株式会社林原生物化学研究所・研究センター・基礎細 胞研究部門
特集疾患の制御―臨床から免疫へ―
総 説新しいタイプのヒト制御性 T 細胞(HOZOT)の発見と臨床応用
中 村 修 治Discovery of a new type of human regulatory T cell lines, HOZOTs, and their clinical application
Shuji NAKAMURACell Biology Institute, Research Center, Hayashibara Biochemical Laboratories, Inc. (Received May 25, 2009)
summary
Recently, we have discovered a new type of regulatory T (Treg) cell, designated HOZOT, by co-culturing human umbilical cord blood cells with mouse stromal cell lines. There are three unique characteristics of HOZOT ; the ˆrst one is the induction method using the unfractionated cell population, the second is a phenotype of CD4+CD8+, the third is
a multifunctional property of killer, suppressor, and helper activities. HOZOT can be deˆned as a new type of Treg cells because of its immunosuppressive activity in allogeneic MLR and of its phenotype and functions distinct from conven-tional Treg cells. HOZOT exerts cytotoxic activities against mouse stromal cells as well as human tumor cells such as colon carcinoma. Moreover, IL10/RANTES/IL8 are deˆned as signature cytokines of HOZOT due to the remarka-bly high production of them. Given the unique properties of HOZOT, especially anti-tumor and immunosuppressive ac-tivities, HOZOT should be utilized for clinical application for the treatment of cancer and immunological disorders. Key words―HOZOT; regulatory T cells; stromal cells; CD4+CD8+; IL10
抄 録 ヒトの臍帯血の細胞をマウスのストローマ細胞と共培養することにより従来知られていなかった種類の制御性 T 細胞(Treg)が誘導されることが明らかになった.HOZOT と命名されたこの T 細胞には 3 つの大きな特徴があ る.1 つは誘導法でヒト臍帯血の単核球細胞をマウスストローマと共培養し xenogeneic な系を用いる点,2 つめは CD4+CD8+のダブルポジティブの表現形を示す点,3 つめはキラー活性,サプレッサー活性,ヘルパー活性をあ わせ持つ多機能性である点である.HOZOT はアロ MLR 抑制機能をもつことから Treg として定義されたが表現 型や免疫抑制作用機序などから従来の Treg と異なっている.またマウスストローマに対してキラー活性を示すだ けでなくヒトの大腸癌細胞などある種の腫瘍細胞に対してもキラー活性が判明した.また,HOZOT はサイトカイ ン産生という面から IL10/RANTES/IL8 の高産生細胞として従来の T 細胞と大きく異なる.これらの特徴のう ち特に抗腫瘍活性とサプレッサー活性を同時にもつことから機能や分化への興味とともに臨床応用が期待される. I. は じ め に か つ て は そ の 存 在 す ら 疑 問 視 さ れ た 抑 制 性 T ( suppressor T ) 細 胞 が 制 御 性 T ( regulatory T : Treg ) 細 胞 と 名 前 を 変 え て 復 活 し た1,2). さ ら に FOXP3 という Treg を規定する遺伝子の発見を機 に Treg の研究と応用は一気に加速された感があ る3).FOXP3 は,かつて CD8+の抑制性 T 細胞を IJ で規定したにもかかわらず遺伝子として同定で き な か っ た の と は 対 照 的 に 明 確 な 遺 伝 子 と し て CD4+CD25+Treg の機能と分化の特異性を規定し ている4).しかし一方で Treg には FOXP3 を発現し ない種類も存在することが報告されてきた5). 我々は造血幹細胞を研究する過程で偶然にも新し い タ イ プ の Treg 細 胞 株 HOZOT を 見 出 し た6). HOZOT はヒト臍帯血の単核球細胞をマウススト ローマ細胞と共培養することで誘導される.この誘 導法は未精製画分を用いること,IL2 など増殖因 子は加えないこと,抗原提示能の弱いと考えられる ストローマ細胞を用いることなどの特徴を持つ.誘 導された HOZOT は CD4+CD8+の double positive
図 1 HOZOT の 3 つの特徴
HOZOT のユニークな 3 つの特徴を図示した.誘導法,表 現型,多機能性のいずれにおいても従来の T 細胞とは異な ったユニークな特徴を持つ.
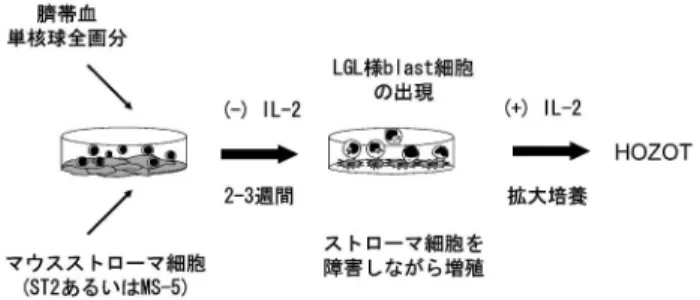
(DP)の表現型を示す点で従来の成熟 T 細胞と異 なっており,さらに FOXP3+,CD25high,GITR+, CTLA4+などの表現型,allogeneic MLR(混合リ ンパ球反応)抑制機能を持つことから新規 Treg 細 胞として分類できるもののキラー(細胞障害)活性 あるいは条件によってはヘルパー(免疫促進)活性 も合わせ持っている(図 1).このような従来の Treg と異なった特徴は HOZOT の機能や分化への 興味を増すとともに臨床への応用が期待される.本 総説では HOZOT の発見の経緯とその特徴また臨 床応用の可能性について述べる. II. HOZOT の発見 HOZOT は臍帯血から誘導される Treg 細胞株で ある6).まずその発見の経緯を説明する7). 1. 臍帯血を使った解析 臍帯つまりへその緒は胎児が誕生するまで母親と 酸素や栄養の交換をしたり老廃物の処理をしたりす るための重要な器官である.臍帯血には成人の末梢 血と同様な血液細胞が流れているが成人の血液細胞 といくつかの点で異なっている.そのひとつは未熟 な段階の細胞を多く含む点であり,特に血液細胞の なかで最も未熟な造血幹細胞を多く含む.そのため 臍帯血は造血幹細胞移植のソースとして臨床応用さ れている.HOZOT は臍帯血中の造血幹細胞を増殖 させることを目指してストローマ細胞との共培養系 を模索していたなかで偶然誘導されてきた.つまり 造血幹細胞として CD34 陽性画分,コントロールと して残りの CD34 陰性画分をそれぞれストローマ細 胞と共培養した.するとまったく予期しなかったこ とであるがコントロールの CD34 陰性の培養からブ ラスト化した T 細胞が出現し,この細胞を解析す ることで新規制御性 T 細胞として確立できたので ある.その後の解析で CD34 陰性画分あるいは全単 核球画分どちらを用いても HOZOT の誘導には差 がないことが判明した. 2. ストローマ細胞の特徴と利用 HOZOT の誘導にはストローマ細胞との共培養が 欠かせない.ストローマ細胞とは実質細胞に対する 間質細胞あるいは支持細胞のことであるが厳密な定 義となると難しい.一般には線維芽細胞と考えれば よいが脂肪前駆細胞として扱われることもあり内皮 細胞やマクロファージさらには上皮細胞を対象とし て含める場合がある8).我々は実験的には培養フラ スコに付着性の非造血系(CD45 陰性)および非内 皮細胞系(CD31 陰性)の細胞をストローマ細胞と して扱っている.HOZOT の誘導に用いるストロー マ細胞は細胞株として樹立されたマウスの骨髄ある いは頭蓋冠由来の細胞である.抗原提示細胞である マクロファージや樹状細胞と比較した場合 MHC のクラス I と CD80 の発現はみられるものの MHC クラス II 陰性,CD86 陰性という違いがある.近 年ストローマ細胞に新たな関心が集まっている.そ れはひとつには間葉系幹細胞として多分化能をもち 再生医療としての応用が期待されるからである.ま た T 細胞に直接あるいは他の細胞を介して間接的 に免疫系に働きかけている証拠があり免疫抑制細胞 としての応用が考えられている9,10).特に HOZOT 誘導と関連して興味深いことはストローマ細胞が Treg の誘導に関わっているという報告があること である11,12). 3. 誘導法とその特徴 実 際 の HOZOT の 誘 導 は 図 2 の よ う に し て 行 う.まず臍帯血から単核球を分離し単核球全画分を 前もって培養したストローマ細胞の上に播く.増殖 因子などを加えることなくそのまま共培養を 2~3 週間続けるとストローマ細胞が障害を受けるのと同 時に blast 化した large granular lymphocyte (LGL) 様の細胞が増殖してくる.ストローマ細胞が障害さ れてなくなるのに伴い新しいストローマ細胞上に blast 細胞を継代して拡大培養を続ける.Blast 出現 から 1~2 週間で増殖が衰えてくるがさらに IL2 を加えることで拡大培養が可能となる.培養中は常 にストローマ細胞との共培養を維持し,凍結保存あ るいは実験使用時にはストローマ細胞の debris 等 を Ficoll-Paque で除去して用いる.Blast 細胞の出
図 2 HOZOT 誘導法 HOZOT の誘導法を簡略化して示した.臍帯血より Ficoll-Paque 遠心法により分離された単核球の全画分をマウススト ローマ細胞と共培養する.IL2 などの増殖因子を加えるこ となく 23 週間培養を続けるとストローマ細胞に対して細胞 障害を示しながら増殖する blast 細胞の出現がみられるよう になる.しばらく IL2 無添加で培養を続けた後,さらに IL2 を加えることで長期の拡大培養が可能となり HOZOT として樹立できる6). 現は常にみられるわけではなく通常 HOZOT 誘導 の効率は 10ほどである. 以上述べた HOZOT 誘導法には従来の T 細胞誘 導法と比べ次のような特徴がある.◯単核球の全画 分を用いる,◯マウスストローマ細胞と共培養する, ◯増殖因子を加えない.まず単核球全画分を用いる 点であるが,一般の T 細胞分化誘導系が CD3 陽性 あるいは CD4 陽性などの精製画分を用いて行われ る点で大きく異なる.単核球全画分には T 細胞以 外に B 細胞,NK 細胞,単球,樹状細胞などが含ま れ多様な細胞種の相互作用の結果 HOZOT が誘導 されると考えている.またマウス細胞と共培養する つまり xenogeneic な刺激で T 細胞を誘導する点も 非常にユニークな T 細胞誘導法である.マウスの かわりにヒトのストローマ細胞を使ったデータはな いが xenogeneic な刺激が強く入ることで HOZOT が誘導できた可能性は高い.また増殖因子特に IL 2 を添加しないで T 細胞の分化誘導を行うのも特徴 である.IL2 の添加は一度樹立された HOZOT を 拡大培養するには必要であるが樹立の初期段階では 不要でありむしろ IL2 添加が HOZOT 誘導にマイ ナスに働くようである. III. 表現型の特徴 1. CD4+CD8+ 一 番 の 特 徴 は CD4+CD8+ の DP 表現型を示す点である.胸腺内の T 細胞のよう に未熟な T 細胞では DP 細胞は一過性に出現する が分化成熟した T 細胞では DP の表現型は極めて まれである.HOZOT 細胞は CD4/CD8 の表現型で は CD4+CD8+,CD4+CD8dim,CD4-CD8+の 3 画 分から構成されており特に DP 細胞を必ず 2060 含んでいる.しかも DP は活性化 T 細胞の場合の ように一過性の発現ではなく安定に維持される.
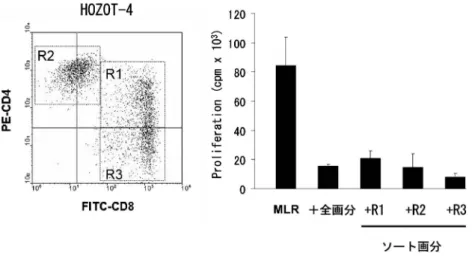
2. Treg 表現型HOZOT が Treg であるという 証拠はまずその表現型にみられた.一般にいわれて いる Treg の 表現型 とは CD25high,GITR+ ,cyto-plasmic CTLA4+,FOXP3+であるが HOZOT は いずれの抗原の発現も陽性である.ただし FOXP3 に関しては 2030のみ陽性である.FOXP3 を高 率にしかも強発現する natural Treg (nTreg)や in-duced Treg (iTreg)の場合と異なり FOXP3 の発現 は抗 CD3/CD28 刺激でも増強されない. IV. 多機能性 T 細胞として 1. サプレッサー活性 HOZOT が Treg 様の表現型を示すことから機能 面でも Treg の活性を示すか検討した.免疫抑制の アッセイ系として樹状細胞を stimulator とし CD4+ CD25-細胞を responder とした allogeneic MLR の 系を用いた.このアッセイ系に HOZOT を加える と顕著な抑制効果がみられた.HOZOT は CD4/ CD8 の表現型からみると heterogeneous な細胞株で あるが 3 つの画分(CD4+CD8+,CD4+CD8dim, CD4-CD8+)をソーティングして調べるといずれ の画分にもサプレッサー活性が見られた(図 3). つまりサプレッサー活性が nTreg と表現型が似て いる CD4+CD8dimにのみに担われているのではな いことを示す.サプレッサー活性を担う因子につい て検討をしたところ既に報告のある細胞表面抗原 あるいは液性因子である GITR,PD1,IL10, TGFb,indoleamine 2,3dioxygenase,galectin10 などの関与は否定された13).特に最近抑制性因子と して注目されている HLA-G の発現が HOZOT に 見られることからその関与を検討したが抗体の中和 実験データからは否定された14). 2. キラー活性 HOZOT が blast 細胞として出現・増殖する過程 で共培養に用いたストローマ細胞に対して細胞障害 を示すつまり xenogeneic なキラー T 細胞として増 殖する.この活性はヒトの CD8,CD3 に対する抗 体で阻害されまたマウスストローマ細胞上の H2 class I 抗原に対する抗体でも阻害されることから MHC 拘束性を持った通常のキラー T 細胞と同様の メカニズムが働いていると考えられる.In vitro で は障害を受ける細胞は正常細胞に限らずマウスの腫
図 3 Allogeneic MLR に対するサプレッサー活性
HOZOT のサプレッサー活性を allogeneic MLR により検討した.HOZOT4 は左図のように CD4/CD8 の表現型を基にすると 3 つの画分より構成されている.HOZOT4 全画分と R1(CD4+CD8+),R2(CD4+CD8dim),R3(CD4-CD8+)をソーティング
した画分(ソート画分)について MLR のアッセイを行ったところいずれのソート画分にもサプレッサー活性が確認できた13).
図 4 ヒトがん細胞に対する in vitro のキラー活性
ヒトのがん細胞に対する HOZOT4 のキラー活性を正常細胞に対する活性と対比させて示した.ST2 はマウスストローマ細胞,
NHDF は正常ヒト皮膚線維芽細胞,Colo320DM と LoVo と WiDr はヒトの大腸がん細胞,MKN5 はヒトの胃がん細胞である. 活性をkilling で示した.HOZOT4 は ST2,ヒト大腸がん細胞ならびに胃がん細胞に対して killing 活性を示したが,NHDF に 対しては活性はみられなかった.抗 CD8 抗体により ST2 に対する活性は阻害されたがヒトがん細胞に対しては阻害がかからな かった. 瘍細胞株にまでおよぶ.さらに驚いたことにはヒト の腫瘍細胞株に対してもキラー活性を示すことが明 らかになった.標的細胞としては大腸がん,胃が ん,乳がん,メラノーマなどの付着性のがん細胞株 に活性を示すものの白血病細胞など浮遊系の腫瘍細 胞株に対しては活性はみられていない.ヒトの正常 細胞として皮膚線維芽細胞や内皮細胞などの細胞株 を用いて調べたが活性はなかった(図 4).ヒト腫 瘍細胞株に対する細胞障害のメカニズムは明らかに マウスの細胞に対する場合と異なっており CD8 や CD3 に対する抗体では阻害がかからない.まだ充 分解明できていないが腫瘍細胞に対する選択性が高 ければ臨床的にも抗腫瘍効果を期待できる. HOZOT は形態的に LGL 様つまり NK 細胞様で あるが K562 に対するキラー活性が見られないこと から NK 細胞とは区別される.また,表現型も NK 細胞マーカーの CD16,CD56,CD161 発現がみら れないか低い.ただし granzyme A および B,per-forin などの細胞障害性の分子は豊富に発現してい る. 3. ヘルパー活性 一見不思議なことであるが HOZOT はサプレッ
図 5 IL10 遺伝子座の intron に存在する STAT 応答性の enhancer
HOZOT では抗 CD3/CD28 抗体刺激時に IL2 を加えることで相乗的に IL10 産生が促進される.このとき IL2/STAT5 の signaling axis の下流に存在する IL10 遺伝子座の enhancer が働いている.この enhancer は IL10 遺伝子座の intron 4 に位置す る STAT 応答性エレメント(I-SRE)で,epigenetic な制御をうけることで IL10 産生能が制御されている17).
サー活性と同時に免疫促進つまり responder T 細 胞の増殖を促進するヘルパー活性も持っている. Allogeneic MLR の系では樹状細胞を stimulator と して用いているが樹状細胞のかわりに抗 CD3 抗体 刺激を行うと HOZOT は responder T 細胞の増殖 を促進する.このとき nTreg はこのような活性を 示さない.また,このヘルパー活性に IL2 が関与 していないことは HOZOT 自身が IL2 を産生しな いこと IL2/IL2R 抗体の blocking が効果のない ことから確認している.おそらく細胞表面抗原を介 した TT interaction の結果と思われるがはっきり した分子の同定はできていない.同じ T 細胞がヘ ルパー活性とサプレッサー活性を共に発揮する例は NKT 細 胞 や germinal center に 存 在 す る follicular helper T cell などの報告があるが15),HOZOT のよ うに同じ responder 細胞に対して刺激の違いだけで 相反する作用を発揮する例はいまだにないと思われ る. 4. IL10 の産生 HOZOT は抗炎症性サイトカイン IL10 を非常 に多く産生する.HOZOT はストローマ細胞と共培 養したとき,あるいは抗 CD3/CD28 抗体で刺激し たときのどちらでも高いレベルの IL10 産生を示 す.この高産生機構には従来知られていなかった IL2/STAT5 のシグナル伝達経路が重要な役割をし ていることを見出した16).HOZOT の IL10 産生 は IL2 を加えることで相乗的に促進されるが,そ の効果は IL2/STAT5 signaling axis を介して IL 10 遺伝子座の 4 番目の intron に存在する STAT 応 答性エレメントが enhancer として働くことによる という事実を見出した(図 5).さらにこの enhan-cer が epigenetic な制御(メチル化脱メチル化)を 受けることにより Treg の中でも IL10 高産生のも のと低産生のものを区別できることが明らかになっ た17).FOXP3 を高発現する nTreg や iTreg におい ては IL2/STAT5 signaling axis が FOXP3 遺伝子 座の intron にある enhancer に働いて FOXP3 の発 現を誘導していることを考えると HOZOT の IL 10 産生誘導と類似のメカニズムを使っていると考 えられる18).ただし HOZOT においてはこの sig-naling axis は FOXP3 誘導には働いていない.
5. ケモカイン産生 IL10 以外に注目すべきサイトカイン類として IL8 および RANTES の 2 つのケモカインがあげら れる.これらのケモカインは他のケモカインと比べ 特に産生量が多くまた産生メカニズムにかなり特徴 がみられる.RANTES はストローマ細胞あるいは TCR の刺激などがない状態でも mRNA あるいはタ ンパクとして高いレベルで蓄積されており刺激に対 し て す ば や く タ ン パ ク を 分 泌 す る . こ の と き mRNA 発現はむしろ刺激前より低下するという興 味深い現象がみられる.一方 IL8 は典型的な誘導 型発現パターンを示し無刺激でほとんど発現のない 状態からストローマ細胞刺激で強く,抗体刺激でさ らに強く mRNA とタンパクの発現が誘導される. ただしこのときタンパク合成阻害剤である cyclo-heximide を加えると mRNA の発現が上昇するとい う superinduction の現象がみられる.RANTES の mRNA およびタンパクを蓄積する性質,IL8 の superinduction という現象はいずれも HOZOT に特 異的というわけではないが HOZOT を特徴付ける 性質・現象として重要と考えておりメカニズムの解
表 1 HOZOT と代表的な Treg 細胞との比較
特 徴 種 類
HOZOT nTreg iTreg Tr1 Th3 NKT
表現型 CD4/CD8 3 つの亜集団より 構成される 1) CD4+CD8+ 2) CD4+CD8dim 3) CD4-CD8+ CD4+CD8- CD4+CD8- CD4+CD8- CD4+CD8- 異なるサブセッ トが存在する (CD4+CD8-, CD4-CD8+, CD4-CD8-) CD25 高発現 高発現 高発現 発現無し 発現無し 発現無し FOXP3 一部発現 高発現 高発現(一過性) 発現無し 発現無し 発現無し
他のマーカー GITR, CTLA4 GITR, CTLA4 GITR, CTLA4 Va24
細胞障害活性 有り 無し 無し 無し 無し 有り
免疫抑制機構 主として液性因子 細胞間相互作用 細胞間互作用 IL10 TGFb ILTGFb, IFNg4, IL10,
IL10 産生 高い 低い 低い 高い 低い 高い
由来 (臍帯血から誘導)不明 胸腺 (活性化で誘導)末梢 胸腺 胸腺 胸腺,末梢
HOZOT と 5 種類の Treg 細胞について表現型,機能,由来について比較した.nTreg は natural Treg, iTreg は induced Treg を意味する.
明が HOZOT を理解する手がかりになると考える. 6. アナジー Treg の 特 徴 は サ プ レ ッ サ ー 活 性 と と も に ア ナ ジー状態を維持していることが挙げられる.アナ ジーとは TCR 刺激に対して増殖不応答でありまた IL2 の産生がみられない状態をいう19).一般にヘ ルパー T 細胞のアナジーは TCR 刺激に対して IL 2 産生に限らずほとんどのサイトカインの産生が不 応答状態となるのに対して Treg 特に HOZOT の場 合は IL2 産生抑制に対し IL10 産生などのある種 のサイトカイン産生はなんら問題なく充分応答がみ られる.Treg のアナジー維持機構は免疫抑制機構 と表裏の関係にあり共通の分子機構が働いている可 能性がありその解明は免疫学の大きな課題として残 されている.nTreg の場合には FOXP3 がアナジー とサプレッサー活性を規定する分子と想定されてい るが20),HOZOT の場合は FOXP3 発現は 2030 にとどまることを考えると FOXP3 以外の分子がア ナジーとサプレッサー活性を制御している可能性が 高い. V. 他の Treg との比較 HOZOT と他の代表的な Treg との比較を表 1 に 示した.nTreg は胸腺で分化し FOXP3 の恒常的で 強い発現を示す一方,iTreg は末梢で抗原刺激など をうけて活性化することで誘導されるが FOXP3 の 発現も抑制機能も一過性で不安定である.Tr1 は IL10 の高産生を示す点で HOZOT とよく似てい る21).Th3 は TGFb の産生で特徴付けられる22). NKT も Treg の一種として分類されるがこれは IL 4,IL10 などのサイトカインが抑制的に働くから である23).いずれの Treg にしても HOZOT と比べ CD4/CD8 の表現型とキラー活性の有無で大きく異 なるものの明確な分類には nTreg における FOXP3 のように HOZOT を規定する分子の同定が必須で ある. VI. In vivo での役割 HOZOT は in vitro で培養によって誘導された T 細胞である.成熟 T 細胞としては CD4+CD8+とい うユニークな表現型から考えると HOZOT が単な る in vitro の人工産物という可能性もある.In vivo に HOZOT に対応する counterpart が存在するかど うか臨床応用や疾患との関連を考える場合に重要に なってくる.CD4+CD8+という表現型に注目する と腸管上皮に存在する IEL (Intraepithelial lympho-cyte)がその候補のひとつである24).マウス IEL に は LGL 様の形態をした CD4+CD8+細胞が存在し IEL の 10ほどを構成する.腸管には様々な種類 の Treg が存在し経口免疫寛容や腸内菌叢に対する ホメオスタシス維持に関わっていること25),また IEL のマーカーといわれる CD103 が HOZOT に発 現していることから HOZOT を腸管由来の Treg と 考えることも可能である.ただし IEL 中の DP 細 胞 は HOZOT と 異 な り CD8 の 発 現 が CD8aa の homodimer で あ り , CD8ab を 持 つ HOZOT の
図 6 HOZOT の T 細胞生物学における位置付け T 細胞の分類における HOZOT の位置付けを図示した. CD4+CD8+の表現型を特徴とした HOZOT はサプレッサー 活性,キラー活性,ヘルパー活性いづれも保持した多機能性 T 細胞と考えられる. counterpart とは必ずしもいえない.腸管ではない がヒト健常成人の末梢血中には 510ほどの高い 割合で DP の T 細胞を持つ例が何例かあることを 我々は観察しておりこういった細胞が HOZOT の in vivo counterpart の可能性も考えている. VII. 臨床応用へのみち HOZOT の示すサプレッサー活性あるいはキラー 活性を in vivo でも発揮させることで臨床応用を期 待している(図 6).キラー活性の臨床応用の一歩 として in vivo 抗腫瘍活性を NOD/SCID マウスを 用いて検討している.ヒト大腸がん細胞株 WiDr を 腹腔に移植し HOZOT を投与する腹膜播腫モデル で検討したところ HOZOT 投与で WiDr の腹膜播 腫 が 顕 著 に抑 制 で き る こ と を 確 認 し た . 一 方 in vivo でのサプレッサー活性をみるためにはやはり マウスの HOZOT counterpart を見出し疾患モデル での治療を検討する必要がある. VIII. 今後の課題 我々が現在取り組んでいる主要な課題は 3 つあ る.ひとつは前項でも述べたようにマウスモデルの 開発である.マウスのリンパ球を単純にマウススト ローマ細胞と共培養しても blast 細胞の出現はみら れず培養法やマウス系統の選択など何らかの工夫が 必要である.もうひとつの課題は誘導メカニズム の 解 明 で あ る . そ の た め に も 現 在 10 程 度 の HOZOT 誘導効率を 100近くまで上げ安定したア ッセイ系を確立する必要がある.この点に関しては 培養法の改良でかなり効率を上げることが可能にな ってきている.3 つめは HOZOT のマーカー遺伝子 あるいは規定遺伝子の同定である.現在 HOZOT の マ ー カ ー と し て CD4+CD8+ 表 現 型 , IL 10 / RANTES 高 発 現な ど を 指標 に し てい る が明 確 に HOZOT を決定する分子・遺伝子がない状況である. nTreg や iTreg の FOXP3 に相当する規定遺伝子の 同定が HOZOT 研究発展の鍵と考えている. IX. ま と め HOZOT の発見とその後の研究を通して感じたの は驚きの連続であったということである.陰性コン トロールからの blast の出現,CD4+CD8+ という 異例な表現型,サプレッサーとヘルパーという相反 する活性の検出,免疫していないにもかかわらずヒ ト腫瘍細胞に対してキラー活性を示すことなど予想 外の結果が次々と現れてきた.このような多様な性 質を示す細胞の研究はその意義解釈には困難が伴う が,我々はむしろ HOZOT という細胞が大きな発 展性を持っていると考えている.かつて生命科学の 歴史の中では enzyme hunting や gene hunting とい った新しいものの発見・同定が活発に行われた時代 があった.今の時代は Treg や樹状細胞の発見, iPS の作製にみられるように cell hunting の興奮の な か に あ る の か も し れ な い . 我 々 の 見 出 し た HOZOT もその一つとして研究に臨床にと発展させ てゆきた. 謝辞本総説の内容は株式会社林原生物化学研究 所研究センター基礎細胞研究部門を中心に行われた 研究についてまとめたものです.共同研究にあたり まして倉敷成人病センターの山o史行先生,岡山大 学医学部腫瘍病理学教室の近藤英作先生に深謝いた します.また研究内容につきましてご助言ご指導を いただきました産業技術総合研究所の辻典子先生に 深謝いたします. 文 献
1) Sakaguchi S. : Regulatory T cells in the past and for the future. Eur J Immunol 38 : 901 937, 2008.
2) Sakaguchi S. : Naturally arising CD4+ regula-tory t cells for immunologic self-tolerance and negative control of immune responses. Annu Rev Immunol 22 : 531562, 2004.
3) Hori S, Nomura T, Sakaguchi S. : Control of regulatory T cell development by the transcrip-tion factor Foxp3. Science 299 : 10571061, 2003.
4) Ziegler SF. : FOXP3 : of mice and men. Annu Rev Immunol 24 : 209226,2006.
5) Shevach EM. : From vanilla to 28 ‰avors : mul-tiple varieties of T regulatory cells. Immunity 25 : 195201, 2006.
6) Nakamura S, Suzuki M, Sugimoto A et al. : IL 2independent generation of FOXP3(+)CD4 (+)CD8(+)CD25(+) cytotoxic regulatory T cell lines from human umbilical cord blood. Exp Hematol 35 : 287296, 2007.
7) 中村修治,木畑正義抗がん作用,免疫抑制
作用をもつ血液細胞「HOZOT」. BIONICS. 2007 年 3 月号 pp8487.
8) Svensson M, Kaye PM : Stromal-cell regulation of dendritic-cell diŠerentiation and function. Trends Immunol 27 : 580587, 2006.
9) Uccelli A, Pistoia V, Moretta L. : Mesenchymal stem cells : a new strategy for immunosuppres-sion?Trends Immunol 28 : 219226, 2007. 10) Nauta AJ, Fibbe WE. : Immunomodulatory
properties of mesenchymal stromal cells.Blood 110 : 34993506, 2007.
11) Di Ianni M, Del Papa B, De Ioanni M et al. : Mesenchymal cells recruit and regulate T regulatory cells. Exp Hematol 36 : 309318, 2008.
12) Poggi A, Zocchi MR. : Role of bone marrow stromal cells in the generation of human CD8+ regulatory T cells.Hum Immunol 69 : 755759, 2008.
13) Suzuki M, Sugimoto A, Harashima A et al. : Novel mechanisms of suppressor activity ex-hibited by cytotoxic regulatory T cell lines, HOZOT.Exp Hematol 37 : 92100, 2009. 14) Feger U, Tolosa E, Huang YH et al. : HLA-G
expression deˆnes a novel regulatory T-cell sub-set present in human peripheral blood and sites of in‰ammation. Blood 110 : 568577, 2007. 15) Marinova E, Han S, Zheng B. : Germinal
cen-ter helper T cells are dual functional regulatory cells with suppressive activity to conventional CD4+ T cells. J Immunol 178 : 50105017, 2007.
16) Tsuji-Takayama K, Suzuki M, Yamamoto M et al. : IL2 activation of STAT5 enhances production of IL10 from human cytotoxic regulatory T cells, HOZOT.Exp Hematol 36 : 181192, 2008.
17) Tsuji-Takayama K, Suzuki M, Yamamoto M et al. : The production of IL10 by human regula-tory T cells is enhanced by IL2 through a STAT5responsive intronic enhancer in the IL 10 locus.J Immunol 181 : 38973905, 2008. 18) Zorn E, Nelson EA, Mohseni M et al. : IL2
regulates FOXP3 expression in human CD4+ CD25+ regulatory T cells through a STATde-pendent mechanism and induces the expansion of these cells in vivo. Blood 108 : 15711579, 2006.
19) Schwartz RH. : T cell anergy. Annu Rev Im-munol 21 : 305334, 2003.
20) Sakaguchi S, Yamaguchi T, Nomura T et al. : Regulatory T cells and immune tolerance.Cell 133 : 775787, 2008.
21) Roncarolo MG, Bacchetta R, Bordignon C et al. : Type 1 T regulatory cells. Immunol Rev 182 : 6879, 2001.
22) Weiner HL. : Induction and mechanism of ac-tion of transforming growth factor-beta-secret-ing Th3 regulatory cells. Immunol Rev 182 : 207214, 2001.
23) Taniguchi M, Harada M, Kojo S et al. : The regulatory role of Valpha14 NKT cells in innate and acquired immune response.Annu Rev Im-munol 21 : 483513, 2003.
24) Cheroutre H. : Starting at the beginning : new perspectives on the biology of mucosal T cells. Annu Rev Immunol 22 : 217246, 2004. 25) Tsuji NM, Kosaka A. : Oral tolerance :
intesti-nal homeostasis and antigen-speciˆc regulatory T cells.Trends Immunol 29 : 532540, 2008.