コーヒー(Coffea arabica, Coffea canephola var. robusta)の機能性食品素材としての利用に関する研究
142
0
0
全文
(2) 博士学位論文 コーヒー (Coffeaarabica, C o f f e ac a n e p h o l ava 巴r o b u s t a )の 機能性食品素材としての利用に関する研究. 7年 3月 平成 1. 近畿大学大学院農学研究科 応用生命科学専攻(指導:藤田藤樹夫教授) 岩井和也. 2.
(3) (和文題目). コーヒー CCojj旬 。rabicα,Coffeacanepholav a r .r o b u s t a )の 機能性食品素材としての利用に関する研究. 近畿大学大学院農学研究科 応用生命科学専攻 岩井和也 (指導:藤田藤樹夫教授). (英文題目). U t i l i z a t i o no fC o f f e e( C o l 作αa r a b i c a,Col 作 。c anψh o l av a r .r o b u s t a ) a saP h y s i o l o g i c a lF u n c t i o n a lFoodM a t e r i a l .. KazuyaIwai. March2005. KinkiU n i v e r s i t y GraduateS c h o o l, D i v i s i o no fA g r i c u l t u r a lS c i e n c e M a j o r :AppliedB i o s c i e n c e ( A d v i s o r :P r o f . T o k i oF u j i t a ). Submittedt ot h eGraduateS c h o o l,KinkiU n i v e r s i t y ,t of u l f i l lt h e requirementf o rt h eD o c t o r a t eD e g r e e . 3.
(4) コーヒー (Coffeaarabica,C o j j 匂 c αn e p h o l av a r .r o b u s t a )の 機能性食品素材としての利用に関する研究. U t i l i z a t i o no fC o f f e e(C~併a a r a b i c a,C o j j 匂 c a n e p h o l av a r .r o b u s t α ,) a saP h y s i o l o g i c a lF u n c t i o n a lFood乱1 a t e r i a l . 岩井和也. 目次. 緒 論 . . . . ・ ・ . . . . . . . . ・ ・ . . . . ・ ・ … . . . . ・ ・ . . . . ・ ・ . . . . ・ ・ . . . . ・ ・-……… ・・ . . ・ ・ … . . . . ・ ・ . . . . ・ ・ …. . 6 H. 第一章. H. H. H. H. H. H. H. H. H. H. H. コーヒー抽出残 j 査に含まれるアラピノガラクタンの分離と酵素分解. 物ガラクトオリゴ糖の機能性評価....・ ・ . . . . ・ ・ . . . . ・ ・ . . . . ・ ・ . . ・ ・ … . . . . ・ ・ . . . . . ・ ・ …1 1 H. H. H. H. H. H. H. 1-1.緒言 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 1 . 実験材料および実験方法… . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 3 1・2 …. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 0 1 3 . 実験結果および考察....・ ・ H. ・ 4 . まとめ...・ ・・・ . . . . . ・ ・・・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・・・ . . 3 1 1 H. 第二章. H. H. H. H. H. H. H. H. H. H. H. H. H. H. コーヒ一生豆に含まれるクロロゲン酸類の分離とその機能性評価 3 3. 2・ 1 . 緒 言 . . . . ・ ・ … . . . . ・ ・ … . . . . ・ ・ …. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 3 H. H. H. 2・2 .実験材料および実験方法 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 4. 2・3 .実験結果および考察....・ ・ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 6 H. 4 . まとめ…...・ ・ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6 8 2・ H. 第三章. クロロゲン酸を用いた機能性成分の酵素合成....・ ・ … ・・ . . . . . ・ ・ …. 6 9 H. H. H. H. 3・1.緒言 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6 9 4.
(5) 3 2 . 実験材料および実験方法 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7 2. 3・4 . まとめ...・ ・・・ . . . . . ・ ・ . . . . . ・ ・・・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . … .113 H. H. H. H. H. H. H. H. H. H. H. H. H. 総 合 考 察 … ・ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .1 1 5. S u m m a r y . . . .・・ …. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 2 1 H. 説 話 宇 . . . ・ ・ . . . . . ・ ・・・ . . . . . ・ ・・・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . . . . ・ ・ . . … … .127 H. H. H. H. H. H. H. H. H. H. H. H. H. H. 参 考 文 献 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .1 28. 5.
(6) 緒論. コーヒーは古くから曙好品として世界中で愛飲されてきた。近年、日本でも. 0万 t余りのコーヒ一生豆が輸入され、 260万 kLのコーヒー飲料が製造 年間 4 されている。それに伴い、製造現場から排出される様々な廃棄物が発生してい る 。 この現状に対して製造者側は、技術的・経済的に可能な最大限の環境対策を 講じることが求められ、環境対策としてどのような対応を行っているのか、そ れが技術的、経済的に最大限可能なものであるのか常に検証し、環境報告書、 環境会計等を通じて社会に情報を提供し、消費者から正当な評価を受けること 査や、 が求められている。コーヒー飲料製造工場から排出されるコーヒー抽出残j 最終製品にならない欠点豆などは排出される量が多いことから、これらコーヒ 一生豆由来の未利用資源を有効活用する手段の確立が求められている。 査には、マンナンやア コーヒー飲料製造工場から排出されるコーヒー抽出残 j. 0 ' " ' ' 5 0 % (乾燥重量)含まれており ラピノガラクタンなどのヘミセルロースが 4 査は、ボイラーの燃料や堆肥などに再 有望な資源である。しかしコーヒー抽出残 j 利用されるケースがほとんどであり、バイオマス資源の有効利用と呼ぶために は更に付加価値の高い利用方法を考えることが必要である。 そこで著者らはコーヒー立、あるいはコーヒー抽出残直に含まれるアラピノ ガラクタンが、. 0・(1→ 3 ) -結合からなる主鎖と 0・ ( 1 →6 ) -結合からなる側鎖で構成. I Iアラピノガラクタンであることに注目した。この残j 査を酵素分解 されたType してオリゴ糖を得ることができれば、プロバイオティクスとしての利用だけで なく、糖鎖工学における有用素材として利用できると期待した。 第一章では、コーヒー豆由来のアラピノガラクタン分解微生物を探索し、. A s p e l 宮i l l u sルm i g a t u sN o . 2 3 2が生産するガラクタナーゼを精製して酵素化学的諸. 6.
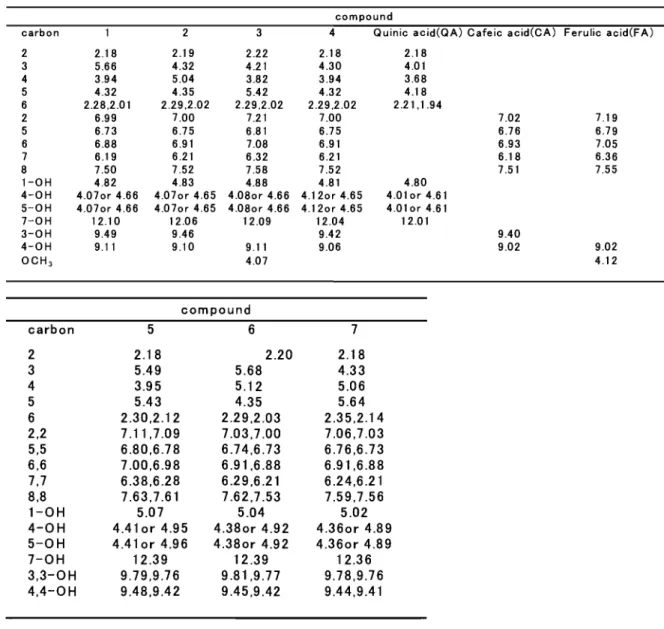
(7) 性質を明らかにした。つぎに本酵素をアラピノガラクタンに作用させて得られ た生成物の機能性も評価したヘ 一方、コーヒー豆を賠煎する前に形を崩した欠け豆や未成熟豆など等級規格 外のコーヒー豆は最終製品にはならず、有効利用されていないのが現状である。 そこで著者は、これら規格外の豆を機能性食品素材として利用する方法を考え、 豆に含有されるクロロゲン酸類に注目した。 カフェー酸はベンゼン環(フェニル、 C 6 ) に炭素 3個のプロパン ( C 3 ) が結 合した基本骨格をもっ化合物群(フェニルプロパノイド;桂皮酸誘導体)の一. C 6 C 3 J 1単位で構成されているものから、 2 " ' 4単位が縮合したリグナ 種で、 r ン、さらに多数縮合したリグニンなどの形で植物界に広く分布している 2)。キナ. a f f e o y l qmmc a c i d (CQA, 酸にカフェー酸がモノエステル化したものが c C h l o r o g e n i ca c i d )、ジエステル化すると d i c a f f e o y lq u i n i ca c i d(diCQA)となる ( F i g .1 )。. 正ぞ. HO、 / " ' -. ~人 γγ~ 'OH. HO 〆. OH. ¥f. Q u i n i ca c i d(QA). C a f f e i ca c i d(CA). にぷH :. み H. C a f f e o y lq u i n i ca c i d(CQA). o~号. む 了10/¥J三. 3, 5・d iC a f f e o y lq u i n i ca c i d ( 3, 5-diCQA). 2・C a f f e i ca c i dp h e n e t h y le s t e r (2-CAPE). F i g .1 .Ty p i c a lh y d r o x y c i n n a m i ca c i da n di t sd e r i v a t i v e s . 7.
(8) クロロゲン酸類はタケ類 3)、ノラニンジン (D a u c u sc a r o t a ) 4 )、キク科植物. 、マ. 5 8 ). l e xp a r a g u a r i e n s i s ) 9 )、オナモミ (x a n t h i u mo c c i d e n t a l e )10,11)、ナシ ( P y r u s テ茶 ( J a p p a )13)、 ペ ル ー 産 生 薬 P a j a r oB o b o ( T e s s a r i a p y r i f o l i a )12)、 ゴ ボ ウ Urctium l. P r u n u sd o m e s t i c a i n t e g r i f o l i a )14) 、 プ ル ー ン (. ) 1 5 )、. ヤーコン ( S m a l l a n t h u s. S p o n d i a sm o m b i n ) 1 7 )、熱帯産果実 s o n c h i f o l i u s )1 6 )、ウルシ (. 1 8 )などに含まれている. ことが知られており、様々な生理機能を有することから近年注目されている物 質である。コーヒ一生豆にもクロロゲン酸類が含まれているが、私たちが飲用 するコーヒーは賠煎した豆を抽出して飲用するために、高温で熔煎する過程で クロロゲン酸類は糖類と反応して香気成分あるいは褐色色素成分へ変化する。 したがって、賠煎が進むにつれてクロロゲン酸類は急速に消失してしまう。し かし、飲用に適さない等級外の生豆からクロロゲン酸類を取り出し、機能性成 分として利用していくことは未利用資源の有効利用へとつながり、循環型社会 に貢献すると期待している。 第二章では、クロロゲン酸類が成熟の早い段階から豆に蓄積されることと、 コーヒーの品種の違いによりクロロゲン酸類の含量が異なることを明らかにし た。また、生立からクロロゲン酸異性体の単離方法を確立し、得られたクロロ ゲン酸異性体の生理活性を i nv i t r oで、比較し、 diCQAに高い活性を見いだした。 さらにクロロゲン酸を機能性食品素材として缶コーヒーに添加することを考え、 缶コーヒー中のクロロゲ、ン酸類の挙動について分析した. 。. 1 9 ). 第三章では、クロロゲン酸を原材料により高い生理機能を持つ物質へ酵素変 換する方法を確立し、得られた生成物の機能性を検討した。 カフェー酸にキナ酸以外の物質がエステル結合したカフェー酸エステル類は. F i g .1 )、多様な生理活性を持つことが報告されている。 植物に広く存在し (. C a f f e i ca c i dp h e n y l e t h y le s t e r(2-CAPE) はプロポリスに含まれる生理活性物 質で、抗酸化、抗菌、抗炎症作用、腫蕩細胞増殖抑制効果があると報告されて. 8.
(9) いる. 2 0 2 2 ) 。古代エジプトでは死体をミイラ化するための防腐・保存用に、また. 東欧では外科、皮膚科、耳鼻咽喉科、歯科、婦人科、獣医科などのさまざまな 医療領域に用いられてきた. 2 3 )。 C henら 24)はプロポリスから単離した. 2-CAPE. がヒト白血病培養細胞 HL-60にアポトーシスを誘導する機構を調べ、 2-CAPE がカスパーゼ 3の活性化と、. B c l 2の発現を抑制し Baxの発現を活性化するこ. とによりアポトーシスを誘導していること明らかにした。 多様な生理活性をもっ 2-CAPEや diCQAを化学合成する方法として、たとえ ばカフェー酸を塩化チオニルなどで塩素化した後、 2 P h e n e t h y la l c o h o l ( 2・pA) と反応させる方法など多数報告されている. 2 5 2 7 )。しかし反応に使用した酸触媒. や塩素化剤を除去する必要があり、環境に優しい製造方法という点で問題が残 る。さらに、 diCQAの化学合成方法は著者が調べた限りでは未だない。 そこでまず A s p e r g i l l u sj a p o n i c u sが生産する CQAe s t e r a s e を用いて CQA と 2-PAから 2-CAPEへの変換条件を検討した。この研究過程で本酵素が加水分 解反応以外にあらたにエステル置換反応と縮合反応を触媒することを見いだし た。そしてエステル置換反応で、最も高い 2-CAPE変換率 (CQAに対するモル変 換率 50%)が得られた。一般に水存在下の環境で加水分解酵素を作用させると、 加水分解反応が優先的に進行することが知られている。本酵素が水共存下でエ ステル置換反応を高収率 ( 5 0 % )で進行できたのは、エステル置換生成物が 2-PA 層に移行して水層に存在する酵素との接触を免れたためと考えられる。 つぎ、に V i n y lc a f f e a t eと CQAから diCQAに酵素変換する方法を考えた。予 備実験として市販の V i n y lcinnamateと CQAから Cinnamoylc a f f e o y lq u i n i c. a c i d(CCQA)に変換できる市販リパーゼを探索したところ、リバーゼ、 ALのみが CCQAに変換した。確立した最適変換条件下での CCQAモル変換率は 19.2%で i n y lc a f f e a t eと CQAから diCQAへの変換反応を触媒しな あった。本酵素は V かったが、酵素を広く探索すれば、 diCQAに変換できる酵素を見つけることが. 9.
(10) できると確信している。. 2-CAPEや diCQAは多様な生理活性をもっ物質で、天然物には微量しか含ま れていない。これらの物質を安定供給する方法として、コーヒ一生豆から抽出 した CQAを基質として 2-CAPEにワンポット反応で酵素変換できることを明. s t e r a s e らかにした。また diCQAについても生豆から抽出した CQAを CQAe で、加水分解し、得られたカフェー酸を V i n y lc a f f e a t e に化学変換した後、 diCQA へ酵素変換できる見通しをつけることができた。. 1 0.
(11) 第一章. コーヒー抽出残澄に含まれるアラピノガラクタンの分離と酵素分解物. ガラクトオリゴ糖の機能性評価. 1-1.緒言 アラビノガラクタンは水溶性ヘミセルロースの一種で、全ての植物に共通す る細胞壁構成成分であり、ガラクトースからなる主鎖にアラピノースもしくは ガラクトースからなる側鎖が結合した多糖類の総称である。アラピノガラクタ ンは構造の違し、から町r p e -1と Eに大別されている. r p e -1アラビノガラ. 2 8 )。司. クタンは 0・ u→ 4 )・ガラクトピラノシド結合をもっ主鎖と α-L-アラピノース残 基を有する側鎖が結合している。Ty p e I Iアラピノガラクタンは、 0・ u→3 )・ ガ ラクトピラノシド結合からなる主鎖と α-L-アラピノースで修飾された 0・ u→ 6 )・ガラクトシド結合からなる側鎖で構成され、ごく微量のキシロース、ラムロ ース、フコース、グルクロン酸などを含んでいるのが特徴である ( F i g . 2 )。 Ty p e I Iアラビノガラクタンは、アズキ種子. 2 9 )、果実のパルプ成分 30-31)、当. A n g e l i c aa c u t i ! o b a )3 2 )、ゼニアオイ (Malvam a u r i t i a n a ) 3 3 )などの他に、コーヒ 帰 ( C o j j 匂 c anephorav a r . ー豆匹乳部に多く含まれている。たとえばロブスタ種 ( r o b u s t a ) のTy p e I Iアラピノガラクタン含量は 48% (乾燥重量あたり)で、ア. ラピカ種 ( C o j j 初 α r a b i c a ) より 3%ほど多い. 3 4 )。. HO HO. F i g .2 .S t r u c t u r eo fa r a b i n o g a l a c t a nf r o mc o f f e eb e a n . 1 1.
(12) 粉砕したコーヒー賠煎豆を熱水抽出した残澄には、セルロース、(ガラクト)マ ンナン、アラビノガラクタンなどの多糖類が 40'""50%以上(乾燥重量)残存し ている。そこでコーヒー豆中の多糖類、特にアラピノガラクタンを酵素的に分解 することにより得られたオリゴ糖がプロバイオティクスや糖鎖工学における有 用素材して利用できると期待した。 コーヒー豆に含まれる多糖類を酵素分解する方法として、清澄なコーヒー抽 出液を製造するために細胞壁溶解酵素などを使用した例がある. 35). が、酵素分解. 物を利用する目的でコーヒー豆に含まれる多糖類を酵素処理した報告はまだな. し 、 。 細菌や糸状菌由来の Endo-s・ ( 1 →C・ D-ガラクタナーゼ (EC3 . 2 .1 .8 9 )はTy p e EアラピノガラクタンやTy p e I Iアラビノガラクタンータンパク質を分解しない. と報告されている. 3644)。 F e l l e r i nら 46) は 、. A s p e l 宮i l l u sn i g e rの培養液上清から. Endo-s・ ( 1 →6 )・ガラクタナーゼを精製している。この酵素は、 0・ ( 1 →6 )結合を ( 1 → 6)-D-ガラクトピオー 主鎖とするアラビノガラクタンを特異的に分解し、 0・. : 0 . 2 6のモル比で生成した。しかしアラピノースを除去し スとガラクトースを 1 たブドウ由来のガラクタン中の 0・ ( 1 → 6)-D-ガラクタンを 50%分解したが、 0・ ( 1 →3 ) -結合したガラクトース側鎖は分解せずにそのまま残存したと報告している。. Hashimotoら 47) は、ふすま培養した R h i z o p u sn i v e u sが生産した Endo-s・ ( 1 → 3 ) D・ガラクタナーゼ (EC3 . 2 .1 .9 0 ) の部分精製酵素が、コーヒー豆由来のアラ ピノガラクタンを数種類のガラクトオリゴ糖とガラクトース、アラピノースに 分解することを見いだしているが、その分解率は 8.9%に留まっている。. F u j i t aらは. 、コーヒ一生豆から分離した A s p e r g i l l u sルm i g a t u sNo.232が. 48,49). 生豆由来のアラピノガラクタンをアラピノース、ガラクトース、ガラクトオリ. .ルm i g a t u sNo.232が生産す ゴ糖に分解することを見出しており、本実験では A る酵素を電気泳動的に単一バンドまで精製し、酵素化学的諸性質を明らかにし. 1 2.
(13) た 。. 1 2 . 実験材料および実験方法. 1 2・1.コーヒー抽出残澄よりアラピノガラクタンの抽出 donesiaAP-l)のコーヒー抽出残 j 査 試料にはインドネシア産ロブスタ種(In を使用し、 Hashimotoら 47) の方法を参考にアラピノガラクタンを抽出した。 査 100g に蒸留水 ( 1 5 0 0 ' " ' " '2000m I)を加え高圧で加熱抽出 コーヒー抽出残 j. ( 1 2 1C、2-4時間)した。抽出液を減圧濃縮し、生じた沈殿を除去した上清に 0. 3 ' " ' " ' 4倍量のエタノールを加えて多糖類を沈殿させた。得られた沈殿物を少量の 水に溶解して水に透析後、凍結乾燥したものを粗アラピノガラクタンとした。. . 0 g 粗アラピノガラクタンは以下の手順で精製した。粗アラピノガラクタン 2 を 0.2M水酸化ナトリウム溶液 100ml に溶解し、窒素気流を通じながら室温. ( 2 5C) で 3時間、ついで 5 5 ' " ' " ' 6 0Cで 3時間撹枠した。冷却後、硫酸で pH7.5 0. 0. に調整しクロロホルム 80ml、酢酸エチル 120ml及びジエチルエーテル 120ml で順次抽出した。水層を蒸留水に透析したのち、凍結乾燥したものを精製アラ ピノガラクタンとした。. 1 2・ 2 .精製アラピノガラクタンの TLC分析 精製アラピノガラクタンの構成糖を TLCで分離検出した。試料 10mgと蒸留 水 2mlをネジ付き試験管に入れ、 80Cで 30分間予備加温した。ついで、 O.lN 0. トリプルオロ酢酸溶液 2ml加えて 100Cで 60分間加水分解した。得られた試料 0. 溶液は室温で冷却し、減圧乾回したものを少量の蒸留水に溶かしてサンプルと した。市販アラピノースとガラクトースを標準物質として使用し、シリカゲル. 60F254プレート (Merck) にスポットした。これを酢酸エチル:酢酸:H2U/3:. 13.
(14) 3:1 ( v : v : v ) で展開後、リンモリブデン酸で発色させた。. 1 2・3 .精製アラピノガラクタンの平均分子量 精製アラピノガラクタンの分子量をゲル漏過クロマトグラフィーで求めた。. 50mMNaCI溶液で膨潤させた SephadexG-200ゲルろ過カラム ( φ1 .15cmX 80.715cm) に試料溶液を展開した。溶出液中のアラビノガラクタンは、フェノ ールー硫酸法. 5 0 )で検出した。標準物質には. P u l l u l a n (昭和電工 TKK製 、. MW212, 000、 1 1 2, 000、22, 800、 1 1, 800および 5, 9 0 0 ) と Bluedextran2000 を用いて、その溶出量と標準物質の分子量から回帰式を作成した。回帰式は以 下の式を用いた。. Mobility=Ve-VoNt-Vo Vo=Voidvolume:ゲルベッドの保水量 Ve=Elutionvolume:サンプルの溶出量 Vt=ゲルベッドの体積. 1 2・4 . その他の試料 カラマツ由来アラピノガラクタンは東京化成工業側、他の多糖類(アラビア ガム、カードラン、マンナン、カルボ、キシメチルセルロース、アミロース)は. SigmaChemical C o .,( S tL o u i sMO) より購入した。 SP-SephadexC-50 と SephadexG-I00は Pharmacia (U p p s a l a, Sweden)より、 U l t r a f r e e 4 (限外櫨 過膜ユニット)は日本ミリポア(槻より購入した。その他の試薬はナカライテス ク(槻より購入した。. 1 4.
(15) 1 2・5 .As per g i J J usfumigatusNo.232の培養 A .ルm i g a t u sNo.232 ( F i g .3 ) 胞子懸. 106spores/mD を Table1に示し 濁液 ( た培地に接種し、 40Cで 7日間培養後、 0. 培養ろ液を粗酵素液とした。. 1 2 6 . 酵素活性の測定 1% (w/v) ア ラ ピ ノ ガ ラ ク タ ン を 0.02M酢酸緩衝液 ( pH4.6) に溶解し たものを基質とした。基質 0.1mlに酵. .1mlを添加後、 45Cで 30min F i g .3 .L i g h tands c a n n i n gm i c r o g r a p h s 素溶液 0 0. 問反応させた後、生成した還元糖量を. ofAルmiga ωNo.232.. SomogyiNe l s o n 法でガラクトースを 基準として定量した。組酵素液 1ml当たり 1min聞に 1μM 還元糖(ガラクト ースを基準として)を遊離させ る活性を 1 単位と定義した。. T a b le1 .C o m p o s i t i o no fc u l t u r emedium. Ara b i n o g a l a c t a n M e t h i o n i n e K2HP0 4 7H2 0 MgS04 ・ KCI pH. タンパク量はウシ血清アルブ ミンを標準物質として Lowry 法 51)によって定量した。. 0 . 5 0 . 1 0 . 1 0. 0 5 0. 0 5 6 . 0. 1 2 7.粗酵素溶液の調製 A .ルm i g a t u sNo.232をカラマツ由来アラピノガラクタン添加培地 ( Table1 )5. Lに 400Cで 7日開通気撹枠培養 (0.5vvm、 100rpm) した。培養液をガーゼと o . 5 0 )で、各 2回ずつ糠過して菌体と胞子を除去した。滴液は 80% 漉紙( 東洋滴紙 N 飽和 (NH ) 2 S04で塩析し、得られた沈殿は 0.02M酢酸緩衝液 ( p H 4 . 6 )で、透析後、 4. Ul t r a f r e e 4限外滅過ユニッ トを用いて濃縮した。これを組酵素溶液とした。 1 5.
(16) 1・2・8 . 陽イオン交換カラムクロマトグラフィー 透析した酵素溶液 ( 1 2 0 m ! ) を 20mM酢酸緩衝液 (pH4 . 6 ) で平衡化した. SP-SephadexG-50 カラム (2.8X40cm) に吸着させた。同緩衝液で洗浄後、 NaCl( 0 " ' 0 . 5 M )グラジエントをかけてガラクタナーゼを溶出した ( 6 0 m l l h r )。 溶出液は 3mlずつ分取し、タンパク量とガラクタナーゼ活性を測定して、活性 画分を回収した。. 1 2・9.Sephade 玄 G -100によるゲル滴過 得られた活性画分は 0.02M酢酸緩衝液 (pH4 . 5 ) で平衡化した Sephadex. m l l h r ) した。溶出液は 0.8ml G-100カラム(1.5X100cm)を用いてゲ、ル櫨過(15 ずつ分取しタンパク量とガラクタナーゼ活性を測定し、活性画分を回収した。. 1 2・1 0 .電気泳動と分子量 12.5%アクリルアミドゲルによる SDS-PAGEによって分子量を測定した。分 子 量 マ ー カ ー に は myosin (200, 000Da)、 E .c o/ i 由 来 0・g a l a c t o s i d a s e. ( 1 1 6, 250Da)、ovalbumin ( 4 5, 000Da)、c a r b o n i canhydrase ( 3 1, 000Da)、 t r y p s i ni n h i b i t o r( 2 1, 500Da)、lysozyme ( 14, 400Da)、a p r o t i n i n( 6, 500Da) をイ吏用し、 CoomassieB r i l l i a n tb l u eR-250(Rapids t a i nCBBk i t,ナカライテス ク)で、ゲ、ルを染色した。. 1 2・1 1.ガラクタナーゼの最適 pHと温度および安定性 a ) 最適 pH pH2 . 0 " ' 8 . 0に調製した緩衝液 ( p H 2 . 0 " ' 3 . 5,50mMg l y c i n e H C lb u f f e r; c e t a t eb u f f e r;p H 6 . 5 " ' 8 . 0,50mMphosphateb u f f e r ) p H 4 . 0 " ' 6 . 0,50mMa. 16.
(17) にアラビノガラクタンを 1%( w / v )となるように溶解させたものを基質に用いて 酵素活性を求めた。. b)pH安定性 上記緩衝液にガラクタナーゼを透析 ( 4C、24時間)後、最適 pHで酵素活性 0. を測定し、最も活性が高かった pHでの活性値を 100%として相対活性で表した。. c )最適温度. 20mM酢酸緩衝液 ( p H 4 . 6 ) に 1%( w / v ) アラピノガラクタンを添加したもの 0. 0. を基質とし、 35Cから 85Cの間で 30分間反応させ、最も活性が高かった温度 を 100%として相対活性で表した。. d)熱安定性 0. 3 5 ' " ' " '85Cで 30分間酵素溶液をインキュベートした後冷却、最適温度で酵素活. 性を測定した。最も活性が高かった値を 100%として相対活性で表した。. e )金属塩が酵素活性に与える効果 2 2 2 2 +、Cu +、Mg2+、Al 8種類の金属イオン (Mn2 +、L i +、Fe 3 +、Hg2+) +、Zn. と EDTA 、 D T T ( D i t h i o t h r e i t o I ) を 1mM添加したときのガラクタナーゼ活性を、 対照(無添加)を 100%とする相対値で表した。. 。酵素動力学的特徴 . 6 ) に溶解した 1 ' " ' " '10% ( w / v ) 生豆由来アラピノガ 20mM酢酸緩衝液 (pH4 ラクタンもしくはカラマツ由来アラビノガラクタンを基質とし、 45Cで 30分間 0. r -Burk プロットで表示し 酵素反応させたときの還元糖生成速度を Lineweabe. 1 7.
(18) K m値と Vmax値を求めた。. d基質特異性 9種類の基質(コーヒー豆由来アラピノガラクタン、カラマツ由来アラピノ. ガラクタン、アラビアガム、カードラン、キシラン、マンナン、カルボ、キシメ. p H 4 . 6 ) チルセルロース、ペクチンおよびアミロース)を 20mM酢酸緩衝液 ( に 1% (w/v) となるように溶解し酵素活性を測定した。生豆由来アラビノガラ クタンを基質としたときの酵素活性を 100%として相対活性値で表した。. 1 2・ 1 2 . アラピノガラクタン分解試験 0.5% (w/v)アラピノガラクタンを溶解した 20mM酢酸緩衝液 ( p H 4 . 6 )70ml 0. にガラクタナーゼを終濃度 4mg添加後、 45Cで反応させた。反応液を 0 . 5、1、2、. 3、4、5、6、12、24、48および 72時間後に 1mlずつ取り出し、 SomogyiNe l s o n 法で還元糖を定量した。また取り出した反応液に 99.5%エタノールを 3倍量加 ( 1 →3 )ーD-アラピノガラクタン え、遠心分離(10000rpm、10min)して未反応の 0・ を沈殿させた。この上清に含まれる全糖量をフェノール硫酸法で、定量して、分解 率を求めた。. 1 2 1 3 .酵素反応生成物の薄層クロマトグラフィー試験 20mM酢酸緩衝液 ( p H 4 . 6 )に 1% (w/v)コーヒー豆由来アラピノガラクタンと ガラクタナーゼ (40mU/m I)を加え、 45Cで反応させた。一定時間ごとにサン 0. プリングし 2M酢酸を加えて反応を停止しさせた。反応液をシリカゲ、ル薄層フ。 レートにスポットし 1 p r o p a n o l:e t h a n o l:H201 7:1:2 ( v : v : v ) で展開後、 硫酸を噴霧、加熱してスポットを発色させた. 18. 5 2 ).
(19) 1 2・1 4 . オリゴ糖資化試験 ( 1 →3 ) D -アラビノガラク 酵素反応生成物の微生物資化試験を行った。まず 0・. タンをガラクタナーゼで処理して、得られた分解物を活性炭カラムクロマトで ( 1 →6 ) D -ガラクトピオースに単離した。 アラビノース、ガラクトースおよび 0・. 生 成 物 は TLC 分 析 で 同 定 し た 。 資 化 試 験 に は 腸 内 細 菌 4 属 7 種 10 株 ( B i f i d o b a c t e r i u m 属 3 種 7株 、. T ' a b l e2 .C u l t u r es t r a i n sf o rv i a b i l i t yt e s t . S t r a i n B i f i d o b a c t e r i u mb r e v eJCM1 1 9 2 T B i f i d o b a c t e r i u mb r e v eJCM1 2 7 3. L a c t o b a c i l l u s 属. 2 種. E s c h e r i c h i a c o l i 1 株、. 2 株、 S e r r a t i α. marcescens1株)を用いた ( T a b l e. B i f i d o b a c t e r i u mb r e v eJCM7 0 1 6. 2 )。. B i f i d o b a c t e r i u mb i f i d u mJCM1 2 5 5 T B i f i d o b a c t e r i u mb r e v eJCM1 2 0 9. B i f i d o b a c t e r i u m属の培地には. B i f i d o b a c t e r i u mb r e v eJCM1 2 5 4 B i f i d o b a c t e r i u mlongumJCM1 2 1 7 T. Caseinpeptone(DifcoL a b . )10.0g、. L a c t o b a むi l l u sp l a n t a r u mNBRC3 0 7 0. Yeaste x t r a c t( D i f c o )5 . 0 g、 Meat. B i f i d o b a c t e r i u mb i f i d u m NBRC1 4 2 5 2 E s c h e r i c h i ac o/ i NBRC3 0 7 0. e x t r a c t( D i f c o )5 . 0 g、K2HP043.0g、. S e r r a t i am a r c e s c e n sNBRC3 0 4 6. Tween801.0ml、精製水 1000ml、 0. pH6.8 を使用した。高圧滅菌 (121C、20分)後、漏過滅菌したアスコルビン c y s t e i nHClを最終濃度1.0%および 0.05%になるよう無菌的に添加した。 酸と L 1→ 6 )D -ガラク 炭素源(グルコース、アラピノース、ガラクトースおよび 0・ (. トビオース)も櫨過滅菌し、最終濃度 0.5% となるよう添加した。 0. 供試株はグルコースを炭素源とした同培地で前培養 (37C、24時間)し、1.0. X106c f u l t u b e となるように接種した。資化性の有無または強弱は培養液の pH の低下で判定した。つまり、 pH4.5未満のものを(+++)、 満のものを(++)、 ものを(土)、. pH5.0以上 5 . 5未満のものを(+)、. pH6.0以上のものを(-)で表示した。. 1 9. pH4.5以上 5 . 0未. pH5.5以上 6 . 0未満の.
(20) 1 3 .実験結果および考察. 1 3・1.コーヒー抽出残漬からアラピノガラクタンの抽出 コーヒー抽出残澄 100g(水分 7 %)を 1000μm 未満に破砕し 、蒸留水 1500ml に懸濁後 、1 2 1Cで 3時間加熱抽出した。得られた抽出液上清に 4倍量のエタノ 0. ールを加え、粗アラ ピノガラクタン. 2. 48g ( dryweight) を沈殿させた(収率. 2 . 7%)。組アラピノ ガラクタンから精製 アラピノガラクタン. g .4 . Ara b i n o g a l a c t a n仕 omc o f f e ebean. F i. 1 .08g を得た ( 収率. c r u d ea r a b i n o g a r a c t a n; B, p u r i f i e da r a b i n o g a r a c t a n . A,. %)( F i g .4 .A, B )。 1 .1. 1 3 2 .アラピノガラクタン ( A G ) 酸加水分解物の TLC分析 3種類のアラピノガラクタン(粗 AG, レーン 5 ;精製 AG, レーン 7 ;市販 カラマツ由来, レーン. 9 )を酸加水分解し 、得られた分解物を TLC分析 したと. ころ 、いずれのアラピノガラクタンからもアラピノースとガラク トースのみ検 出された。 この結果から 、コーヒー抽出残漬から単離した標品はアラピノガラ クタンであることを明らかにした ( Fig.5) 。. 2 0.
(21) 1, a r a b i n o s e. 2, g a l a c t o s e. 3,. a r a b i n o s e + g a l a c t o s e 4, h y d r o l y s a t e so f crude c o f f e e bean AG+. a r a b i n o s e +. y d r o l y s a t e so fcrudec o f f e e g a l a c t o s e;5,h y d r o l y s a t e so fc o f f e ebeanAG beanAG;6,h. 1. 2. 3. 4. 5. 6. 7 8. 9. F i g . 5 .TLCo fh y d r o l y s a t e so fc o f f e ebean. +a r a b i n o s e+g a l a c t o s e;7,h y d r o l y s a t e so f p u r i f i e dc o f f e ebeanAG;8,h y d r o l y s a t e so f. andl a r c hwooda r a b i n o g a l a c t a n(AG)by. l a r c hwoodAG;9,h y d r o l y s a t e so fl a r c h. t r i f l u o r o a c e t i ca c i dh y d r o l y s i s .. woodAG+a r a b i n o s e + g a l a c t o s e. 1 3 3 . コーヒー抽出残漬から抽出したアラピノガラクタンの平均分子量 精製アラピノガラクタンを SephadexG-200 カラムクロマトグラフィーに供. F i g .6 )。 ゲルろ過から求めた精製ア したところ、ブロードなピークを示した ( ラピノガラクタンの分子量(モード値)は 122, 000、分子量分布は 122, 0 0 0 ' " ' ' 7, 500 で、 あった ( F i g . 7 )。北畠はカラマツから抽出したTypeI Iアラピノガラクタンの 分子量 を測定し、シベリア産 1 8, 400、北海道産 18, 700、長野県産 2 1, 400と報 告している. 53)が、これらの値と比較するとコーヒー抽出残澄から抽出したアラ. ビノガラクタンの分子量は大きかった。 また分子量分布も 122, 0 0 0 ' " ' ' 7, 500で 、 カラマツ由来のアラピノガラクタンと比較するとぱらつきが大きかった。. 2 1.
(22) ト一一-1. ・. 10. ・,・. A r a b i n o 1cUn. 亘. 号10 S ~ 0. 8. 色. 。. 〈. 主. 0. 8. ・. 10 0. 2. 20. 40. 80. 80. 10 S. 100120. o. ・. E l u t i o nv o l u m( m) 1. 0. 2. 0. 4. ・. 0. 8. 0. 8. K v. F i g .7 .Molecularweighto f. F i g .6 . GPC-chromatography o f. arabinogalactanfromc o f f e e. arabinogalactanfromc o f f e ebean.. 1 3・4 .A .1imn伊白lsNo.232の培養 A. ルmigatusNo.232は 0.5%アラビノガラクタン を唯一の炭素源 とす る培地で. ) D -ガラク タナーゼ活性は菌糸 の増殖 ととも 良好に成育 し 、 培養液中の 0・u→ 3 に上昇 し 、 培養 7日後 に もっとも 高く なった。. 1 3・5 .酵素の精製 培養 ろ液 中のガラ クタナーゼは 4段階のステ ップで精製した ( T a b l e3 )。 Table3 .Summaryo fp u r i f i c a t i o no fgalactanasefromA .fumigatusNo.232. Volume(ml }. P r o t e i n Enzyme A c t i v i t y ( m,). u m i 主 壷 ]. c u l t u r ef i l t r a t e ammoniums u l p h a t e SP-SephadexG-50 SephadexG-100. 4500. 0 4 7 8 . 1 1 3 5 . 0 1 2 6 . 6. 7 2 5 . 1 7 0 . 0 2 2 . 1 9. 2. 1 3 5 . 0 3 4. 4 3 3 . 1 3 2 . 9. S p e c i f i c Y i e l d ( " ) A c t i v i t y ( u n i t s ! m,) 0. 1 9 0 . 4 9 1 .5 0 3 . 5 7. 1 0 0. 0 9 . 7 3 . 0 1 .3. F o l d. 1 .0. 2. 6 8. 1 1 9. 2. %s o l u t i o no ft h ea r a b i n o g a l a c t a n ( w / v ) f r o mc o f f e eb e a n Thes t a n d a r da s s a yr e a c t i o nc o n t a i n e da1. i n20mMa c e t a t eb u f f e r( p H4 . 6 ),a n dwasi n c u b a t e dwi出 t h ee 回 戸n ep r e p a r a t i o na t40C f o r 0. 3 0 r n i ni nat o t a lvolumeo fO. 4 m 1. Th er e a c t i o nr n i x t u r ewass t o p p e dbya d d i n gO. 2 r n lo fN e l s o n‘ s t . r e a g e n. 22.
(23) 硫安塩析で、得られた沈殿を透析後、 SP-SephadexG-50カラム ( 2 . 8X40cm) O' " ' ' O . 5 M ) グラジエン トをかけて溶出 したところ、 1本の に吸着させ、 NaCl ( ガラクタナーゼピークが得られた ( F i g . 8A)。 活性画分を濃縮し、 Sephadex G-100カラム(1.5X100cm) で、ゲ、ル癒過したところ 、 1本のガラクタナーゼピ ークが得 られた ( F i g.8B)。この精製酵素は SDS-PAGEで 、 単一バンドを示し ( F i g. 9 )、1 9 . 2倍精製され、回収率は 1 .3%であった。. J. ドー・4. R. 0 .1 『. 回. 1 . 0. 円 芯.、. . .. ノ/. ~05. o. ー. o. 0 .5. D) 4 0 0 必 伺 叫m前吋). 0 . 8 0 .5 0 . 4. 0 . 2 0 .1. o. a. ー← 島町市副~II枇/同). 4. トデ圃. 1 2 : 3 z i. Jγ. 5ω 0 . 0. 2 .5. : : l B 。m. o. 4 0. ~ぅ ω ω. ∞. 1. - -岳宮卯晴 嵐対頃削 ぽ枇/川). F i g. 8 .P u r i f i c a t i o no fg a l a c t a n a s e仕omA伊 e t 宮i l l u sルm i g a t u sN o . 2 3 2 . A: S P S e p h a d e xG・50. B: S e p h a d e xG-100. 2 3. a〈. 。. 1 2 01 4 0. 臥必∞ 叫UTば 岬. 一← k I I r J. ! ?. ー← k I I r J.
(24) 1 3 6 . ガラクタナーゼの分子量. A. B (kD a ). ー -2∞ ・ ー116. 分子量マーカーと 一緒に SDS-PAGE した精製酵素の分子量は 86, 000Daと推 定された ( F i g . 9)。 一方 、 Sephadex G-100 ゲ、 ル滅過カラムクロマトグラフ. .圃圃・. -. 6 6 . 2. ・ ー. 2 1 . 5. ・ ー. 1 4 . 4. ・ ・ ー 4315. G a l a c t a n a s e ( 8 6 kD a ). ィーで求めた n a t i v e酵素の分子量は 80, 000Da で あ っ た こ と か ら 、 A .. ルmigatusNO.232が生産したガラクタナ. ・ ー 6.5. ーゼは分子量 80, 0 0 0 " ' ' 8 6, 000のモノマ ータンパク質であることを明らかにした。 No.232株が生産するガラクタナーゼ 000Da)は 、 I r p e xl a c t e u s の分子量 ( 86, の ガ ラ ク タ ナ ー ゼ (51, 000Da) や A s p e r g i l l u sn i g e r( 66, 000Da)が生産する. 酵素と比較して大きかった。 しかし A. F i g .9 . SDS-PAGEo fp u r i f i e dg a l a c t a n a s e. f u m i g a t u sN O . 2 3 2 . f r o mA. LaneA, p u r i f i e dg a l a c t a n a s e( 2μg . ) L a n eB,M o l e c u l a rw e i g h tm a r k e r so fp r o t e i n s t a n d a r d s .. 0 0 0 " ' ' f u m i g a t u s が生産する酵素は、 65, 000Daと報告されており 130,. 5 4 ・6 0) 、本酵. 素の分子量も既報の結果と類似していた。. 1 3 7 .精製酵素の最適 pHと温度および安定性 本酵素の最適 pHは 4 .6で 、 pH 8 . 0以上もしくは pH2 . 0以下では 10%以下 の活性しか示さなかった。また pH4.0で最も安定であり、 pH8.0以上では活性 l 由来のガラク s p e r g i l l u sn i g e r 4s を示さなかった。本酵素の最適反応 pH4 . 6は A. r p e xl a c t e u s52) ( pH4 . 6 )、Rhizopusn i v e u s47) (pH5 . 0 )と タナーゼ ( pH4. 6)や I 0. 同じように酸'性域にあった。既報のガラクタナーゼは pH4 . 0, 4Cで 24時間保. 24.
(25) 存したときも安定で、あったが、本酵素の pH安定域は p H 3 . 5 ' " ' ' 5 . 0の範囲で狭か った。 、 50C以下で 30min保持しても失 本酵素の最適温度は pH4.6のとき 45Cで 0. 0. F i g . 1 0 )。既 活しなかった。しかし、 70Cで 30min保っと 10%以下に失活した ( 0. 報のガラクタナーゼは 50C以下でも失活しなかったが、アッセイ系のインキュ 0. 1 0分)ため、単純に比較できない。本酵素は 50Cで 30分 ベート時聞が短い ( 0. インキュベートしても、活性を 100%保持していたことから、既報の酵素より熱 安定性は高いと思われる。 1 0 0. B. 3 8 0 さ. 。 わ 20. o. 35. 4 5. 5 5 6 5 7 5 T e m p e r a t u r e e c ). 85. F i g .1 0 .EffectofpHandtemperatureonstabilityandactivityofgalactanase fromA . f u m i g a t u sNo.232. . tA,E f f e c to fpHona c t i v i t y( ・: g l y c i n e H C I Ther e a c t i o nc o n d i t i o n sa r ed e s c r i b e di nt h et e x. b u f f e r ,企 : a c e t a t eb u f f e r ,. : p h o s p h a t eb u f f e r)ands t a b i l i t y ( O : g l y c i n e H C Ib u f f e r ,. . d .: a c e t a t e. , 口: p h o s p h a t eb u f f e r ) .B, E f f e c to f t e m p e r a t u r eona c t i v i t y(.)加ds t a b i l i t y ( O ) . b u f f e r. 1 3・8 .金属塩が酵素活性に与える効果 2 1mMMn2 +と Cu + は精製ガ、ラクタナーゼ活性を 60.2%と 72.4%に低下させ、 Hg2+は酵素を完全に失活させた ( F i g .1 1 ) EDTAは活性を全く阻害しなかった 0. ことから、本酵素は活性の発現に金属を要求しないと考えられる。. 2 5.
(26) 1 2 0. i i t1 0 0. . . h. ま. 8 0. ~. 8 0. •・ • 2. ミ 40 a : 20. o . " c ト. ザゲペや. 仇 r i ' ' < J ザ. ず. . ! < . , i i -. ・ ず ザ ザ ザ ザ 。 ; ' > " +(:0ザ. ψ dJ d e t b V ψ . . 0-. ー. . . ~.. E 企. F i g .1 1 .E f f e c to fv a r i o u scompoundsong a l a c t a n a s ea c t i v i t y . Thee 回 y mewasp r e i n c u b a t e dw i t hl m Mv a r i o u sc h e m i c a l sa t4 5" Cf o rI hi n2 0 mM a c e t a t eb uf I e r . R e s i d u a la c t i v i t i e sw e r et h e na s s a y e du n d e rt h es t a n d a r dc o n d i t i o n s .. 1 3・9 .酵素の動力学的特徴 基質にコーヒー豆とカラマツ由来のアラピノガラクタンを用いて、基質濃度 が酵素活性に与える効果を検討した。本酵素の生豆由来アラピノガラクタンに. 4 .74mg/mD 対する Km値 O.89mg/mlで、カラマツ由来アラピノガラクタン ( に比べて小さかったことから、本酵素はコーヒ一生豆由来アラピノガラクタン. F i g .1 2) 。本酵素は s( 1→ 3 ) D -ガラクトシル結合 に高い親和性をもっていた ( をもっ主鎖を加水分解して、アラピノガラクタン側鎖は分解しない。一方、カ. ・. Lr c hwood. F i g . 1 2 . Lineweaver'Burkp l o t so f 1 .5. g a l a c t a n a s e仕omA. f u m i g a t u sNo.232. . ,. a g a i n s ta r a b i n o g a l a c t a n .. 〉. ¥、. ・・,・. . rb i n o Ic t . n. ,, . x. Km: 0. 89m / m l ' / ' 1o l /mi n / " , 3. 1 8μ1. -2. 2. 3. 4. -0. 5 l/S. 26. c o f f e eb e a n ;企 , larchwood..
(27) ラマツ由来アラピノガラクタンは、生豆由来アラビノガラクタンよりも 0・ ( 1 →6 ) D -ガラクトシル結合をもっ長い分岐側鎖が結合している. 4 7 .6 1 ) ので、本酵. 素が作用しにくいと考えた。本酵素の Km値 は 既 報 の A .f u m i g a t u s生産ガラ クタナーゼと比べて低く、アラビノガラクタンに対して高い親和力をもつこと が示唆された。 九四値は生豆由来アラピノガラクタン (3.56μM/minlmg) よりカラマツ由来 アラピノガラクタン ( 6 . 5 4 μ Ml minlmg) が高かった。. 1 3・ 1 0 . ガラクタナーゼの基質特異性 本酵素はアラピノガラクタン(コーヒー豆およびカラマツ由来)以外にアラ ビアガム、 0・ ( 1 →3 ) D -グソレカン結合をもっカードランからも還元糖を遊離した. ( F i g . 1 3 )。キシランやマンナン、ベクチンからも分解物が若干確認された。分 解物の糖組成を分析したところ、いずれもガラクトースが検出されたことから 基質に混在していたガラクタンが分解されたと考えられる。本酵素は 0・ ( 1 →4 )・ グルコシド結合や α・ u→4 )・グルコシド結合をもっカルボ、キシメチルセルロー スやアミロースには作用しなかった。. 27.
(28) 120 ;100. 主 g. 40 20. S u b s t r a t e. U n k a g e. A Ara bi n o g a la c t a n ( f ヤ omc o ' 仔e eb e a n ). s-(l→ 3 ) -GaI βー { 1→6 ) -GaI αー { 1→ 3 ) A r a. B A r a b i n o 伊l a c t a n ( 作oml a r c h附 o d ). αー { 1→ 3 ) A r a. C C a r d r a n. βー { 1→3 } ー G l c. D Gum a r a b i c. βー { 1→3 ) -GaI βー { 1→ 6 ) ーGal αー { 1→3 ) A r a. E P e c t i n. βー { 1→ 4 } ーGal. F X y l a n. s-(l→4 ) X y. n a n G Man. s-(l→4 ) -Ma n. s-(l→ 3 } ーGal s-(l→6 ) -GaI. H C a r b o x y m e t y lc e l l u l o s eβー{ 1 + 4 ) G l c. o. αー{ 1→ 4 } ーG l c. Am y l o s e. A B C. 0 E F G. H. F i g .1 3 . Substrates p e c i f i c i t yo fgalactanaseonthevarioussubstrates. Reactionmixtures( 1.0ml)containedvariouspolysaccharides( 1% , w/v)andthe. l )in20m Macetatebuffer(pH4. 6 ) . enzyme(O.04units/m. 1 3・ 1 1 . s・ ( 1→3 )・ D・アラピノガラクタン分解試験. O . 5% ( w / v )アラピノガラクタンを基質として本酵素を反応させたとき、反応に 伴って還元糖が遊離し、反応後 24時間で、ほぼ定常状態になった。このとき遊離. 1 . 6. . . 。 c e a. 1 0 0. 1 . 4 80. 1 .2. , U 旬 ,R 、 。 . 、 ,,. z. o. . ‘. o, 画 、 』. 60 ・ 4 2 -. ・ ー. 塁 王. 0 . 8. ~",:;. 0. 6. 国. E. ;! ! I. 40 唱 ー 包 @. 0 . 4. 20. @. a : 0 . 2. o. o. c o f f e ebeanbygalactanase. o n s i s t i n gof70mlofa R e a c t i o nm i x t u r e,c. 。 O.5%(w/v)arabinogalactansolutionin20rnM. o. 昌. F i g .1 4 .Hydrolysisofarabinogalactanfrom. o 20. ,. 40. 60. 80. I n c u b a t i o nt i m e ( h r ). • R e d u c i n supr( m g ! ml ). ・D e c r a d a t i o nr a t i o( , , ). 28. a c e t a t ebuf I e r ( p H 4 . 6 )andg a l a c t a n a s e ( 4 m g o. p r o t e i n ), werei n c u b a t e da t4SC ..
(29) した還元糖は1.34mg/mlであった。遊離した全糖量(エタノールで、 沈殿しなか った糖類)を定量し分解率 を求めたところ、反応 24 時間後で 63.0%に達した ( F i g . 1 4 )o Hashimoto は. 47 )コーヒー豆由来のアラピノガラクタンを. Rhizopus. nlveusの生産する酵素で分解しているが、本酵素活性は Hashimotoらの 7.1倍 で、 あった。 1 3・1 2 . アラピノガラクタン酵素分解物の TLC分析. コーヒー豆由来のアラピノガラクタンに精製酵素を加えてインキュベートし、 経時的にサンプリングした反応液を薄層クロマトグラフィーで分析した結果、 反応初期の段階からアラピノース、ガラクトースおよびオリゴ糖とほぼ同じ位 . 3 6 ) が出現した ( F i g .1 5 )0 Rf0.36のス 置にスポット (Rf0.67,Rf0.62,Rf0. ・・. a・ 4・・~ ~・ a・ a a a・ J~・. o0.25 0.5. 1 2 6 1 2 24 R e a c t i o nt i m e(hr) M1 M2 M3. F i g .1 5 . TLCo fh y d r o l y s i sp r o d u c t sfrom a r a b i n o g a l a c t a nby g a l a c t a n a s e . Ar e a c t i o n m i x t u r e( 1. 0ml )c o n t a i n i n ga r a b i n o g a l a c t a n仕omc o f f e ebean(l%, w / v )andt h e enzyme 0 ( O. 04U/m l )i n20m Ma c e t a t eb u f f e r(pH4 . 6 )wasi n c u b a t e da t40 C. Ati n t e r v a l s,t h e r e a c t i o nwass t o p p e dbyb o i l i n gande a c hsample( 15 μ1 )wast h e ns p o t t e donaTLCp l a t e .. s( 1→ 6 ) -D G a l a c t o b i o s e( r e a g e n t ) . Markers( M ): Ml, Ara b i n o s e; M2, G a l a c t o s e ;M3,. 29.
(30) ポットをかき取り抽出後、活性炭カラムクロマトグラフィーで精製した。得ら. 0 0C、2時間加熱)、もし れた物質を酸分解(1M トリプルオロ酢酸によって 1 0. .c o l i由来 sガラクトシダーゼで加水分解して、生成物がガラクトース 2 くは E. 量体であることを確認した。またこのスポットは市販 s-( 1→ 6 ) D -ガラクトピオ ースと同じ Rf値を示したことから、 Rf0 . 3 6のスポットをガラクトピースと同 ( 1 →6 ) D -ガラクトピオースと微量のガ 定した。また、酵素反応開始初期から 3・. R f0 . 1 5 )と思われるオリゴ糖を蓄積したことから、コーヒー ラクトトリオース ( 豆由来のアラビノガラクタン側鎖の重合度は 2 ' " ' " ' 3個と推察される 。従って本酵 素は α・ ( 1 →3 )・あるいは s -( 1→ 3 ) -結合を選択的に切断してアラピノース、ガラク ( 1 →6 )・結合をもっオリゴ糖が残る形式 トースを遊離させ、分解されなかった 0・ で分解されると考えている ( F i g .1 6 ) 。この分解形式は、 R .n i v e u sが生産するガラ クタナーゼの分解様式と大きく異なっており. 4 7 )、むしろ. 5 A .n i g e r 4 やよ L a c t e u s52) ). が生産するガラクタナーゼの分解形式に酷似していた。. F i g . 1 6 . Diagrammatic representation o fd e g r a d a t i o n o fc o f f e e bean a r a b i n o g a l a c t a n by g a l a c t a n a s e . Ar rows i n d i c a t e t h a t t h e a c t i o n o f g a l a c t a n a s e .. 30.
(31) 1 3・1 3 .腸内細菌によるオリゴ糖の資化性 コーヒー抽出残透から調製したTy peIIアラビノガラクタンを本酵素で分解し、 得られた 8・(1→ 6 )D -ガラクトピオースとグルコース、アラピノース、ガラク トースの腸内細菌による資化性を、. PYF液体培地を用いた嫌気培養 ( 3 7Cで 9 6 0. 時間)した培養液の pH変化から判定した。 0・ ( 1→6 )D -ガラクトビオースは. B i f i d o b a c t e r i u mbreve、B i f i d o b a c t e r i u mb i 沼dum 、B i f i d o b a c t e r i u mlongum、 L a c t o b a c i l l u splantarumによって資化され酸を生成した ( T a b l e4 ) ことから、 腸内の善玉菌の生育を促進し酸を生産させることにより悪玉菌の増殖を抑制す るプロバイオティックな効果が期待される。 f f e c to fg l u c o s e,a r a b i n o s e,g a l a c t o s eand s・ ( 1 →6 )・ D g a l a c t o b i o s eon T a b l e4 . E t h ev i a b i l i t i e so fv a r i o u smicroorganisms S t r a i n B i f i d o b a c t e r i u mb r eνeJCM1 1 9 2 T B i f i d o b a c t e r i u mb r e v eJCM1 2 7 3 B i f i d o b a c t e r i u mb r e v eJCM7 0 1 6 B i f i d o b a c t e r i u mb i f i d u mJCM1 2 5 5 T B i f i d o b a c t e r i u mb r e v eJCM1 2 0 9 B i f i d o b a c t e r i u mb r e v eJCM1 2 5 4 B i f i d o b a c t e r i u ml o n g u mJCM1 2 17T L a c t o b a c i l l u sp l a n t a r u mNBRC3 0 7 0 B i f i d o b a c t e r i u mb i f i d u mNBRC1 4 2 5 2 E s c h e r i c h i ac o l iNBRC3 0 7 0 S e r r a t i am a r c e s c e n sNBRC3 0 4 6. G l u c o s e G a l a c t o s eA r a b i n o s e. +++ ++ ++ ++ ++ ++ +++ ++ +++ + +. ++ ++ ++ + ++ + ++ + ++ + ++ + +++ +++ ++ +++ +++ + +. s-(1→ 6 )ーD G a l a c t o b i o s e. +++ ++ ++ ++ +++ +++ +++ +++ +++. 1 4 . まとめ A .ルm i g a t u sNo.232の培養液から s-U→ 3 ) D -ガラクタナーゼ (EC3 . 2 .1 .9 0 ). をイオン交換クロマトグラフィー (SP-Sephadex G -50)、ゲ、ル櫨過クロマトグ ラフィー (SephadexG-I00)によって、電気泳動的に均一になるまで精製した。 0. 精製酵素の分子量は 86kDaで、最適 pHは 4 . 0、温度は 45Cであった。本酵素 は pH4.0、45C以下で最も安定で、 Hg2+イオンにより完全に失活した。 0. 3 1.
(32) ガラクタナーゼの M i c h a e l i s定数 (Km) は、コーヒー豆由来アラピノガラク タンを基質にしたとき O.89mg/mlで、最大反応速度 ( l t う l 1a x )は 3 .16units/mgで あった。本酵素は、TypeI Iアラビノガラクタンのような s-( 1 →3 )結合や α ー(1 →3 )結合を加水分解し、酵素反応の初期段階からアラピノース、ガラクトース、. xo型分解様式をもっ酵素と思われる。 ガラクトビオースに分解したことから、 e ヒト腸内フローラ構成菌種に対するこれらの糖の資化性を調べた。. 0・(1→. 6 ) D -ガ ラ ク ト ピ オ ー ス は B i f i d o b a c t e r i u mb r e v e 、B i f i d o b a c t e r i u mb i f i d u m、 B i f i d o b a c t e r i u mlongum、L a c t o b a ci l / u sp l a n t a r u mに資化され多量の酸を生成して培 養液の pHを低下させた。一方腸内有害菌である E s c h e r i c h i ac o/ iおよび S e r r a t i a. ma 陀 白c e n sにも資化されたが、酸は生成されなかった。これらの結果からコーヒ 一生豆から抽出したアラピノガラクタンを酵素分解して得たオリゴ糖には、ヒ ト腸内フローラの改善効果をもつことを明らかにすることができ、プロバイオ ティクスとしての応用が期待される。. 32.
(33) 第二章. コーヒー生豆に含まれるクロロゲン酸類の分離とその機能性評価. 2 -1.緒言. コーヒ一生豆の品質は、栽培種、栽培地、収穫期、精製方法(水洗式、乾式)、 輸送、保存など様々な要因で決定されるが、輸入生豆の品質は、成熟するまで に摘果したり、強風などで落下してしまった未熟果の混入割合によって大きく 左右される。等級の低い生豆ロットには欠点豆の混入率が高く、とくに未熟果 の混入割合がコーヒーの等級を決定する最も大きな要因となっている。 含まれている。現在商業的に栽培 コーヒ一生豆にはクロロゲン酸類が 4"-'10%. C o f f e aa r a b i c a )とロブスタ種 ( C o f f e a されているコーヒーの多くはアラピカ種 ( c a n e p h o l av e r .r o b u s t a )で、ロブスタ種の生豆には 80"-'100mg/gのクロロゲン酸 類が含まれており、アラピカ種と比較してその含有量は有意に高いことが明ら かにされている. 6 2 )。このクロロゲン酸類に注目して、機能性食品素材として積. 極的に利用していくためには、まずコーヒ一生豆中のクロロゲン酸含有量を、 果実の成熟度、産地、銘柄ごとに正確に把握しておく必要がある。そこでまず コーヒー果実の成熟度と生豆中のクロロゲン酸類量の関連性を調べた。. DeMenezes63)は、コーヒー豆の成熟に従って全クロロゲン酸類 (CQA、FQA、 diCQA) に占めるクロロゲン酸 (CQA) の割合が高くなると報告している。つ diCQA) の含有比率が高く、これが まり未成熟果ではジカフェオイルキナ酸 ( コーヒーの品質とくに味覚バランスに悪い影響を与えるといわれている。これ ら等級の低いコーヒー豆は市場に出まわらないで廃棄されることが多い。そこ で我々は、これら低級豆に含まれるクロロゲン酸類を機能性食品素材として活 用する方法を検討した。 クロロゲン酸類は、顕著な抗酸化活性 64,65)、肝保護活性 66・ 側 、 HIVインテグ ラーゼ阻害 69-72)、マクロファージの活性化. 3 3. 7 3 )など様々な生理機能を有している。.
(34) また 5-CQAにはリノレン酸の脂質過酸化抑制効果. 64). が 、 1 , 3-diCQAには γ線. 照射によって引き起こされる 5-CQAの酸化抑制効果が報告されている. 65) 。. やフェルラ酸 (FQA)、diCQA 異 しかし、コーヒ一生豆に含まれている CQA 性体のもつ様々な機能性を同 一条件下で比較した研究は著者の知る限りまだな 、FQA 、diCQA 異性体を単離し、それらの い。 そこで、コーヒ一生豆から CQA 機能性を lnげtro, 同一条件下で比較した。 さらにクロロゲン酸類を機能性食品 素材として応用していくにあたって、消費者に馴染みゃすい缶コーヒーへの応 用を考え、缶コーヒーに添加したクロロゲ、 ン酸類の製造工程中で、 の変動につい て分析した。. 22.実験材料および実験方法 2・2 -1 .試料 鹿児島県内で栽培されているアラピカ種果実を成熟度の異なる時期に採取し、 測色色差計 (CR-300、MINOLTA株式会社)を用いて色差 (Lab値)を測定. F i g .1 7,1 8 )。 した。そして色調の違いに基づいて採取果実を 7段階に分類した (. . . E. I. 司. F. ..ll~寸1υ什 lìキ斗.J 千十↓↓い. 寸皿. . . . . . . ー. L. I V. V. V I. . . . . . .. . .. /"弘、:::1><、三〉く、主オ1'"1 0. V s. F i g .1 8.Colortoneo fc o f f e ec h e r r y. F i g.1 7 .Coffeecherrysamples.. 34.
(35) 2 2・2 . コーヒー果実. Co 符' e ec h e r r i e s( G r o u p1. . . , 四 ). から生豆の単離 まず果肉とパーチ メント(種殻)付き のコーヒー豆に分離 M u c i l a g e. した後、得られたパ ーチメントコーヒー を天 Hで 1 日乾燥し. P a r c h m e n t. てから、パーチメン トとシルバースキン. F i g .1 9 .Preparationf o rthesamples(wetp r o c e s s i n gmethod).. (種皮)を除去して 生豆を得た ( F i g .1 9 )。. 2 2・3 .官能試験 テストロースター(プロバット社、 BRZ-4) を用いて 7試験区から得た生豆 をミディアムローストになるように賠煎した後、コーヒーミル (BONMAC、. BM-570M) で細挽きに粉砕した。官能試験には、粉砕した賠煎豆 10gに熱水 150ml を注ぎ、ペーパーフィルターを用いて抽出したコーヒーを用いた。官能試 験はパネラー 10名で、行った。カテゴリーは酸味、苦味、渋味、濃厚感、後味の. 5っとし、尺度は -3'""+3の 7段階に分けて絶対評価した。また異味、異臭の 評価とその強度は 0 ' " "+6の 7段階で評価した。. 2 2・4 . クロロゲン酸類の分離定量 生豆は液体窒素で凍結後、粉砕機(分析粉砕機 R-8、日本理化学機械株式会. 35.
(36) 社)を用いて1.0mm のふるいを通過するサイズに粉砕し試料とした。試料は 70%( w / v )メタノールで 20分間還流抽出をおこない、 3回抽出を繰り返した後、. 定容した。限外ろ過した試料中のクロロゲン酸類(異性体組成)は、 HPLCで 定量した. 。定量は市販クロロゲン酸 (5-CQA) を用いて C l i f f o r dらの吸光度. 74). 比に基づいておこなった. 7 5 )。この方法で定量した. 5-CQAの添加回収率は 9 4 . 9. ' " ' '1 0 3 .9%(n=5)で、あった。. 2-2・5 . 市場コーヒ一生豆中のクロロゲン酸異性体の組成. 供試試料には 17産地、 3 1銘柄のコーヒーを用いた ( T a b l e5 )。. T a b l e5 .An a l y t i c a lsampleso fc o f f e eb e a n .. c ' o f f e aa r a b i c a BRAZILSANTOSN o . 2 #1 8 4 /5 #14/16 BRAZILSANTOSNo. COLOMBIA EXCELSO COLOMBIA MARAGO COLOMBIA SUPREMO CUBA CRYST AL MOUNTAIN DOMINICA EL SALVADOR ETHIOPIA MOCHADJIMMAH ETHIOPIAMOCHASIDAMO .. p GUATEMALA E GUATEMALASHB HAWAII KONA EXTRA FANCY HAWAII KONA N o . 1 INDIA JAMAICA BLUE MOUNT AIN N o . 1 JAMAICA PEABERRY JAMAICA PRIMEWASHED KENYA AA .. p MEXICO E NICARAGUA PERU CHANCHAMAYO TANZANIA KIBO. C o f f e ac a n 伊h o l av a r .r o b u s t a INDONESIA WIB INDONESIAAP-1 INDONESIA EK-1 INDONESIAMANDHEUNG INDONESIA TORAJA IVORY COAST UGANDA VIETNAM. . w. . w. 36.
(37) 2 2 6 . コーヒ一生豆からクロロゲン酸類の単離. 等級の異なる生豆からクロロゲン酸類を単離した。供試生豆には 2002年に収 穫 さ れ た イ ン ド ネ シ ア 産 ロ ブ ス タ 種 を 使 用 し 、 等 級 の 低 い 生 豆 に は EK-1 grade4(ジャワ島産)を、等級の高い生豆には WIB (スマトラ島産)を用いた。. カフェー酸、キナ酸、 3, 4-dihydroxyhydrocinnamicacid ( 3, 4-diOHHCA) は SIGMAAldrich (St.Louis, MO)、 SephadexLH2 0は AmerciamB i o s c i e n c e s AB (Uppsala,Sweden) から購入した。他の分析用試薬はナカライテスク(京. 都)から購入した。. 2 2 7 . クロロゲン酸類の単離方法. コーヒ一生豆中のクロロゲン酸類は Uritaniらの報告を参考に単離した. 7 6 )。. 2種類の供試生豆 100gを粉砕し、 70%( v / v ) メタノール水溶液 1Lを加えて 800Cで 20分間、 3回抽出を繰り返した。抽出液は 50ml まで減圧濃縮後、 40C. に 48時間保ってクロロゲン酸類とカリウムイオン、カフェインが等モルずつ会 合した黄褐色の沈殿を得た. 76,7 7 )。この沈殿物に飽和酒石酸溶液を加えて生成し. た酒石酸カリウムの白色沈殿を除去した。つぎにクロロホルムを添加してクロ 2 0 カラム ロホルム層に移行したカフェインを除去した。水層を SephadexLH(450X26m mi . d . )に供し、 0.2%( v / v ) 酢酸溶液にメタノール濃度勾配 ( 0・70%). をかけて展開した (60ml/h)。溶出液は 325nmの吸光度を測定した。得られた ピークを回収、濃縮した後、分取 HPLCPLC-561system (GLSciencesI n c ., ) で目的のピークを分取した。分離条件を Table6に示した。. T a b l e6 .P r e p a r a t i v ec o n d i t i o no fc h l o r o g e n i ca c i d s c o l u m n c o l u m ntemp m o b i l ep h a s e g r a d i e n t d e t e c t i o n. I n e r t o s i lODS-32 5 0x1 9mmi . d .(GLS c i e n c e sI n c . ) 4 00C s o l v e n t sA( 2 0 %CH30Hi n0 . 2 %a c e t i ca c i d )a n d8( m e t h a n ol ) 0 . 0m i n,A=100%,6 0 . 0m i n,A :8 = 1 : 1,7 0 . 0m i n,8=100, %8 0 . 0m i n,A=100% UV3 2 6n m .. ※A I Is a m p l e sw e r ee l u t e da t1 5ml /m i n .. 3 7.
(38) 各ピークフラクションは再び SephadexLH2 0 カラムクロマトグラフィーを おこなって精製した。精製クロロゲ、ン酸類は凍結乾燥を行ってから -20oCで保 存した。. 2 2・ 8 . 単離したクロロゲン酸類の構造解析 高 速 原 子 衝 突 イ オ ン 化 質 量 分 析 (FAB-MS) には JEOLTandem-MStation. JMS-700TKMmasss p e c t r o m e t e r (JEOLL t d .,)を用いた。. uvスペクトルは. U3310 s p e c t r o m e t e r (日立製作所)、 lHNMRスペクトルは JEOLGSX-500 (500MHz) (JEOLL t d .,)を用いて測定した。 2 2 9 .1 , I D i p h e n y l 2 p i c r y l h y d r a z y l (DPPH) ラジカル捕捉活性 DPPHラジカル捕捉活性は、 Yoshikawaら 78)の方法に従って測定した。 O.lM 酢酸緩衝液 (pH5 . 5 )2.0mlに 500m l / Lエタノーノレ・ O.lM酢酸緩衝液(pH5 . 5 )に溶. .0mlと 0.2mMDPPH (ナカライテスク)-エタノー 解したクロロゲン酸類溶液 2 ル溶液1.0mlを添加して、室温、暗黒条件下で正確に 30分保った後、その溶液 の吸光度 (517nm) を測定した。クロロゲン酸類の濃度を変えて吸光度を測定 し、横軸にクロロゲン酸類濃度、縦軸に吸光度をプロットしてグラフを描き、 近似曲線から DPPHラジカルの吸光度(クロロゲン酸類無添加のコントローノレ). e d u c t i o n ) を算出し を 50%減少させるのに必要なクロロゲン酸類濃度 (50%r た。比較対照にはキナ酸、アスコルビン酸、. α ・トコブエロール、. 3, 4-dihydroxyhydrocinnamica c i d( 3, 4-diOHHCA) を使用した。. 2・ 2・ 1 0 . スーパーオキシドアニオン消去活性 anthine-Xanthineo x i d a s e (XOD) スーパーオキシドアニオン消去活性は X 系を用いてスーパーオキシドフリーラジカルを産生させる Matushigeら 79)の方. 0 . 2 )700μlに 3mMキサンチン 50 法に従って測定した。 0.05MNa2C03(pH1. 38.
(39) μl、 3mMEDTA50μl、1.5mg/ml午血清アルブミン 50μl、0.75mM ニトロ ブルーテトラゾリウム (NBT) 50μl とクロロゲン酸類溶液 50μlを添加して. 0分間保った。この溶液に 1 u n i t / m lXOD (ナカライテスク)を 50μl 室温で 1 添加し 25Cで 20分間保ってから、 6mMCuChを 50μl添加して反応を停止さ 0. せ、吸光度 (560nm) を測定した。クロロゲン酸類の濃度を変えて吸光度を測 定し、横軸にクロロゲン酸類濃度、縦軸に吸光度をフ。ロットしてグラフを描き、 近似曲線からコントロール(クロロゲン酸類無添加)の吸光度を 50%減少させ. 5 0%red u c t i o n ) を算出した。比較対照には るのに必要なクロロゲン酸類濃度 ( 4-dihydroxyhydrocinnamica c i d( 3, 4-diOHHCA) を用いた。 カフェー酸、 3,. 2 2・11.チロシナーゼ阻害活性 -チロ クロロゲン酸類のチロシナーゼ阻害活性は Tada80}らの方法をもとに L シンと L-DOPAを基質に用いて行った。. a )L ーチロシンを基質としたチロシナーゼ阻害活性. 0.55mML -チロシンと 10mM リン酸緩衝液 (pH6 . 8 )、マッシュルームチロ 0 u n i t s、クロロゲン酸類を含む反応液 2mlを 37Cで 1 0分間反応さ シナーゼ 6 0. せた後、 475nmの吸光度を測定した。チロシナーゼ阻害活性はコントロールに 対する比をチロシナーゼ阻害活性とし、次式によりもとめた。. A-(B-C)]/AX100 チロシナーゼ阻害活性(%)=[. A=供試試料を添加していない反応液の吸光度(コントロ ール). Bニ供試試料を添加した反応液の吸光度. C=サンフ。ルの吸光度 クロロゲン酸類の濃度を変えてチロシナーゼ阻害活性をもとめ、横軸にクロ. 39.
(40) ロゲン酸類濃度を、縦軸にチロシナーゼ阻害率をプロットしてグラフを描き、 回帰式からチロシナーゼ活性を 50%阻止するのに必要なクロロゲン酸類濃度を. e d u c t i o n )。比較対照にはアルブずチン、アスコルビン酸、カフェ 算出した (50%r ー酸を用いた。. b )L-DOPAを基質としたチロシナーゼ阻害活性 0.85mML-DOPAと 19mM リン酸緩衝液 (pH6 . 8 )、マッシュルームチロシ 0 u n i t s、クロロゲン酸類を含む反応液 2mlを 25Cで反応させ、 15秒間 ナーゼ 6 0. 隔で 3分間生成したドーパキノンの吸光度 (475nm) を測定した。この値を横 軸に測定時間を縦軸にフ。ロットして得られた回帰式を求め、この式から1.75分 と2 . 7 5分の 475nmの吸光度を算出した。チロシナーゼ阻害活性はコントロー ルに対する比をチロシナーゼ阻害活性とし、次式よりもとめた。 チロシナーゼ阻害活性 (%)=(A-B)/AX100 A =供試試料を添加していない反応液(コントローノレ)の1.75分と 2 . 7 5分. の吸光度の差. B=供試試料を添加した反応液の1.75分と 2 . 7 5分の吸光度の差 クロロゲン酸類の濃度を変えてチロシナーゼ阻害活性を求め、横軸にクロロ ゲン酸類濃度を、縦軸にチロシナーゼ、阻害活性をプロットしてグラフを描き、 回帰式からチロシナーゼ活性を 50%阻止するのに必要なクロロゲン酸類濃度を 算出した。 比較対照にはアルブチン、アスコルビン酸、カフェー酸を用いた。. 2 2 ・1 2 . ヒアルロニダーゼ阻害活性 ヒアルロニダーゼ阻害活性は、 Morgan Elson法を応用した前田ら を参考に測定した。. 40. 81)の方法.
(41) 100mM酢酸緩衝液 (pH4 . 0 ) に溶解したクロロゲン酸類溶液 200μ1にヒア 0 u n i t sを添加し、 37Cで 20分間予備加温した。同緩衝液に溶解 ルロニダーゼ、 4 0. した 0 . 0 1%(w/v)compound48/80 ( ナカライテスク)を 200μl加えて、 37Cで 0. 20分間インキュベートしてヒアルロニダーゼ、を活性化させた。この溶液に最終 4mg/mlとなるようヒアルロン酸を加え、 37Cで 20分反応させた後、 O. 4N 濃度 O. 0. NaOH を 200μl加えて氷冷し反応を停止させた。 0.5NNaOH に溶解した 0. 4.95%(w/v) ホウ酸溶液を 200μl反応液に加え、 1 00Cで 3分間加温した後、再. Dimethylaminobenzaldehyde試薬を 6ml加え、 37C び氷冷した。この溶液に p0. で 20分間反応させた後、 585nmの吸光度を測定した。ヒアルロニダーゼ阻害 活性は次式によりもとめた。 ヒアルロニダーゼ阻害活性(%)=[ ( A B ) ( C D ) ] / (A B )X 100. A=供試試料を添加していない反応液の吸光度(コントロ ール). B=酵素を添加していない反応液の吸光度 ( B l a n k ) C=試料溶液の吸光度 D =供試試料、酵素を添加していない反応液の吸光度(試料 Blank) クロロゲン酸類の濃度を変えてヒアルロニダーゼ阻害活性をもとめ、横軸に クロロゲン酸類濃度を、縦軸にヒアルロニダーゼ、阻害活性をフ。ロットしてグラ フを描き、回帰式からヒアルロニダーゼ活性を 50%阻止するのに必要なクロロ. e d u c t i o m )。また、比較対照にはクロモグリク ゲン酸類濃度を算出した (50%r + ) c a t e c h i nを用いた。 酸ナトリウム (DSCG,抗炎症剤)と (. 2 2・1 3 . ガン細胞増殖抑制活性 クロロゲン酸類のヒト由来培養細胞に対する増殖阻害活性を測定した。供試. 41.
(42) 細胞にはヒト由来の接着ガン細胞である KB細胞(ヒト鼻咽頭ガン由来)と. MCF-7細胞(ヒト乳ガン由来)、浮遊ガン細胞である U937細胞(ヒト組織球 性リンパ腫由来)を使用した。 KB細胞は島根県立保健環境科学研究所. 持田恭. 博士より分譲していただいた。 MCF-7細胞は ATCC (HTB22,USA) から、. U937細胞はヒューマンサイエンス財団(大阪)から入手した。正常細胞とガン 細胞に対する増殖阻害活性を比較するためにヒト由来接着性 WI-38細胞(肺繊 維芽細胞由来正常細胞)と、これを SV40 ウイルスで形質転換しガン化させた. WI-38VA13sub2RA細胞 (VA細胞)をヒューマンサイエンス財団から入手し た 。. U937細胞は、 100m l / L牛胎児血清 (FBS,FlowL a b o r a t o r i e s )RPMImedium (1640培地、 SigmaAld r i c hC o . ) を用いて 37C、5%炭酸ガス解卵器の中で培 0. l / L 子牛血清 (CS,Flow L a b . ) -E a g l e ' s minimal 養した。 KB 細胞は 20m. e s s e n t i a lmedium (MEM培地、日水製薬)で、 MCF-7細胞と WI-38細胞, WI38VA細 胞 は 100m l / L 牛胎児血清- MEM培地を用いて、 U937 細 胞 は 100ml/LFBS-RPMImediumを用いて 37C、5%炭酸ガス解卵器の中で培養し 0. 0 0 μg / m lおよ た。全ての培地にはペニシリン 100U/mL、ストレプトマイシン 1 0 μ g/mlを添加した。 びカナマイシン 6 供試培養細胞をクロロゲン酸類添加培地で培養し、ガン細胞増殖抑制活性を. WS T -1法 82)で測定した。そして、ガン細胞の増殖を 50%抑制させるのに必要な e d u c t i o n、I C 5 o ) を算出した。 クロロゲン酸類濃度 (50%r U937) は 96穴組織培養用マイクロプレート ( F a l c o n ) の各 浮遊性ガン細胞 ( 6c w e l lに 1X10 e l l s / m lに調整した細胞懸濁液を 50μ1分注して 370Cで 3時間保. った後、目的の濃度に希釈したクロロゲン酸類溶液 50μ1を添加して、全量を. 100μlとした。 24時間培養後、 WST-1試薬(株式会社同仁化学研究所)を 10 μl添加し 3時間インキュベートした後、プレートアナライザー(東洋測器株式. 42.
(43) 会社)で主波長 450nm、副波長 630nmの吸光度を測定して、細胞の生存率を算 出した。 接着性培養細胞 (KB、MCF-7、WI-38、WI-38VA) は 96穴組織培養用マイ クロプレート ( F a l c o n ) の各 w e l lに 、 5X103c e l l s / m lに調整した細胞懸濁液を 0. 100μI分注して 37Cで 1日前培養し、底面に培養細胞の単層を形成させた。 培養液上清を除去した後、培地で目的の濃度に希釈したクロロゲン酸類溶液を. 100μl添加した。 24時間培養後、 WST-1試薬を添加して、細胞の生存率を算出 した。ガン細胞増殖抑制活性は以下の式から求めた。 ガン細胞増殖抑制活性(%) ={[(サンプル無添加培養液の 450nmの吸光度)(培地の 450nmの吸光度)]-[(サンフ。/レ添加培養液の 450nmの吸光度)-(サンプ ル添加培地の 450nmの吸光度)]I[(サンプル無添加培地の 450nmの吸光度)-. 0 0 . (培地の 450nmの吸光度)]} x1 また、クロロゲン酸類添加培地で培養した U937 細胞を Hoechst33258. Ald r i c h )で、染色し、蛍光顕微鏡を用いて核の変化を観察した。 (Sigma-. 2 2・ 1 4 .抗変異原性試験 ウムラック AT(日本抗体研究所)キットを用いて、 0・ガラクトシダーゼ活性 を指標として抗変異源活性を測定した。供試菌s.かphimuriumTA1535/pSK1002 には、突然変異に関わる umu遺伝子に転写・翻訳の開始部位を欠く l acZ遺伝. muC'-' l acZ融合遺伝子が形質転換法で導入されている。変異原 子を融合した u 物質によって SOS反応が誘発されると、 umu遺伝子フ。ロモーターの調節下にあ. muC'-' l acZ 融合遺伝子が発現して 0・ g a l a c t o s i d a s e活性を持つ umuC'-' L acZ るu 雑種蛋白が形成される。したがって、 sg a l a c t o s i d a s e活性は SOS反応すなわ ち変異原性の強さを反映している。. 96穴マイクロプレートに終濃度 0.85mMになるように調製したクロロゲン酸. 4 3.
(44) 10%(w/v)DM80 に 溶 解 し た 7 種 類 の 変 異 原 物 質 T r p・ p・ 1. 類と. 4d i m e t h y l 3-amino-1,. τ ' rpp 2. 5Hp y r i d o [4, 3 -b l i n d o l e. ・. [ 4, 3・b l i n d o l e. p y r i d o・ 3-amino-1・methyl-5H-. ・. AF-2. (a l p h a 2 f u r y l 5 n i t r o・2-furanacrylamide). 2-AA(2-Amino- anthracene). 2 B[a]pU, ・. benzopyrene). 4-NQO (4-nitroquinoline N-Oxide) . IQ. (2-amino-3-methylimodazo-[ 4, 5 f ] q u i n o l i n e )をそれぞれ 10μl加えた。変異原 a i t oら 86)の報告を参考 物質の最終濃度は Yamadaら 83)、Yoshimotoら 84,85)、8 に Table7に示した。 AF2を添加した w e l lには Nutrientbrothで前培養した 0. S .t y p h i m u r i u mTA1535/p8K1002 を 100μl加え 37Cで 2時間インキュベートし. た。残り 6種類の変異原物質(官p・ P-1、Tr p・ P-2、2-AA、B[a]p、4-NQO、 IQ) はヒト肝臓に取り込まれてから代謝活性化されるので、 8-9MIXとともに試料. . をインキュベートしてから菌液を添加した。 37Cで 120 分間保った後、 S 0. かphimuriumTA1535/p8K1002の生産する菌体外 0・ガラクトシダーゼ活性を測 定した。 供試試料のみ添加した反応液の酵素活性を 100とし、供試試料と変異原物質を 添加した反応液の活性を相対活性で表示した。 T a b l e7 .F i n a lc o n c e n t r a t i o no f r e a g e n t s . R e a g e n t. F i n a lc o nc . (n g / w e l l ). *Tr p P l( 3 a m i n o l, 4 d i m e t h y l 5 H p y r i d o [ 4 . 3 b ] i n d o l e ) * T r o P 2 ( 3 a m i n o l m e t h v l 5 H o v r i d o r 4 . 3 b l i n d o l e ) A F 2 ( a l p h a 2 f u r y l一5 n i t r o 2 f u r a n a c r y l a m i d e ) * 2 A A ( 2 A m i n o a n t h r a c e n e ) * B [ a ] p ( 1 . 2 b e n z o p y r e n e ) * 4 N Q O ( 4 n i t r o q u i n o l i n eN O x i d e ) * I Q ( 2 a m i n o 3 m e t h y l i m o d a z o [ 4 . 5 f] q u i n o l i n e ) * T h ea n t i m u t a g e n i c i t vw a st e s t e dw i t hS 9m i x .. 4 4. 2 0 1 0 0 3 3 0 2 0 4 5 0.
図


関連したドキュメント
め測定点の座標を決めてある展開図の応用が可能であ
(2) カタログ類に記載の利用事例、アプリケーション事例はご参考用で
「心理学基礎研究の地域貢献を考える」が開かれた。フォー
このため、都は2021年度に「都政とICTをつなぎ、課題解決を 図る人材」として新たに ICT職
注:一般品についての機種型名は、その部品が最初に使用された機種型名を示します。
えて リア 会を設 したのです そして、 リア で 会を開 して、そこに 者を 込 ような仕 けをしました そして 会を必 開 して、オブザーバーにも必 の けをし ます
経済学研究科は、経済学の高等教育機関として研究者を
なお、関連して、電源電池の待機時間については、開発品に使用した電源 電池(4.4.3 に記載)で
