Nagoya City University Academic Repository
学 位 の 種 類 博士 (薬学) 報 告 番 号 甲第1718号 学 位 記 番 号 第352号 氏 名 山影 祐子 授 与 年 月 日 平成 31 年 3 月 31 日 学位論文の題名 生後脳におけるリーリン特異的切断の機構、および、その阻害によるアル ツハイマー病態抑制効果の解明 論文審査担当者 主査: 松永 民秀 副査: 服部 光治, 頭金 正博, 田中 正彦
名古屋市立大学学位論文
生後脳におけるリーリン特異的切断の機構、
および、その阻害による
アルツハイマー病態
抑制効果の解明
平成
30 年度(2019 年 3 月)
名古屋市立大学大学院薬学研究科
病態生化学分野
山影 祐子
一. 本論文は 2019 年 3 月、名古屋市立大学大学院薬学研究科において審査さ れたものである。 主査 松永 民秀 教授 副査 服部 光治 教授 副査 頭金 正博 教授 副査 田中 正彦 准教授 二. 本論文は、学術情報雑誌に収載された次の報文を基礎とするものである。 【基礎となる報文】
1. Yuko Yamakage, Hitomi Tsuiji, Takao Kohno, Himari Ogino, Takashi Saito, Takaomi C. Saido, and Mitsuharu Hattori
Reducing ADAMTS-3 inhibits amyloid β deposition in App knock-in mice Biol. Pharm. Bull., 42, 354-356. (2019)
2. Yuko Yamakage, Michinao Kato, Aya Hongo, Himari Ogino, Keisuke Ishii, Takumi Ishizuka, Takana Kamei, Hitomi Tsuiji, Tomomi Miyamoto, Hisashi Oishi, Takao Kohno, and Mitsuharu Hattori
A disintegrin and metalloproteinase with thrombospondin motifs 2 cleaves and inactivates Reelin in the postnatal cerebral cortex and hippocampus, but not in the cerebellum.
Mol. Cell. Neurosci., 100, 103401. (2019)
三. 本論文の基礎となる研究は、服部光治教授の指導の下に名古屋市立大学大 学院薬学研究科において行われた。
本論文では、以下の略語を用いた。
ADAMTS : a disintegrin and metalloproteinase with thrombospondin motifs ApoER2 : apolipoprotein E receptor 2
Aβ : amyloid beta
App : amyloid precursor protein BSA : bovine serum albumin
CRISPR: clustered regularly interspaced short palindromic repeats CR : cysteine-rich domain
DI : disintegrin domain
DMEM : Dulbecco's modified Eagle's medium FBS : fetal bovine serum
FL : full-length
HEK : human embryonic kidney HRP : horseradish peroxidase KI : knock-in
KO : knock-out
MP : metalloprotease domain P/S : penicillin/streptomycin PBS : phosphate buffered saline
PNP/PL : procollagen N-proteinase/protease and lacunin pro : prodomain
PVDF : polyvinylidene difluoride RR : Reelin repeat
RT-PCR : reverse transcription polymerase chain reaction
SDS-PAGE : sodium dodecyl sulfate polyacrylamide gel electrophoresis SEM : standard error of the mean
SP : signal peptide spc : spacer domain TAM : Tamoxifen
TFRC:Transferrin receptor C TS : thrombospondin type 1 motif
VLDLR : very low density lipoprotein receptor WB : Western Blotting
目次 第一章 序論 1 第二章 実験材料と実験方法 5 2-1. 試薬および抗体 5 2-2. 実験動物 5 2-3. 細胞培養 5 2-4. 発現プラスミド 5 2-5. 遺伝子導入 10 2-6. 限外濾過によるタンパク質の濃縮 10 2-7. Western Blotting 法 10 2-8. panADAMTS-2/3/14 ポリクローナル抗体の樹立 11 2-9. Tamoxifen 溶液の調製 11 2-10. 免疫染色法 11 2-11. 逆転写 PCR 法 12 2-12. 統計解析 14 第三章 ADAMTS-3 の減少によるアルツハイマー病態抑制効果の解明 15 3-1. 結果 3-1-1. ADAMTS-3 を減少させれば、大脳皮質で野生型 Aβ の沈着を抑制 または遅延できる 16 3-2.考察 20
第四章 成体脳でリーリンN-t 切断を担う酵素の解明 22
4-1. 結果 4-1-1. in vitro で、ADAMTS-2 はリーリン N-t 切断活性を有する 23
4-1-2. ADAMTS-2 と ADAMTS-3 のリーリン N-t 切断活性には、Disintegrin domain や最も N 末端側の Thrombospondin motif が重要である 26
4-1-3. ADAMTS-2 KO マウスの作製 29 4-1-4. ADAMTS-2 は成体期の大脳皮質および海馬において、 リーリンN-t 切断と不活化を担う 29 4-2. 考察 32 第五章 総括 34 引用文献 36 謝辞 39
第一章
序論
記憶、思考や学習など高次機能を発揮するには神経ネットワークの形成と機 能発現が必須である。分泌タンパク質リーリンは、神経細胞の樹状突起形成を正 に制御すること (Niu et al., 2004; Matsuki et al., 2010)、神経細胞における長期増 強を誘導し、記憶能力を向上させること (Chen et al., 2005; Rogers et al., 2011)、 成体脳における神経新生に対して促進的に働くこと (Pujadas et al., 2010; Teixeira et al., 2012) が知られており、脳機能に重要なタンパク質である。また、精神神 経疾患との関連も報告されており、リーリン機能の低下がアルツハイマー病、統 合失調症、自閉症の発症あるいは増悪に関与すると考えられている。
リーリンがアルツハイマー病に対して抑制的に働くことを示唆する研究結果 は数多く報告されている。例えばin vitro の研究で、リーリンはアミロイド β ペ プチド (Aβ) の産生 (Hoe et al., 2006; Rice et al., 2013)、凝集 (Pujadas et al., 2014; Durakoglugil et al., 2009)、および毒性発現 (Pujadas et al., 2014; Durakoglugil et al., 2009)を強力に抑えることが報告されている。また、アルツハイマー病モデルマ ウスを用いた研究では、リーリンヘテロ欠損にすると症状が悪化し (Kocherhans et al., 2010)、逆にアルツハイマー病モデルマウスにリーリンを過剰発現すると症 状が改善すること (Pujadas et al., 2014)、成体特異的にリーリンを欠損させたマ ウスはAβ の毒性に対して脆弱になること (Lane-Donovan et al., 2015) が報告さ れている。したがって、ヒトの脳内でリーリンの量や活性を増強できれば、アル ツハイマー病の発症や進行が抑制できると考えられる。しかし、リーリンタンパ ク質の脳内への直接投与や、リーリン遺伝子を脳へ導入する方法はいずれも人 への侵襲が大きく、現実的な治療法ではない。そこで本研究では、リーリンの活 性調節機構に着目した。 リーリンは3,461 アミノ酸から成る巨大なタンパク質で、N 末端領域、8 回の 繰り返し構造 (リーリンリピート: RR)、C 末端領域から構成される (D’Arcangelo et al., 1995) (図 1)。リーリンは受容体であるアポリポタンパク質受容体 2 (ApoER2) や 超 低 密 度 リ ポ タ ン パ ク 質 受 容 体 (VLDLR) に 結 合 す る (D’Arcangelo et al., 1999; Hiesberger et al., 1999)。これにより細胞内タンパク質 Dab1 がリン酸化され、下流シグナルが活性化される (Lee and D’Arcangelo, 2016; Wasser and Herz, 2017)。リーリンは分泌後に、RR3 内 (N-t) と RR6-7 の間 (C-t)、 C 末端領域内 (WC) で特異的な切断を受ける (Lambert de Rouvroit et al., 1999; Kohno et al., 2015)。このうち N-t 部位での特異的な切断 (N-t 切断) は、リーリン
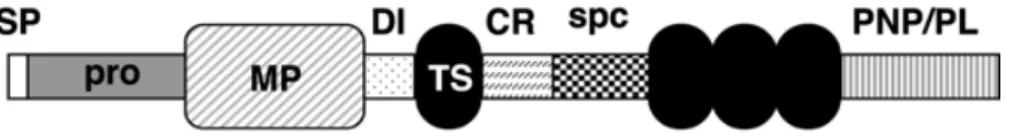
の生物活性を著しく減弱させる (Kohno et al., 2009)。したがって、リーリンの N-t 切断を担う酵素を阻害すれば、リーリン機能を増強できると考えられる。 図1 リーリンの模式図とシグナル伝達機構 リーリンはN 末端領域、8 回の繰り返し構造、C 末端領域から構成される。リーリンが受容体に 結合すると、細胞内タンパク質のDab1 がリン酸化され、続く下流シグナルが活性化される。 これまでに当研究室では、リーリンN-t 切断を担う酵素として、A Disintegrin And Metalloproteinase with Thrombospodin motifs-3 (ADAMTS-3) を 同 定 し た (Ogino et al., 2017)。ADAMTS-3 は分泌型のメタロプロテアーゼである。ADAMTS-3 は、N 末端側から signal peptide (SP)、prodomain (pro)、metalloproteinase domain (MP)、disintegrin domain (DI)、1 番目の thrombospondin type 1 motif (TS)、cysteine-rich domain (CR)、spacer domain (spc)、2 番目〜4 番目の TS、そして procollagen N-proteinase/protease and lacunin module (PNP/PL)というドメインで構成されてい る (図 2)。
図2 ADAMTS-3の構造の模式図
N 末端側から signal peptide (SP)、prodomain (pro)、metalloproteinase domain (MP)、disintegrin domain (DI)、1 番目の thrombospondin type 1 motif (TS)、cysteine-rich domain (CR)、spacer domain (spc)、 procollagen N-proteinase/protease and lacunin module (PNP/PL)で構成されている。
生体における ADAMTS-3 のリーリン N-t 切断への寄与を明らかにするため、 ADAMTS-3 knock-out (KO) マウスを解析した結果、ADAMTS-3 は胎生期と幼若 期において、大脳皮質および海馬でリーリン不活化を担う主要な酵素であるこ とが明らかとなった(Ogino et al., 2017)。 そこで、1 つ目の研究課題として、ADAMTS-3 の阻害によりアルツハイマー 病態を抑制できるか否かの解明を目指した。 1 つめの研究課題の進行と同時に、ADAMTS-3 以外にリーリン N-t 切断を担 う酵素の解明も目指した。この理由は、研究を進める過程で、ADAMTS-3 は成 体期においてもリーリンN-t 切断を担うが、他の酵素の寄与が無視できないこと が判明したためである。精神神経疾患の治療のためには、成体脳におけるリーリ ン N-t 切断機構の更なる解明が必要であると考えた。そこで 2 つめの研究課題 として、成体脳でリーリンN-t 切断を担う酵素の解明を目指した。
第二章
2-1. 試薬および抗体
試薬は、特に記載のない限り富士フィルム和光純薬株式会社から購入し、 Milli-Q (Merck Millipore) に溶解した。マウス抗リーリンモノクローナル抗体 G10 (Merck Millipore)、ウサギ抗 Dab1 ポリクローナル抗体 (当研究室鮫島が作製)、 マウス抗β-Actin モノクローナル抗体 AC15 (Sigma-Aldrich)、マウス抗 Myc モノ クローナル抗体 9E10 (Sigma-Aldrich)、マウス抗 Aβ モノクローナル抗体 82E1 (株式会社免疫生物研究所)、Horseradish peroxidase (HRP) 標識二次抗体 (GE Healthcare)、蛍光標識二次抗体 (Thermo Fisher Scientific) を使用した。
2-2. 実験動物
Rosa26-CreERT2 (B6.129-Gt(ROSA)26Sortm1(cre/ERT2)Tyj/J) マ ウ ス は The Jackson
Laboratory より購入した。Adamts-3floxマウスの作製方法は以下の通りである。ま
ず、ADAMTS-3 KO(Adamts3tm1a(KOMP)Wtsi-3) マウスを国際ノックアウトマウスプ
ロジェクトから入手した。その後、理研バイオソース研究センターより入手した B6-Tg(CAG-FLPe)36 マウスと交配し、Adamts-3floxマウスを得た。
2-3. 細胞培養
ヒト小児腎癌由来細胞HEK293T 細胞は、独立行政法人理化学研究所、脳科学 総合研究センターの御子柴克彦チームリーダーより御供与頂いた。HEK293T 細 胞は10% (v/v) Fetal bovine serum (FBS, Corning)、50 µg /mL Penicillin および 50 U/mL Streptomycin (P/S, Nacalai tesque) を含む Dulbecco's modified Eagle's medium (DMEM) を用いて、37 ˚C、5 % CO2存在下で培養した。
2-4. 発現プラスミド
以 下 で 作 製 し た プ ラ ス ミ ド の 塩 基 配 列 は 全 てApplied Biosystems™ 3130 Genetic Analyzer (Applied Biosystems) により確認した。また、使用した制限酵素 はTakara Bio、TOYOBO、およびNew England Biolabsから購入した。
2-4-1. ADAMTS-3-Myc 発現プラスミド
pENTR22.1 ベクターに挿入された ADAMTS-3 全長発現プラスミド DNA を Dharmacon より購入した。まず、ADAMTS-3 in pENTR22.1 を HpaI/XbaI を用い てpcDNA3.1 Zeo (+) (Thermo Fisher Scientific) に挿入した。次に ADAMTS-3-Myc
発現プラスミドをクローニングするため、全長 ADAMTS-3 を開始コドンから BamHI 認識配列まで、BamHI 認識配列から終止コドンまでの 2 分割に区切って 考え、順に①、②と名付けた。 ①のクローニングは、鋳型として ADAMTS-3 in pcDNA3.1/Zeo (+) を用いて BamHI 間を切り出した。②のクローニングは、鋳型として pcDNA3.1/Zeo (+) に 挿入された ADAMTS-3 を用い、2899 塩基〜3609 塩基までの PCR 産物 A を得 た。PCR 産物 A の BamHI〜XbaI 間を切り出し、pcDNA3.1 (+)/Myc-His A (Thermo Fisher Scientific) の BamHI〜XbaI 間に挿入して作製した。更に、②を挿入したプ ラスミドを BamHI で処理し、①を挿入して ADAMTS-3-Myc 発現プラスミドを 作製した。 ・PCR 産物 A を作製するためのプライマー ADAMTS3-2899-13 5' - CCAGCTCAGTGGAAAACAGGATC - 3' ADAMTS3-3609-Xb-14 (下線部は XbaI 認識部位を示す) 5' - GAGTCTAGATCTCTCTGAGGCTGTTGACC - 3' 2-4-2. ADAMTS-3 M1 変異体発現プラスミド ADAMTS-3-Myc 発現プラスミドを鋳型として PCR 産物 B を作製し、HindⅢ /XbaI を用いて pcDNA3.1(+)/Myc-His A に挿入した。 ・PCR 産物 B を作製するためのプライマー ADTS3-1-Hd-51 (下線部は HindⅢ認識部位を示す) 5' - GCCAAGCTTATGGTGTTCCTGTCACTTTGG - 3' ADTS3-2544-Xb-52 (下線部は XbaI 認識部位を示す) 5' - CACTCTAGACTCGAAGGTATCCAGCTCCCTC - 3' 2-4-3. ADAMTS-3 M2 変異体発現プラスミド ADAMTS-3-MycHis A を鋳型として PCR 産物 C を作製し、HindⅢ/XbaI を用 いてpcDNA3.1(+)/Myc-His A に挿入した。 ・PCR 産物を作製するためのプライマー ADTS3-1-Hd-51 (下線部は HindⅢ認識部位を示す) 5' - GCCAAGCTTATGGTGTTCCTGTCACTTTGG - 3' ADTS3-1815-Xb-56 (下線部は XbaI 認識部位を示す) 5' - GGGTCTAGAGCATTCTTCTGTATTACAAAGC - 3'
2-4-4. ADAMTS-3 M4 変異体発現プラスミド ADAMTS-3-MycHis A を鋳型として PCR 産物 D を作製し、HindⅢ/XbaI を用 いてpcDNA3.1(+)/Myc-His A に挿入した。 ・PCR 産物 D を作製するためのプライマー ADTS3-1-Hd-51 (下線部は HindⅢ認識部位を示す) 5' - GCCAAGCTTATGGTGTTCCTGTCACTTTGG - 3' ADTS3-1257-Xb-54 (下線部は XbaI 認識部位を示す) 5' - GGGTCTAGAAGTCTCATCGCCACACCG - 3' 2-4-5. ADAMTS-2-Myc 発現プラスミド
pCMV-SPORT6 ベクターに挿入された ADAMTS-2 全長発現プラスミド DNA をDharmacon より購入した。これを鋳型として PCR 産物 E を作製し、EcoRI/XhoI を用いてpcDNA3.1(+)/Myc-His A 挿入した。 ・PCR 産物 E を作製するためのプライマー ADTS2-80-R1-35 (下線部は EcoRI 認識部位を示す) 5' - GGGGAATTCATGGATCCGCCGGCGGGAGCC - 3' ADTS2-3718-Xh-22 (下線部は XhoI 認識部位を示す) 5' - GGGCTCGAGGAACTTTCCCGGCTTCTC - 3' 2-4-6. ADAMTS-2 M1 変異体発現プラスミド ADAMTS-2-Myc 発現プラス ミドを鋳型 として PCR 産物 F を作製 し、 EcoRI/XhoI を用いて pcDNA3.1/MycHis A に挿入した。 ・PCR 産物 F を作製するためのプライマー ADTS2-80-R1-35 (下線部は EcoRI 認識部位を示す) 5' - GGGGAATTCATGGATCCGCCGGCGGGAGCC - 3' ADTS2-2653-Xh-26 (下線部は XhoI 認識部位を示す) 5' - GGGCTCGAGCTCGTGGCGCACAGCATC - 3' 2-4-7. ADAMTS-2 M2 変異体発現プラスミド ADAMTS-2-Myc 発現プラスミドを鋳型として PCR 産物 G を作製し、 EcoRI/XhoI を用いて pcDNA3.1/MycHis A に挿入した。
・PCR 産物 G を作製するためのプライマー ADTS2-80-RI-36 (下線部は EcoRI 認識部位を示す) 5' - GGGGAATTCATGGATCCGCCGGCGGGAGCC - 3' ADTS2-1927-Xh-28 (下線部は XhoI 認識部位を示す) 5' - GGGCTCGAGACAGTCCTGGGGGTTGCAGAGC - 3' 2-4-8. ADAMTS-2 M3 変異体発現プラスミド ADAMTS-2-Myc 発現プラスミドを鋳型として PCR 産物 H を作製し、 EcoRI/XhoI を用いて pcDNA3.1/MycHis A に挿入した。 ・PCR 産物 H を作製するためのプライマー mADTS2-1-RI-31 (下線部は EcoRI 認識部位を示す) 5' - TAAGAATTCATGGATCCGCCGGCGGGAGC - 3' mADTS2-1683-Xh-34 (下線部は XhoI 認識部位を示す) 5' - CCCCTCGAGTCGTTTGAGGATATCGGGTG - 3' 2-4-9. ADAMTS-2 M4 変異体発現プラスミド ADAMTS-2-Myc 発 現 プ ラ スミ ドを 鋳型と し て PCR 産物 I を 作製 し 、 EcoRI/XhoI を用いて pcDNA3.1/MycHis A に挿入した。 ・PCR 産物 I を作製するためのプライマー mADTS2-1-RI-31 (下線部は EcoRI 認識部位を示す) 5' - TAAGAATTCATGGATCCGCCGGCGGGAGC - 3' mADTS2-1413-Xh-32 (下線部は XhoI 認識部位を示す) 5' - CCCCTCGAGGGGATCATCCCGCAGGCAGTC - 3' 2-4-10. ADAMTS-14-Myc 発現プラスミド
pCR-XL-TOPO に挿入された ADAMTS-14 全長発現プラスミド DNA を鋳型と し、EcoRI を用いて pcDNA3.1/Zeo (+) (Life Technologies) に挿入した。更にこれ を鋳型として PCR 産物 J を得た。PCR 産物 J の EcoRⅠ-XhoⅠ間を切り出し、 pcDNA3.1 (+)/Myc-His A の EcoRI〜XhoI 間に挿入して作製した。
・PCR 産物 J を作製するためのプライマー
ADTS14-37-R1-41 (下線部は EcoRI 認識部位を示す) 5’ - GGGGAATTCATGGCTTGGCTCCGAGCTCTGC - 3’
ADTS14-3657-Xh-42 (下線部は XhoI 認識部位を示す) 5’ - GGGCTCGAGTGTCACTGGGGAGGTGGCAGG- 3’
2-5. 遺伝子導入
10 cm dish に対し 80 ~ 100%コンフルエントの HEK293 T 細胞に DNA 15 µg/dish とDNA 1 µg あたり 2 µL の Polyethyleneimine "Max" (Polysciences) を Opti-MEM (Thermo Fisher Scientific) 中で混合し、室温で 20 分間インキュベートした。次に、 Opti-MEM に培地交換した HEK293T 細胞に添加した。添加から 5 時間後、Opti-MEM に培地交換し、37 ˚C、5% CO2存在下で48 時間培養した。培養上清はクリ
ーンベンチ内で無菌チューブに回収し、440 g、5 分遠心後、上清を別の無菌チュ ーブに回収した。
2-6. 限外濾過によるタンパク質の濃縮
HEK293T 細胞に発現させて回収した ADAMTS-2、ADAMTS-3、ADAMTS-14 のいずれかを含む培養上清はAmicon Ultra Centrifugal Filter Device 50 kDa cutoff (Merck Millipore) にアプライし、5 倍〜30 倍に濃縮した。ADAMTS-2 欠失変異 体および ADAMTS-3 欠失変異体を含む HEK293T 細胞培養上清の濃縮には、 Amicon Ultra Centrifugal Filter Device 10 kDa cutoff (Merck Millipore) を使用し、1 倍〜30 倍に濃縮した。濃縮倍率が 1 倍のサンプルについては、適宜濃縮後、培 地であるOpti-MEM で希釈した。
2-7. Western Blotting (WB) 法
2-7-1. 使用した buffer
4xSDS-PAGE sample beffer: 250 mM Tris-HCl pH 6.8 (Nacalai tesque), 40% (v/v)
Glycerol, 0.2% (w/v) Bromo phenol blue, 8% (w/v) Sodium dodecyl sulfate (SDS), 20% (v/v)
2-mercaptoethanol (2-ME)
SDS-PAGE electrode buffer: 2.5 mM Tris, 19.2 mM Glycine, 0.01% (w/v) SDS Transfer buffer: 47.9 mM Tris, 39.3 mM Glycine, 20% Methanol, 0.001% (w/v) SDS TBS-T: 20 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.05% Tween® 20
2-7-2. 実験方法
解析するサンプルと4xSDS-PAGE sample buffer を 3:1 で混合した。1 レーンあ たり7~35 µL のサンプルを SDS-ポリアクリルアミドゲル電気泳動 (SDS-PAGE) により分離し、セミドライ式転写機 (Trans-blot SD cell, Bio-Rad) を用いて PVDF 膜 (Merck Millipore) に転写した。転写後、PVDF 膜を 5%スキムミルク (雪印メ グミルク) in TBS-T で 30 分ブロッキングした。その後、PVDF 膜と一次抗体を 含む 5%スキムミルク in TBS-T を 4˚C で一晩反応させた。反応後、TBS-T で 5 分間洗浄することを4 回繰り返した。次に、PVDF 膜と HRP 標識された二次抗 体を含む5%スキムミルク in TBS-T を室温で 1 時間反応させた。反応後、TBS-T で5 分間洗浄することを 4 回繰り返し、Immobilon Western Chemiluminescent HRP Substrate (Merck Millipore) または ECL Select Western Blotting Detection Reagent (GE Healthcare) を用いて可視化し、LAS 4000 mini (Fujifilm) を用いて検出した。 なお、抗リーリン抗体は1:1000、抗 Dab1 抗体は 1:1000、抗 β-Actin 抗体は 1:10000、 抗Myc 抗体は 1:1000、panADAMTS-2/3/14 抗体は 1:1000 で使用した。HRP 標識 二次抗体は1:4000 で使用した。
2-8. panADAMTS-2/3/14ポリクローナル抗体の樹立
Eurofin 株式会社に委託し、日本白色ウサギ 1 羽に ADAMTS-2/3/14 の共通配 列 で あ る panADAMTS-2/3/14 (CNHEDGFSSAFVVA-HETGH 、 cystein 残 基 は conjugation のために付加した) を抗原として免疫した。6 週間 (計 4 回) 免疫し、 全採血することでpanADAMTS-2/3/14 ポリクローナル抗体を得た。
2-9. Tamoxifen (TAM) 溶液の調製
Tamoxifen (Cayman Chemical) をコーン油 (Sigma-Aldrich) に溶解し、20 mg/mL のTAM 溶液を調製した。
2-10. 免疫染色法
2-10-1. 使用した buffer
10 mM sodium citrate pH 6.0: 10 mM Citric acid monohydrate TBS-T: 20 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.05% Tween® 20
2-10-2. 実験方法
マウスを4% PFA in PBSで灌流固定し、全脳を摘出した。更に4% PFA in PBS中 で一晩固定した。続けて、20% Sucrose in PBS、30% Sucrose in PBSの順に固定し た全脳を4 ˚Cで一晩浸漬した。次に、固定した全脳からの一部を切り取り、75% Tissue-Tek O.C.T. Compound (Sakura Finetek) in 30% Sucrose 中に沈め、ドライア イス上で包埋した。包埋した脳からcryostat (CM 1850, Leica Microsystems) を用 いて14 µmの切片を作製し、MASコート付きスライドガラス (松浪) に貼付後、 室温で乾燥させた。得られた脳切片をPBSで5分間、2回洗浄し、10 mM Sodium Citrate pH 6.0中で105 ˚C、5分間加熱処理することで抗原賦活化した。放冷後、 TBS-Tで5分間、2回洗浄し、2% BSA in TBS-Tを用いて室温で1時間ブロッキング した。そして、一次抗体を含む2% BSA in TBS-Tと4 ˚C、一晩反応させ、TBS-Tで 4回洗浄した。蛍光標識二次抗体および2 µg/mL Hoechst 33342 (Thermo Fisher Scientific) を含む2% BSA in TBS-Tと室温で2時間反応させた。TBS-Tで4回洗浄 した後、Fluorescence Mounting Medium (Dako) を用いてスライドガラスに封入し た。作製したサンプルは、BIOREVO BZ-9000 (KEYENCE) を用いて観察した。 なお、抗Aβ抗体は1:100、蛍光標識二次抗体は1:500で使用した。
2-11. 逆転写PCR (RT-PCR) 法
2-11-1. total RNA 抽出
12 ヶ 月 齢 Adamts-3flox/flox;AppNL-F/NL-F マ ウ ス と Adamts-3flox/flox
;Rosa26-CreERT2/+;AppNL-F/NL-Fマウスの海馬からtotal RNA を抽出するために、RNeasy Mini
kit (Qiagen) を用いた。組織に 600 µL の 2-ME 含有 RLT を加え、25G (テルモ) ニ ードルで組織を破砕した。次に、600 µL の 70 % Ethanol を加え、よく混合し、 RNeasy スピンカラムに全量アプライした。8,000 g、25 ˚C、15 秒間遠心した後、 RW1 を 700 µL 加え、再度 8,000 g、25 ˚C、15 秒間遠心した。更に 500 µL の RPE を加え、8,000 g、25 ˚C、15 秒間遠心することを 2 回繰り返した。RNase-free water を30 µL を加え、8,000 g、25 ˚C、1 分間遠心したものを回収して total RNA を得 た。 1 ヶ月齢の野生型マウスと ADAMTS-2 KO マウスの大脳皮質および海馬から RNA を抽出するために、TRIzol (Thermo Fisher Scientific) を用いた。摘出した組 織に1.2 mL の TRIzol を加え、20G (テルモ) ニードルで組織を破砕した。室温で 5 分間静置した後、250 µL の Chloroform を加え、15 秒間、激しく転倒混和した。
室温で3 分間静置した後、12,000 g、4 ˚C で 15 分間遠心した。上層から 750 µL を回収し、700 µL の 2-propanol を加え、転倒混和後、室温で 10 分間静置した。 次に、12,000 g、4 ˚C で 15 分間遠心し、アスピレーターで上清を除いた。ペレッ トに75% Ethanol を 1.3 mL 加え、しっかり混合した後、7,500 g、4 ˚C で 5 分間 遠心し、アスピレーターで上清を除いた。この操作を2 回繰り返した。ペレット を風乾した後、30 µL の RNase-free water を加え、55 ˚C で 10 分間加熱処理する ことでtotal RNA を得た。 2-11-2. cDNA 合成
5 µg の total RNA、1µL の 500 µg/mL Random Primers (Promega) および 4 µL の 2.5 mM dNTP (Takara Bio) を含む混合液 (RNase-free water で計 13 µL に合わせ る) を 65 ˚C、5 分間加熱処理した。その後、氷上で 1 分以上氷冷した。その後、 7 µL の cDNA 合成ミックス (4 µL の 5xRT buffer (Thermo Fisher Scientific)、1 µL の 0.1 M DTT (Thermo Fisher Scientific)、1 µL の 40 U/ µL Recombinant RNase Inhibitor (Takara Bio)、 1 µL の 200 U/ µL Superscript™ III RT (Thermo Fisher Scientific)で計 7µL) を混合した。55 ˚C、60 分間、加熱処理を行ったのち、75 ˚C、 15 分間、加熱処理を行うことで cDNA を合成した。
2-11-3. RT-PCR
Total RNA から逆転写して得た cDNA を鋳型として RT-PCR を行った。RT-PCR には、ExTaq (Takara Bio) と下記プライマーを用いた。ADAMTS-2 を検出するた めに、94 ˚C で 3 分を 1 サイクル、94 ˚C で 20 秒、63 ˚C で 20 秒、72 ˚C で 15 秒 を30 サイクル、25 ˚C で 3 分を 1 サイクルで増幅反応を行った。TRANSFERRIN RECEPTOR C (TFRC)を検出するために、94 ˚C で 3 分を 1 サイクル、94 ˚C で 20 秒、59 ˚C で 20 秒、72 ˚C で 15 秒を 30 サイクル、25 ˚C で 1 分を 1 サイクルで 増幅反応を行った。ADAMTS-2 を標的とした場合は 138 base pairs (bp) の、TFRC を標的とした場合は110 bp の amplicon が得られる。 ・ADAMTS-2 を標的としたプライマー AD2-ex17-18-03 5' - ACAATGTCCTGGAAGATGATGCTGTG - 3' AD2-ex17-18-04 5' - GGTGTACCATCTTGCTGTCCAACCT - 3'
・TFRC を標的としたプライマー Tfrc-4081-01 5' - TGTTTGCTGGTGACAAAGGA - 3' Tfrc-4190-02 5' - CCCTCCCAAAAACCAAAAAT - 3' 2-11-4. 定量的 RT-PCR
Total RNA から逆転写して得た cDNA を鋳型として RT-PCR を行った。RT-PCR にはTaqMan Gene Expression Assays (Thermo Fisher Scientific)を用い、50 °C で 2 分を1 サイクル、95 °C で 20 秒を 1 サイクル、95 °C で 1 秒、60°C で 20 秒を 40 サイクルで増幅反応を行なった。解析には StepOnePlus system (Thermo Fisher Scientific)を用いた。
2-12. 統計解析
統計データは全て、mean ±SEM で表記した。異なる 3 群間を検定する場合に はTukey-Kramer multiple comparison test を用いた。異なる 2 群間を検定する場合 には Student’s t-test を用いた。統計解析には Microsoft Excel、および Prism7 (GraphPad software) を用いた。*p < 0.05, **p < 0.01, ***p < 0.001, n.s., not significant.
第三章
ADAMTS-3 の減少による
3-1. 結果
3-1-1. ADAMTS-3を減少させれば、大脳皮質で野生型Aβの沈着を抑制または
遅延できる
ADAMTS-3 の減少がアルツハイマー病態を抑制できるか否かを解明するため、 アルツハイマー病態を示すマウスで成体特異的にADAMTS-3 を欠損させた。目 的のマウスを作製するため、まず、Adamts-3floxマウスと、TAM 投与により全身
で Cre リコンビナーゼを発現する Rosa26-CreERT2 マウスを交配し、
Adamts-3flox/flox;Rosa26-CreERT2/+マウスを得た (図 3A)。次に、得られたマウスと、アルツ
ハイマー病態型モデルマウスを交配した。アルツハイマー病態型モデルマウス として、ヒト型のAβ を発現する AppNL-Fノックイン (KI) マウスおよび App NL-G-F-KI マウスの 2 系統を用いた (Saito et al., 2014)。AppNL-F-KI マウスは、家族性ア
ルツハイマー病患者で発見された、Swedish 変異 (NL) と、Beyreuther/Iberian 変 異 (F) が導入されている。その結果、ヒトの野生型 Aβ を多く発現する。App NL-G-F-KI マウスは、Swedish 変異と Beyreuther/Iberian 変異に加え、Arcitc 変異 (G)
が導入されている。この結果、凝集性の高い変異型Aβ を多く発現する。Adamts-3flox/flox;Rosa26-CreERT2/+マウスと各アルツハイマー病態型モデルマウスを交配し、
Adamts-3flox/flox;Rosa26-CreERT2/+;AppNL-F/NL-F マ ウ ス と Adamts-3flox/flox
;Rosa26-CreERT2/+;AppNL-G-F/NL-G-Fマウスを得た。それぞれの対照実験には、Cre リコンビ
ナーゼを発現しない Adamts-3flox/flox;AppNL-F/NL-F マウスと Adamts-3flox/flox;App NL-G-F/NL-G-Fマウスを用いた。以下、Cre リコンビナーゼを発現しないマウスを Flox、
Cre リコンビナーゼを発現するマウスを conditional KO: cKO を表記する。これら のマウスが3 ヶ月齢の時に、TAM を 6 mg/40 g マウス体重で 5 日間連続、腹腔 内投与した。定量的RT−PCR により TAM 投与後の ADAMTS-3 mRNA の発現量 を解析した結果、cKO マウスでは Flox マウスと比較して ADAMTS-3 が約 15%減 少していた。TAM 投与後、AppNL-G-F系統のマウスは5 ヶ月齢で、AppNL-F系統の
マウスは 12 ヶ月齢で脳組織を摘出した。AppNL-G-F-KI マウスは 2 ヶ月齢から脳
内へのAβ 沈着を示し、7 ヶ月齢で飽和状態に達する。AppNL-F-KI マウスは 6 ヶ
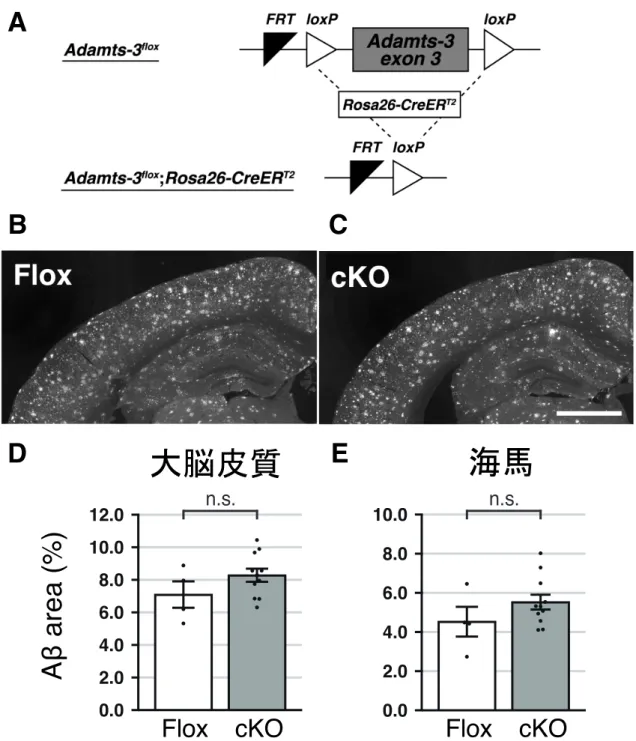
月齢から脳内へのAβ 沈着を示し、24 ヶ月齢まで進行が続く。そのため解析は、 このほぼ中間の時期である5 ヶ月齢と 12 ヶ月齢に設定した。脳組織摘出後、凍 結切片を作製し、抗Aβ 抗体を用いた免疫染色を行いた。そして大脳皮質および 海馬を占めるAβ の割合を ImageJ で定量した。5 ヶ月齢 AppNL-G-F系統では、Flox
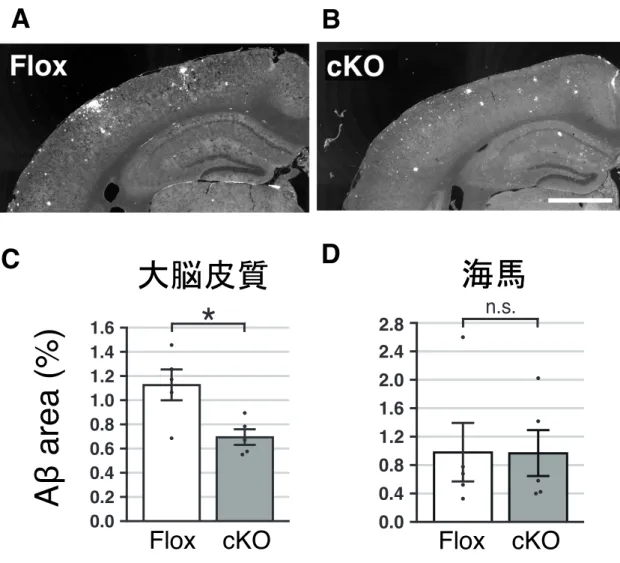
マウスと cKO マウスで脳内を占める Aβ の割合に変化はなかった (図 3B-3E)。 一方、12 ヶ月齢 AppNL-F系統のcKO マウスは、Flox マウスと比較して、大脳皮
質を占めるAβ の割合が約 40%減少した (図 4)。以上の結果より、ADAMTS-3 を 減少させれば、大脳皮質で野生型Aβ の沈着を抑制あるいは遅延できることが示 唆された。
図3 AppNL-G-F-KIマウスでは、ADAMTS-3を減少させてもAβの沈着は抑制されない (A) Adamts-3floxマウスとAdamts-3flox;Rosa26-CreERT2マウスのゲノム配列の模式図。
(B-C) 5 ヶ 月 齢 Adamts-3flox/flox;AppNL-G-F/NL-G-F (Flox) マ ウ ス (B) と Adamts-3flox/flox
;Rosa26-CreERT2/+;AppNL-G-F/NL-G-F (cKO) マウス (C) の脳コロナル切片を作製し、抗 Aβ 抗体を用いて免疫
染色を行った。Scale bar: 1mm.
(D-E), Flox マウスと cKO マウスの大脳皮質 (D)および海馬(E)を占める Aβ の面積の割合を定 量した。棒グラフをmean ± SEM で表した(Flox: n =4、cKO: n = 11)。n.s., not significant.
D
E
A
B
C
A
β
a
re
a
(
%
)
Flox
cKO
図4 AppNL-F-KIマウスでは、ADAMTS-3を減少させると大脳皮質でAβの沈着が抑制される (A-B) 12 ヶ 月 齢 Adamts-3flox/flox;AppNL-F/NL- (Flox) マ ウ ス (A) と Adamts-3flox/flox
;Rosa26-CreERT2/+;AppNL -F/NL-F(cKO)マウス (B) の脳コロナル切片を作製し、抗 Aβ 抗体を用いて免疫染
色を行った。Scale bar: 1mm.
(C-D) Flox マウスと cKO マウスの大脳皮質 (C) および海馬 (D) を占める Aβ の面積の割合を定 量した。棒グラフをmean ± SEM で表した(:n =5 ずつ)。*p < 0. 05, n.s., not significant.
D
A
B
C
A
β
a
re
a
(
%
)
Flox cKO3-2. 考察
本研究では、リーリンのN-t 切断による不活化に着目し、リーリン N-t 切断酵 素として我々が同定したADAMTS-3 の阻害によってアルツハイマー病態が抑制 できるか否かを検証した。結果、成体で ADAMTS-3 を減少させれば野生型 Aβ の大脳皮質への沈着を抑制または遅延できることが明らかとなった。
本研究では、AppNL-G-F-KI と AppNL-F-KI マウスの 2 系統のアルツハイマー病モ
デルマウスを用いて、ADAMTS-3 の減少による Aβ 沈着抑制効果を解析した。 結果、ADAMTS-3 の減少により Aβ 沈着が抑制できたのは、AppNL-F系統のみで
あった (図 3 と図 4)。AppNL-G-F系統のマウスでAβ 沈着が減少しなかった理由と
して、以下の2 つの可能性が考えられる。1 つめは、リーリンが E22G 変異型の Aβ の凝集を抑制できない可能性である。先行研究において、リーリンは野生型 Aβ の凝集を阻害することが報告されている (Pujadas et al., 2014)。しかし、リー リンと Aβ の結合部位は明らかになっておらず、E22G 変異型の Aβ はリーリン との結合性が低くなっている可能性がある。また、E22G 変異型 Aβ は多様な形 態のAβ 繊維を形成すると報告されている (Elkins et al., 2018)。したがって、リ ーリンは野生型を含む一部の Aβ の繊維形成に対してのみ凝集抑制効果を発揮 するのかもしれない。この可能性を検討するため、今後in vitro で E22G 変異型 Aβ の凝集をリーリンが抑制できるか否かを解析すべきだと考えている。2 つめ は、より早い時期からADAMTS-3 を阻害しないと Aβ 沈着を抑制できない可能 性 で ある 。今回の 実験 では、「ヒ トへ の応用 を志向し、成体になってか ら ADAMTS-3 を阻害すること」「成体期において、ADAMTS-3 のリーリン N-t 切 断への寄与は小さくなる傾向があること」の2 点を考慮し、AppNL-G-F系統とApp NL-F系統ともに 3 ヶ月齢で TAM 投与を行なった。しかし、両系統で Aβ 沈着の進 行する速度は異なっており、AppNL-G-F系統ではAppNL-F系統よりもアルツハイマ ー病態が悪化した状態でADAMTS-3 を阻害したこととなる。したがって、例え ばAppNL-F系統で、より遅い時期 (Aβ 沈着が開始する 6 ヶ月以降) から
ADAMTS-3 を減少させた場合の Aβ 沈着抑制効果を解析すれば、2 つめの可能性を検討で きると考えられる。
12 ヶ月齢 Adamts-3flox/flox;Rosa26-CreERT2/+;AppNL-F/NL-Fマウスにおいて、大脳皮
この理由は、12 ヶ月齢では海馬への Aβ 沈着が僅かであったためだと予想して いる。したがって、海馬でも大脳皮質と同様に ADAMTS-3 の減少によって Aβ 沈着が抑制できるか否かを解明するには、12 ヶ月より老齢させたマウスを用い て検討する必要があると考えている。
ADAMTS-3 はリーリン以外の基質もいくつか報告されている (Beckhouche et al., 2016)。したがって、AppNL-F系統で見られたADAMTS-3 の減少による Aβ 沈着抑
制の効果が、リーリンN-t 切断抑制およびリーリン機能亢進を介しているかを明 らかにする必要がある。近年、当研究室ではN-t 切断抵抗型のリーリンを発現す るKI (PADV-KI) マウスを樹立した。PADV-KI マウスでは成体期においても大脳 皮質および海馬でリーリンのN-t 切断が抑制され、リーリン機能が亢進している ことが分かっている。現在はPADV-KI マウスと AppNL-F-KI マウスを交配して得
られたマウスでも、Adamts-3flox/flox;Rosa26-CreERT2/+;AppNL-F/NL-F マウスと同様に
第四章
成体脳でリーリン
N-t 切断を担う
4-1. 結果
4-1-1. in vitroで、ADAMTS-2はリーリンN-t切断活性を有する
生後脳でリーリンの N-t 切断に寄与する酵素の候補として、ADAMTS-2 と ADAMTS-14 が考えられた。その理由は、(1) ADAMTS-3 と同じ ADAMTS ファ ミリーに属し、構造や基質特異性が類似している、(2) 2 ヶ月齢のマウスの脳に おいてADAMTS-2 とADAMTS-14 の mRNA が発現しているという報告がある、 の二つである。
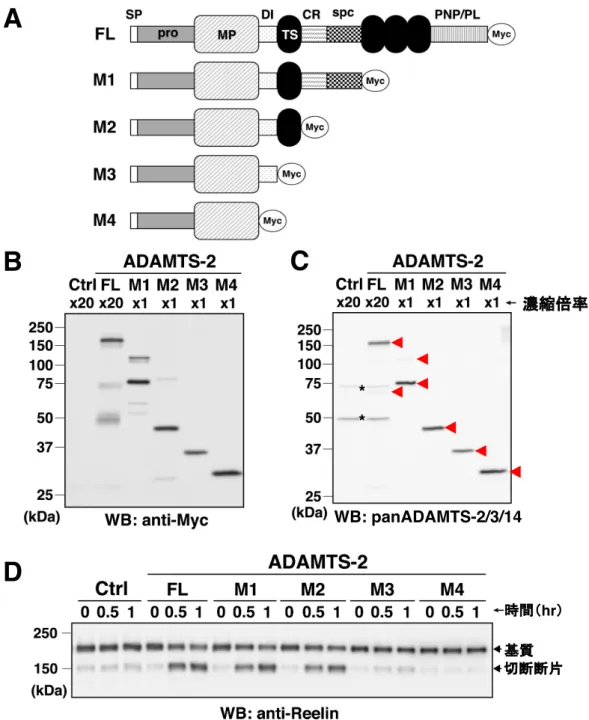
そこで、まずin vitro で、ADAMTS-2 と ADAMTS-14 のリーリン N-t 切断活性 を調べた。C 末端に Myc タグを付加した ADAMTS-2、ADAMTS-3、ADAMTS-14 のいずれかを発現した HEK293T 細胞培養上清と、人工基質 NR3 (リーリンの N 末端領域から RR3 までをもつ) を混合し、一定時間インキュベートした後、 N-t 切断断片である NR2 の量を解析した。ADAMTS-2、ADAMTS-3、ADAMTS-14 を含む培養上清は、WB での検出を容易にするため、適宜濃縮した。ADAMTS-2、ADAMTS-3、ADAMTS-14 の存在量は抗 Myc 抗体を用いて解析を行った。し かし、解析を進める過程で、ADAMTS-3 は C 末端側で複数の切断を受けるため 抗Myc 抗体で検出できないことが判明した。そこで、ADAMTS-2、ADAMTS-3、 ADAMTS-14 の MP 中の共通配列を認識する、panADAMTS-2/3/14 抗体を樹立し た。panADAMTS-2/3/14 抗体と抗 Myc 抗体の 2 つの抗体を用いることで、 ADAMTS-2、ADAMTS-3、ADAMTS-14 の存在量を解析した。
細胞画分では、ADAMTS-2 > ADAMTS-14 > ADAMTS-3 の順に多く発現して いた (図 5A-5B)。ADAMTS-14 は抗 Myc 抗体との反応性と、panADAMTS-2/3/14 抗体との反応性が大きく異なっていた。この原因は不明であるが、ADAMTS-14 が何かしらの翻訳後修飾を受けているためだと考えている。ADAMTS-3 は抗 Myc 抗体を用いた WB の長時間露光によりわずかに検出された。濃縮した培養 上清中での存在量は、ADAMTS-2 の方が ADAMTS-3 よりも多かった (図 5C-5D) 。 ADAMTS-14 は 細 胞 画 分 の 結 果 と 同 様 、 抗 Myc 抗 体 と の 反 応 性 と panADAMTS-2/3/14 抗体との反応性が異なっていたため存在量の評価は困難で あったが、ADAMTS-2 や ADAMTS-3 よりもかなり多いと判断した。また、分泌 されたADAMTS-14 は細胞画分で見られた分子量 (約 150 kDa) よりも大きな分 子量へ移行していた。この理由は不明であるが、何らかの翻訳後修飾を受けてい ると考えられる。
A
次に、濃縮した ADAMTS-2、ADAMTS-3、ADAMTS-14 のいずれかを含む培 養上清とNR3 を混合し、N-t 切断により産生した NR2 を抗リーリン抗体を用い た WB により検出した (図 5E)。結果、ADAMTS-2 を混合したサンプルは ADAMTS-3 を混合したサンプルと同程度の NR2 が存在していた。濃縮した培養 上清に含まれる ADAMTS-2 は ADAMTS-3 よりも多く存在していたため、リー リンN-t 切断活性は ADAMTS-3 の方が ADAMTS-2 よりも高いことが分かった。 一方、ADAMTS-14 を混合したサンプルでは NR2 がほとんど産生されず、 ADAMTS-14 のリーリン N-t 切断活性は ADAMTS-2 や ADAMTS-3 と比較して 著しく低かった。以上の結果より、ADAMTS-2 はリーリン N-t 切断活性を有す ることが明らかとなった。(次頁に続く)
WB: panADAMTS-2/3/14 WB: anti-MycWB: anti-Myc WB: panADAMTS-2/3/14
B
図5 ADAMTS-2はリーリンN-t切断活性を有する
(A-B) HEK293T 細胞に発現させた ADAMTS-2、ADAMTS-3、ADAMTS-14 を抗 Myc 抗体 (A) お よびpanADAMTS-2/3/14 抗体 (B) を用いた WB により検出した。陰性対照実験 (Ctrl) には空ベ クターを導入した細胞を用いた。 (C-D) HEK293T 細胞に発現させて回収した ADAMTS-2、ADAMTS-3、ADAMTS-14 を含む培養 上清を適宜濃縮し、抗Myc 抗体 (C) および panADAMTS-2/3/14 抗体 (D) を用いた WB により 検出した。赤矢頭は特異的シグナル、*は非特異的シグナルを示す。 (E) 濃縮した ADAMTS-2、ADAMTS-3、ADAMTS-14 と NR3 の混合により産生した NR2 を、抗 リーリン抗体を用いたWB により検出した。 hr
E
4-1-2. ADAMTS-2 と ADAMTS-3 のリーリン N-t 切断活性には、Disintegrin
domainや最もN末端側のThrombospondin motifが重要である
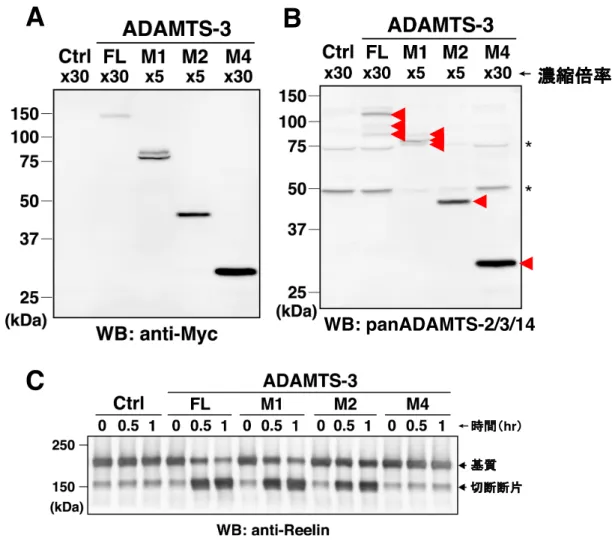
ADAMTS-2 や ADAMTS-3 のどのドメインがリーリン N-t 切断活性に重要であ るかを調べるため、ADAMTS-2 や ADAMTS-3 の C 末端から各ドメインまでを 欠損させ、C 末端に Myc タグを付加した欠失変異体 (M1、M2、M3、M4) を作 製した(図 6A)。そして、これらのリーリン N-t 切断活性を full-length (FL) と比 較解析した。 まず、ADAMTS-2 について、濃縮した ADAMTS-2 の FL と各欠失変異体の存 在量はおおよそ同程度であった (図 6B-6C)。リーリン N-t 切断活性については、 FL、M1、M2 は活性を示したが、M3 は活性が著しく低く、M4 は活性がほとん どなかった (図 6D)。この結果から、ADAMTS-2 のリーリン N-t 切断活性には、 DI および最も N 末端側の TS が重要であることが明らかとなった。 ADAMTS-3 については、M3 が得られなかったため、M1、M2、M4 を用いて 解析を行った。まず、濃縮した培養上清中でのADAMTS-3 の存在量は、M4 > M3 > M1≒FL であった (図 7A-7B)。リーリン N-t 切断活性は、M1 と M2 は活性を示 すのに対し、M4 は活性がほとんどなかった (図 7C)。以上の結果より、ADAMTS-2 と同様、ADAMTS-3 のリーリン N-t 切断活性にも、DI および最も N 末端側の TS が重要であることが明らかとなった。
図6 ADAMTS-2のリーリンN-t切断活性には、Disintegrin domainや最もN末端側の
Thrombospondin motifが重要である
(A) ADAMTS-2 と ADAMTS-3 の FL および欠失変異体 (M1-M4) の模式図。
(B-C) 濃縮した ADAMTS-2 FL と欠失変異体を、抗 Myc 抗体 (B) および panADAMTS-2/3/14 抗 体 (C) を用いた WB で検出した。陰性対照実験 (Ctrl) には空ベクターを導入して得た培養上清 を用いた。赤矢頭は特異的シグナル、*は非特異的シグナルを示す。 (D) 濃縮した ADAMTS-2 FL もしくは欠失変異体と NR3 の混合により産生した NR2 を、抗リー リン抗体を用いたWB により検出した。 WB: panADAMTS-2/3/14
C
hrA
B
D
図7 ADAMTS-3のリーリンN-t切断活性には、Disintegrin domainや最もN末端側の
Thrombospondin motifが重要である
(A-B) 濃縮した ADAMTS-3 FL と欠失変異体を、抗 Myc 抗体 (A) および panADAMTS-2/3/14 抗体 (B) を用いた WB で検出した。陰性対照実験 (Ctrl) には空ベクターを導入して 得た培養上清を用いた。赤矢頭は特異的シグナル、
*
は非特異的シグナルを示す。 (C) 濃縮したADAMTS-2 FL もしくは欠失変異体と NR3 の混合により産生した NR2 を、 抗リーリン抗体を用いたWB により検出した。 WB: panADAMTS-2/3/14 hrA
B
C
4-1-3. ADAMTS-2 KOマウスの作製
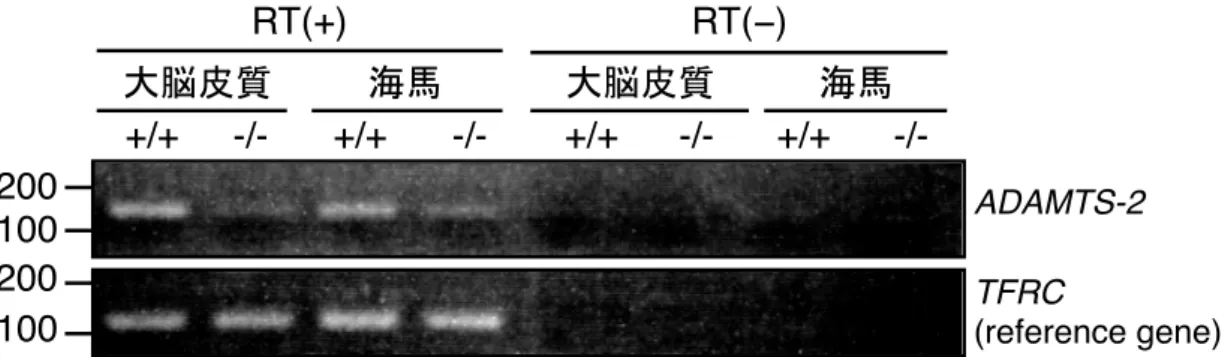
生体での ADAMTS-2 のリーリン N-t 切断への寄与を明らかにするため、 CRISPR/Cas9 システムを利用して ADAMTS-2 KO マウスを作製した。作製した ADAMTS-2 KO マウスは Adamts-2 exon 1 中の 28bp が欠損しており、フレームシ フ ト 変異 が 生じ る こ と が予 想 さ れ る 。作製した ADAMTS-2 KO マウスで ADAMTS-2 が欠損しているかを検討するため、大脳皮質および海馬を用いて RT-PCR を行った(図 8)。結果、ADAMTS-2 KO マウスでは ADAMTS-2 の多くが消失 していた。また、作製したADAMTS-2 KO マウスでは皮膚の脆弱化が見られた。 先行研究で、ADAMTS-2 deficient マウスにおいても皮膚の脆弱化が報告されて いる(Li et al., 2001)。したがって、作製した ADAMTS-2 KO マウスでは機能的 タンパク質が欠損していると考えられる。
図8 ADAMTS-2 KOマウスではADAMTS-2の多くが消失している
RT-PCR 法により 1 ヶ月齢の野生型マウス (+/+) と ADAMTS-2 KO マウス (–/–) の大脳および 海馬におけるADAMTS-2 の発現を解析した。TFRC は reference gene として用いた。DNA のサイ
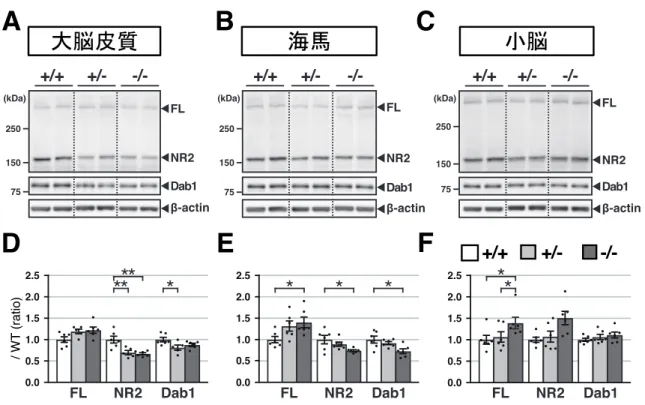
ズマーカー (bp) は泳動結果の左側に示した。 4-1-4. ADAMTS-2 は成体期の大脳皮質および海馬において、リーリン N-t 切断 と不活化を担う 成体脳におけるADAMTS-2 のリーリン N-t 切断への寄与を明らかにするため、 2 ヶ月齢および 4 ヶ月齢の ADAMTS-2 KO マウスの大脳皮質、海馬、小脳にお けるリーリンFL と NR2、Dab1 を WB により解析した。 まず、2 ヶ月齢 ADAMTS-2 KO マウスの大脳皮質では、NR2 量および Dab1 量が 減少した (図 9A と 9D)。Dab1 はリン酸化されると速やかに分解されるため、 Dab1 量の減少はリン酸化の亢進、すなわちリーリンシグナルの増強を意味する。 また、有意な差はなかったが、FL 量は増加する傾向にあった。次に海馬では、 大脳皮質と同様、NR2 量および Dab1 量が減少した (図 9B と 9E)。一方小脳で +/+ -/-200 100 200 100 RT(+) RT(−) ADAMTS-2 TFRC (reference gene) +/+ -/- +/+ -/- +/+
-/-は、NR2 量や Dab1 量に変化は見られなかった (図 9C と 9F)。以上の結果より、 ADAMTS-2 は 2 ヶ月齢のマウス大脳皮質および海馬でリーリン N-t 切断と不活 化を担うことが示唆された。
図9 ADAMTS-2は2ヶ月齢のマウス大脳皮質および海馬でリーリンN-t切断と不活化を担う (A-C) 2 ヶ月齢の野生型マウス (+/+)、ADAMTS-2 ヘテロ欠損マウス (+/–)、ADAMTS-2 KO マウ ス (–/–) の大脳皮質 (A)、海馬 (B) および小脳 (C) におけるリーリン FL と NR2 (画像上段)、 Dab1 (中段)、β-actin (下段) を WB により検出した。 (D-F) 野生型マウスの値を 1 としたときの、ADAMTS-2 ヘテロ欠損マウスと ADAMTS-2 KO マ ウスの大脳皮質 (D)、海馬 (E)、小脳 (F) における FL および NR2、Dab1 の割合を示した。 *p < 0.05、**p < 0.01.
A
B
C
D
E
F
+/+ -/-/ W T ( ra tio )+/-次に、4 ヶ月齢の ADAMTS-2 KO マウスでは、2 ヶ月齢の結果と同様、大脳皮質 と海馬においてNR2 量および Dab1 量が顕著に減少した (図 10A、10B、10D、 10E)。一方小脳では、NR2 量や Dab1 量は減少しなかった (図 10C と 10F)。以上 の結果より、ADAMTS-2 は 4 ヶ月齢マウス大脳皮質および海馬でもリーリン N-t 切断と不活化を担うことが明らかとなった。 図10 ADAMTS-2は4ヶ月齢のマウス大脳皮質および海馬でリーリンN-t切断と不活化を担う (A-C) 4 ヶ月齢の野生型マウス (+/+)、ADAMTS-2 ヘテロ欠損マウス (+/–)、ADAMTS-2 KO マウ ス (–/–) の大脳皮質 (A)、海馬 (B) および小脳 (C) におけるリーリン FL と NR2((画像上段)、 Dab1 (中段)、β-actin (下段) を WB により検出した。 (D-F) 野生型マウスの値を 1 としたときの、ADAMTS-2 ヘテロ欠損マウスと ADAMTS-2 KO マ ウスの大脳皮質 (D)、海馬 (E)、小脳 (F) における FL および NR2、Dab1 の割合を示した。 *p < 0.05, **p < 0.01, ***p < 0.001.
D
E
F
A
B
C
+/+ +/- -/-/ W T ( ra tio )4-2. 考察
ADAMTS-3 は胎生期と幼若期において、リーリン N-t 切断を担う主要な酵素 であることが報告されていた(Ogino et al., 2017)。しかし、ADAMTS-3 以外に成 体脳でリーリン N-t 切断を担う酵素は報告されていなかった。本研究により、 ADAMTS-3 と同じく ADAMTS ファミリーに属する ADAMTS-2 もリーリン N-t 切断を担うことが明らかとなった。またADAMTS-2 は、ADAMTS-3 の寄与が小 さくなる成体期においてもリーリン N-t 切断と不活化を担うことを新たに見出 した。
リコンビナント ADAMTS-2 や ADAMTS-3 を発現する HEK293T 細胞の培養 上清中には、様々な分子量の断片が検出されていた (図 5C と 5D)。ADAMTS-2 や ADAMTS-3 は、リーリンや procollagen 以外にも候補基質が報告されている (Bekhouche et al., 2016)。もし、生体内でも ADAMTS-2 や ADAMTS-3 の部分分 解が行われているのなら、それぞれの基質に対する酵素活性の ON/OFF の切り 替えや、活性の微調整を行う機構が存在するのかもしれない。
ADAMTS-2、ADAMTS-3、ADAMTS-14 のリーリン N-t 切断活性は、ADAMTS-3 > ADAMTS-2 の順に高く、ADAMTS-14 は活性が著しく低かった (図 5E)。 ADAMTS-14 のリーリン N-t 切断活性が低い理由の 1 つとして、作製した ADAMTS-14 が充分に活性化されていない可能性が考えられる。ADAMTS-2 は 自己分解を受けた後、Furin によって Prodomain-Metalloprotease domain 間で切断 を受け、活性化されることが報告されている (Colige et al., 2005)。培養上清中に 含まれているADAMTS-14 は ADAMTS-2 や ADAMTS-3 よりも分子量が大きく、 ADAMTS-14 だけが自己分解を受けていない可能性もある。そのため、使用した ADAMTS-14 が酵素活性を有しているか否かを、リーリン以外の基質を用いて検 討する必要があると考えている。
ADAMTS-2 や ADAMTS-3 のリーリン N-t 切断活性には、DI や最も N 末端側 の TS が重要であることが明らかとなった (図 6 と図 7)。先行研究より、 ADAMTS-2 や ADAMTS-3 は procollagen の切断と成熟化を担うことが知られて いる (Fernandes et al., 2001; Colige et al., 2002)。また、ADAMTS-2 の procollagen 切断活性には、N 末端側から 2 番目〜4 番目の TS が必要であると報告されてい る (Colige et al., 2005)。したがって、ADAMTS-2 や ADAMTS-3 の基質認識機構
は一様ではなく、基質によって様々であることが示唆された。現在、ADAMTS-3 の Mは一様ではなく、基質によって様々であることが示唆された。現在、ADAMTS-3 変異体を作製し、ADAMTS-2 と ADAMTS-は一様ではなく、基質によって様々であることが示唆された。現在、ADAMTS-3 のリーリン N-t 切断機構に ついて、より詳細な解明を進めている。 2 ヶ月齢および 4 ヶ月齢 ADAMTS-2 KO マウスのリーリンおよび Dab1 を解析 した結果、ADAMTS-2 が成体期におけるリーリンの N-t 切断と不活化に寄与す ることがわかった (図 9 と図 10)。一方、小脳では、NR2 量や Dab1 量は減少せ ず、むしろ増加する傾向があった。当研究室では、N-t 切断抵抗型のリーリンを 発現するPADV-KI マウスの小脳でも、大脳皮質および海馬と比較して N-t 切断 が抑制されないという結果を得ている。したがって、小脳でリーリンのN-t 切断 を担うのは全く別の酵素 (群) であると考えている。また、NR2 量や Dab1 量が 増加傾向を示したことに関しては、ADAMTS-2 の基質となる何らかの酵素が活 性化されたことで、リーリンN-t 切断が亢進したと考えている。小脳におけるリ ーリンN-t 切断酵素については報告がなく、今後さらなる解明が期待される。 大脳皮質および海馬では、4 ヶ月齢 ADAMTS-2 KO マウスは 2 ヶ月齢の ADAMTS-2 KO マウスと比較して、NR2 の割合が減少する傾向にあった (図 9 と 図10)。この結果から、ADAMTS-2 は加齢とともにリーリン N-t 切断への寄与が 大きくなる可能性が示唆された。しかし、ADAMTS-2 KO マウスでも完全に N-t 切断が抑制されなかったため、ADAMTS-2 だけでなく ADAMTS-3 も(もしく は、加えて他の酵素も)リーリンN-t 切断を担っていると考えている。したがっ て、リーリンのN-t 切断抑制およびリーリン機能亢進を標的とした精神神経疾患 の治療薬開発では、ADAMTS-2 と ADAMTS-3 の両酵素の阻害が重要であると 推察される。先述の通り、ADAMST-3 や ADAMTS-2 はリーリン以外の基質も報 告されており(Bekhouche et al., 2016)、酵素阻害剤投与による副作用発現が懸念さ れる。ADAMTS-2 や ADAMTS-3 の阻害剤開発にあたっては、ADAMTS-2 およ びADAMTS-3 の基質特異性(例えば、それぞれの基質に対する酵素反応速度や 認識機構)の詳細な解明が必要になるだろう。
第五章
総括
本研究では、アルツハイマー病の発症や進行に対して抑制的に働くことが報告 されているリーリンに着目し、その不活化酵素であるADAMTS-3 の阻害による アルツハイマー病態抑制効果の解明を目指した。また、成体脳でリーリンの N-t 切断と不活化を担う酵素の解明を目指した。そして、以下の知見を得た。 1. ADAMTS-3 を減少させれば、大脳皮質で野生型 Aβ の沈着を抑制あるいは 遅延できる。
2. ADAMTS-3 と同じく ADAMTS ファミリーに属する ADAMTS-2 はリーリン N-t 切断活性を示す。
3. ADAMTS-2 や ADAMTS-3 のリーリン N-t 切断活性には、Disintegrin domain や最もN 末端側の Thrombospondin motif が重要である。 4. 成体期では ADAMTS-3 だけでなく ADAMTS-2 もリーリン N-t 切断と不活化 を担う。 本研究はリーリンの N-t 切断のメカニズムと生理的意義の解明につながるもの であり、ADAMTS-2 および ADAMTS-3 の阻害によるアルツハイマー病の新規 治療法開発の可能性を示唆する。
引用文献
Bekhouche, M., Leduc, C., Dupont, L., Janssen, L., Delolme, F., Vadon-Le Goff, S., Smargiasso, N., Baiwir, D., Mazzucchelli, G., Zanella-Cleon, I., Dubail, J., De Pauw, E., Nusgens, B., Hulmes, D. J., Moali, C., Colige, A. (2016) Determination of the substrate repertoire of ADAMTS2, 3, and 14 significantly broadens their functions and identifies extracellular matrix organization and TGF-β signaling as primary targets. FASEB J., 30, 1741–1756.
Chen, Y., Beffert, U., Ertunc, M., Tang, T. S., Kavalali, E. T., Bezprozvanny, I., Herz, J. (2005) Reelin modulates NMDA receptor activity in cortical neurons. J. Neurosci., 25, 8209–8216.
Colige, A., Vandenberghe, I., Thiry, M., Lambert, C.A., Van Beeumen, J., Li, S. W., Prockop, D. J., Lapiere, C. M., Nusgens, B.V. (2002) Cloning and Characterization of ADAMTS-14, a Novel ADAMTS Displaying High Homology with ADAMTS-2 and ADAMTS-3. J. Biol. Chem., 277, 5756–5766. Colige, A., Ruggiero, F., Vandenberghe, I., Dubail, J., Kesteloot, F., Van Beeumen, J.,
Beschin, A., Brys, L., Lapière, C. M., Nusgens, B. (2005) Domains and maturation processes that regulate the activity of ADAMTS-2, a metalloproteinase cleaving the aminopropeptide of fibrillar procollagens types I-III and V. J. Biol. Chem., 280, 34397–34408
D'Arcangelo, G., Homayouni, R., Keshvara, L., Rice, D.S., Sheldon, M., Curran, T. (1999) Reelin Is a Ligand for Lipoprotein Receptors. Neuron, 24, 471–479. D'Arcangelo, G., Miao, G. G., Chen, S. C., Soares, H. D., Morgan, J. I., Curran, T. (1995)
A protein related to extracellular matrix proteins deleted in the mouse mutant reeler. Nature, 374, 719–723.
Durakoglugil, M. S., Chen, Y., White, C. L., Kavalali, E. T., Herz, J. (2009) Reelin signaling antagonizes beta-amyloid at the synapse. Proc. Natl. Acad. Sci. U. S. A., 106, 15938–15943.
Elkins, M. R., Wang, T., Nick, M., Jo, H., Lemmin, T., Prusiner, S. B., DeGrado, W. F., Stöhr, J., Hong, M. (2018) Structural Polymorphism of Alzheimer’s β-Amyloid Fibrils as Controlled by an E22 Switch: A Solid-State NMR Study. J. Am. Chem. Soc., 138, 9840–9852.
Fernandes, R. J., Hirohata, S., Engle, J.M., Colige, A., Cohn, D.H., Eyre, D.R., Apte, S.S. (2001) Procollagen II Amino Propeptide Processing by ADAMTS-3. Insights on dermatosparaxis. J. Biol. Chem., 276, 31502–32509.
Hoe, H. S., Tran, T. S., Matsuoka, Y., Howell, B. W., Rebeck, G. W. (2006) DAB1 and Reelin effects on amyloid precursor protein and ApoE receptor 2 trafficking and processing. J. Biol. Chem., 281, 35176–35185.
Kocherhans, S., Madhusudan, A., Doehner, J., Breu, K. S., Nitsch, R. M., Fritschy, J. M., Knuesel, I. (2010) Reduced Reelin expression accelerates amyloid-beta plaque formation and tau pathology in transgenic Alzheimer's disease mice. J. Neurosci. 30, 9228–9240.
Kohno, S., Kohno, T., Nakano, Y., Suzuki, K., Ishii, M., Tagami, H., Baba, A., Hattori, M., (2009) Mechanism and significance of specific proteolytic cleavage of Reelin. Biochem. Biophys. Res. Commun., 380, 93–97.
Kohno, T., Honda, T., Kubo, K., Nakano, Y., Tsuchiya, A., Murakami, T., Banno, H., Nakajima, K., Hattori, M., (2015) Importance of Reelin C-terminal region in the development and maintenance of the postnatal cerebral cortex and its regulation by specific proteolysis. J. Neurosci., 35, 4776–4787.
Lambert de Rouvroit, C., de Bergeyck, V., Cortvrindt, C., Bar, I., Eeckhout, Y., Goffinet, A. M. (1999) Reelin, the extracellular matrix protein deficient in reeler mutant mice, is processed by a metalloproteinase. Exp. Neurol., 156, 214–217.
Lane-Donovan, C., Philips, G. T., Wasser, C. R., Durakoglugil, M. S., Masiulis, I., Upadhaya, A., Pohlkamp, T., Coskun, C., Kotti, T., Steller, L., Hammer, R. E., Frotscher, M., Bock, H. H., and Herz, J. (2015) Reelin protects against amyloid β toxicity in vivo. Sci. Signal., 8, ra67.
Lee, G. H., D'Arcangelo, G. (2016) New Insights into Reelin-Mediated Signaling Pathways. Front. Cell. Neurosci., 10, 122.
Li, S. W, Arita, M., Fertala, A., Bao, Y., Kopen, G. C., Långsjö, T. K., Hyttinen, M. M., Helminen, H. J., and Prockop, D. J. (2001) Transgenic mice with inactive alleles for procollagen N-proteinase (ADAMTS-2) develop fragile skin and male sterility. Biochem. J., 355, 271–278.
Matsuki, T., Matthews, R. T., Cooper, J. A., van der Brug, M. P., Cookson, M. R., Hardy, J. A., Olson, E. C., Howell, B. W. (2010) Reelin and Stk25 Have Opposing
Roles in Neuronal Polarization and Dendritic Golgi Deployment. Cell, 143, 826–836.
Niu, S., Renfro, A., Quattrocchi, C. C., Sheldon, M., D'Arcangelo, G. (2004) Reelin promotes hippocampal dendrite development through the VLDLR/ApoER2-Dab1 pathway. Neuron, 41, 71–84.
Ogino, H., Hisanaga, A., Kohno, T., Kondo, Y., Okumura, K., Kamei, T., Sato, T., Asahara, H., Tsuiji, H., Fukata, M., Hattori, M. (2017) Secreted Metalloproteinase ADAMTS-3 Inactivates Reelin. J. Neurosci., 37, 3181–3191.
Pujadas, L., Gruart, A., Bosch, C., Delgado, L., Teixeira, C. M., Rossi, D., de Lecea, L., Martínez, A., Delgado-García, J. M., Soriano, E. (2010) Reelin regulates postnatal neurogenesis and enhances spine hypertrophy and long-term potentiation. J. Neurosci., 30, 4636–4649.
Pujadas, L., Rossi, D., Andrés, R., Teixeira, C. M., Serra-Vidal, B., Parcerisas, A., Maldonado, R., Giralt, E., Carulla, N., Soriano, E. (2014) Reelin delays amyloid-beta fibril formation and rescues cognitive deficits in a model of Alzheimer's disease. Nat. Commun., 5, 3443.
Rice, H. C., Young-Pearse, T. L., Selkoe, D. J. (2013) Systematic Evaluation of Candidate Ligands Regulating Ectodomain Shedding of Amyloid Precursor Protein. Biochemistry, 52, 3264–3277.
Rogers, J. T., Rusiana, I., Trotter, J., Zhao, L., Donaldson, E., Pak, D. T., Babus, L. W., Peters, M., Banko, J. L., Chavis, P., Rebeck, G. W., Hoe, H. S., Weeber, E. J. (2011) Reelin supplementation enhances cognitive ability, synaptic plasticity, and dendritic spine density. Learn. Mem., 18, 558–564.
Saito, T., Matsuba, Y., Mihira, N., Takano, J., Nilsson, P., Itohara, S., Iwata, N., Saido, T. C. (2014) Single App knock-in mouse models of Alzheimer’s disease. Nat. Neurosci., 17, 661–663.
Teixeira, C. M., Kron, M. M., Masachs, N., Zhang, H., Lagace, D. C., Martinez, A., Reillo, I., Duan, X., Bosch, C., Pujadas, L., Brunso, L., Song, H., Eisch, A. J., Borrell, V., Howell, B. W., Parent, J. M., Soriano, E. (2012) Cell-Autonomous Inactivation of the Reelin Pathway Impairs Adult Neurogenesis in the Hippocampus. J. Neurosci., 32, 12051–12065.
謝辞 本研究を遂行するにあたり、日々厳しくも、熱心に御指導と御鞭撻を賜り、 そして研究室にあたたかく迎え入れてくださいました名古屋市立大学大学院薬 学研究科、服部光治教授に心より深謝致します。 本論文の作成にあたり、多大な御指導と有益なる御助言を賜りました名古屋市 立大学大学院薬学研究科、松永民秀教授、頭金正博教授、ならびに田中正彦准 教授に深く感謝申し上げます。 本研究を遂行するにあたり、日頃から細やかに御指導して頂きました名古屋市 立大学大学院薬学研究科、河野孝夫講師に心より御礼申し上げます。 本研究を遂行するにあたり、的確な御助言とご協力を賜りました名古屋市立大 学大学院薬学研究科、築地仁美講師に厚く御礼申し上げます。 本研究を遂行するにあたり、AppNL-FマウスおよびAppNL-G-Fマウスを御供与いた だきました理化学研神経老化制御研究チーム、西道隆臣チームリーダー、なら びに斉藤貴志副チームリーダーに心より感謝致します。 本研究を遂行するにあたり、ADAMTS-2 KO マウスを作製していただきました 名古屋市立大学大学院医学研究科、大石久史教授、ならびに宮本智美先生に深 謝致します。 HEK293T 細胞を恵与していただきました、理化学研究所脳科学総合研究セン ター、御子柴克彦チームリーダーに厚く御礼申し上げます。 5 年間の研究生活を共に過ごし、苦楽を分かち合いました名古屋市立大学大学 院薬学研究科、病態生化学分野の皆様に心より御礼申し上げます。 最後に、いつも研究生活を支え、応援してくださいました両親に心より感謝申 し上げます。