで大腸菌における大量発現系の確立を目
指した。またコドンを変えることで
cooA
遺伝子にユニークな制限酵素の認識部位を 20 ヶ所組み込んだ。これらの制限酵素の部位は、遺伝子の合成及びその後の遺伝子組 み換えを行う際に用いた。コドンを置き換えることによって大腸菌に最適化されたcooA
遺伝子とし、PCR を用いた特殊な方法 (Scheme 1) 59)を利用することで相補的なcooA
遺伝子を得ることが可能であると考え、さらに合成後の塩基配列の確認などの点 から、全長 666 塩基対を前後半の二つの部分に分け、それぞれを図示したような部分 相補的な六つのオリゴ DNA として設計し、個別に合成することにした。なお、図に示 した相補的な部分の結合を示す線は模式的なものであり、水素結合や塩基対の数を示す ものではない。合成した遺伝子はクローニングベクターに組み込んで繋げ、目的とする 完全長のcooA
遺伝子を得ることに成功した。得られた遺伝子は DNA シークエンサー で確認し、完全にcooA
遺伝子であり設計通りに完成したことが明らかとなった。第3節 CooA 発現系の構築
遺伝子の全合成という従来の考えに捕われない新奇な方法によって
cooA
遺伝子が獲 得できたことが確認された。そこで、次にこの遺伝子から実際にタンパク質を発現させ、他のグループが報告した天然型や野生型と一致するか否かを確認することとした。発現 には本研究室で pBluescriptII SK(+) と pET 3a をもとに作製された pBEX ベクターを 用い、ヘム合成の基質となる 5-アミノレブリン酸溶液を加えることでヘムの産生とタ ンパク質の発現を同調させ、菌体に可溶性のヘムタンパク質の存在を示す赤色を確認し た。超音波破砕器によって菌膜破砕を行い、超遠心分離を行って不可溶性分画を取り除 き、イオン交換と分子ふるいカラムとを併用することで赤色を示すヘムタンパク質の単 離精製に成功した。なおこれら一連の発現精製過程の詳細については実験の部に記す。
得 られた ヘ ム タ ン パ ク 質 が CooA で あ る こ と を 確 認 す る た め 、 SDS-PAGE と TOF-MS により分子量を確認し、アミノ酸配列をアミノ酸シークエンサーにより同定 した。さらに吸収スペクトルを測定し、酸化型、還元型、一酸化炭素結合型のすべてに おいて、その極大吸収波長は天然型および野生型 CooA と一致した。以上のことから 得られたヘムタンパク質が間違いなく野生型 CooA であることが確認できた。
Scheme 1. Synthesis of designed CooA gene with recursive PCR method.
第4節 小括
本研究を行う際に、目的タンパク質である CooA を獲得するのは非常に重要である。
当研究室には本研究を行った当初から部位特異的変異を行う環境は確立されており、本 方法はその延長線上にあるとも言える。幾つかのプライマーを部分相補的な領域を持た せて設計し、PCR を行うことで相補的でない部分に対して伸長反応が行われ、完全な 相補的 DNA となるわけであるが、基本原理は PCR の伸長反応であり、プライマーの 設計と反応条件を検討することによって、今まであまり類の見ないような方法で目的タ ンパク質の遺伝子を得ることに成功した。本方法は、身近に発現可能な遺伝子あるいは 菌株が存在しなくても目的タンパク質を得ることが可能となる方法であり、かつ大腸菌 あるいは他の宿主菌による培養を行う場合には、それに最適化した状況で遺伝子を設計 し発現までをスムーズに行える利点がある。なお本方法はすでに完全に確立されており、
当研究室だけに限らず他の研究室でも目的のタンパク質を得るためにこの方法を用い、
細菌由来だけでなくヒトなどの哺乳類由来のタンパク質についても獲得に成功している。
また今回の野生型 CooA に言及するならば、その発現量は他の宿主由来のタンパク質 をそのままの塩基配列で発現させたものに比べて収量は非常に増加したことから、今後、
生体でごく微量にしか発現していないようなタンパク質について研究する際に本方法が 非常に有効であり、大量に発現させ精製が比較的簡単に行えるであろうことは疑う余地 も無い。
第3章 DNA 結合ドメインの及ぼす影響
第1節 序論
CooA は、先に述べたように一酸化炭素を感知するヘムタンパク質であり、転写非活 性型である還元型の立体構造が X 線結晶構造解析によってあきらかとなっているが、
転写活性型である一酸化炭素結合型の構造は未だ解明されていない。そのため、転写活 性化機構は推測の域を出ないが、一酸化炭素がヘム結合ドメインのヘム鉄に結合するこ とによって DNA 結合ドメインの構造変化が誘起され、特異的な塩基配列を認識できる ようになると考えられる。この DNA 結合ドメインの構造変化は相同性の関係上 CRP と非常に近いと考えられ、DNA 結合ドメインがヘム結合ドメインに対して接近した構 造をとり得ることが CRP と DNA との複合体の構造を元に推測された29, 60-62)。CooA の転写活性化において、この DNA 結合ドメインの動きが非常に重要であると考えられ、
一酸化炭素の結合による構造変化が如何なるものかが他のグループによっても推測され ているが未解明のままである63, 64)。本研究では CooA の DNA 結合活性の制御に着目 し、DNA 結合ドメインの及ぼす影響を検討するために、Figure 2 に示すような DNA 結合ドメインを欠失させた変異体、以下 truncated 変異体を調製し、各種分光学的測 定を行うことで、DNA 結合ドメインとヘム結合ドメインとの相互作用の有無、一酸化 炭素が結合する際の DNA 結合ドメインが及ぼすヘム結合ドメインへの影響を調べ、野 生型と各種分光学的測定結果の比較検討を行った。
第2節 truncated 変異体の調製
前章で示した合成によって得られた cooA の遺伝子は大腸菌内での発現に最適化され たものであり、野生型においてその発現量は充分であった。そこで、この遺伝子を用い て以後のすべての変異体を調製することとした。DNA 結合ドメインは先に述べたよう に一本の長いヘリックスを介してヘム結合ドメインと繋がっている。このヘリックスの C 末端には 131 位のメチオニンが存在しており、この部分がヘリックスとドメインと の間に存在する。そこで cooA 遺伝子上で、本来 131 位のメチオニンのコドンの ATG に続いて存在する TTC を、終止コドンである TAA に変えて翻訳を終了させることに
した。この変異の導入には部位特異的変異に用いる方法を適用し、設計したプライマー を用い PCR によって終止コドンを導入することとした。
DNA シークエンサーで塩基配列を確認した後にタンパク質を発現させた。基本的に 多くの部分を欠失させた場合、全体の構造が本来の構造を保持できない場合もある。今 回の場合のようにドメインごとに別れた部分を欠失させた場合では、その可能性は多少 は低くなるもののゼロではない。タンパク質が正しくフォールドされて発現することを 期待し、組み換え遺伝子を含む大腸菌を培養したところ、可溶性分画にヘムタンパク質 に特有の赤色を確認できたので、可溶性分画をイオン交換カラムにていったん精製した 後、ゲルろ過カラムにて再度精製を行った。この際、溶出時間に野生型とは明確な差が 生じたため、分子量が変化していることが明らかとなり、得られたものを SDS-PAGE によって分子量を確認したところ、発現した CooA において確かに DNA 結合ドメイン が欠失していることが確認できた。
Figure 2. Structure of wild-type CooA (left) and a putative structure of truncated CooA (right).
DNA-binding domain in blue (left) is deleted in the truncated structure (right).
第3節 truncated 変異体の吸収スペクトルによる構造変化の追跡
DNA 結合ドメインを欠失させたことによってどのような影響が考えられるであろう か。構造的な変化がどの程度誘起されるのかを検討するために各種分光学的測定を行う こととした。測定には現有する装置を主に用い、野生型と truncated 変異体とを比較 し、DNA 結合ドメインの存在の有無による違いを検討した。
まず紫外・可視吸収スペクトルを測定した。吸収スペクトルの波形には可視領域であ る 400nm 付近にヘムタンパク質に特有の吸収帯である Soret 帯が見られ、極大吸収波 長の数値や波形からヘムタンパク質に含まれるヘム鉄の配位などに関する知見が得られ る。第2章で確認のため測定した野生型の吸収スペクトルからは、酸化型、還元型およ び一酸化炭素結合型のいずれにおいても6配位低スピン状態のヘム鉄を含むことが示唆 された (Figure 3)。
Figure 3. UV-Vis absorption spectra of wild-type CooA. Ferric, ferrous, and CO-bound forms are
shown as thick black, thin black, and thick glay lines, respectively. Q-band regions are
expanded 5-fold.
そこで次に truncated 変異体について吸収スペクトルを測定して、野生型のものと 比較してみた (Figure 4)。精製後の標品は野生型の時と同様、酸化型のスペクトルを示 し、その波形および極大吸収波長の値は野生型のものとほとんど変化がなく、酸化型に おいてヘム周辺構造には DNA 結合ドメインの欠失による影響は見られない。次に還元 型についてであるが、こちらも酸化型と同様に波形や極大吸収波長のいずれも野生型と ほとんど差が見られず、やはり DNA 結合ドメインの欠失による影響は見られない。最 後に一酸化炭素結合型であるが、CooA の機能的な意味を考えると DNA 結合ドメイン と一酸化炭素の結合には何らかの因果関係が存在しうる可能性が非常に高い。そこで当 然のことながら吸収スペクトルにおいても何らかの違いが見られると期待した。しかし ながらその波形や極大吸収波長には酸化型、還元型と同様にほとんど変化がなく、
DNA 結合ドメインが直接的に一酸化炭素の結合に関与していないように見えた。
Figure 4. UV-Vis absorption spectra of truncated CooA. Ferric, ferrous, and CO-bound forms are
shown as thick black, thin black, and thick gray lines, respectively. Q-band regions are
expanded 5-fold.
このことはヘム鉄の配位数などを変えるほどの大きな変化が生じていないことを示唆 しているとも言え、DNA 結合ドメインが欠失していても全体的な構造にはあまり影響 を及ぼしていない、つまり野生型と比較してタンパク質の巻き違いなどが起きていない ことを示しているとも言える。吸収スペクトルの結果からは以上のように DNA 結合ド メインがヘム周辺構造に全くと言っていいほど影響を及ぼしていないことが示された。
しかしながら吸収スペクトルにおいてはヘム周辺構造の微細な変化を追跡することは困 難であり、ヘム鉄の配位子など詳細を知ることは難しい。そこでヘム周辺構造の変化を より詳細に調べるために、共鳴ラマンスペクトルを測定し、野生型と truncated 変異 体とのヘム周辺環境の違いを調べた。
第4節 共鳴ラマン分光法にみる DNA 結合ドメイン欠失の及ぼす構造変化
ラマン分光法は、赤外分光法と同じく振動分光法の一種であり、分子の振動エネルギー を測定することによって分子構造を探る手段である。一般に、分子の電子吸収帯に一致 する励起光を照射することによって、ラマン散乱光を十万倍以上に増強することが出来、
その電子遷移に関与している発色団の構造情報のみが得られる。これが共鳴効果である。
タンパク質のような複雑な構造においても、ヘムタンパク質のように吸収スペクトルに おいてヘムなどの発色団由来の特有な吸収帯を有していれば、共鳴効果を得るためにそ の波長に近い光で励起することで発色団自身および周辺の構造の情報のみを主に得るこ とが出来る極めて優れた分光法である。また共鳴効果を用いることにより、測定試料が 微量であっても測定が可能であるなど、ヘムタンパク質の研究を行う際には非常に有用 な方法である。測定結果に関してはミオグロビンによる同定をはじめ65)、様々なヘム タンパク質の測定により、どの振動ピークがどの結合部分に起因するものであるかも容 易に判断できる。以上の理由から、共鳴ラマンスペクトルを測定することによって、ヘ ム周辺環境の微細な変化、特に軸配位子に及ぼす影響を詳細に検討した。なお、測定方 法に関する詳細な情報は実験の部に記載した。
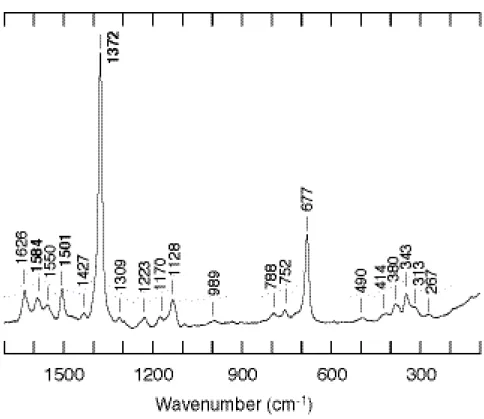
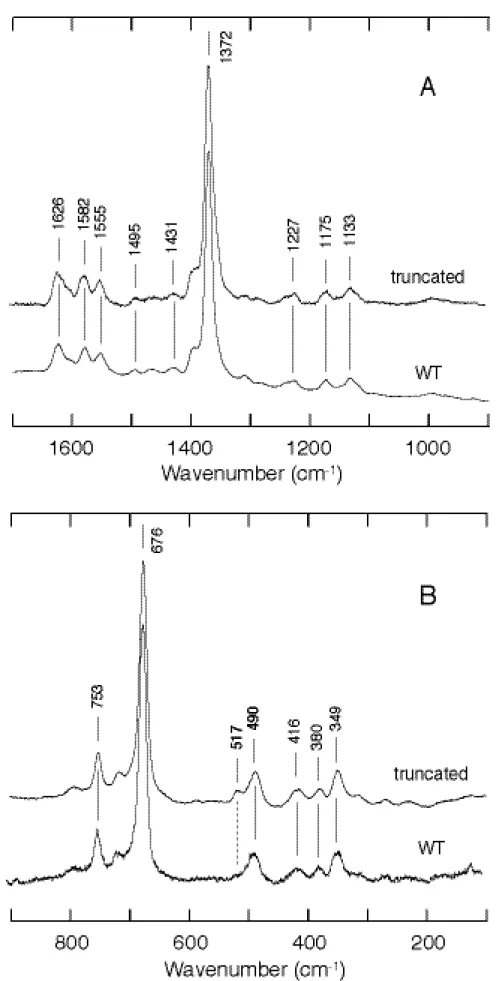
まず、野生型について精製標品の測定を行った。吸収スペクトルの結果から CooA は 酸化型、還元型および一酸化炭素結合型のいずれにおいても6配位低スピン状態をとる と考えられる。共鳴ラマンスペクトルを測定したところ、スピン状態と配位状態に敏感
な高波数領域において、酸化還元状態のマーカーバンドである
ν
4 が 1372 cm-1に見ら れたことから、精製後の標品がやはり酸化型をとっていることがわかる (Figure 5)。ま たν
3 およびν
2 がそれぞれ 1501 cm-1と 1584 cm-1に出現したことから、ヘム鉄が6 配位低スピン状態であることも確認できた。また、6配位のヘム鉄を含む他のヘムタン パク質について明らかとなっている波数と比較したところ、軸配位子について、特にν
2 を比較した結果、一方にはシステインがもう一方にはイミダゾール基類似のものが 配位していると考えられた (Table 1)。このことは先に述べた他のグループによる部位 特異的変異の結果とも一致した。Table 1. Resonance Raman frequencies of CooA and other heme proteins.
Abbreviations : Cyt b2, Cytochrome c oxidoreductase ; RLM, rat liver microsomal
Figure 5. Resonance Raman spectrum of wild-type CooA in the ferric form.
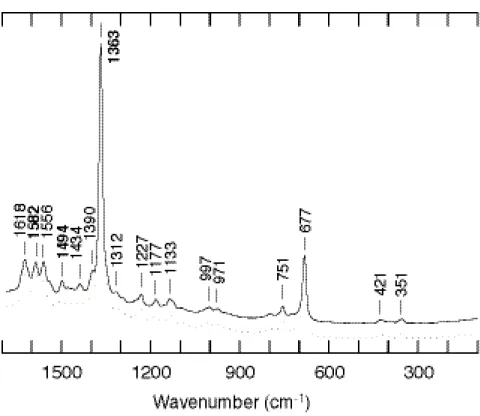
次に還元型について述べる。すでに X 線結晶構造解析により、構造が明らかとなっ ていることから、ヘム鉄は6配位で低スピン状態にあると考えられる。共鳴ラマンスペ クトルから、ヘム鉄が還元状態にあることを示すバンドである
ν
4 が 1363 cm-1に見ら れた (Figure 6)。さらにν
3 およびν
2 がそれぞれ 1494 cm-1と 1582 cm-1に出現した ことから、結晶構造解析の結果と同様に溶液中でも6配位低スピン状態であることが明 らかとなった。また逆にスペクトルから他のヘムタンパク質と比較し構造を検討してみ ても、やはりヒスチジン由来の軸配位子の存在を示唆することとなり (Table 1)、得ら れたスペクトルが確かに野生型 CooA のものであることがわかる。また酸化型におい てはヘム鉄に 75 位のシステインが配位しており、それがヘム鉄が還元されることによっ て 77 位のヒスチジンに置換されることが提唱されてきたが、共鳴ラマンの結果もそれ を支持するものとなった。Figure 6. Resonance Raman spectrum of wild-type CooA in the ferrous form.
最後に一酸化炭素結合型について述べる。還元型に一酸化炭素が結合すると、二つ存 在する軸配位子のうちの一方が一酸化炭素によって置換され、6配位低スピン状態をと ることが吸収スペクトルから推測される。このヘム鉄と一酸化炭素との結合については 500 cm-1付近に伸縮振動ピークが見られることがわかっており、その波数から一酸化 炭素がどの程度の強さで、またどのような形で配位しているのか、さらにはヘム鉄に配 位し続けている近位にあたる軸配位子が何であるかや周辺にどのような残基が存在する かまで推測可能となる。野生型 CooA においては還元型で 77 位のヒスチジンとアミノ 末端に位置する 2 位のプロリンが配位しており、二つの軸配位子は共に窒素原子が配 位した形状をとり、比較的性質も似ていると考えれる。現にプロリンの配位は当初はヒ スチジンの配位であると言われており、他のグループの研究においても未だ一酸化炭素 結合型における近位軸配位子は推測の域を出ない。本研究においても、ヘム鉄と一酸化 炭素の結合の伸縮振動を示すバンドが 490 cm-1に検出されたが、この値は他の研究グ ループと一致し、またそれ以上のことを同定することは不可能であった (Figure 7)。
Figure 7. Resonance Raman spectrum of wild-type CooA in the CO-bound form.
ただ、実際に測定することによって以下のことが判明した。当研究室では、今までに ミオグロビンなどを扱い共鳴ラマンスペクトルを測定してきた。今回の野生型 CooA については、ミオグロビンと同様の条件で励起させようとした場合、通常の励起光出力 では配位している一酸化炭素を光解離させてしまうことが明らかとなったため、出力を 1 mW に下げて測定を行わなければならなかった。この現象は後述する一酸化炭素の解 離定数を求める実験により、CooA の CO に対する親和性が低いためであると解明され た。以上が野生型について得られたものであり、その波数や波形は他のグループの報告 とも一致した。
そこで今度は truncated 変異体についても同様に測定を行った。図は高波数側と低 波数側に分けて野生型と比較して示した。この変異体については吸収スペクトルのみが 他のグループによって報告があるものの61)、その他の測定および解析はなされておら ず、如何なる結果が出るか非常に興味が持たれた。
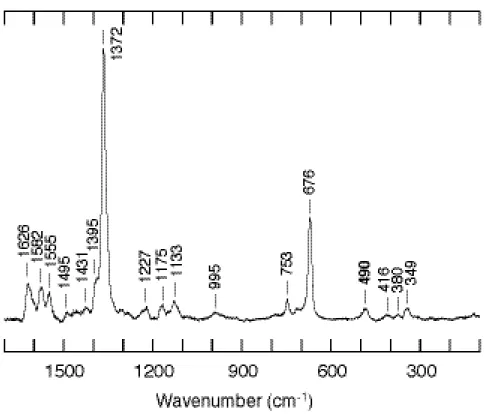
精製後のサンプルを測定すると、酸化還元マーカーバンドである
ν
4 が 1373 cm-1に 検出 されたことから、野生型と同様に精製後は酸化型をとっていることがわかる (Figure 8)。またν
3 およびν
2 が 1501 cm-1と 1584 cm-1に見られ、野生型の酸化型と 一致したことから、酸化型においては DNA 結合ドメインの影響はほとんどないことが わかった。このことは CooA が還元型であるときに一酸化炭素が結合して転写活性型 になることを考えれば妥当であり、むしろ変化がないということから、転写に関係ない 状態においてはその活性化を制御しているヘム周辺構造は DNA 結合ドメインの動きに 左右されず安定性が保持されていると言える。次に還元型を測定したところ、野生型と同様に還元状態を表すマーカーバンドである
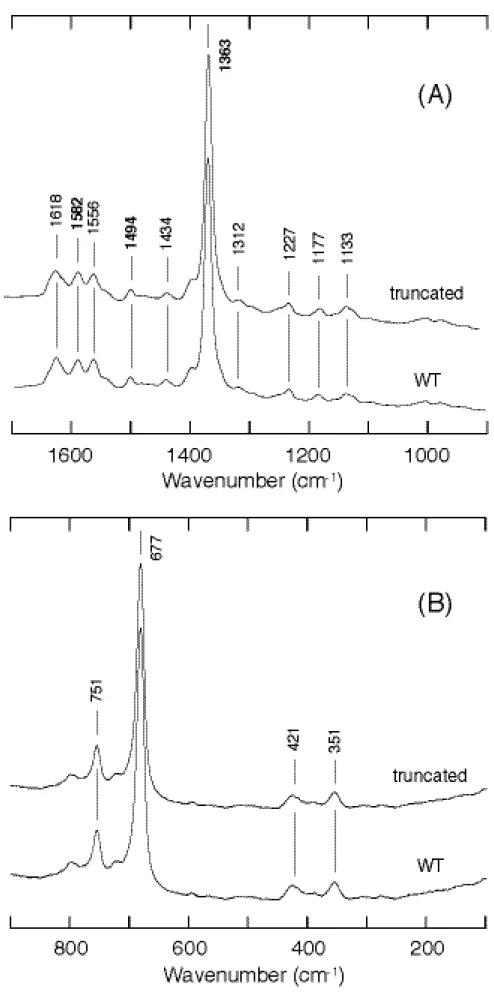
ν
4 が 1362 cm-1に、またν
3 およびν
2 が 1493 cm-1と 1582 cm-1に見られた (Figure 9)。この値も野生型の還元型とほぼ一致した。つまり、還元型においても野生型と同様 にヘム鉄は 6 配位低スピン状態にあり、DNA 結合ドメインの欠失による軸配位子の変 化は起こっていないと考えられる。このことは、ヘム鉄の還元によって酸化型から還元 型へと移行した CooA は、ヘム鉄の還元という現象だけによっては、DNA 結合ドメイ ンの構造変化が誘起されず、またヘム周辺構造は酸化型の時と同様、非常に安定に保た れていることがわかる。Figure 8. Resonance Raman spectra of ferric CooA in high frequency (A) and low frequency (B)
regions.
Figure 9. Resonance Raman spectra of ferrous CooA in high frequency (A) and low frequency (B)
regions.
では、一酸化炭素結合型ではどうであろうか。CooA の還元型の立体構造から考える と、ヘム結合ドメインと DNA 結合ドメインは離れていることから、やはり変化がない のではと考えられる。しかし機能という面から考えると、DNA 結合ドメインが欠失し たことはヘム鉄への一酸化炭素の結合というシグナルを伝達する相手を失ったこととな り、結合に関しても何らか影響があってしかるべきだとも思われる。吸収スペクトルで はほとんど変化が見られなかったことから、共鳴ラマンスペクトルにおいて何らかの変 化が見られ、シグナルが伝達されていることが証明できることを期待した。
一酸化炭素結合型にして truncated 変異体の共鳴ラマンスペクトルを測定したとこ ろ、吸収スペクトルのようには一致せず、野生型とは異なる結果を示した。一酸化炭素 が配位することに由来する鉄と炭素原子の伸縮振動を示すラマンバンドが、野生型と同 様に 489 cm-1に見られるが、その高波数側に、同じく一酸化炭素配位由来と考えられ るバンドが 518 cm-1に出現した (Figure 10)。これは野生型には存在しないものであり、
DNA 結合ドメインが欠失したことによると思われる。このバンドはヘム鉄と一酸化炭 素の変角振動ほどは高い値を示さなかったことから、やはりヘム鉄と一酸化炭素の伸縮 振動であると帰属した。伸縮振動が二本検出された点については主に二つ考えられるこ とがある。一つはヘム鉄一つに対して一酸化炭素が二つ配位した形をとる場合、もう一 つは一酸化炭素の結合様式に二種類が存在し、異なる二つのバンドとして検出される場 合である。共鳴ラマンの結果からだけではそれがどちらにあたるのかを明言するのは難 しい。そこで一つのヘムあたりにどれだけの一酸化炭素が反応するのか、またその反応 性に野生型との違いがあるかを調べることにした。
Figure 10. Comparison of resonance Raman spectra of CO-bound CooA in high frequency (A)
and low frequency (B) regions.
第5節 一酸化炭素の結合性の変化
反応に関与する外来性の配位子を検討する際、ヘムタンパク質の研究でよく行われる 方法はシアン滴定である72)。これは酸化型のヘム鉄に対してシアンを配位させ、その 変化を追跡するもので、シアンを徐々に滴下し、ヘムタンパク質の吸収スペクトルの変 化を解析して外来性の配位子が配位可能なヘムポケットの数や性質を調べるものである。
本研究では還元型に対する一酸化炭素の結合を調べるために、この方法を応用すること が適当ではないかと考えた。実験に関する詳細はここでは省略するが、一酸化炭素の飽 和溶液を調製し、気密セルに徐々に滴下していき、吸収スペクトルの変化を追跡する。
シアン滴定と同様に吸収スペクトルの変化を追跡し解析を行うことで反応に関与する一 酸化炭素の数や親和性を同定した。解析方法に関する詳細は実験の部に記載した。
まず野生型について測定を行った。野生型は共鳴ラマンスペクトルを測定する際に、
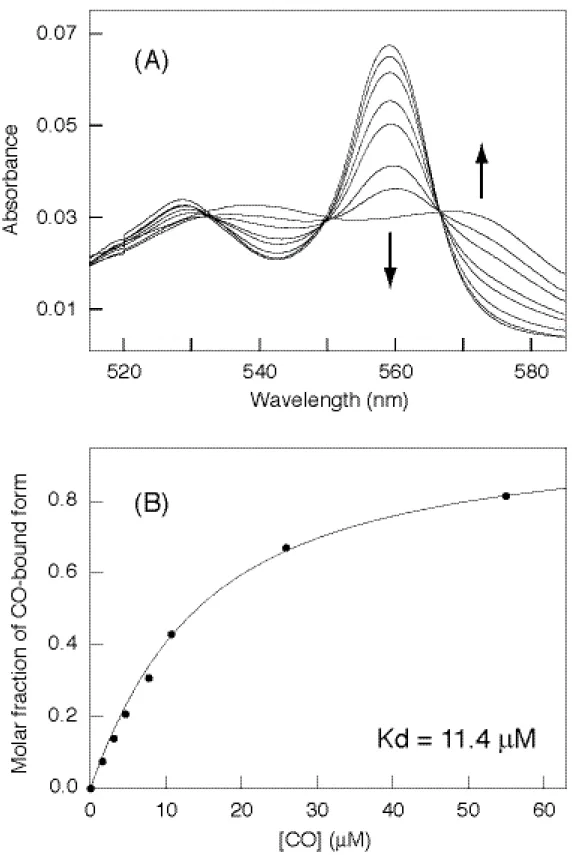
ラマンの励起光であるレーザーの照射によって一酸化炭素が非常に光解離しやすかった ことから、ヘム鉄への結合親和性は非常に低いものであることが推測された。実際に滴 定していった際の吸収スペクトルの変化を示す (Figure 11A)。スペクトルは等吸収点 をとって変化しており、ヘム鉄と一酸化炭素との結合が一段階であることがわかる。こ のスペクトル変化において特に吸光度変化の大きな波長において変化の割合を算出し、
加えた一酸化炭素の濃度変化に対して変化率をプロットすると、図に示したような点と して描くことが出来、理論曲線をフィットさせることで一酸化炭素の解離定数である Kd が算出できた (Figure 11B)。野生型 CooA における Kd 値は 11.4
µ
M となり、ミ オグロビンに比べると約 500 倍も結合しにくいことがわかった (Table 2)。この値は他 のセンサータンパク質のものと近く、容易にはガス分子が結合しないということが、セ ンサータンパク質に共通の性質であると推測できる。また理論曲線は反応に関与する一 酸化炭素の数が一つであることを示しており、一つのヘム鉄つまり単量体一つに対して 一酸化炭素が一つ結合していることを示している。Figure 11. CO titration analysis of wild-type CooA. Spectral changes in CO titration (A) and plots
for molar fraction of CO-bound form against CO concentrations (B).
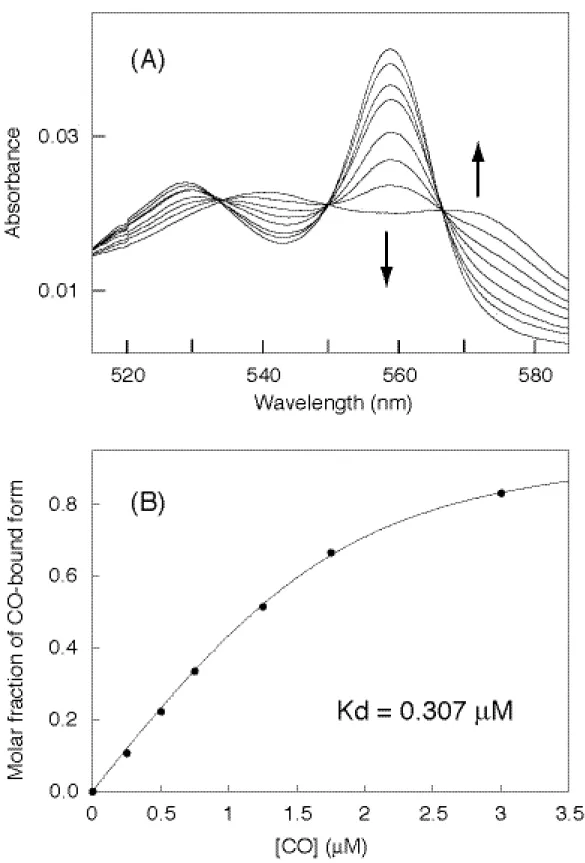
次に truncated 変異体についても野生型と同様に測定を行った。共鳴ラマンの測定 の際には、野生型ほどの容易な光解離は見られなかったことから、その結合親和性は野 生型よりも高いことが推測された。実際に滴定した結果、そのスペクトル変化は野生型 の時と同様に等吸収点をとったことから反応は一段階であることがわかる (Figure 12A)。また変化率を一酸化炭素濃度変化に対してプロットした結果から、一酸化炭素 のヘム鉄への配位数は野生型と同様に単量体一つに対して一酸化炭素が一つであり、解 離定数が 0.31
µ
M となることが算出された (Figure 12B and Table 2)。このことは野 生型と比べると、反応に関係する一酸化炭素の数は同じであり、DNA 結合ドメインが 結合親和性に非常に影響を及ぼしていることがわかる。この親和性の違いは、先の共鳴 ラマンの測定の際の光解離の結果とも矛盾しない。また truncated 変異体の共鳴ラマ ンスペクトルで帰属されたヘム鉄と一酸化炭素との二本の伸縮振動ピークは、一酸化炭 素の結合が二分子を意味するのではなく、二つの異なる結合様式が存在することがこの 研究より明らかとなった。Table 2. Dissociation constants (Kd) of CO in various heme proteins.
Figure 11. CO titration analysis of truncated CooA. Spectral changes in CO titration (A) and plots
for molar fraction of CO-bound form against CO concentrations (B).
第6節 小括
DNA 結合ドメインの役割として、当然のことながら DNA への結合があるもののそ の構造的な意義を、ドメインを欠失させることによって明らかとすることが出来た。こ れは、精製過程でのヘムの脱離や精製時のイオン交換カラムでの分離の際に大きな変化 が現れなかったことから、構造が完全に変わってしまうようなタンパク質の巻き直しが 起こっていないことが分かる。その上で、酸化型および還元型においては吸収スペクト ル、共鳴ラマンスペクトルのいずれにおいても野生型と truncated 変異体の間に有意 な差が見られなかったことから、活性型である一酸化炭素結合型以外では DNA 結合ド メインがヘム結合ドメインと直接的な関係にはないことを示している。しかしながら一 酸化炭素結合型においては共鳴ラマンスペクトルにおいてヘム鉄と一酸化炭素の結合に 顕著な差が見られた。また一酸化炭素の親和性を調べた実験においては、約 40 倍もの 差が認められ、DNA 結合ドメインの有無による違いが一酸化炭素の結合性の違いとし て示された。ヘム鉄へ一酸化炭素が結合し、そのシグナルが DNA 結合ドメインの構造 変化を誘起するであろう CooA の転写活性化機構を考慮し、単純に軸配位子の一酸化 炭素による置換という形で制御されていると考えられていたが、今回の実験によって一 酸化炭素の結合もまた DNA 結合ドメインの存在によって制御されていることが明らか となり、本研究が、DNA 結合ドメインとヘム結合ドメインという二つの離れた位置に 存在するドメイン間に相互作用が存在することを証明した初めてのケースとなった。な お、本実験を遂行するために確立した一酸化炭素滴定法は、ガス分子を定量的に測定す ることが可能である極めて希少な方法であり、今後も様々な領域において有用であると 思われる。
第4章 遠位軸配位子の機能解明
第1節 序論
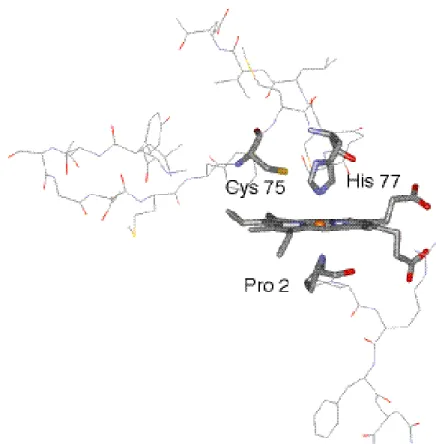
前章で DNA 結合ドメインの一酸化炭素結合性への影響が検討された。その結果、転 写活性化機構は単純な軸配位子の置換ではない可能性が示唆された。では還元型で存在 する二つの軸配位子の役割は如何なるものであろうか。先に触れたように CooA にお けるヘム鉄の配位構造は非常に複雑であり、酸化型における近位軸配位子であるとされ る 75 位のシステインは、ヘム鉄の還元に伴い 77 位のヒスチジンに置換される。さら に遠位側は 2 位のプロリンが同型二量体間で相補的に軸配位子として存在している (Figure 13)。
Figure 13. Structures of heme and proximate residues in the ferrous CooA (pdb :1ft9). Pro 2 belongs to the other subunit.
今まで、一酸化炭素結合型に光照射して一酸化炭素の解離型を非常に短い単位で時間 分解測定し、ヒスチジンが配位していることが提唱されてきたが79, 80)、プロリンも一 酸化炭素結合型で配位している可能性があり、未だ確定していない。そこで一酸化炭素 結合型には Scheme 2 に示す二つの形態をとる可能性があると考え、本研究では、ま
ず一酸化炭素結合型における軸配位子が何であるかを、truncated 変異体の時と同様、
吸収スペクトル、共鳴ラマンスペクトル、および一酸化炭素滴定によるスペクトル変化 の三種類の分光学的測定を行うことによって解析し、同定することとした。またそれに より性質の類似したプロリンとヒスチジンという二つの軸配位子の役割がそれぞれどの ようなものであるかを検討した。
Scheme 2. Coordination structure of the heme in the ferric, ferrous, and CO-bound CooA.
第2節 遠位軸配位子の吸収スペクトルによる解析
構造が明らかとなっているのは非活性型の還元型である以上、著者を始め研究の目は 当然のように転写活性型である一酸化炭素結合型に向けられる。一酸化炭素結合型にお ける軸配位子については、他のグループが行った光照射の経時変化から、77 位のヒス チジンであることが示唆されている。そこで、まず遠位である 2 位のプロリンに変異 を導入し、各種測定を行い、プロリンの置換による影響を検討した。なおプロリンは先 に述べたようにアミノ末端に位置している。他の研究グループにおいて、2 位のプロリ ンをチロシンとヒスチジンに置換した変異体が報告されており49, 81)、他の測定結果と 比較するために 2 位のプロリンをヒスチジンに置換した変異体、以下 P2H 変異体、を 部位特異的変異により調製し、まず吸収スペクトルを測定した。
精製後の標品は野生型の時と同様にやはり酸化型のスペクトルを示したが、その波形 は極めて異なるものとなった (Figure 14)。
Figure 14. UV-Vis absorption spectra of P2H CooA. Ferric, ferrous, and CO-bound forms are
shown as thick black, thin black, and thick gray lines, respectively. Q-band regions are
expanded 5-fold.
野生型と同じ 420 nm 付近に Soret 帯が見られるもののその強度は弱くなり、代わっ て 390 nm 付近にショルダーとして別のピークが出現しているのがわかる。またこの ピークと連動して 650 nm 付近にも新たにピークが出現していることがわかる。この 390 nm 付近に出現した幅広の吸収は、シトクロム P450 にカンファーが結合したもの およびそのモデルタンパク質と類似しており82-84)、P450 と同様システインが配位した 5配位高スピン状態をとっていることがわかる。高スピンでは 650 nm 付近に吸収帯 が現れることがわかっており、この点からも P2H 変異体にはシステインが配位した5 配位高スピン成分が存在することが確認された。しかしながら完全にすべてが5配位成 分ではなく、野生型と同じ6配位も混ざった状態で存在していることがわかる。これは 置換したヒスチジンのアミノ末端あるいはイミダゾール窒素が配位した状態をとりうる ことを示している。
次に還元型であるが、酸化型の場合とは非常に異なった結果を示し、野生型と似通っ た波形を示した。これは酸化型および還元型の両方においてプロリンは配位しているも のの、ヘム鉄の酸化状態によって遠位軸配位子の親和性に違いが生じていることを示し ており、アミノ末端の配位は還元状態に対しては比較的安定に保持されると考えられる。
では、一酸化炭素結合型においてはどうであろうか。吸収スペクトルは還元型と同様、
野生型とほとんど違いが見られない。このことは、一酸化炭素結合型における軸配位子 が 77 位のヒスチジンであり、本来一酸化炭素が配位することによって置換される残基 を変異導入によって置換したところで、顕著な変化が現れないのは当然であると考える ことも出来る。しかしながら、先に述べたように近位のヒスチジンと遠位のプロリンの アミノ末端の性質は類似しており、P2H 変異体において、導入されたヒスチジンの側 鎖あるいはアミノ末端が保持されている可能性も完全には否定できない。そこで、より 詳細に検討するために、truncated 変異体の時と同様、共鳴ラマンスペクトルを測定 した。
第3節 遠位軸配位子の共鳴ラマンスペクトルによる解析
精製過程で CooA は野生型や truncated 変異体において酸化型で存在しており、酸 化型の軸配位子である 2 位のプロリンが置換された P2H 変異体は安定性が低下し、発 現量も少なく精製が困難であったため、共鳴ラマンを測定するのは難しく、そのスペク トルには多少のノイズが認められた。しかしながら、酸化状態を示すバンドである
ν
4 が 1373 cm-1に検出されたことから確かに酸化型をとっていることが判明し、吸収ス ペクトルにおける5配位高スピン成分由来のものと思われるバンドがν
3 とν
2 で 1493 cm-1と 1578 cm-1に検出でき、吸収スペクトルの結果と一致した (Figure 15A)。なお、低スピンよりも高スピンの方がラマンバンドの強度としては強く現れる傾向にあり、
P2H 変異体において6配位成分がわずかにショルダーとして見られているものの、そ の存在比は単純にバンドの強度比では言えないことをここでは明記しておきたい。また ノイズで分かりにくいものの、低波数領域にも多少の変化が見られる (Figure 15B)。
これはスピン状態が変化しており、ヘム鉄がポルフィリン平面から多少なりとも引っ張 り出され、ポルフィリン骨格に歪みが生じた結果であると考えられた。
次に還元型について共鳴ラマンスペクトルを測定した。吸収スペクトルの測定結果か ら野生型との間には違いがないと思われたが、遠位ではあるものの軸配位子を置換して おり、何らかの変化が見られることを期待した。しかしながら、共鳴ラマンスペクトル の波形および各バンドの波数は P2H 変異体と野生型との間でほぼ一致し、遠位軸配位 子の置換による還元型の配位構造への影響は見られない。この結果は確かに吸収スペク トルの結果を支持するものではあるものの、2 位のプロリンの存在意義が何たるかを解 明する上で、未解決な問題として残った。そこで一酸化炭素結合型においては共鳴ラマ ンスペクトルに何らかの違いが見られると考えた。
Figure 15. Resonance Raman spectra of P2H CooA in the ferric form in high frequency (A) and
low frequency (B) regions.
前章で触れたように、ヘム鉄と一酸化炭素の結合については伸縮振動が 500 cm-1付 近に検出される。P2H 変異体について測定を行ったところ、ヘム鉄と一酸化炭素の伸 縮振動は 490 cm-1に検出され、野生型とほとんど変化が見られなかった (Figure 16)。
この振動の波数はすなわち結合のエネルギーを現しており、近くに妨げとなる残基が存 在すればその値も当然変化する。またより結合しやすくなればその結果も反映される。
これは先に truncated 変異体の際に見られた変化を考えれば理解しやすい。この DNA 結合ドメインを欠失させた変異体では、外来性配位子が結合可能な部位であるヘムポケッ トに直接的には影響を及ぼしていないはずの DNA 結合ドメインを欠失させただけで、
一酸化炭素結合型の共鳴ラマンスペクトルには大きな変化が現れたのである。しかしな がら、本来軸配位しているアミノ酸を置換した P2H 変異体においてほとんど変化が見 られなかったということはどういうことであろうか。2 位のプロリンは、やはり単なる 一酸化炭素に置き変わるまでの代理としての役割だけなのであろうか。共鳴ラマンスペ クトルの結果からはヘム鉄への配位構造においてはほとんど変化が見られず、これ以上 の知見を得ることは出来なかった。では、一体このプロリンの役割は何なのであろうか。
Figure 16. Resonance Raman spectrum of P2H CooA in the CO-bound form in low frequency region.
第4節 遠位軸配位子の置換による一酸化炭素結合性の変化
共鳴ラマンを測定した際、P2H 変異体では truncated 変異体の時と同様、野生型と 異なり光解離しにくい現象が見られた。つまり一酸化炭素の結合親和性が高くなってい る可能性が示唆された。そこで著者は一つの点に注目した。それはセンサータンパク質 と呼ばれるものに共通のある値である。今現在までにセンサータンパク質について、そ の幾つかは一酸化炭素との結合親和性が測定解析なされており、その値はいずれも近い ものである。一方、ミオグロビンやヘモグロビンなどのグロビンファミリーは 100〜
1000 倍程度の高い親和性を示す (Table 2)。著者はこの低い親和性こそがセンサータン パク質が機能する上で重要であると考え、CooA においてこの親和性を制御する役割を 担っているのが 2 位のプロリンではないかと考えた。そこで P2H 変異体について一酸 化炭素の結合親和性を調べるために truncated 変異体と同様に一酸化炭素滴定を行い、
そのスペクトル変化を追跡した。測定の方法は truncated 変異体の際と同様である。
吸収スペクトル変化が等吸収点を持つことから、反応は一段階であることがわかる (Figure 17A)。吸光度変化から一酸化炭素結合型の割合を算出し、一酸化炭素濃度に対 してプロットしたところ、反応に関与する一酸化炭素の数は一つとなり野生型と一致し たものの、解離定数 Kd は大きく異なり、野生型では 11.4
µ
M であったのに対し、P2H 変異体においては 0.053
µ
M となった (Figure 17B)。Figure 11. CO titration analysis of P2H CooA. Spectral changes in CO titration (A) and plots for molar fraction of CO-bound form against CO concentrations (B).
第5節 小括
野生型と比較すると P2H 変異体では約 200 倍も一酸化炭素の結合親和性が高くなっ ており、この値はむしろグロビンファミリーのそれと近い。吸収スペクトル、共鳴ラマ
ンスペクトルの結果と照らし合わせると、野生型と結合様式に変化はなく、ヘム鉄一つ あたり一分子の一酸化炭素が結合し、その結合親和性のみ大きく影響されていることか ら、野生型における一酸化炭素の結合は Scheme 3 のようであると考えられる。よっ て、2位のプロリンは一酸化炭素の結合性を制御する役割を担っており、一酸化炭素が 容易には結合できず、生体内で一定の濃度に達したときにのみ機能発現が起きる。
CooA においてはそれが転写活性化であり、ガス分子がタンパク質の機能を制御する上 で、構造変化の引き金となるガス分子結合段階で親和性を調節することによって複雑な メカニズムをうまく制御しているのだと思われる。今回、解明した 2 位のプロリンの 役割は、未だ他のグループによっては解明されておらず、本研究によってセンサータン パク質に共通の性質として結合親和性を掲げることによって、今後さらに発見されるで あろうガス制御転写因子をはじめとするセンサータンパク質において、その指針となる であろうことは間違いない。