九州大学学術情報リポジトリ
Kyushu University Institutional Repository
アレルギー関連因子の評価法の開発とその利用に関 する研究
高杉, 美佳子
九州大学農学研究科食糧化学工学専攻
https://doi.org/10.11501/3135077
出版情報:Kyushu University, 1997, 博士(農学), 課程博士 バージョン:
権利関係:
の
アレルギ一関連因子の評価法の開発と その利用に関する研究
高 杉 美 佳 子
199 8
アレルギ一関連因子の評価法の開発とその利用に関す る 研究
目 次
第1章
緒論
.…・・…・・・…..… ・・…・・…・・……・・・…-…一…H・H・-…・・一……… H ・ H ・ - … . . . .1 第2章 ラットおよびヒトにおけるアレルゲン特異的および
非特異的抗体の分析 . . . . ・H・-……. . . . ・H・....・H・ . . . …・… . . . .……一一…………・….
5
第 3 章 アレルゲン特異的抗体発現のクラスおよびサブクラス依存性 .…・.21
第4章 MAST法を用いたアレルゲン特異的19Aおよび�IgG定量法の開発 …ー…一. . . ・H・..…・. . . .. . . . .. . . .. . . ..
3 4
第5章 ヒト血清および唾液中のアレルゲン特異的および非特異的抗体発現パターンの解析 . . . . ・ H ・ . . . ... ... . . .. . . ... . .
44
第6章 食品のアレルゲン活性評価法の検討 . . . ・H・..…・・……・………. . . ・ H ・ . . . ・ H ・ - …
60
第7章 マウスリンパ球を用いた免疫調節機能因子検定系の開発 ・……・…・・. 71 第8章 マウス牌臓細胞の抗体産生に及ぼす食品成分の影響 … . . … . . . .. . . ・ H ・ . . . 81 第9章 総括 .…・・・……一…一一……一一…… . . . .. . . ..... ... . ... . . ... . . .. . . . ..
101
文献
.…-…・・・・……H・H ・ - … ー ・…・・・……一……...・H・...・H・ -…・・・・………110
第1章 緒論
近年、 わが国では住環境の変化に加え、 食生活の内容が豊かになり多様化し てきた。 このことは小児の食生活においても同様で、 乳児の離乳の早期化、 乳 幼児および小児の高栄養化などが食物アレルギー患者増加の原因の一部と考え られている。 また、 乳幼児や小児の食物アレルギーが原因で成人後も食物や環 境因子に対してアレルギー症状を示す者が多いため、 食物アレルギーは小児だ けでなく成人にとっても切実な問題となっている(Casimir
et
a1., 1993)。 アレル ギーは免疫機能の過剰反応によって引き起こされるが、 Coombs and Gell ( 1968) はアレルギ一反応を四つのタイプに分類しており、 食物アレルギーや環境アレ ルギーはI型アレルギーによるものが多い。 1型アレルギーは、 マスト細胞や好 塩基球表面に結合したアレルゲン特異的IgEがアレルゲンにより架橋され、 ヒ スタミンやロイコトリエンなどのケミカルメディエーターが放出されることに より誘導される(Metcalfe, 1991)。 血清中のIgE濃度は、 健常人では非常に低い が、 アレルギー患者においてしばしば高濃度で検出される(Bjorkstenet
a1.,1983)。 したがって、 血清中の総IgEおよびアレルゲン特異的IgEの測定は食物 アレルギーの発症機構を理解する上で必要不可欠であると考えられる。
IgEの測定にはradioaller gosorbent test (RAST) およびその応用法(Adler
et
a1.,1991; William
et
a1., 1992) や皮膚受身アナフィラキシー試験(PCA) が用いられて いる(Ovary 1958; Sed gwick and Holt, 1983)。 これらの方法はIgE検出のための感 度は充分であるが、 RASTは放射性同位元素を使用するための特別な施設が必 要であり、 PCAは定量性に欠け、 一度に多くのサンプルを分析することが困難 である。 酵素抗体法(enzyme-linked immunosorbent assay ; ELISA) はこれらの方 法の代替法としてヒト、 マウスおよびラットIgEの検出にも利用されている(Doke et a1., 1989; Fehlner et a1., 1991; Fritsche and Bonzon, 1990)。 しかし、 通常
のELISA法の感度では抗原特異的IgEの検出には不十分である。 そこで、 より
検出感度の高いアビジン-ビオチン法を用いてIgEの定量を行い、 アレルゲン
特異的IgEの発現について解析した。
IgEだけでなくその他のクラスの抗体もアレルギ一応答に関与している可能性 がある。 19Aには二つのサブクラスがあり、 血清中だけでなく、 分泌型19A (sIgA) は腸管粘膜の分泌液や唾液、 涙にも合まれており(Watanabe
et
a1.,1984)、 腸管粘膜中の s1gAは腸管からのアレルゲンの吸収を抑制することにより アレルギ一反応を抑制する(Shorter and Tomasi, 1982)0 IgGは血中に最も多量に 合まれている抗体であり、 アレルギーの減感作療法により誘導される抗体でI型 アレルギーの原因抗体である19Eと競合することによりアレルギー抑制的に働 いたり(Aalbers e
et
aJ., 1993)、 終生免疫の獲得に関与している。 19Gには四つの サブクラスが存在しており、 補体の活性化能やナチュラルキラー細胞の活性化 能に若干の違いが認められるが(Janewa)ら1996)、 機能の違いについては不明な 点も多い。 19 M は初期免疫に関与する抗体であり、 抗原感作を受けると一番最 初に産生される。 これらの抗体はアレルギーの発症および抑制に関与している と考えられるが、 抗体の発現をクラス別に総合的に解析したものはほとんどな い。 そこで、 本研究では健常人およびアレルギー患者血清中のアレルゲン特異 的および非特異的抗体の発現パターンを解析した。 また、 腸管分泌液中の19A 発現に関する情報も非常に限られており、 腸管内のアレルゲン特異的19Aの発 現を調べることは困難である。 そこで、 ELISAによる唾液19A定量法を確立し、唾液中の19A発現を解析することにより腸管免疫系の活性の推測を行った。
M ultiple ant ig en simultaneous tes t (MAST) は多項目のアレルゲン特異的19E の同時定量が可能な方法であり、 RASTの代替法として利用されている(Agata
et al., 1993; Brown et al., 1985; Miller et al., 1984)。 そこで、 多くのアレルゲンに 対する抗体の発現を同時に解析するため、 MAST法を用いた血清および唾液中 のアレルゲン特異的19Aおよび19G定量法を確立し、 アレルギー患者および健 常人の血清あるいは唾液中のアレルゲン特異的抗体発現について解析した。
食物アレルギーは食品中のアレルゲンと呼ばれるタンパク質によって引き起 こされるため、 アレルギー予防にはアレルゲンの除去が有効な手段となる。 そ のためには食品中のアレルゲンの同定が重要であるが、 食品は加熱や加圧など の加工を加えることによりその物性が変化し、 抗原性や栄養価が変化すること
2
がある(Kilshaw et a1., 1982; Nakamura et aJ., 1993; Varshney et a1., 1991)。 また、
加工食品にアレルゲン合有成分が添加される場合もあり、 正確なアレルゲンの 検出法を確立する必要がある。 加うるに、 低アレルゲン食品や 代替食品の 開発
を行う場合、 in vitroにおけるアレルゲン活性の測定は臨床試験を行う前の予備 試験としても必要であり、 in vitroでの食物アレルゲンの検出に、サンドイツチ
ELISA (Levieux and Venien, 1994; 1ρvegrove et a1., 1993; Makinen -Kijunen and Palosuo, 1992)、inhibition ELISA (Kilshaw et aJ., 1982)、radioimmunoassay
(Paganelli and Levinsky, 1980 ) などの方法が用いられている。卵、 大豆および牛乳はアレルギーを引き起こす主要食品と言われており、 卵 ではオボアルブ、ミン(OVA )" オボムコイド、 オボトランスフェリン、 リゾチー ムなどが(Ebbehoj et aJ., 1995 )、 大豆ではGJy mBd 30K などが(Ogawa et a1. , 1991) アレルゲンタンパクとして同定されており、 食品中の OVA抗原活性およ び 大豆タンパクの抗原活性を 調べることは卵アレルギーや 大豆アレルギーの予 防に役立つと思われる。 また、 アレルギー患者は代替食品を利用することも多 いの で、 本研究では低アレルゲン食品あるいは代替食品開発の ための基礎情報 の獲得および抗原活性評価法の 開発を目的として、 分離大豆タンパク質のペプ シンおよび微生物由来プロテアーゼ、分解非消化性画分であるUDFの抗原活性、
および降コレステロール食品として開発された大豆タンパク質を用いたマヨネー ズ製品のOVA抗原活性を ELISA、inhibition ELISA により評価した。
19AやIgGの産生促進などの クラス特異的抗体産生調節はアレルギーの予防 や生体防御に有効であると推察される。 生体内では、 Bリンパ球はTリンパ球 やマクロファージなどの免疫関連細胞と相互作用しており、 Bリンパ球からの 抗体産生は、 インターロイキン-4(IL-4)、IL-5、 インターフエロン y(IFN-y) お
よび IFN-αなどのサイトカインによってクラス特異的に制御されている
(Gauchat et aJ., 1991; Ochel et aJ., 1991; Pene et a1., 1988; Rousset et al., 1991)。 ま
た、マクロファージや抗原提示細胞などの接着細胞はIL-1を産生しており(Borish et a1., 1996)、 Tリンパ球を刺激することにより、抗体産生に影響を及ぼ すことが考えられる。 これらのサイトカインの他に、 マイトジェンや生体およ
3
び食品成分が抗体産生調節機能を有しており、 産生応答が抗体のクラスにより 異なることが報告されているが(Lim et a1., 1994a, 1996; Kaku et a1., 1997)、 これ らの実験はラットリンパ球を用いたものであるが、 抗体産生調節機構の解明に は抗体などの免疫関連試薬の充実しているマウスや ヒトの系を用いることが望 ましい。 そこで、 マウス牌臓細胞を用いた免疫調節機能因子検定系の開発を試 みた。
ラクトフェリンやカゼインなどの乳タンパクはin vitroにおいて抗体産生を合 めた免疫応答調節機能を有することが報告されているが(Yamada et a1., 1989a,
1990a, 1991a)、 その作用機構は不明であり、 マウス牌臓細胞の抗体産生に及ぼす 影響については明らかにされていない。 そこで、 マウス牌臓細胞の抗体産生に 及ぼす乳タンパクの影響を検討し、 その作用機構を解明するためにサイトカイ ンの関与について検討した。
不飽和脂肪酸はリンパ球の増殖を抑制することにより免疫応答に影響を及ぼ すことが報告されているが(Calder et a1., 1992; Soyland et a1., 1993; Tonkin et a1.,
1978)、 抗体産生に及ぼす影響についての研究は非常に限られており、 いずれも ラットリンパ球を用いたものである(Lim et aJ., 1996; Yamada et a1., 1996)。 そこ で、 脂肪酸がマウス牌臓細胞の抗体産生に及ぼす影響について検討した。 多価 不飽和脂肪酸の二重結合は酸素により容易に酸化され、 それによって生じる脂 質酸化物がアレルギーなどの免疫応答に影響を及ぼすことや(Doke et a1.,
1989)、 天然抗酸化成分であるセサミンおよびαートコフエロールを摂食させたラッ
トでは肺のロイコトリエンc4の産生能が減少すること(Gu eta1., 1994, 1995) な どが報告されている。 また、 Yamada et a1. (1996) はアラキドン酸のIgE産生促 進作用の発現がαートコフエロールにより阻害されることを報告している。 これ らの報告は、 抗酸化剤が脂肪酸の抗体産生調節活性を修飾する可能性を示して いる。 そこで、 脂肪酸と種々の抗酸化剤を共存させたときの抗体産生の変化に ついても検討した。
4
第2章 ラットおよびヒトにおけるアレルゲン特異的および 非特異的抗体の分析
第1節 緒 百
食物アレルギーは小児だけでなく成人にも深刻な問題とな っている。 アレノレ ギ一反応は四つのタイプに 分類されるが(Coombs and Gell, 1968)、 食物アレル ギーや環境アレルギーはI型アレルギーによるものが多い。 I型アレルギーは、
マスト細胞や好塩基球表面に結合したアレルゲン 特異的IgE がアレルゲンによ り架橋され、 ヒスタミンやロイコトリエンなどのケミカルメディエーターが放 出されることにより誘導される(Metcalfe, 1991)。 血清中のIgE濃度は、 健常人 では非常に低いが、 アレルギー患者においてはしばしば高濃度で 検出される
(Bjorksten et a1 ., 1983)。 したがって 、 血清中の総IgEおよび アレルゲン特異的 IgEの測定は食物アレルギーの研究に重要である。 IgEの測定には
radioallergosorbent test (RAST)およびその応 用 法(Adler
et
a1., 1991; Williamet
a1., 1992)や皮膚受身アナフィラキシー試験(PCA) が用いられている(Ovary
1958; Sedgwick and Holt, 1983)。 これらの方法はIgE 検出のための感度は充分で あるが、RASTは放射性同位元素を使用 するための特別な施設が必要であり、
PCAは定量性に欠け 、 一度に 多くのサンプルを分析することが困難である。
酵素抗体 法(enzyme-linked immunosorbent assay; ELISA)はこれらの方法の代 替法としてヒト、 マウスおよびラットIgEの検出にも利用されている(Doke
et
a1., 1989; Fehlneret
a1., 1991; Fritsche and Bonzon, 1990)。 しかし、 通常の ELISA 法の感度では抗原特異的IgEの検出には不十分である。 そ こで 、 より検出 感度 の高いアピジンービオチン法を用いてラットIgEの定量を行い、 アレルゲン特異的IgEについて解析した。
IgEはアレルギーの発症に重要な役割を果たしているが、 IgEだけでなくその 他のクラスの抗体も アレルギ一応答に関与している。 分泌型IgA(slgA)は腸管 粘膜の分泌液や唾液 、 涙にも合まれてお り(Watanabe
et
a1., 1984)、 腸管粘膜中5
の sIgAは腸管からのアレルゲンの吸収を抑制することによりアレルギ一反応を
抑制する (Shorter and Tomasi, 1982 )
。
しかし、 腸管 分泌液中の19A発現に関す る情報は非常に限られており、 また、 腸管内のアレルゲン特異的19Aの発現 を調べることは困難である。 そこで、 sIgAの発現に 関する情報を得るために ELISAによる唾液19A の定量を試みた。
第2節 材料と方法
第1項 一般試薬
ß-ラクトグロプリン(ß-L G)およびオボアルブ、ミン(OVA)は Sigma社(St.
Iρuis , MO)より購入し、 分離 大豆タンパク質(ニューフジ プ口氏Soy)は不二製 油(大阪,日本)より提供された。 抗原特異的抗体の定量のため、 βLGおよび OVAは50mM炭酸緩衝液で50μgfmlとなるように調製して使用した。 Soyは
50mM炭酸緩衝液に1 mg/mlとなるように懸濁し、 12.5∞xgで20分間遠心分 離した上清を使用した。
ヒトの唾液および血清サンプルは九州大学食糧化学工学科の学生(男性3人、
女性4人、 年齢21 �26歳)よりインフォームドコンセントを得て採取した。 血 清は、 真空採血管により採血して室温で30分間放置した後、 1 ,500x gで15 分間
遠心分離して調製した。 唾液は、 吐き出し法により採取して75xgで20分間遠 心分離した上清を用いた。 サンプルは全て実験に用いるまで-200Cで保存した。
ラット関連抗体では、 ヤギ抗ラットIgE(εchain)はBethyl社(Montgomery,
CA)より、 ビオチン 標識マウス抗ラットIgE (εchain)および ラットIgE標準抗体 をZymed社(SanFrancis co, CA)より購入した。
ヒト関連抗体では、 ウサギ抗ヒトIgA(α-chain)F (ab')2断片、 ウサギ抗ヒト IgG (y-chain) F (ab')2断片、 ウサギ抗ヒトIgE(ε-chain)、 西洋ワサビペルオキシ
ダーゼ(HRP) 標識ウサギ抗ヒトIgA F (ab')2断片、 HRP標識ウサギ抗ヒト(y-
chain)、 HRP標識アビジンおよび ヒト標準抗体はDako社(Glostrup, Denmark) より、 ビオチン標識抗体である ア フイニティー精製ビオチン標識ヤギ抗ヒト
6
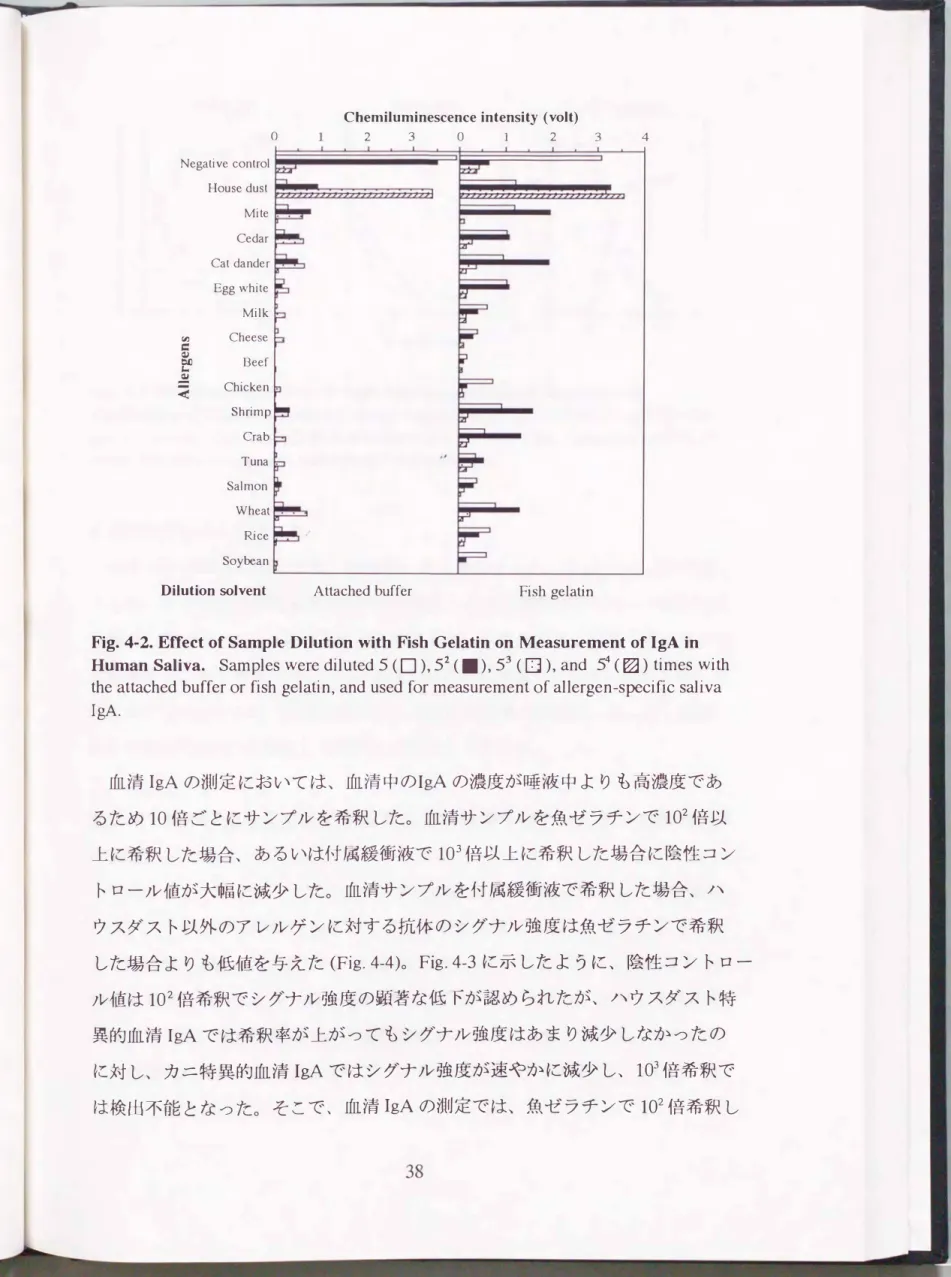
IgE (ε)はKPL社(Gaithersburg, MD)より購入した。 固相抗体は 50 mM炭酸緩 衝液で、 HRP標識抗体およびビオチン標識抗IgEは2%魚ゼラチン溶液でそれ
ぞれ1,0ü0倍に希釈して用いた。
ポリアクリルアミドゲル電気泳動(PAGE)のために、 アグリルアミドモノ
マー 、 トリス(ヒドロキシメチル)アミノメタンおよびグリセロールは ナカライ テスクより、 N, N'-メチレンビスーアクリルアミド、 ドデシル硫酸ナトリウム
(SDS)、 ブロモフエノーノレブ、ルーおよび2-メルカプトエタノールは和光純薬(大
阪,日本)より、 過硫酸アンモニウム(APS)、 N,N, N, N'-テトラメチルエチレン ジアミン(TEMED)および分子量マーカーであるSDS-PAGE standards low range は Bio Rad社より購入した。 また、 電気泳動装置としてミニプロティアン
II 1-D 、
電源としてモデル200 /2.0パワーサプライをBio Rad社より購入して使 用した。
ウエスタンプロッテイングのために、 装置はミニトランスプロットセルをBio Rad社より、 転写用のニトロセルロース膜は Cellulosenitrat(E)をSchleicher
&
Schuell社(Dassel, Germany)より購入した。
イムノプロッティングのために、 ブロッキング液はブロックエースを大日本 製薬(大阪,日本)より、 HRP標識ウサギ抗ヒトIgE をDako社より購入した。
HRP標識抗体は 1/10ブロックエースで1,000倍に希釈して使用し、 洗浄液は Tween 20 をナカライテスク(京都3日本)より購入し、 0.05%Tween 20 合有生理 リン酸緩衝液(TPBS)を調製して使用した。 検出には Amersham社
(Buckinghamshire, England)よりECL western blotting detection reagents を購入 して使用した。
第2項 免疫
4週齢のBrown Norway系雄ラットをセアック吉富(吉富,日本)より購入し、
飼育中はカゼイン20%を合むOVA無添加食を与えた。 oおよび21日目に3%
Al (OH)3 - 0.15 M NaClに懸濁したOVA を50μg/rat腹腔内に投与した。 24日目 にジエチルエーテルで、麻酔し、 腹部大動脈より採血を行った。 採取した血液は
7
3,000 x gで 15分間遠心して血清を調製した。
第3項 血清中の抗体の分画
血清 中の抗体の分画のため、 カラム充填剤はBio-Gel
HT
Hydroxylapatiteを、溶出用緩衝液の濃度勾配作成装置はModel 385 Gradient Formerを Bio-Rad社 (Hercules, CA)より購入した。 カラム装置はModuline™ Medium-Pressure
L aboratory Column (150 x 10 mm, 12 ml bed volume)をアミコン 社(東京,日本 ) より、 フラク ションコレクターはLKB . Redi Fracを Pharmacia社(Uppsala,
Sweden)より購入した。 溶出用緩衝液は、 0.05 M NaCl添加10 mMリン酸緩衝 液(pH 7ムA緩衝液 )および0.3 M NaCl添
別
500 mM リン酸緩衝液(pH 5.8; B緩衝液)を使用した。
カラムにハイドロキシアパタイトを充填し、 A緩衝液で一晩カラムを平衡化 させた後、 ラットあるいはヒト血清を 500μi添加してA緩衝液を流し、 分取を
開始した。 A緩衝液16 mlで洗浄した後、 吸着した IgEを30 mlのA緩衝液およ びB緩衝液の直線濃度勾配(リン酸緩衝液 ;10 mM "-' 500 mM、 NaCl; 0.05 M "-' 0.3 M、 pH; 7.5 "-' 5
.8)により溶出した。
さらに 、B緩衝液16 mlで洗浄し、
合 計60本の画分を得た。 流速は2.8 mllmin、 分取量は1.6ml/tubeの条件下で行っ た。 分 画した血清は実験に使用するまで40Cで保存した。プロテイン Aカラム キット(Pierce社製 ; Rockford, IL)を用いて 、 ヒト血清中 のIgE 画 分中に存在する IgGを除去した。 実験はキットのプロトコールに従っ
て行った。 カラムを室温に戻して保存緩衝液を流出させた後、 結合緩衝液でカ ラムを平衡化させた。 分画した血清と結合緩衝液を1:1に混合してカラムに添
加し、 血清が完全に流出した後に結合緩衝液でカラムに吸着していないタンパ ク質を流出させ、 この溶液を SDS-PAGEおよびイムノプロッティングに用いた。
第4項 ELISA法による抗体の定量
Immuno-plate Maxi sorp C96をNunc社(Roskilide, Denmark)より購入した。
ELISA プレー ト洗浄液はTPBSを使用し、 魚ゼラチンはSigma社(St.lρuis,
8
MO)より購入し、 TPBSで2%になるように希釈し、 ブロッキング溶液、 HRP 標識抗体、 サンプルおよび標準抗体希釈液として使用した。 発色基質溶液は、
0.0069るHρ2合有0.2Mクエン酸緩衝液(pH 4 .0)、 超純水および6 mg/ml 2, 2'-ア ジノー ビス(3-エチノレベンゾチア ゾリンー6-スルホン酸)ジアンモニウム塩( ABTS) 溶液を 10:9: 1で混合して用い、 反応停止液には 1.5%シユウ酸溶液を用いた。
また、 吸光度の測定には東ソ一社製( 東京,日本)のマイクロプレート分光光度計、
MRP- A4iを使用した。
血清IgEの定量においては、 血清中のIgE濃度が低いので検出感度を上げる ためにアビジンービオチン法を用いた。 ラット血清IgEの測定においては、 50
mM炭酸緩衝液で1,000倍希釈した固相抗体を100μi、 もしくは食品タンパク溶
液150μlを96well ELISA プレートに添加し、 370Cで1時間保温した。 つぎに、
ブロッキング溶液を300μl添加して、 40Cで一晩ブロッキングした後、 ブロッ キング溶液で希釈したサンプルまたは 標準抗体 50μlを添加し、 37 0Cで1時間 保温した。 さらに、 ブロッキング溶液で2,000倍に希釈したビオチン標識抗ラッ
トIgE 100μlを添加して 1時間保温した後、 ブロッキング溶液で5,000倍に希釈 したHRP標識アビジン100μlを添加して1時間保温した。 ビオチン標識抗体お よびHRP標識アビジン添加における反応温度は 4、 20、 370Cで行った。 最後に、
発色基質溶液 100μlを添加して370Cで15分間保温した後、 1.5%シュウ酸溶液 を 100μlを添加して反応を停止させ、 マイクロプレート分光光度計で415 nmの
吸光度を測定した。 各反応過程の聞はTPBSでプレートを3回ずつ洗浄し、 サ ンプル添加以後の洗浄は 4 回ずつ行った。
ヒト血清IgEの定量においては、 血清サンプルおよび標準抗体を添加後、 40C で 1時間反応させ、 ブロッキング溶液で1,000倍に希釈したビオチン標識抗 ヒ トIgEを 100μlずつ添加して40Cで1時間保温した。 つぎに、 ブロッキング溶
液で 5,000倍に希釈した HRP標識アビジン100μlを添加して 40Cで30分間保 温した後、 発色反応を行ったo サンプル添加以後の洗浄は4 回ずつ行ったo
ヒト唾液19Aの定量は通常の ELISA 法で 行った。 サンプルおよび標準抗体を 添加後、 ブロッキング溶液で1,000倍に希釈したHRP標識抗ヒトIgA抗体100
9
μiを添加して1時間反応させた後発色反応を行った。 各反応過程の反応温度はブ ロッキング以外は全て37 0Cで行い、洗浄は3回ずつ行った。
第5項 SDS-PAGEおよびイムノプロッティング
サンプル緩衝液は超純水:0.5 Mトリスー塩酸緩衝液(pH6.8) : 509る グリセロー
ル:19るブロモフェノーノレブルー:2-メルカプトエタノール=20 : 5 : 8
: 1
: 2に混 合して調製した。 泳動用緩衝液は、トリス(ヒドロキシメチル )アミノメタン6mg/ml、 グリシン28.8 mg/mlおよび SDS 1 mg/mlとなるように超純水に溶解した。
ウエスタンブロッティング用緩衝液は、 トリス(ヒドロキシメチル )アミノメタ ン3 mg/mlおよびグリシン 14.1 mg/mlとなるように超純水に溶解し、使用直前 にこの溶液とメタノールを 4: 1に混合した。
ゲルの銀染色には、シルバーステインプラスキットをBio R ad社より購入し、
キットのプロトコールに従って操作を行った。 ゲルの固定化には、メタノール:
酢酸:fixative enhancer concentrate :超純水=200
: 40 : 40 :
120に混合してfixative enhancer solution (FES)を調製して用いた。 染色には、超純水 35 ml、
silver complex solution 5.0 ml、reduction moderator solution 5.0 ml、lmage
development reagent 5.0 mlを混合し、使用直前に 室温 の development accelerator solution 50 mlを加えて 、充分に撹持することにより、staining solution (SS)を 調
製した。SDS-PAGEはLaemmli(1970)の方法にしたがい、49る濃縮ゲルおよび129る分 離ゲルを用いて泳動を行った。 サンプルはサンプル緩衝液と容量比1 : 1で混合 し、1 000Cで 5分間加熱後、1ウェルあたり 10μlずつ添加して200V、30分間
泳動した。
SDS-PAGE終了後、ゲルを FESに浸して固定化し、超純水で、2回洗浄した。
調製したSSにゲルを浸し、おだやかに撹持しながら染色を行った。 染色終了後、
5%酢酸溶液にゲルを浸して反応を停止させた。
ウエスタンプロッティングは 100V、1 時間の条件下で行った。 転写後、40C で一晩ブロッキングを行い、HRP-標識抗体を添加して室温で2時間静置して反
検出 ECLに よる蛍光発色はキットのプロトコールに従って行った。
応させた。
試薬1および2を1 : 1に混合してニトロセルロース膜に添加し、 1分間反応させ 各 1分間現像した。
て蛍光発色させ、 8分間ポラロイドフィルムに感光した後、
操作の聞は全てTPBSで10分ずつ3回の洗浄を行った。
結 果 第3節
ELISA法によるラットIgE測定に及ぼす反応温度の影響 第1項
OVA特異的19Bおよびバツ クグランドの吸光度をFig.2-1に示した。
19Eを合ま ないパックグランドの吸光
度は、 370Cでイン キュベートを行った場合、 総19Eの測定においては0.12、反応温度を変えたときのラット血清中の総19E、
しかし、 反応
OVA特異的19Eの測定においては0.15とかなり高い値を示 した。
パックグランドの値は総19Eの測定においては0.03、
温度を40Cに した場合、
OVA特異的 OVAで免疫したラット としてい
ま た、
OVA特異的19Eの測定においては0.0 7にまで減少した。
IgEの測定値は、 反応温度を370Cにした場合、
0 8 6 4
2
11 ハU ハU ハU
ハU
(23『)同切回ωロ刀ω色m,d刊kp。
じニ________.
1.0
令0.8
吋マ
"-' 0.6
同同叫。心
] 0.4
H 0
0.2
0.0 0
0.0 0 20 30 40
Reaction temperature (OC)
40 10 20 30
10
Fig.2・1. Effect of Reaction Temperature on the Determination of Total and
Ovalbumin-specific Rat Serum IgE by the A vidin-biotin Method. The total and
OV A-specific IgE contents in sera from 3 rats immunized with OV A (・) and in sera from
3 non-immunized rats (0) were determined at 4, 20, and 370C. Each A415 value was
deduced from the background level measured each reaction temperature ( ・ ) .
ないラットのどちらにおいても高濃度のOVA特異的IgEが検出されており、 免
疫による抗原特異的IgEの発現量の違いは認められなかった。 これに対して、
反応温度を下げた場合、 免疫していないラット血清中のOVA特異的IgEのシグ ナルは免疫したラットのシグナルより速やかに減少し、 反応温度が40Cの場合 ではOVA特異的IgEの発現量に明らかな差が認められた。
第2項 ハイドロキシアパタイトカラムを用いたラットおよびヒト血清の 分画
Immunized Non-immunized
0.20
370C 370C
0.15
0.10
(23刊)同切同ω回一ωω島mad刊除。0.05 O.∞
0.15 0.10 0.05
0.00
0 20 40 60 。 20 40 60
Fraction number
100
50
忠
信
』 ω
�
箇ロ
100
。
50
80 0
Fig.2・2 EtIect of Reaction Temperature on the Determination of OV A-specific IgE
in Sera Fractionated by a Hydroxyapatite Column. The OV A-specific IgE contents
in the hydroxyapatite column fractionates of sera obtained from a rat immunized with
OVA and a non-immunized rat were determined by the ELISA method at 4 and 370C.
ッ
一フ
第1項で示した 反応温度による測定結果の違いを明らかにするために、
それぞれの画分のOVA特異 ト血清をハイドロキシアパタイトカラムで分 画し、
的IgE量を40Cおよび370Cの反応条件下でアビジンービオチン法により測定した。
Fig.2-2に示したように、 免疫していないラット血清においては、 370Cで OVA これに対して、 免 特異的シグナルが検出されたが、 40Cでは検出されなかった。
疫したラット血清においては、 370CでOVA特異的シグナルのピークがいくつか 番最後のピークのみが消失した。
検出されたが、 40Cでは
三F、ーー
の溶出位置の違いを解析するためには高濃度のIgEを合んだ血清が必要である。
OVA特異的IgEには複数のピークが検出されたが、
Fig.2-2の結果より、
(民)償・む匂ロ凶
100
1∞
。 50 50
(回同守司)同凶同ωロ℃ω《凶mEd刊k,。
。
ゥ,U
1i
nv ハu
nu
ハUぷU ハU
A
0.2 0.0
0.4
0.2
ハU ハU ハU
1.0 0.8 0.6 0.4
fo ハU
(虫マ〈)同関岡ωロ刀ω色白・ロ伺mwahc∞
(@eHM23刊)-30H
50
Fraction number
40 30 10 20
Fig.2・3. Measurement of Total and Antigen-specific IgE in Human Serum
Fractionated by a Hydroxyapatite Column. Total (A) and antigen-specific IgE (B)
contents were detennined by the ELISA method. (・); OVA-specific, (0); soybean
specific IgE.
そこで、血清IgEの抗体価が高く、以前に大豆アレルギーと診断されたヒトの 血清について同様の検討を行った。 Fig 2- 3に示したように、総IgEにおいては 17 および 22番目に 2 つのピークが認められた。 OVA特異的IgEにおいては14、
17、19、22、26番目にピークが認められ、特に19番目のピークが最も高かった。
また、 Soy特異的IgEにおいては16、19、28 、46、5 0番目にピークが認められ、
特に28 番目のピークが最も高く、抗原 の種類によりIgEの溶出ピークが異なる ことが明らかと なった。
第3項 イムノプロッティングによるヒトIgEの分析
Fig.2-3で抗原特異的IgE抗体価が最も高かった19 および28 番目の画分につ いて、 SDS-PAGEおよびイムノプロッティングを用いて解析を行った。 また、
IgEに対する抗体はIgG と交叉反応することがあるのでプロテインA により
IgGを除去した画分についても同様の検討を行った。 OVA特異的IgEピーク画 分および Soy特異的IgEピーク画分には分子量約74kDaのIgE重鎖と思われる バンド、分子量約51kDaのIgG重鎖と思われるバンド、 および分子量約23 kDaの軽鎖と思われるバンドを合む複数のバンドが検出されたが、プロテイン Aで処理したそれぞれのピーク画分では51kDaおよび23kDaのバンドは消滅 し、74kDaの濃いバンドおよび85 kDaの薄いバンドが検出された(Fig. 2-4 A)。
イムノプロッティングを行った結果、OVA特異的IgEピーク画分およびSoy 特異的IgEピーク画分では 74kDaと 85 kDaの二つのバンドが検出され、プロ テインA で処理したそれぞれのピーク画分では分子量約85 kDaのバンドは消滅
し、74kDaのバンドのみが検出された(Fig. 2-4 B)。
第4項 ELISA法によるヒ卜唾液IgAの検出
成人ドナー5人の唾液中の総19A、ß-LG、OVA および、Soy特異的19A を
ELISA法で測定した結果をFig.2-5 に示した。 5人のドナーにおいて、唾液中の 総IgA濃度は190- 470μg/mlで検出されており、その発現量の違いは約2.5倍で あった。 代表的な食物アレルゲンであるß-LG、OVAおよび、Soy に対する唾液
A
Heavy chain (ε)�
Heavy chain (y)一一炉
Light chain-・P
B
2 3 4
-
‘ 'ー一戸『濁...a:.-ニ以
一、写�.-!.;i _-
ゐ園田園田•
Ar
‘晶噌副島2 3
...-司ーー
e .
4
Fig.2・4. Detection of IgE in Human Serum by SDS-PAGE and Immunoblotting.
Serum fractionated by a hydroxyapatite column was applied to SDS-PAGE and immunoblotting. Panel A shows silver staining of polyacrylamide gel and panel B shows ECL detection of immunoblotted membrane. Lane 1, peak fraction of OVA- specifìc IgE� lane 2, peak fraction of soybean-specific IgE� lane 3, protein A -treated peak fraction of OV A-specific IgE� lane 4, protein A-treated peak fraction of soybean-specific
IgE.
19Aの濃度を測定した結果、 各ドナーの分泌パターンは類似した傾向を示したが、
その発現量は2.7
- 50
ngfmlの範囲で検出されており、 発現量の違いは約19倍と 総19A量より個人差が大きいことが明らかとなった。 また、 総IgAに対する抗 原特異的IgAの割合は1%
(ドナーC、 OVA特異的19A)から12%
(ドナーD、600 60
Total Iß・lactoglobulin
宙
空 4∞ -
司)
( 、g 皆司 、、 40
J45u
ロ曲胴S轟
2∞
‘ 場ロ山
5回u ーd
20
4田。到圃
tJ
4回。同到£
。 。
A B C D E A B C D E
Donors Donors
60 60
Ovalbumin " I Soybean
-、g E 司、
P40
ー ( 、
、 宮 皆-
、J、 40
JO 5 図。
田2
, a u .
却
J 。E
曲5u
-S
20
4回むぞ・司
£
4恒。・可司£
。 。
A B C D E A B C D E
Donors Donors
Fig.2・5. Determination of Total and Food Component-specific IgA in Human Saliva. In adition to total IgA, ß-LG-, OVA- and s oy b ean -spe c ifi c IgA in human saliva obtained f rom fi ve donors (A -E) were dete口nined by ELISA.
Soy特異的19A)であり、 ドナーCは、 他のドナーが三種類の抗原において総
19Aに対する比率が8%以上であるのに対し、 5%以下と食品成分特異的唾液 19Aの発現率が低い傾向が認められた。
一人のドナーの唾液を午前10時から午後10時まで3日間連続して採取し、 総
唾液I�生および食物抗原としてSoy特異的IgA発現量の日内変動を調べた。 Fig.
2-6に示すように、 総IgAは107-5,400 f.tg/ml、 Soy特異的IgAは5 - 224 ng/ml の範囲で検出されが、 その発現量にサンプル採取時刻依存性は認められなかっ
た。
16
第4節 考 察
多くの物質が合まれている溶液の中 ELISA法(Engvall and Perlman, 1971) は、
から種々のタンパク質や生体成分を特異的に検出する方法として最も頻繁に用 ウ いられている。 抗体を用いる実験系では抗体の非特異的吸着を防ぐために、
シ血清アルブ、ミンや乳タンパクによるブロッキング操作が行われる(Yamada
et
アレルギーに関する実験を行う場合にはブロッキン しカ瓦し、
a1., 1989b, 1991a)。
グ剤として使用する乳タンパク自身がアレルゲンとして検出される可能性があ アレルゲン性の低い魚ゼラチンをブロッキング剤として用いることが るので、
本実験においても魚ゼラ ある(Fritsche and Bonzon, 1990; Yamada
et
a1., 1996)。チンをブロッキング剤あるいは抗体および、サンプルの希釈液として使用した。
アビジンービオチン法は通常のELISA法よりバックグランドレベルが高くなる バックグランドレベルをさげるために洗浄回数の増加や反応温度 傾向にあり、
戸、J ハU 1i
ロ
企.�・�.
t込 ロ
�口llO
•
。
D
�
•
•
•
ロAUo aロ
会-,
h
G
口 0
• Ie
企
II 巴�
F
‘
ロAo
a
pbLE
-A
- thh fL
4 3 2 1 0 」 3 2 ハU ハU ハU ハu ハU nv ハU ハU 噌'i 唱'i 噌Ei 唱ai 噌Ei 'ai 噌si 噌Ei
(-shュ)回。判定bgω回。ω司丘
E
よ
�
24 22
20 18
16 12 14
10
Time
Fig.2・6. Time Courses of Total and Soybean-specific IgA Contents in Human
Saliva. Saliva were col1ected from a donor for successive 3 days. Open and closed
symbols are the result of total and soybean-specific IgA. Circles are the results of the
first, triangles are the second, and squares are the third day, respectively.
の検討が必要である。 OVAで免疫したラットおよび免疫していないラット血清 中の総IgEおよびOVA特異的IgEを測定した結果、 反応温度370Cでは免疫し ていないラット血清中にも高濃度のOVA特異的IgEが検出され、 免疫による抗 原特異的IgEの発現量の違いは認められなかった。 反応温度を下げた場合、 免 疫していないラット血清中のOVA特異的IgEシグ、ナルは免疫したラットのシグ
ナルより速やかに減少し、 反応温度が40Cの場合ではOVA特異的IgEの発現量 に明らかな差が認められた。 反応温度による測定結果の違いを明らかにするた めに、 抗体の精製に利用されているハイドロキシアパタイトカラム分画(Juarez
Salinas et aJ.,
1986; Langone, 1982)を行って検討した。 反応温度が370Cの場合、免疫していないラット血清中には二つのOVA特異的IgEピークが検出されたの に対し、 免疫したラット血清中には三つのOVA特異的IgEピークが検出された。
反応温度が40Cの場合、 免疫していないラット血清中のOVA特異的IgEピーク は消滅し、 免疫したラット血清中のOVA特異的IgEピークは三番目のピークの みが消滅したが、 他のピークのシグナル強度の減少などは認められなかった。
これらの結果は、 アビジンービオチン法の非特異的シグナルを低減させるために は反応温度を下げることが有効であることを示している。 Masonand Williams (1980)は抗原抗体複合体の解離定数は抗体、 pHおよび温度に依存して結合定数 よりも変化することを報告している。 本研究で認められた低温での非特異的シ グナル減少の原因は、 抗体の非特異的結合の減少、 あるいは非特異的に結合し た抗原抗体複合体の解離促進などが考えられる。
免疫したラット血清中のOVA特異的IgEには複数のピークが検出されたが、
このピークの違いを解析するためには高濃度のIgEを合んだ血清が必要であっ た。 そこで、 血清IgEの抗体価が高く、 以前に大豆アレルギーと診断されたヒ
トの血清についてもハイドロキシアパタイトカラム分画を行った。 その結果、
総IgEおよび抗原特異的IgEにおいて複数のピークが認められること、 抗原の 種類によってピークの位置が異なることが明らかとなった。 それぞれの抗原特 異的IgEのピーク画分をプロテインAでさらに精製してSDS-PAGEおよびイム
ノプロッティングを行った結果、 SDS-PAGEでは分子量約74 kDaの濃いバンド
および分子量約85 kDaの薄いバンドが検出された。 分子量およびイムノプロッ ティングの結果から74
kDaの濃いバンドが19E重鎖であると推察され、 85 kDa
のバンド、についてはさらなる検討が必要であると考えられる。 また、 それぞれのピーク画分の19E重鎖の分子量が同じであったことから、 ハイドロキシアパ タイトカラム分固による溶出ピークの違いは19E重鎖の分子量の違いによるも のではないことが明らかとなった。
Juarez-Salinas et al. (1986)はハイドロキシアパタイトカラム分画により軽鎖の 分子量が異なる19Gアイソタイプが分画可能であることを示しているが、 今回 の実験からは軽鎖の分子量に明らかな違いは認められなかった。 Lyczak etaJ.
(1996)はヒトにはC末端付近の違いにより四つの19Eアイソタイプが存在し、
マウスミエローマ細胞株を用いてこれらのアイソタイプを発現させて解析し、
一つのアイソタイプが75 kDaの重鎖を持つことを示している。 この結果は本実 験の19E重鎖の分子量とほぼ一致している。 また、 Batista
et al.
(1995)はIgE重 鎖に結合している糖鎖に違いがあることを報告している。 糖鎖や機能の違いを 合めてヒト19Bの多様性についてはさらに検討する必要があると思われる。分泌液中の19Aの発現に関する情報を得るため、 唾液中の総19Aおよび食物 アレルゲン特異的19A定量法を確立し、 5人のドナーについてその発現を調べた。
その結果、 唾液IgAの発現量には個人差があること、 アレルゲンの種類によっ て総19Aに対する割合が異なることが明らかとなった。 唾液の分泌量は 持続的 な運動により減少すること(Mackinnon and Jenkins, 1993)、 食品や意識の影響を 受けて変化すること(Edgar, 1992; Tachiyashiki
et aJ.,
1992a, 1992b)などが 報告さ れているo ここで認められた唾液19Aの発現量の違いが、 唾液分泌量の変動に 基づいている可能性が考えられたので、 一人のドナーの唾液19A発現の経時的 変化を3日間にわたって 調べた。 その結果、 唾液IgAの発現量はある程度変動 することが明らかとなったが、 発現量にサンプル採取時刻依存性は認められな かった。 これらの結果より、 唾液19Aの発現量には個人差があることが明らか となり、 さらに多くの症例について検討することにより唾液19Aを利用したア レルギー簡易診断法の開発やアレルギ一発症機構の解明のための基礎情報が得19
られる可能性が示された。
第3章 アレルゲン特異的抗体発現のクラスおよび サブクラス依存性
第1節 緒 日
食物アレルギーは小児だけでなく成人に も深刻な問題となっている。 食物ア レルギーは免疫機能の過剰反応によって引き起こされるが、 免疫機能に関与し ている抗体には19A、 IgD、 IgE、 IgG、 IgMの五種類 がある。 アレルギ一反応は Coombs and Gell (1968)によって 四つのタイプに 分類されているが、 そのうち IgEはI型アレルギーの原因抗体であり、
食
物アレルギーや環境アレルギーはI型アレルギーによるものが多い。 I型アレルギーはマスト細胞や好塩基球表面に 結合したアレルゲン特異的IgEがアレルゲンにより架橋され、 ヒスタミンやロ イコトリエンなどのケミカルメデイエー ター が放出されることにより誘導され る(Metcalfe, 1991)。 血清中のIgE濃度は、 健常人では非常に低いが、 アレルギー 患者においてしばしば高濃度で検出される(Bjorks ten
et
a1., 1983)。19Aは血清中だけでなく、 分泌型19A (sIgA)は腸管粘膜の分泌液や唾液 、 涙 にも合まれており(Watanabe
et
a1., 1984)、 腸管粘膜中の sIgAは腸管からのアレ ルゲンの吸収を抑制することによりアレルギ一反応を抑制する(Shorter andTomasi, 1982)0 19Aには二つのサブクラスが存在するが(Depelchin et a1. ,
1994)、 機能の違いやアレルギー との関連についてはほとんど明らかにされてい ない。
IgGは血中に最も多量に合まれている抗体であり、 アレルギーの減感作療法に より誘導される抗体で I型アレルギーの原因抗体であるIgEと競合するこ とによ りアレルギー抑制的に働いたり(Aalbers e
et
a1., 1993)、 終生 免疫の獲得に関与し ている。 IgGには四つのサブクラスが存在しており、 補体の活性化能やナチュラ ルキラー細胞の活性化能に若干の違いが認められるが(Janewa�ら1996)、 機能の 違いについては不明な点も多い。IgMは初期免疫に関与し、 抗原感作を受け ると最初に産生される抗体である。
これらの抗体はアレルギー の発症および抑制に関与していると考えられるが、
これらの抗体の発現について総合的 に解析した研究はほとんどないため、 本 研 究で は成人およびアレルギー患者血清中のアレルゲン特異的抗体のクラス およ びサブクラス依存性について検討した。
第2節 材料と方法
第1項 一般試薬
ß-ラクトグロプリン(βLG)はSigma社(St.lρuis,
MO)より、
卵粗製アルブ、ミ ン(Alb)はナカライテスク(京都,日本)より、 スギ花粉抗原(Cedar)は三光純薬 (東京,日本)よりアラスタットアレルゲンスギ抗原試薬を購入し、 分離ダイズタンパク質(ニューフジ プ口氏Soy)は不二製油(大阪,日本)より提供された。
血清サンプル は九州大学食糧化学工学科 の学生4人(年齢22 �27歳)および 国立療養所南福岡病院小児科でアレルギーと診断された患者8人(年齢o � 15 歳)よりインフォームドコンセントを得てから採取した。 血清は、 真空採血管を 用いて 採血して室温で30分間放置し、 1,500x gで15分間遠心分離して調製し、
実験に用いるまで-200Cで保存した。
抗体は西洋ワサ ビベルオキシダーゼ(HRP)標識ウサギ抗ヒトIgAのF (ab')2 断片、 HRP標識ウサギ抗ヒトIgG (y-chain)のF (ab')2断片、 HRP標識ウサギ抗 ヒトIgM(μ-chain)のF (ab')2断片、 HRP標識アビジンおよびヒト標準抗体を Dako社(Glostrup, Denmark)より購入した。 ま た、 IgE 定量のためアフイニ ティー精製 ビオチン標識ヤギ抗ヒトIgE (ε)をKPL社(Gaithersburg, MD)より 購入した。 サブクラス抗体の定量に は、 抗ヒトIgAl、 IgA 2、 IgG l、 IgG 2、
IgG3、 IgG4およびHRP 標識ロバ抗ヒツジ IgG (H+L)をBinding Site社 (Birmingham, England)より購入して用いた。
第2項 ELISA法による各クラス抗体の定量
E L ISA用として、 Immuno-plate Maxi sorp C96をNunc社(Roskilide,
Denmark)より購入した。 EL1SAプレート洗浄液 はTween 20をナカライテスク より購入し、 0.05%Tween 20合有生理リン酸緩衝液 ( TPBS)を調製して使用し
た。 魚ゼラチン はSigma社より購入し、 TPBSで2%になるように希釈し、 ブロッ キング溶液 、 HRP標識抗体、 サンプルおよび 標準抗体希釈液として使用した。
発色基質溶液 は、 0.006%
H202合有0.2 M クエン酸緩衝液 (pH 4.0)、 超純水およ び6mg/ml 2, 2'_アジノービス(3-エチルベン ゾチアゾリン-6- スルホン酸) ジアンモ
ニウム塩(ABTS)溶液を10:9: 1で混合して用い、 反応 停止 液 には1.59るシュウ 酸溶液を用いた。 また、 吸光度 の 測定には東ソ一社製(東京,日本) のマイクロプ
レート分光光度計 、 MRP-A4iを使用した。
血清19E の定量にお いては、 血清中の19E濃度が低いので検出感度を上げる ためにアビジンービオチン法を用いた。 抗原特異的抗体定量の ための抗原溶液と
して、ß-L Gおよび A1bは50mM炭酸緩衝液で50μg/mlとなるように調製した。
Soyは50mM炭酸緩衝液で1mg/mlとなるように懸濁し、 12,500
x
gで20分間 遠心分離した上清を使用した。 スギ抗原は50mM炭酸緩衝液で106倍に希釈し て用いた。 これらの抗原溶液を150μlずつ96 well EL1SAプレート に添加し、370Cで1時間保温した。 ブロッキング溶液を300μiずつ添加して 40Cで一晩ブ ロッキング、した後、 ブロッキング溶液で希釈した血清または標準抗体を50μ1ず つ添加し、 40Cで1時間反応させた。 つぎ、に、 ブロッキング溶液で1,000倍に希 釈したビオチン 標識抗 ヒト19Eを1∞μlずつ添加して40Cで1時間反応させた。
ブロッキング溶液で5,000倍に希釈したHRP標識 アビジンを100μlずつ添加し て 40C に30分間保持した後、 発色基質溶液を100μlずつ添加し、 370Cで15分
間反応させた。 最後に、 1.59るシュウ酸溶液を100μ1 添加して反応を停止させ、
マイクロプレート分光光度計で 415 nm の吸光度を測定した。 各反応過程の問は TPBSでプレートを3回ずつ洗浄し、 サンプル添加以後の洗浄は 4回ずつ行った。
19A、 IgGおよび19M の定量は通常のEL1SA法で行った。 サンプルおよび 標 準抗体を添加後、 ブロッキング溶液で1,000倍に希釈したHRP標識抗ヒト19A、
19Gおよび19Mを100μiずつ添加して1時間反応させた後に発色反応を行った。
各反応過程の 反応温度はブロッキング 以外は全て370Cで行い、 洗浄はTPBSで
3回ずつ行った。
第3項 ELISA法による各サブクラス抗体の定量
抗原溶液を150μlずつ96 well ELISA プレートに添加し、370Cで1時間保温 した。 ブロッキング溶液を300μlずつ添加して40Cで一晩ブロッキングした後、
ブロッキング溶液で希釈した血清を 50μlずつ添加し、40Cで1時間反応させた。
つぎ、に、ブロッキング溶液で10,∞0倍に希釈した抗 ヒトサブクラス 抗体を100 μlずつ添加して40Cで1時間反応させた。 さらに、ブロッキング溶液で2,000倍 に希釈したHRP標識抗ヒツジIgG を100μ!ずつ添加して40Cで1時間反応 後、
発色基質溶液を100μlずつ添加して370Cで15分間反応させた。 1.5%シュウ酸 溶液を100μl添加して反応を停止させ、マイクロプレート分光光度計で415nm の吸光度を測定した。 各反応過程の聞はTPBSでプレートを3回ずつ 洗浄し、
サンプル添加以後の洗浄は4回ずつ行った。 サブクラスの抗体価は各被験者の IgGlおよびIgA1のA415値を1 としたときの相対値 として表した。
第3節 結 果
第1項 健常人におけるアレルゲン特異的抗体発現パターンの解析
本学科学生4人の血清中のアレルゲン特異的抗体量を ELISA により定量した 結果をFig.3-1に示した。 MA STで アレルゲン特異的血清 IgEを測定した結果、
donor AおよびBではスギ花粉に対するIgEが検出されており、donor Cおよび Dではいずれのアレルゲンに対しでもIgEは検出されなかった。 Fig.3-1に示す
ように、ß-LG、Soy、Cedar特異的IgEは4人のドナー全てに検出され、Alb 特
異的IgEはdonor Cでは検出されず、donor A、Bで高い傾向が認められた。 IgA では、donor A およびCのAlb、 ß-LG、Cedarに対する抗体が高い傾向が認め
られ、donor CのSoy に対する抗体は検出されなかった。 IgG では、donor A 以 外のドナーのAlbおよびß-LG に対する抗体が高い傾向が認められ、donor A は 他のドナーに比べて 抗体の発現量が低い傾向が認められた。 IgMでは、donor B
24
IgE(略Iml) IgA(nglml)
3 4 _ 5 _ -1 _ 0 _ 1 _ 2 _ 3 4 5 _ � 6
10 -
10 10 10 10 10 10 10 10 10 10 10 10 10 10
-1 0 1 2 3 4 5 -1 0 1 2 3 4 5 6
10 10 10 10 10 10 10 10 10 10 10 10 10 10 10
IgG(昭Iml) IgM (nglml)
Alb
Soy
ß-凶
Cedar
Soy
Cedar Alb
ß-lβ
EEω国司
Fig.3・1. Expression of Allergen-specific Antibodies in Sera of Healthy Adults. Sera were collected from 4 donors and applied to ELISA.
(口);donor A,
(圏);donor B,
(図);donor C,
(・);donor D.
3 3
2
(HEOM巴省S回。ωOω同ωkFZSω出ß-LG
A1b
。
Cedar Soy
。
(何日間。凶同)恒国畠ロ。ωO回同ωkZ富山ω出
。 C D
B A D
Donor
B C A
。
Fig.3・2. Expression of Allergen-specific IgG Subclωses in Sera of Healthy Adults.
The data of IgG2, IgG3, and IgG4 are expressed as the relative value of the data of IgG 1.
(口);
IgGl,
(圏);IgG2,
(図);IgG3,
(・);IgG4.
およびCのSoyに対する抗体が検出されなかった。
Fig.3- 2にIgGサブクラスの発現を検討した結果を示した。 AlbおよびβLG ではIgG 4の発現が、 SoyではIgGlの発現が優位で、あり、 CedarではIgG lと
IgG4が同程度に発現していることが明らかとなった。 また、 いずれのアレルゲンにおいてもIgG2とIgG3の発現は同程度であった。
Fig.3-3に19Aサブクラスの発現を検討した結果を示した。 Albおよびß-LG ではIgAl とIgA2が同程度に発現し、 Cedarおよび SoyではIgAlの発現が優位 である傾向が認められたが、 donorCのβLGに対する19A2の発現のみ低かっ た。
第2項 アレルギー患者におけるアレルゲン特異的クラス抗体発現パター
ンの解析
アレルギー患者のうち、 卵アレルギー患者patientA
'"'-' D
(年齢0'"'-'1歳)の血2
s
J H
E 国盟 E
O
�
沼芯 E
ω
a
s 国
・
l
。 Alb
Soy
円 円 円
A B C
。-LG 2
1 3
(,4 司 1 1
o s g ‘固' H Cedar
-4。・可
4 a
円 円 円 円 円 1 壱 凶 .5
q砂 ,
D A B C D 。
Donor
Fig.3・3. Expression of Allergen-specific IgA Subclasses in Sera of Healthy Adults.
The data of IgA2 is expressed as the ralative value of the data of IgAl. (口); 19A1, (圏);
19A2.
清中のアレルゲン特異的抗体量をELISAにより定量した結果をFig.3-4に示し
た。 PatientAおよびBは卵に対してのみ、 patientCおよびDは卵以外に牛乳 および大豆に対してアレルギー症状を示す。 Fig.3-4に示すように、 patientAお よびBはAlbおよびß-LGに対するIgEを有しており、 patientCおよびDは4
種類のアレルゲン全てに対するIgEを有していた。 IgAにおいては、 Soyに対す る抗体の発現がpatientAおよびBで認められず、 Cedarに対する抗体の発現が patient Bで認められなかった。 A1bおよびß-LG特異的I!Þ生はいずれのpatient においても発現していた。 IgGは、 いずれのアレルゲンに対しでも高濃度で検出 される傾向を示したが、 Soyに対する抗体がpatientBで検出されなかった。
IgMは、 patientDで発現が低い傾向を示し、 A1bに対する抗体は検出されなかっ た。 アレルギー患者のうち、 環境アレノレゲンに対してアレルギー症状を示す
patient E
,..____,H
(年齢7,..____,15歳)の血清中のアレルゲン特異的抗体量をELISAに より定量した結果をFig.3-5に示した。 MASTによりpatientEおよびFは環境 アレルゲンに対するIgEが検出されており、 patientGおよびHは環境アレルゲ
ン以外に卵白および大豆に対するIgE が検出されている。 Fig.3-5に示すように、
アレルゲン特異的IgEの発現量は全体的に低い傾向を示し、 A1bに対するIgE は patient EおよびGに検出され、 ß-LGに対するIgEは全てのpatientで検出され た。 IgAにおいては、 A1bに対する抗体はpatient全員に検出されており、 ß-LG およびSoyに対する抗体は2人のpatientにしか発現が認められず、 Cedarに対 する抗体の発現がpatientEで認められなかった。 IgGはいずれのアレルゲンに 対しでも高濃度で検出される傾向を示した。 IgM はアレルゲンの種類やpatient による発現量の違いは少ない傾向を示したが、 patientEの Soyに対する抗体は 検出されなかった。
第3項 アレルギー患者におけるアレルゲン特異的IgGのサブクラス 依存性
Fig.3-6に卵アレルギー患者血清中のIgGサブクラスの発現を検討した結果を
示した。 A1bおよびβLGではIgGlの発現が最も強く、 次いで、IgG3あるいは
IgE(nglmJ) IgA(ngl凶)
1 ,,,U ,(\ I ,(\2 ,(\3 . (\4 . (\ 5 -1 0 1 2 3 4 .5 6
10 10 - 10 -
10
-10 10 � 10 J 10 - , 10 v 10 1 10 L. 10 � 10
斗10
コ10
0A1b
βLG
Soy
Cedar
S Meg- -d司
-・・・・・圃・・・・・圃・・・・・・・・・・・・・・・・・・・・...
“““““““““““““““L 4
A1b
ß-LG
Soy
zwifØfi?p/A?Jþiß/?(1'j7A'
ー 1 • (\ 0 • (\ 1 _ _ 2 3 _ 4 5 - -1 0 1 2 3 4 弓 民
10 10 10 10 10 10 10 10 . )O v
10 10 � 10 --'
10 喝 IO J
lO u
IgG(ngl凶) IgM(ngl凶)
Cedar
Fig.3・4. Expression of Allergen-specific Antibodies in Sera of Egg Allergy Patients.
Sera were collected from 4 egg allergy patients and applied to ELISA.
(口)�patient A,
(函)�
patient B, (図)� patient C, (・)� patient D.
IgE(nglmJ) IgA(ngl凶)
ー 1 _ 0 _ 1 2 _ 3 _ 4 5 -1 0 1 _ 2 _ 3 _ 4 5 6
10 10 10 10 10 10 10 10 10 10 10 10 10 10 10
』-・E・-・E・-圃圃圃園田・圃圃E・E・圃
“““““““““““““““““負““““““ι.
「一一一一一一一一一一一一一一一
A1b
也ρしW
C
Rw凶ιZ司d司
ß-凶
件LGSoy
A1b
Soy
mm初防初防 r--・圃園田E・E・
ー1 0 1 2 3 4 5 -1 0 1 2 3 4 5 6
10 10 10 10 10 10 10 10 10 10 10 10 10 10 10
IgG(ngl凶) IgM(ngl凶)
Cedar
Fig.3・5. Expression of Allergen-specific Antibodies in Sera of Allergy Patients
Sensitive to Environmental Allergens. Sera were collected from 4 allergy patients
sensitige to environmenta1 allergens and applied to ELISA.
(口)�patient E, (圏); patient
F,
(図);patient G,
(・);patient H.
IgG4の発現が強い傾向が認められた。 しかし、 patients Aにおいては他の
patientに比べてβLGに対するIgG4の発現が強かった。 SoyおよびCedarでは IgGlの発現がかなり優位であることが明らかとなった。
Fig.3-7に環境アレルギー患者血清中のIgGサブクラスの発現を検討した結果 を示した。 AlbではIgGlおよびIgG4の発現が同程度、 もしくはIgG4の発現が 強い傾向が認められ、 IgG3の発現も比較的IgGlの発現量と近い傾向が認められ た。 ß-LGではpatientにより発現パターン が異なっていたがIgG4の発現が強い 傾向が認められた。 しかし、 patientEにおいては他のpatientに比べてβLGに 対するIgG4の発現が弱かった。 SoyではIgGlの発現が優位である傾向が認めら れたが、 patientFにおいてはIgG3の発現が、 patientHにおいてはIgG4の発現
が強かった。CedarではIgGl およびIgG4の発現が優位で、ある傾向が認められた。
第4項 アレルギー患者におけるアレルゲン特異的IgAのサブクラス 依存性
Fig.3-8に卵アレルギー患者血清中のIgAサブクラスの発現を検討した結果を 示した。 卵アレルギー患者においては、 いずれのアレルゲンにおいても恥位の 発現はIgAlの半分以下であり、 IgA1の発現がかなり優位であることが明らかと なった。 また、 IgA2の発現の割合はpatientおよびアレルゲンの種類により若干 異なっていた。
Fig.3-9に環境アレルギー患者血清中の19Aサブクラスの発現を検討した結果 を示した。 環境アレルギー患者においては、 Albに対するIgA1 および19Azの発 現が同程度であったが、 他のアレルゲンでは19A1の発現が優位で、あった。 特に、
SoyおよびCedarに対するIgA2の発現の割合が低い傾向が認められた。
第4節 考 察
健常人およびアレルギー患者について血清中のアレルゲン特異的抗体量を測 定した。 その結果、 アレルギー症状が認められないdonorAおよびBにおいて、
2
書
H
a s gω o
。
凶E τ T ω a 4 a
Z 3 ,
I
。
Alb
Soy
n n n n
A B C D
ß-LG
Cedar
n n n n
A " B C
Patient
D 2
1 8
。同) 例a 1 1 a
0 S
ロ5
。oll
l
ta
-g ω ee b
。
Fig.3・6. Expression of Allergen-specific IgG Subclasses in Sera of Egg Allergy
Patienú句・The data of IgG2, IgG3, and IgG4 are expressed as the relative value of the data of IgG 1.
(ロ); IgGl, (圏); IgG2, (図); IgG3, (・);
IgG4.4 3
( 官o
同H ) se
2 a
制a
回。ω 。
。
個凶圃
a2ee 3 2
。
Alb
Soy
四
E F G
ß-LG
Cedar
H E F G
Patient
H 4 3
2
;円
o
同H )
e l町la 、
oo a
世