ツルボの胞子及び配偶体の形成とカロースの消長
10
0
0
全文
(2) 佐藤嘉彦・松元啓介 (ヒアシンス科)にツルボ属を収. スパラガス目)を設け,この目の-科Hyacinthaceae 容する研究者(HuBER. 1969等)もいる。ユリ科(広義)の発生学的な調査はこれまでに. 数多くなされてきた。ツルボ属は発生学的にはたいへん興味深い属で,この属にはEn_ dymion. typeに従って匪嚢が形成される種が含まれている(DAVIS. 1966. ;. DAHLGEN. &. CLIFFORD1982)。この腫嚢形成の様式は,少なくともユリ科ではツルボ属の他には,ユ キザサ(Smilacina)属とAgraphis属及びEndymion属でしか知られていないものであ & CLIFFORD1982)0. る(DAHLGEN. 被子植物の小胞子形成過程と大胞子形成過程でのカロース(Callose)の存在が,それ ぞれWATERKEYN. (1962)とRoDKIEWICZ. 良 G6RSKA-BRYLASS. (1967)によって示され. てから,胞子や配偶体の形成過程でのカロースの存在様式の調査はときおり行われてきた。 しかし調査された植物はまだ少数にすぎない。カロースはP-1,. 3結合によるグルマー. ス(glucose)の重合体であるが,これが細胞壁に沈着すると高分子物質に対する壁の透過 性は減少することが実験的に示されている(HESLOP・HARRISON. 1966)。カロース性の壁. をもっ細胞は周囲を囲む細胞と情報の交換が完全ではないが,ある程度制限されることに なると考えられる。それ故にカロースの壁-の沈着ほ,ある特定の細胞を周囲にある細胞 群から孤立化させる働きをもっと考えられる。つまりこの現象は細胞や組織の分化と何ら かの関係を持っものと推論される(SAT古. 1986)。また,大胞子母細胞が行う減数分裂の. 2回の分裂の過程や,その終了後にみられるカロースの存在様式には よる相違がある(KAPIL&TIWARI. 腫嚢形成の様式に. 1978)と思われる。そればかりでなく,被子植物. の腫嚢形成様式の中ではもっとも普遍的な単胞子性8核タデ型の中でも変異が認められる (SAT6. & ARIMA. 1984. ;SAT6. &. TAKEDA. 1985)。. そこで筆者等はツルボ(Scillascilloides. (LINDL.)DRUCE)を材料として,その胞. 子と配偶体の形成様式及びその過程でみられるカロースの壁-の沈着様式についての調査 を行ってきた。この論文ではこの調査で得られた結果について報告する。. 材料及び方法 1984年の8月から9月に,横浜国立大学の構内に自生しているツルボから多数の曹と花 を採り,. F AA. (Formalin-Acetic-Alcohol)で固定した。胞子及び配偶体の形成過程. を調査するために,固定した材料を第三ブ≠ルアルコール(te,i. butyl. alcohol)とエ. チルアルコール(etbyl に包埋し,. alcohol)のシリーズで脱水した後にパラフィン(paraffin) 6-8〟mの厚さの連続切片を作った。この切片を-イデン-イン氏の鉄ミョ. ウバン.へマトキシリン(HEIDENHAIN's ーン(fast ニリン. iron. alum. bematoxylin)とファースト. green)で重染した。カロースの検出にあたっては,同様に作成した切片をア ブルー(aniline blue)の水溶液で染色し( SMITH&McCuLLY. グリ. 1978),蛍. 光顕微鏡下で観察した。アニリンブルーで処理するとカロースは淡黄色の蛍光を発し, 微量でも容易に検出できる。.
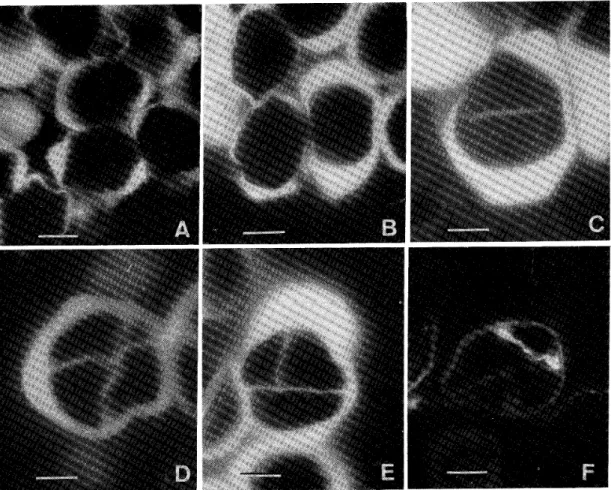
(3) ツルボの胞子及び配偶体形成 観. 察. 胞子及び配偶体の形成 1.雄性配偶体の形成 薪壁。小胞子母細胞が減数分裂を始める頃には,薪壁は4細胞層で構成されている。そ layer)及びじゅうたん細. れは表皮(epidermis),内被(endotbecium),中間層(middle. 胞層(tapetum)である。減数分裂を終了する頃には中間層はなくなり,じゅうたん細胞 も多核になっている。荊が裂関する直前にはじゅうたん細胞層もなくなり,東壁は表皮細 胞層と内被おのおの1層となる。表皮細胞は扇平な形に変形しており,内被細胞の壁には 線状肥厚(fibrous. thickening)が発達している。 1 A)の多くは,減数分. 小胞子及び花粉の形成。荊内に分化した小胞子母細胞(Fig.. 裂第一分裂に入る頃には,やや長球形になっている。第一分裂の紡錘体はこの長軸に沿っ. 小胞子母細胞は分裂して, (Fig.. 1. 1 B)。第一分裂の進行にともない細胞板が形成され(Fig・. て作られる(Fig.. 2個の小胞子二分子細胞になる。二分子は減数分裂第二分裂. 1 D)を経て小胞子四分子になる。第二分裂でも細胞板が形成され,細胞質の分. 裂がおこる。四分子には,二分子の一方の細胞に作られた隔壁がもう一つの細胞の隔壁に 並行なもの(Fig.. Fig.. 1E)と直角なもの(Fig.. 1. Formation. 1F)がある。その後四分子細胞はばら. in Scilla scilloides. C. Microsporocyte which nearly of meiosis. first division D. Second division of meiosis. E. Four the completes of meiosis. decussately. F. Four isobilaterally. tetrad cells, arranged tetrad cells, arranged G. Young in first division has the the just been pollen grain, which of microspore of microspore. A. Microsporocyte.. H.. over.. of. the. Mature. vegetative. B.. pollen cell.. and. First. pollen. division. grain. ;. (scale. bar. the. generative. -lo〟m). cell. is. surrounded. by the. cytoplasm. C),.
(4) 4. 佐藤嘉彦・松元啓介 ばらに離れ,それぞれが小胞子としての機能を持つ。 つまりツルボの小胞子は漸次膜形成型(successive type)に従って形成される。 ばらばらになった小胞子は大形の栄養細胞と小 形の生殖細胞に分裂し,二細胞性の花粉に発達す る。生殖細胞は形成直後には花粉の壁に接してい る(Fig.. 1G)が,紡が裂関する前に花粉の壁か. ら離れ,栄養細胞の細胞質の中に遊離する(Fig. 1 H)。花粉は2細胞の状態で柱頭に運ばれる。. 2.雌性配偶体の形成 腫珠o雌蕊は3枚の心皮で構成され,子房は完全 に3室に分かれている。その各室には1個の腫珠が生 じる。大胞子母細胞が減数分裂を終わった時には腫珠 はほぼ倒生の状態になっており,膝嚢が完成した時 には完全な倒生の状態に達する。突起として生じた Micropylar. Fig.2.. and egg. stropbiole. II, ;. Micropyle. S,. Details Inner OI,. ;. :. F,. ;. integument. Strophiole.. が分化する。胞原細胞は数回体細胞分裂を行い,. Fertilized. integument Outer. 若い膝珠の先端の表皮下に,通常は1個の胞原細胞. ovule. of. region. M, ;. 数個の壁細胞(parietal. cells)と1個の大胞子 母細胞を作る。すなわち,ツルボは厚層型の珠心. (crassinucellate. nucellus)を持っている。この珠心を2枚の珠皮が覆うが,外珠皮は 内珠皮よりも遅れて分化する。大胞子母細胞が減数分裂を終える頃には,両珠皮とも珠心. を覆う程度にまで伸長している。しかし,珠孔は内珠皮だけで形成される(Fig.. 2)0. 両珠皮の構造ははとんど例外なく先端部を除いて,内珠皮は2細胞層で,外珠皮は2-3 細胞層で構成されている。外珠皮の先端部は減数分裂終了後に,. stropbioleと呼ばれる大 2)を作り始める.珠心の表皮系は腫嚢が完成しても残っており. きな塊状の肥厚(Fig. (Figs.. 2,. 4C),特に珠心先端部の表皮細胞は並層分裂を行い,. 2-3層に発達して. いることが多い。 大胞子及び腫嚢の形成。胞原細胞の分裂によって作られた大胞子母細胞(Fig. は減数分裂第一分裂(Fig.. 3B)を行い,大胞子二分子(Fig.. 分子は続いて減数分裂第二分裂を行い大胞子四分子(Fig.. 3 A). 3C)になる.この二 3 D)になる。. 4個の四分子. 細胞は常に線状に配列している。四分子を構成している4個の大胞子は珠孔側のものから つぶれる(Fig.. 3. E)が,最も合点側のものはつぶれずに機能を持ち腫嚢に発達する。. この機能を持った大胞子は珠心の軸にそって伸長し,その核は細胞質の分裂をともなわず に分裂し,. 2核の膝嚢(Fig.. 4A)になる。この2核は腫嚢の両極にそれぞれ移動し,. 2核の間には大きな液胞が発達する.腫嚢はさらに伸長し,その両極で核はそれぞれ核分 裂を行い,. 4核の腫嚢(Fig.. 4B)が作られる。各極にある2核はそれぞれもう一度分. 裂し,両極にそれぞれ4核を持つ8核の腫嚢が作られる.珠孔側の4核のうちの3核は卵 装置を構成する卵細胞と2個の助細胞のそれぞれの核になり,合点側の4核のうちの3核.
(5) 5. ツルボの胞子及び配偶体形成. Fig.. 3. Megasporogenesis. in Scilla. A・. B・. Megasporocyte・. just. finished. division tetrad. bar. of cells・. the. first. scilloides.. division. micropylar. C. Two dyad cells, which have of meiosis・ D. Four The meiosis, tetrad cells. second dyad finished yet. E. Four cell is not quite to degenerate (scale of tetrad begin. division of. in themicropylar. meiosis Two. First. cells. already.. -lo〟m). は3個の反足細胞の核になる。各極の残りの核はそれぞれ中央細胞に移動し,. 2個の極核 4C)の卵. になる.この2核は受精前に融合して1核になる.この完成した腫嚢(Fig. 装置は小形であるが,反足細胞は3個とも大きく発達し,反足装置は腫嚢全体の1/4か ら1/3の大きさを占める。このようにツルボの膝嚢は単胞子性8核タデ型の発生様式に 従って形成される。. カロースの消長 1.雄性配偶体の形成 小胞子母細胞が減数分裂第一分裂に入り,細糸朔(1eptotene. stage)を過ぎる頃にな. ると,小胞子母細胞の長軸の極付近の細胞壁にカロース性の蛍光が初めて認められるよう になる(Fig.. 5A)。第一分裂の進行にともない蛍光を発する範囲は拡大し,第一分裂. が終わる前には小胞子母細胞の壁全体にカロースが沈着する(Fig.. 5 B)。このカロース. は減数分裂が終了するまで存続する。小胞子二分子を分けている隔壁にも,壁の形成後た だちにカロースの沈着が始まる(Fig.. 5. C)。減数分裂第二分裂の後に,小胞子二分子. の各細胞に作られる隔壁にもカロースが沈着し,小胞子四分子の各細胞は完全にカロース 性の壁で覆われる(Fig.. 5D,. E)0. ばらばらになった小胞子からは,まったくカロースは検出できない。しかし小胞子が栄 養細胞と生殖細胞に分裂すると,この2細胞を分けている壁には,一時的ではあるが,か.
(6) /h). 佐藤嘉彦・松元啓介. Fig.. 4. Megagametogenesis A.. Nucellus. sac.. C.. with Nucellus. in Scilla two-nucleate with. mature. scilloides. embryo embryo. sac.. B.. Nucellus. with. four-nueleate. embryo. sac.. ならずカロース性の蛍光がはっきりと観察される(Fig.. 5. F)。生殖細胞が花粉の壁か. ら離れて栄養細胞の中に遊離すると,生殖細胞の壁からカロースを検出することはできなく なる。その後少なくとも花粉が前の中にある間は,花粉の中にカロースの存在を認めるこ とはできない。 2.雌性配偶体の形成 大胞子母細胞が減数分裂に入る前には,それ自身の壁からも,珠心を構成している他の どの細胞の壁からも,カロースを検出することはできない。大胞子母細胞が減数分裂第一 分裂に入り,細糸期に達すると,大胞子母細胞の合点側の細胞壁からカロ`」スがまず検出 される(Fig.. 6A)。第一分裂の進行とともにカロースの沈着の範囲は拡大し,この分. 裂の中期(metaphase)になると大胞子母細胞の壁全体にカロースが沈着する(Fig. B)。このカロースは減数分裂の2回の分裂が終わるまで存続している。大胞子二分子細胞 を分けている隔壁にも,形成と同時にカロースは沈着する(Fig.. 6C)。減数分裂第二. 分裂の後で二分子細胞のそれぞれに作られた隔壁からも,カロースは検出される(Fig. 6 D)。結局大胞子母細胞,二分子細胞及び四分子細胞はそれぞれ完全にカロース性の壁. で囲まれていることになる(Figs.. 6D,. E)。四分子の珠孔側の3細胞が消え最も合. 点側の大胞子が伸長しはじめると,その大胞子からカロースはまったく検出できなくなる。 以降どの時期の腫嚢からもカロースの存在を示す反応を観察することはできないo. 6.
(7) ツルボの胞子及び配偶体形成. Fig.. 5. Deposition A.. tetrad F.. of. callose. Microsporoeyte. Cells,. Young. (scale. bar. B.. arranged. pollen. grain,. during microsporogenesis in Scilla scilloides. Mierosporocyte in division. D. Four C. Two dyad cells. decussately. tetrad E. Four isobilaterally. cells, arranged in wbicb. the. first. division. of microspore. has. just. finished.. -lo〟m). 考 ツルボの外珠皮の先端には でも知られている(DAⅢLGEN. 察. strophioleが作られるが,この構造はツルボ属の他の種 良 CLIFFORD. 1982)。この構造は単子葉植物だけでなく,. 双子葉植物でも知られているが,これが発達する分類群は多くはない。それ故に. stro-. phioleはツルボ属を特徴付ける構造の一つと考えてよいであろう。 ツルボの珠心には壁細胞(parietal. cells)が作られ,大胞子及びその形成過程の細 胞や発達中及び完成した腫嚢の側部には,珠'L、を作る栄養細胞が存在している.さらに 珠}L、の表皮細胞は並層分裂を行い数層に発達する。. FAVRE_DucIIARTRE. (1984)は裸子. 植物の珠心や被子植物の前の構造と被子植物の珠心の構造を比較して,種子植物の珠心は 単純なものはど進歩的なものであると系統学的な考察を行っている。多くの分類学者がツ.
(8) 8. 佐藤嘉彦・松元啓介. __∴_ Fig・. 61 Deposition A・. of callose B・. Megasporocyte.. tetrad ce11s・ already.. E・ Four. (scale bar. :_十=_. during. in Scilla. megasporogenesis. Megasporocyte. tetrad ce11s・. in division・. Micropylar. two. C・. cells. Two. scilloides.. dyad. of tetrad. D.. Cells,. begin. Four. degenerate. to. -lo〟m). ルポ属を収容しているユリ科の珠心はかなり多様性に富んでいる。. FAVRE_DロCEARTRE. (1984)によって提唱された珠心構造の系統学的な考察が正しいならば,ツルボの珠心は かなり原始的なものとみなすことができる。. 雄性配偶体の形成及びこの過程でのカロースの沈着様式は,単子葉植物に一般的に見ら れるものと変わりはないo. しかし雌性配偶体の形成及びこの過程でのカロースの沈着様式. については議論すべき点がある。 ツルボ属では,腫嚢がEndymion いる(DAVIS. 1966. ;. typeの発生様式に従って形成される種が知られて. DAHLGEN&CLIFFORD. 1982)。この発生様式は. Scillatype. と呼ばれることもあり,二胞子性の発生様式の一つである。二分子細胞のうち,合点側の ものは多核の状態に発達することはあるが,腫嚢を完成させることはなく,珠孔側のもの だけが完全な腫嚢にまで発達するo. しかし,ツルボの腫嚢は単胞子性8核タデ型の発生様. 式に従って形成され,二分子細胞のうちの一つがつぶれたり,四分子細胞のうちの二つが 発達を始めると思われる現象を観察することはできなかった。 KAPIL&TIWARI. (1978)によると,単胞子性の発生過程ではカロースは大胞子母. 細胞及び二分子細胞や四分子細胞を構成する各細胞の壁の大部分に沈着するが,壁によっ ては部分的にカロースが沈着しないこともある。また彼等によると,二胞子性の発生過程 ではカロースの沈着は二分子細胞を分ける隔壁だ桝こ限られる。. SAT6. &. TAKEDA. は単胞子性の発生様式にもかかわらず二分子細胞を分ける隔壁と四分子細胞を分ける3枚 の隔壁にしかカロースが沈着しない例を報告し,さらにこれは二胞子性の発生様式と関連 するものであろうと推論している。ツルボでは,大胞子母細胞及び二分子細胞や四分子細. (1985).
(9) 9. ッルポの胞子及び配偶体形成 胞を構成している各細胞が完全にカロース性の壁で覆われており,カロースの沈着しない 部分は存在していない。これとまったく同じカロースの沈着様式は,単胞子性8核タデ型 &ARIMA. の腫嚢形成を行うヤプラン(ユリ科一広義)でも知られている(SAT6. 1984)。カ. ロースの沈着様式の変異が腫嚢形成様式とどの様に関連を持つかについては,やっと知識 が集り始めたところであるが,ツルボの腫嚢形成過程でのカロースの沈着様式には二胞子. 性の腫嚢形成過程での沈着様式との共通性を,いままでのところでは見出すことはできな い。. このように,雌性配偶体の形成過程及びこの過程でのカロースの沈着様式から,ツルボ の腫嚢形成は単胞子性であり,二胞子性との関連を議論できる情報は何もないと思われる。. ツルボ属は単胞子性の勝義形成を行うものと二胞子性のものが混在している分類群である といえる。 膝嚢の反足装置の構造の多様性についてはよく知られている(. MAI7ESIWARI. 1950. ;. DAVIS. DAVIS(1966)によってまとめられたユリ科の発生学的な特徴の記述には,反足. 1966)。 装置は,. Scillanonscriptaでは細胞化せず3核で構成されるが,. S.pratensis. では. 3細胞で構成され,さらに各細胞は著しく伸長すると記されている.ツルボは. S.pra・. tensisと同じ反足装置をもつと思われる。またこれと同じ反足装置はユリ科(広義)の中 でもツルボ属とはあまり類縁の近くないと思われるホトトギス(Tricyrtis)属で一般的. に見られるものである(OGURA. 1964,1966)。このように,ツルボ属は腫葉形成様式ばかり. でなく腫嚢の構造に関しても,系統学的に大変興味深い分類群である。. 参 CRONQtJ[ST,. A., 1981 Press,. versity DAⅢ」GEN,. M.. Press, DAVIS,. An. T.. New. 良 H.. G. L., 1966. angiosperms.. H., 1969. CLIFFORD,. Systematic. of flowering. of classification. 1982. The. :. monocotyledons. pp.. 697-734.. :. Columbia. plants.. Uni-. a. comparative. study.. Academic. scopy. :. an. MAEESHWARI,. and. Cytoplasmic. Wiley,. angiosperms, In. phylogeny.. New. M.. B.. JoHRl. York.. (ed.),Embryology. of. Berlin.. Springer・Verlag, continuities. Samenmerkmale Mihchen. J., 1973 R.. Homologies. during. formation. spore. in. flowering. plants.. 65172.. Die. N. A. of the. embryology. 1984. Slaatssamml.. HuTCHTNSON,. KAPIL,. T.. M.,. 25. Endeavour. R.. 献. York.. I., 1966. HESLOP・HARRISON,. Bol.. system. 文. London.. FAVRE・DucHARTRE,. HuBER,. integrated. 考. The. N.. 8. families. T[wARl,. of flowering. 1978. Plant. An. P., 1950. der. introduction. Oxford. plants.. at. the. Liliiforae.. Clarenson. investigations embryological and Inter. Rev. Cytol. 53 : 291-331.. of integration.. assessment. Verwandtschaftsverh註1tnisse. und. MillerL.. 219_538.. :. to. the. embryology. of. angiosperms.. Press,. fluorescence. McGraw・Hill,. Oxford. microI. New. York. OGURA,. H., 1964. (Biol.) 30 OGURA,. H.,. :. 1966. 0n. the embryo. of two. sac. species. of Tricyrlis.. Sci, Rep.. To^hoku. Univ.. Ser.. 219-222. 0n. the・. embryo. sac. of. Tricyrtis. macranlhopsis. Masam.. Sci.. Rep.. T∂hoku. IV.
(10) 佐藤嘉彦・松元啓介. 10. Univ,. R.. RoDOKIEWICZ, dividing Y". SAT6,. Sec. Y.. SAT6, et. IV. Ser.. (Biol.) 32. 良 A. G6RSKA・BRYLASS, in the. cell. 1986. Embryo. II. 33. :. 良 T.. 1984. Sieb.. praecox. M・. SM-I,. M.. a. 46. :. WAKERKEYN,. E.. Slain A.,. 0ccurrence. of. of. Pittosporum. in the walls. callose. Nalurwissenschaften. Development. 1980. 59. lobira.. 54. Sci.. of meiotically. 499-500.. :. Repts.. Yokohama. Nail.. Univ.. of. Zucc.. of callose. Sci. Repts.. McCtJLLY,. 53. of. the. Yokohama. Enhancing. 1978. Technology. 0utline. spore. and. in. gametophyte. Liriope. Wang. platyphylla. 116-128.. :. Occurrence. 1985 et. MI. structures.. TAKHTAJAN,. formation. sac. Jour. Jar. Bol.. Y・ 良 K・ TAKEDA,. SAT6,. 1967. of Orchis.. ovule. (in press). AR[MA,. Tang.. 31-34.. :. :. during. Nail. Univ. aniline. sac. embry_o. blue. Sec.. development. 1Z. 32. fluorescent. :. in Stachyurus. 35144. staining. of. cell. wall. 79-85.. classification. of flowering. plants. Bot.. Rev.. Helloborous. et. (Magnoliophyta).. 225_359. L.,. Tradescanlia.. 1962. Les Cellule. parois 62. :. microsporocytaires 225_255.. de. nature. callosique. chez.
(11)
図



+2

関連したドキュメント
内部構造(Fig.3-D2-4, Plate 2):花被の腺毛(D2)は(7. virgatumのものと同様で,頭細胞は球形または軸方向
しかしながら生細胞内ではDNAがたえず慢然と合成
の多くの場合に腺腫を認め組織学的にはエオヂ ン嗜好性細胞よりなることが多い.叉性機能減
1)血管周囲外套状細胞集籏:類円形核の単球を
「サントリー天然水」は、大容量及び小容量(500ml
本装置は OS のブート方法として、Secure Boot をサポートしています。 Secure Boot とは、UEFI Boot
副校長の配置については、全体を統括する校長1名、小学校の教育課程(前期課
体長は大きくなっても 1cm くらいで、ワラジム シに似た形で上下にやや平たくなっている。足 は 5
