Caco-2 細胞 sheet における Poly-L-arginine の水溶性
高分子薬物の透過促進機構に関する研究
目次
緒言 ・・・ 1
第 1 編 水溶性高分子薬物の in vivo ラット腸管からの吸収及び in vitro Caco-2 細 胞 sheet を介した透過に対する PLA の効果 第 1 章 種々腸管部位からの FD-4 の吸収に対する PLA の促進効果 第 1 節 小緒言 ・・・ 6 第 2 節 実験方法 1-2-1 試薬 ・・・ 8 1-2-2 動物実験 ・・・ 8 1-2-3 静脈内投与 ・・・ 8
1-2-4 In situ closed loop 実験 ・・・ 9
1-2-5 血漿採取法 ・・・ 9 1-2-6 血漿中 FD-4 濃度測定 ・・・ 9 1-2-7 動態学的解析 ・・・ 10 1-2-8 Immunoblot analysis ・・・ 10 1-2-9 統計解析 ・・・ 12 第 3 節 結果 1-3-1 腸管各部位における FD-4 の吸収に対する PLA の効果 ・・・ 13 1-3-2 腸管各部位における PLA の吸収促進効果に 対するタンパク質分解酵素阻害剤の影響 ・・・ 16 1-3-3 腸管各部位における occludin の発現量 ・・・ 19 第 4 節 小括 ・・・ 20 第 2 章 Caco-2 細胞 sheet における経上皮電気抵抗、FD-4 透過性、 TJ 関連タンパク質の局在性及び発現量に対する PLA の影響 第 1 節 小緒言 ・・・ 22 第 2 節 実験方法 2-2-1 試薬及び抗体 ・・・ 23
2-2-2 細胞培養 ・・・ 23 2-2-3 経上皮電気抵抗 (TEER) の測定 ・・・ 24 2-2-4 FD-4 透過実験 ・・・ 25 2-2-5 FD-4 透過経路の観察 ・・・ 26 2-2-6 MTT assay ・・・ 26 2-2-7 Immunofluorescence microscopy ・・・ 26 2-2-8 Immunoblot analysis ・・・ 27 2-2-9 統計解析 ・・・ 27 第 3 節 結果 2-3-1 Caco-2 細胞 sheet における TEER 及び FD-4 透過性に対する PLA の影響 ・・・ 29 2-3-2 Caco-2 細胞 sheet における TJ 関連タンパク質の 局在性に対する PLA の濃度による影響 ・・・ 32 2-3-3 Caco-2 細胞 sheet における AJ 関連タンパク質の 局在性に対する PLA の影響 ・・・ 35 2-3-4 Caco-2 細胞 sheet における TJ 関連タンパク質の 発現量に対する PLA の影響 ・・・ 36 2-3-5 PLA による透過促進効果の可逆性の検討 ・・・ 37 第 4 節 小括 ・・・ 40 第 2 編 PLA による TJ 関連タンパク質の細胞間隙部位からの消失機構 第 1 節 小緒言 ・・・ 42 第 2 節 実験方法 3-2-1 試薬及び抗体 ・・・ 45 3-2-2 細胞培養 ・・・ 45 3-2-3 エンドサイトーシス阻害剤含有溶液の 調製並びに適用 ・・・ 45 3-2-4 TEER 測定並びに FD-4 透過実験 ・・・ 46 3-2-5 Erythritol 透過実験 ・・・ 47 3-2-6 Immunofluorescence microscopy ・・・ 48 3-2-7 統計解析 ・・・ 48
第 3 節 結果 3-3-1 エンドサイトーシス阻害剤存在下での Caco-2 細胞 sheet における TEER 及び FD-4 透過性に対する PLA の影響 ・・・ 49 3-3-2 エンドサイトーシス阻害剤存在下での Caco-2 細胞 sheet における TJ 関連タンパク質の局在性に対する PLA の影響 ・・・ 51 3-3-3 クラスリン介在性エンドサイトーシス阻害剤 剤存在下での Caco-2 細胞 sheet における erythritol 透過性に対する PLA の影響 ・・・ 55 3-3-4 PLA 適用後における occludin の細胞内局在部位の同定 ・・・ 56 第 4 節 小括 ・・・ 58 第 3 編 PLA により引き起こされる Caco-2 細胞内のリン酸化タンパク質の同定 並びに機能解析 第 1 節 小緒言 ・・・ 60 第 2 節 実験方法 4-2-1 試薬及び抗体 ・・・ 62 4-2-2 細胞培養 ・・・ 62 4-2-3 抗体アレイ法 ・・・ 62 4-2-4 MAPK 阻害剤の調製 ・・・ 63 4-2-5 TEER 測定並びに FD-4 透過実験 ・・・ 63 4-2-6 Recovery 試験 ・・・ 64 4-2-7 Immunoblot analysis ・・・ 64 4-2-8 統計解析 ・・・ 65 第 3 節 結果 4-3-1 PLA 適用による各種タンパク質の リン酸化レベルの解析 ・・・ 66 4-3-2 PLA による水溶性高分子薬物の透過促進効果 に対する MAPK 阻害剤の影響 ・・・ 69 4-3-3 TJ の再形成と MAPK の関連性 ・・・ 70
第 4 節 小括 ・・・ 73
結論 ・・・ 75
謝辞 ・・・ 80
1
緒言
一般に薬物の脂溶性度の低下や分子量の増加に伴い、上皮粘膜からの薬物の吸 収性は低下する。特に生理活性ペプチドやタンパク質性医薬品のような水溶性 高分子薬物は、上皮粘膜からほとんど吸収されず、生物学的利用率は極めて低 い。従って、このような薬物の投与には注射といった侵襲的な方法しかなく、 利便性が低い上に患者への負担が大きいと考えられる。そのため、水溶性高分 子薬物を上皮粘膜から効率的に吸収させるドラッグデリバリーシステムは、注 射による投与法の代替システムとして注目されている。上皮粘膜からの水溶性 高分子薬物の吸収を高める方策の 1 つに吸収促進剤の利用が挙げられる。これ までに界面活性剤や胆汁酸、脂肪酸などの様々な物質が吸収促進剤の候補とし て検討されてきた1-5)。しかし、それらのほとんどは薬物の吸収は大きく改善す るものの、膜タンパク質の漏出や細胞の脱落等の傷害性が認められ、それらの 吸収促進剤候補物質による薬物吸収の増大と粘膜に対する傷害性の間にはパラ レルな関係があると報告されている3,6)。 近年、新たな吸収促進剤の候補物質として、chitosan やその誘導体、dendrimer のようなポリカチオン性物質や NO 供与体、claudin modulator が見出され、これ らは上皮粘膜にほとんど傷害を与えることなく、水溶性高分子薬物の吸収を改 善することが報告されている7-11)。 このような背景の中、Natsume らは水溶性高分子薬物の吸収を改善させる吸収促進剤の候補物質として poly-L-arginine (PLA) を見出し12)、検討を行ってきた。
これまでの研究で、PLA はラット鼻粘膜において、モデル水溶性高分子薬物で ある fluorescence isothiocyanate dextrans (FD-4, 10, 40, 70, 150, MW 4,400-16,7000)
2
及び recombinant human granulocyte colony-stimulating factor (rhG-CSF) の吸収性
を改善することが報告されている 13,14)。また、家兎摘出鼻粘膜を用いた研究で
は、FD-4 の吸収性の改善が主に細胞間隙経路を介したものであることが明らか
となっている15)。これは、PLA が細胞間隙部位に存在する tight junction (TJ) 関
連タンパク質の occludin 及び ZO-1、adherence junction (AJ) 関連タンパク質の E-cadherin を細胞間隙部位から消失させ、細胞間隙経路が開口することにより、
FD-4 の吸収を増加させる15)。この開口メカニズムのさらなる検討の結果、TJ 関
連タンパク質の occludin の脱リン酸化と ZO-1 のリン酸化が関係しており 16)、
PLA の吸収促進メカニズムには kinase や phosphatase のようなリン酸化・脱リン
酸化反応に深く関わる酵素が関与することが予想された。しかし、家兎摘出鼻 粘膜を用いた研究では、動物間での抗体の交差性、タンパク質量の不足により、 さらなる詳細な解析が困難であった。そのために、PLA による細胞間隙経路の 開口メカニズムに関しては未だに不明な点が多い。 TJ は上皮細胞や内皮細胞シートにおいて、アピカル側に認められ、隣接する 細胞同士を接着している。TJ の形成は Furuse らにより同定されたタンパク質群 である TJ 関連タンパク質が重要な役割を果たしており、TJ 関連タンパク質とし て、四回膜貫通タンパク質である claudins が TJ の構築に重要な要素の一つであ ることが示されている17,18)。Claudins は 27 のファミリーから成り、claudins の発 現により細胞間隙の物質の透過が制御されていることが示されている 19,20)。ま
た、tight junction associated MARVEL proteins (TAMPs) と呼ばれるファミリーも 存在する。これも claudins と同様に四回膜貫通タンパク質であり、occludin や tricellulin、MARVEL D3 が知られている21-23)。Occludin は TJ 関連の膜貫通タン パク質として、最初に発見されたタンパク質であり、多くの研究が行われてき
3
クアウトモデルにおいて、occludin が TJ の形成や維持において必須のタンパク
質ではないことが明らかとなった 27,28)。その一方で、水溶性高分子薬物の透過
の制御において、occludin は重要な役割を果たしていることが明らかとなってい
る 29,30)。加えて、細胞シート上には 3 つの細胞が交わる点にも TJ が存在する。
このような部位には claudins や occludin ではなく、tricellulin と呼ばれる四回膜貫 通タンパク質が発現しており、三細胞間における物質の透過を制御している。 マウス乳腺上皮細胞株の Eph4 を用いた RNA 干渉による tricellulin のノックダウ ン研究では、三細胞間のみならず、細胞間の TJ の構造も破綻することから、TJ
の形成においても重要な役割を持つことが予想されている 22)。また、上記に示
した TJ 関連の膜貫通タンパク質は、細胞質において、ZO-1 や multiple PDZ domain protein (MUPP1) のような scaffold タンパク質と結合し、アクチン骨格に繋ぎ止
められている31-33)。
これまでに、カプリン酸ナトリウムが PLC 依存的な IP3/DAG 経路及び MLCK
を介して細胞間隙経路を開口させることが報告されている34)。Claudin modulator
は直接的に TJ 関連タンパク質である claudin-4 と結合することで、claudin-4 を
TJ から選択的かつ可逆的に消失させる35)。ポリカチオン系の吸収促進剤である
chitosan では、インテグリン受容体を介した Src 及び Focal adhesion kinase (FAK)
の活性化により、細胞間隙経路の開口が誘導されることが報告されている 36)。 一方、PLA による細胞間隙経路の開口に関わる詳細なメカニズムは不明である が、細胞間隙経路の開口は、TJ 関連タンパク質のリン酸化及び脱リン酸化によ り引き起こされ、それらタンパク質が細胞内へ移動して TJ 部から消失すること が予想される。従って、TJ 関連タンパク質の局在性や発現量の変化、細胞内で 変化が認められるタンパク質のリン酸化レベルの変化と細胞間隙透過性との関 連性を検討することは、PLA の吸収促進メカニズムの解析を行う上で極めて重
4 要である。 本研究の目的は、これまでに明らかにされていない PLA による水溶性高分子 薬物の吸収促進メカニズムを明らかにすることである。そのため、TJ のメカニ ズム解析に汎用されている系の 1 つである Caco-2 細胞を中心に検討を行うこと にした。Caco-2 細胞はヒト結腸癌由来の細胞株であり、薬物の透過並びに TJ の 機能を評価する研究に数多く使用されているため、情報も豊富である。すなわ ち、Caco-2 細胞は PLA の細胞間隙開口メカニズムを評価する上で非常に有用な 材料であると考えられる。しかし、PLA がラット鼻粘膜や家兎摘出鼻粘膜と同 様に腸管上皮においても吸収促進効果を示すかどうかはほとんど明らかとなっ ていない。そこで、第 1 編では in vivo ラット腸管及び in vitro 培養細胞評価系を 用いて PLA による水溶性高分子薬物の吸収促進効果を評価することを目的とし て、第 1 編第 1 章では、ラット腸管各部位に対する PLA の吸収促進効果を、in situ closed loop 法にて検討した。また、第 1 編第 2 章では in vitro での評価系として
Caco-2 細胞 sheet を用いて、水溶性高分子薬物の透過性、細胞傷害性及び TJ 関 連タンパク質の局在性並びに発現量に対する PLA の影響を検討した。これらの 検討により、PLA がラット鼻粘膜や家兎摘出鼻粘膜と同様に水溶性高分子薬物 の吸収促進効果を示すかどうかを明らかにした。第 2 編では、PLA により引き 起こされる TJ 関連タンパク質の消失機構に関して検討した。一般的に TJ 関連 タンパク質の細胞間隙部からの消失には、TJ 関連タンパク質の分解やエンドサ イトーシスによる細胞内への内在化が知られている。そこで、PLA による水溶 性高分子薬物の FD-4 の透過促進効果及び TJ 関連タンパク質の局在性の変化に 対するエンドサイトーシス阻害剤の影響を評価し、どのようなメカニズムで TJ 関連タンパク質が消失するかを明らかにした。第 3 編では、PLA により引き起 こされる細胞間隙経路の開口及びその後の TJ の再形成には細胞内でのタンパク
5 質のリン酸化及び脱リン酸化が深く関与していることが予想されたため、PLA の適用によりリン酸化レベルの変化が認められるタンパク質を抗体アレイ法を 用いて評価した。さらに、抗体アレイ法で同定されたリン酸化タンパク質に対 する特異的な阻害剤を用いて、PLA による細胞間隙経路の開口との関連性、並 びに TJ の再形成との関連性を調査した。
6
第 1 編 水溶性高分子薬物の in vivo ラット腸管からの吸収及び
in vitro Caco-2 細胞 sheet を介した透過に対する PLA の効果
第 1 章 種々腸管部位からの FD-4 の吸収に対する PLA の促進効果
第 1 節 小緒言
Ohtake ら及び Natsume らは、既に in vitro 家兎摘出鼻粘膜並びに in vivo ラット 鼻粘膜において、PLA がモデル水溶性高分子薬物である FD-4 の吸収を改善する ことを報告している 14,15)。しかし、PLA の吸収促進効果は鼻粘膜上皮細胞及び 眼組織37) において研究されているが、消化管各部位において、PLA が水溶性高 分子薬物の吸収を促進するかどうかはほとんど検討されていない。さらに、腸 管粘膜は鼻粘膜上皮とは異なり、食物由来の様々な物質を効率的に取り込むこ とに適した構造、機能を有している。主に水分を吸収するために微絨毛が発達 していない結腸に対し、主な薬物吸収部位である小腸前半部分の十二指腸や空 腸は微絨毛構造が発達しており、表面積が著しく大きい。そのため、PLA によ る薬物の吸収促進効果には部位差が生じることが予想される。加えて、腸管部 位ごとに細胞間隙に存在する TJ 関連タンパク質の発現の違いや消化酵素の存在 も無視することはできない。特に消化酵素の存在は PLA の分解を引き起こすこ とがこれまでに示されている38)。 第 1 章では、腸管上皮細胞においても PLA が吸収促進効果を示すかどうかを 確認するために、水溶性高分子モデル薬物である FD-4 の吸収促進効果を in situ closed loop 法を用いてラット腸管で評価した。さらに、PLA による吸収促進作
7
討した。また、腸管各部位において高分子薬物の透過に重要な役割を果たして いる occludin の発現量をウエスタンブロット法を用いて評価し、PLA による吸 収促進効果の部位差の解明を試みた。
8
第 2 節 実験方法
1-2-1. 試薬
Fluorescein isothiocyanete (FITC) – dextran (MW 3.75 kDa, FD-4), poly-L-arginine (MW44.3 kDa, PLA) 、 aprotinin from bovine serum 及 び urethane を SIGMA-ALDRICH (St. Louis, MO) より購入した。ドデシル硫酸ナトリウム
(SDS) を和光純薬工業株式会社 (Osaka, Japan) より購入した。BCA protein assay
reagent kit 及び Super Signal West Dura Extended Substrate を Thermo scientific
(Waltham, MA) より購入した。Mouse anti occludin を Invitrogen (Carlsbad, CA) よ
り購入した。 HRP-conjugated goat anti mouse IgG を Santa Cruz (Santa Cruz, CA) より購入した。その他の試薬は特級品を用いた。
1-2-2. 動物実験
Wistar 系雄性ラット (8 週齢, BW 250 g – 300 g) を埼玉実験動物供給所 (Saitama, Japan) から購入した。ラットを 3 匹/cage で飼育し、自由に食餌及び水
を摂取できるようにした。馴化のために 1 週間飼育し、実験 16 時間前から絶食 させた。本研究におけるラットを用いた全ての実験は、城西大学動物実験規定 に従い、全学動物実験管理委員会の承認を得て行った。 1-2-3. 静脈内投与 Wistar 系雄性ラットの腹腔内に urethane (1 g/kg, 250 mg/mL) を投与し、麻酔し た。ラットを固定台に仰臥位固定した後、腸管投与と同様の条件にするため、 腸管へ外科的処置を施し、左右の頚静脈を露出した。調製した FD-4 生理食塩液 (2 mg/kg, 2 mg/mL) を右頚静脈より投与した。
9 1-2-4. In situ closed loop 実験
Wistar 系雄性ラットを urethane (1 g/kg, 250 mg/mL) の腹腔内投与により麻酔 した後、開腹手術を行った。胃幽門部から下 5 cm を十二指腸部、胃幽門部から 下 10 cm から 15 cm までを空腸部、盲腸の上 5 cm から 10 cm までを回腸部、盲 腸より下 5 cm を結腸部とし、それぞれ 5 cm の loop を作製した。作製した loop の両端に切り込みを入れ、37℃に温めた生理食塩液 20 mL と空気 5 mL をシリン ジで注入し、各腸管内容物を洗浄除去した。十二指腸への手術の際には、loop 内への胆汁の流入を防ぐため、胆管部にメディカルポリエチレンチューブ (内径 0.28 mm, 外形 0.61 mm) を挿入して作製した Bile fistula から胆汁を体外へ排出 させた。FD-4 溶液 (20 mg/kg, 10 mg/mL), PLA (0.05, 0.25 及び 0.5%) 含有 FD-4 溶液 (20 mg/kg, 10 mg/mL), 100 µg/mL aprotinin 含有 FD-4 溶液 (20 mg/kg, 10 mg/mL) 及び 100 µg/mL aprotinin と 0.5% PLA 含有 FD-4 溶液 (20 mg/kg, 10 mg/mL) を各部位の腸管 loop 末端から経口用ゾンデを用いて、腸管 1 cm あたり 100 μL 投与した。体温を維持するために作製したループを体内へ戻し、ミヘル 針で閉腹した。 1-2-5. 血漿採取法 投与後、へパリンで処理した注射筒を用いて、左頚静脈より 0.15 mL ずつ経時 的 (0, 5, 15, 30, 45, 60, 90, 120, 180 及び 240 分) に血液を採取した。得られた血 液を直ちに遠心分離 (20,400 x g, 5 min, 4°C) し、血漿を得た。 1-2-6. 血漿中 FD-4 濃度測定 血漿 50 μL をホウ砂リン酸緩衝液 (pH 8.5) で 100 倍希釈し、サンプル中の FD-4 の蛍光強度を分光蛍光光度計 (RF-1500, Shimadzu, Kyoto) を用いて励起波
10 長 495 nm、蛍光波長 515 nm で測定した。 1-2-7. 動態学的解析 FD-4 の体内動態を 2-コンパートメントモデルに従って解析した。血中濃度を 非線形最小二乗法 (アルゴリズム;Damping Gauss-Newton 法) により解析した。 静脈内および腸管内投与における最高血中濃度 (Cmax)、最高血中濃度到達時間 (Tmax) を血中濃度プロファイルより求めた。モーメント解析法により血漿中濃度 -時間曲線下面積 (AUC0-4 h) を (1) 式から算出した。
AUC
0-4 h=
∫ Cdt
04 ・・・(1) ここで、C は血漿中 FD-4 濃度 (μg/mL) である。 また、生物学的利用率 (F0-4 h) を (2) 式より算出した。F
0-4 h(%)=
AUCtest⁄DtestAUCi.v.⁄Di.v.
×100
×ここで、Dtest及び Di.v. はそれぞれ腸管内及び静脈内投与量であり、AUCtest及び
AUCi.v. はそれぞれ腸管内及び静脈内投与における血漿中濃度-時間曲線下面積 (AUC) である。 1-2-8. Immunoblot analysis ラット腸管 (十二指腸、空腸、回腸、結腸) を 5 cm 摘出し、生理食塩液 20 mL で腸管内を洗浄後、切開した。粘膜側を上にして氷上 phosphate-buffered saline ・・・(2)
11
(PBS) 内に配置した擦りガラスの上に置き、もう一枚の擦りガラスでこすり合
わせながら上皮細胞を採取した。採取した腸管上皮細胞を液体窒素で満たした 乳鉢に加えて凍結し、ホモジナイズした。Tris buffer 溶液 (20 mM Tris-HCl pH 7.4, 5 mM MgCl2, 1 mM EDTA, 0.3 mM EGTA, protease inhibitor cocktail) を 1 mL 加え、
26 ゲージの針と注射筒 (1 mL) を用いて全量の吸い取りと吐き出しを 5 回繰り
返し、さらにホモジナイズした。そのホモジネートを遠心分離 (1,000 x g, 5 min) し、未破砕細胞などを沈殿させ、上清のみを採取した。その後、上清を超遠心 分離 (43,000 x g, 30 min) し、沈殿したペレットを膜フラクションとし、上記の Tris buffer 溶液を加えて再懸濁した。BCA protein assay を用いてタンパク質量を
均一に補正した。サンプルを 2 × SDS sample buffer と 1 : 1 の割合で混合し、95°C の 水 浴 で 10 分 間 加 熱 し 、 こ れ を SDS-PAGE の サ ン プ ル と し た 。 12%
polyacrylamide gel 内へ調製したサンプル 10 μL を添加し、電気泳動を行った (200
V, 40 min)。その後、泳動したゲルを polyvinylidene fluoride (PVDF) 膜に転写した
(70 V, 120 min)。PVDF 膜を 3%スキムミルクを含む PBS-T (Phosphate-buffered
saline containing 0.1% Tween20) でブロッキング (室温、60 min) した後、PVDF
膜を occludin (1 : 1000) 及びβ-actin (1 : 1000) に対する一次抗体を含む PBS-T 中 で 4°C で一晩インキュベートした。インキュベート後、PBS-T で 15 分間振盪し て洗浄し、これを 3 回繰り返した。続いて、これらの一次抗体に対する二次抗 体 (HRP-conjugated anti mouse IgG, 1:1000) を含む PBS-T で PVDF 膜を室温で 1 時間インキュベートした。PVDF 膜を PBS-T で 15 分間振盪して洗浄し、これを 3 回繰り返した。Super Signal West Dura の enhancer solution と stable peroxide
solution を 1 : 1 で混合後、PVDF 膜へ塗布し、生じた化学発光を LAS-1000 (GE
Healthcare, UK) を用いて検出した。Occludin 及びβ-actin のバンド強度を Image J software program (NIH) を用いて定量し、各サンプルの occludin のピクセル強度
12
をβ-actin のピクセル強度で補正した。
1-2-9. 統計解析
2 群間の比較には Student’s t-test を用いて解析し、p < 0.05 を有意とみなし
13
第 3 節 結果
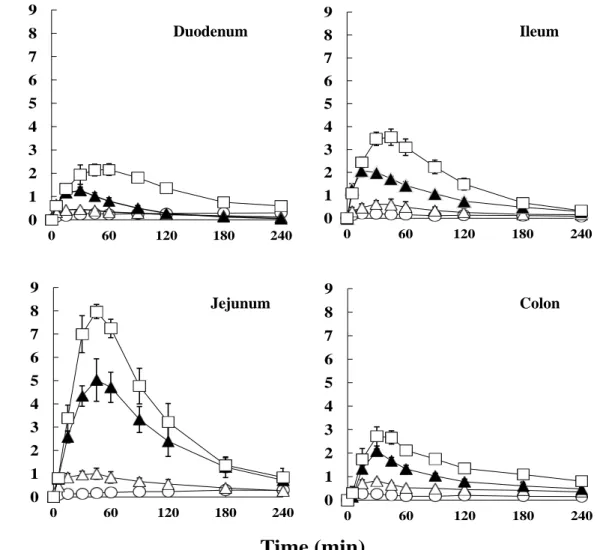
1-3-1. 腸管各部位における FD-4 の吸収に対する PLA の効果 Figure 1 にラット腸管各部位に種々濃度の PLA を FD-4 と共に投与した後の 血漿中 FD-4 濃度推移を、Table 1 に得られた FD-4 の動態学的パラメータを示す。 全ての腸管部位において、PLA は control と比較して FD-4 の吸収を増大させた。 また、PLA の濃度に依存して吸収促進効果は増大した。0.5% PLA を投与した時、 腸管各部位のバイオアベイラビリティの順序は、回腸 > 空腸 > 十二指腸 ≧ 結 腸であった。これらの結果は、PLA が濃度依存的に FD-4 の腸管吸収を促進し、 さらに、吸収促進効果には部位差があることを示唆している。14 0 1 2 3 4 5 6 7 8 9 0 60 120 180 240 0 1 2 3 4 5 6 7 8 9 0 60 120 180 240 0 1 2 3 4 5 6 7 8 9 0 60 120 180 240 0 1 2 3 4 5 6 7 8 9 0 60 120 180 240
P
las
ma c
on
ce
n
tr
ati
on
of
F
D
-4 (
μg
/mL
)
Time (min)Fig. 1 The plasma concentrations of FD-4 following intra-intestinal administration into the closed-loop in rats.
● : Control (FD-4 only), ○ : FD-4 with 0.05% PLA, △ : FD-4 with 0.25% PLA, □ : FD-4 with 0.5% PLA,
Each data point represents the mean and standard error (n=3-7)
a
Duodenum Ileum
15
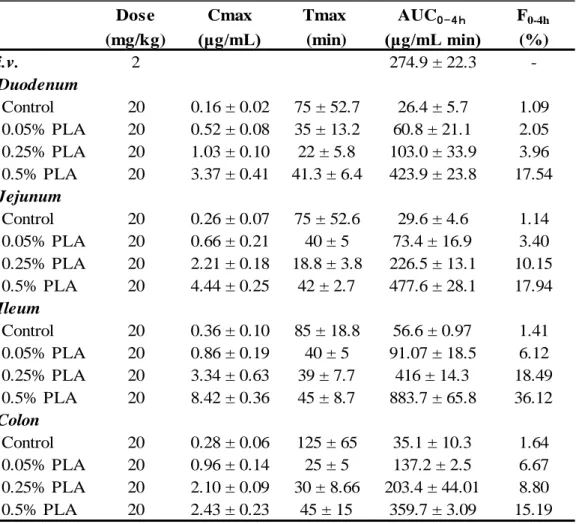
Dose Cmax Tmax AUC0 - 4 h F0-4h
(mg/kg) (μg/mL) (min) (μg/mL min) (%) i.v. 2 274.9 ± 22.3 -Duodenum Control 20 0.16 ± 0.02 75 ± 52.7 26.4 ± 5.7 1.09 0.05% PLA 20 0.52 ± 0.08 35 ± 13.2 60.8 ± 21.1 2.05 0.25% PLA 20 1.03 ± 0.10 22 ± 5.8 103.0 ± 33.9 3.96 0.5% PLA 20 3.37 ± 0.41 41.3 ± 6.4 423.9 ± 23.8 17.54 Jejunum Control 20 0.26 ± 0.07 75 ± 52.6 29.6 ± 4.6 1.14 0.05% PLA 20 0.66 ± 0.21 40 ± 5 73.4 ± 16.9 3.40 0.25% PLA 20 2.21 ± 0.18 18.8 ± 3.8 226.5 ± 13.1 10.15 0.5% PLA 20 4.44 ± 0.25 42 ± 2.7 477.6 ± 28.1 17.94 Ileum Control 20 0.36 ± 0.10 85 ± 18.8 56.6 ± 0.97 1.41 0.05% PLA 20 0.86 ± 0.19 40 ± 5 91.07 ± 18.5 6.12 0.25% PLA 20 3.34 ± 0.63 39 ± 7.7 416 ± 14.3 18.49 0.5% PLA 20 8.42 ± 0.36 45 ± 8.7 883.7 ± 65.8 36.12 Colon Control 20 0.28 ± 0.06 125 ± 65 35.1 ± 10.3 1.64 0.05% PLA 20 0.96 ± 0.14 25 ± 5 137.2 ± 2.5 6.67 0.25% PLA 20 2.10 ± 0.09 30 ± 8.66 203.4 ± 44.01 8.80 0.5% PLA 20 2.43 ± 0.23 45 ± 15 359.7 ± 3.09 15.19
Table 1 Pharmacokinetic parameters of FD-4 following intra-intestinal administration into the closed loop in rats.
a: F0-4 h = (AUCtest / Dosetest) / (AUCi.v. / Dosei.v.) × 100
16
1-3-2. 腸管各部位における PLA の吸収促進効果に対するタンパク質分解酵素
阻害剤の影響
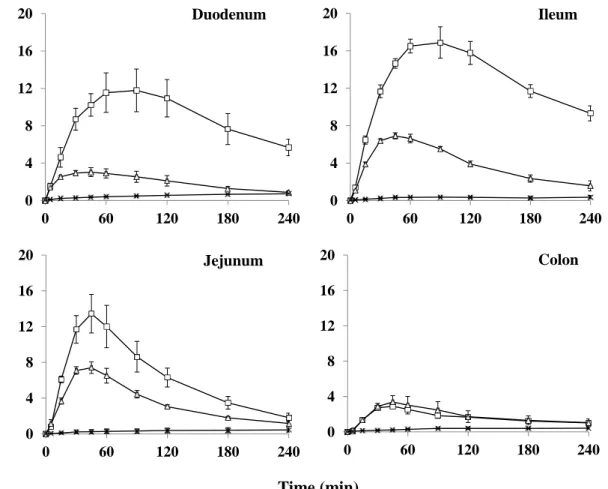
Figure 2 にラット腸管各部位に 0.5% PLA 及び aprotinin を FD-4 と共に投与し た後の血漿中 FD-4 濃度推移を、Table 2 に得られた FD-4 の動態学的パラメータ を示す。なお、FD-4 は酵素的な分解が認められていない39)。0.5% PLA 溶液に aprotinin を加えた FD-4 溶液を投与した場合、十二指腸、空腸及び回腸のいずれ の部位においても、0.5% PLA を含む FD-4 と比較して血漿中 FD-4 濃度は増大し た。特に十二指腸及び空腸において、バイオアベイラビリティが約 3.5 倍増加し た。また、回腸は、1.8 倍程度であった。これに対し、結腸は aprotinin の有無に かかわらず、FD-4 の血中濃度推移がほとんど変化しなかった。この結果は、十 二指腸、空腸及び回腸において、FD-4 の吸収促進効果の部位差を生じる要因の 一つとして、PLA の分解速度の違いにあることを示唆している。
17 0 4 8 12 16 20 0 60 120 180 240 0 4 8 12 16 20 0 60 120 180 240 Time (min) P la sm a co ncent ra tio n o f F D -4 ( µ g/ m L ) 0 4 8 12 16 20 0 60 120 180 240 0 4 8 12 16 20 0 60 120 180 240
Fig. 2 Plasma concentrations of FD-4 following administration at various sites of the intestinal tract in rats.
×: Control (FD-4), △ : FD-4 with 0.5 w/v% PLA , □ : FD-4 with 0.5 w/v% PLA and 100 µg/mL aprotinin Each data point represents the mean ± S.E. (n= 3-4).
Duodenum Ileum
18
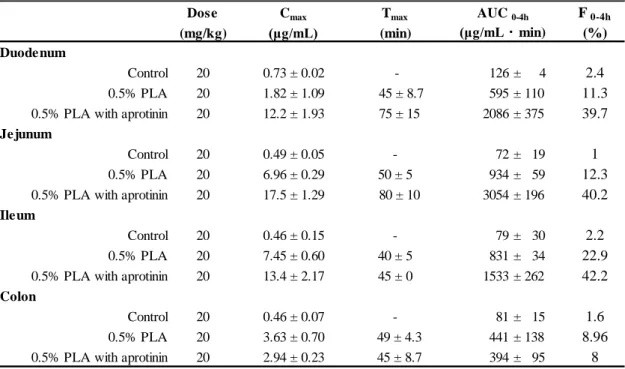
Table 2 Pharmacokinetic parameters of FD-4 following intra-intestinal administration into the closed loop in rats.
Dose Cmax Tmax F 0-4h
(mg/kg) (μg/mL) (min) (%)
Duodenum
Control 20 0.73 ± 0.02 - 126 ± 4 2.4
0.5% PLA 20 1.82 ± 1.09 45 ± 8.7 595 ± 110 11.3
0.5% PLA with aprotinin 20 12.2 ± 1.93 75 ± 15 2086 ± 375 39.7
Jejunum
Control 20 0.49 ± 0.05 - 72 ± 19 1
0.5% PLA 20 6.96 ± 0.29 50 ± 5 934 ± 59 12.3
0.5% PLA with aprotinin 20 17.5 ± 1.29 80 ± 10 3054 ± 196 40.2
Ileum
Control 20 0.46 ± 0.15 - 79 ± 30 2.2
0.5% PLA 20 7.45 ± 0.60 40 ± 5 831 ± 34 22.9
0.5% PLA with aprotinin 20 13.4 ± 2.17 45 ± 0 1533 ± 262 42.2
Colon
Control 20 0.46 ± 0.07 - 81 ± 15 1.6
0.5% PLA 20 3.63 ± 0.70 49 ± 4.3 441 ± 138 8.96
0.5% PLA with aprotinin 20 2.94 ± 0.23 45 ± 8.7 394 ± 95 8
AUC 0-4h
(μg/mL・min)
a: F0-4 h = (AUCtest / Dosetest) / (AUCi.v. / Dosei.v.) × 100
19 1-3-3. 腸管各部位における occludin の発現量 Figure 3 に腸管各部位の上皮細胞における occludin の発現量を示す。ラット腸 管各部位の上皮細胞における occludin の発現レベルは、十二指腸、空腸、回腸 ではほぼ同等であったが、結腸において強く発現していた。 0 0.4 0.8 1.2 1.6 2 2.4 Oc c lu d in p ix e l de ns it y β-actin Occludin
Fig. 3 The expression of occludin in the epithelium of the rat intestinal segments.
(A) Occludin and β-actin were detected by a Western blot analysis.
(B) The densitometry ratios representing the occludin expression in the epithelium. Each data column represents the mean and standard error (n=9).
(B) (A)
20
第4節 小括
ラット腸管各部位において、PLA は濃度依存的に FD-4 の吸収を増加させた。 また、PLA は回腸において最も高い吸収促進効果を示し、次いで空腸、十二指 腸と結腸の順であった。この結果は鼻粘膜と同様に腸管上皮においても PLA は 水溶性高分子薬物の吸収を改善できることを示唆している。 しかし、PLA による FD-4 の吸収促進効果には腸管各部位によって著しい違い が認められた。この吸収促進効果の部位差として考えられる要因には、タンパ ク質分解酵素量、TJ 関連タンパク質の発現レベル及び吸収表面積が腸管各部位 で異なることが挙げられる。結腸の吸収促進効果が最も小さかった主な要因と して、吸収表面積が他の部位に比べ小さいことが主な要因と考えられる 以前に、PLA を用いた FD-4 の経鼻吸収実験において、鼻腔内の鼻粘液中に含 まれる酵素により、PLA が分解されることが報告されている 39)。これと同様に 腸管上皮にも、膵臓由来のプロテアーゼ及び腸管上皮細胞から分泌されるプロ テアーゼによって PLA が分解される可能性がある。そこで、トリプシンやキモ トリプシンなどを阻害する aprotinin を用いて、FD-4 の吸収性に対する PLA の効 果の検討を行った。その結果、aprotinin 併用により、十二指腸、空腸及び回腸 における FD-4 のバイオアベイラビリティは PLA のみの適用と比較して、著し く増大した。この結果は十二指腸、空腸及び回腸において、PLA による吸収促 進効果に部位差が生じた主な要因が、消化酵素による PLA の分解であり、腸管 各部位での上皮粘液中のタンパク質分解酵素濃度が異なることにより40,41)、PLA の吸収促進効果が異なったと考えられた。すなわち、消化酵素の多い十二指腸 と小腸上部では、aprotinin が PLA を分解する酵素を阻害し、腸管内での PLA の 安定性が保たれ、上皮での PLA の濃度が高く維持されために、吸収促進効果が 改善し、FD-4 の血中への移行量が PLA 併用群と比較して増大したと考えられ21 た。 一方、結腸では消化酵素の濃度が小さいことから、aprotinin 併用による FD-4 吸収の増大は認められなかったと考えられる。 TJ 関連タンパク質の一つである occludin の発現量は結腸で最も高かったこと から、結腸での PLA による吸収促進効果が最も低かったのは、吸収表面積が小 さいことに加え、occludin の発現量が高いことも部分的に関与しているかもし れない。 以上より、腸管におけるPLAの吸収促進効果には部位差があり、それには吸収 表面積、消化酵素の分泌量、さらには部分的にoccludinの発現量の違いが関係し ていた。
22
第 2 章 Caco-2 細胞シートにおける経上皮電気抵抗、FD-4 透過性、
TJ 関連タンパク質の局在性及び発現量対する PLA の影響
第 1 節 小緒言
これまで PLA による透過促進メカニズムの解析は、家兎摘出鼻粘膜及びラッ ト鼻粘膜が用いられてきた。しかし、動物間での抗体の交差性やタンパク質量 の不足により、PLA による促進機構の更なる詳細な解析は困難であった。それ ゆえ、TJ の研究に汎用されている腸上皮を本研究で使用するには、PLA が腸粘 膜に対しても鼻粘膜上皮と同様に吸収促進効果を示すことを確認する必要があ る。そこで、腸管吸収実験を行った結果、鼻粘膜と同様に PLA は水溶性高分子 薬物の吸収を促進したため、PLA による水溶性高分子薬物の吸収促進メカニズ ムの更なる解析をヒト結腸癌由来の細胞株である Caco-2 細胞を用いて検討する ことにした。Caco-2 細胞は TJ 関連タンパク質に対する酸化的ストレスや細菌感 染の影響に関する研究や吸収促進剤の評価に数多く利用されているため、情報 も豊富である。すなわち、Caco-2 細胞は PLA の細胞間隙開口メカニズムを分子 生物学的に評価するのに、最適な材料であると考えられた。 そこで第 2 章では、PLA により引き起こされる水溶性高分子薬物の透過促進 メカニズムを明らかにするために、Caco-2 細胞シートにおける経上皮電気抵抗、 FD-4 透過性及び TJ 関連タンパク質の局在性に対する PLA の影響を検討した。 TJ 関連タンパク質として鼻粘膜上皮で検討した occludin 及び ZO-1 に加え、主 に低分子薬物やイオンの拡散を制限する重要なタンパク質である 4 回膜貫通タ ンパク質 claudin-4、3 細胞間での TJ の形成に重要な役割を果たしている tricellulin に関しても評価した。23
第 2 節 実験方法
2-2-1. 試薬及び抗体
Caco-2 細胞培養時に用いた Dulbecco’
s Modified Eagle Medium (DMEM)、牛胎
児血清 (FBS)、非必須アミノ酸 (NEAA)、Gluta-MAXTM
-1、Antibiotic-antimycotic、
Hank’s Balanced Salt Solution (HBSS)、HEPES Buffer Solution (HEPES) 及び
Trypsin-EDTA を GIBCO (MD, U.S.A) より購入した。Anti mouse occludin、anti
mouse claudin-4、anti rabbit ZO-1、anti rabbit tricellulin、Alexa Fluor 488 goat
anti-rabbit IgG、Alexa Fluor 594 donkey anti-mouse IgG を Invitrogen (Carlsbad, CA)
より、anti rabbit GAPDH、HRP-conjugated Goat anti-mouse IgG、HRP-conjugated Goat anti rabbit IgG を Santa Cruz (Santa cruz, CA) より購入した。その他の試薬は
市販の特級品を用いた。
2-2-2. 細胞培養
Caco-2 細胞を American Type Culture Collection (MD, U.S.A) より購入し、実験
には継代数 68-78 代の細胞を使用した。CO2インキュベーター内で 5% CO2存在
下、37°C で細胞を培養した。1 日もしくは 2 日おきに培地を交換し、細胞がセ ミコンフルエントの状態に達したとき Trypsin-EDTA を用いて継代した。
Caco-2 細胞 sheet の調製には、polycarbonate 膜製の Transwell(Corning, USA)
を使用した。Figure 4 に Transwell の構造を示す。1.0×105
cells/cm2になるように polycarbonate 膜上に細胞を播種した。培地換えは、実験に用いるまで 1 日もしく
24 2-2-3. 経上皮電気抵抗 (TEER) の測定
Transwell にて 21-28 日間培養し、TJ を形成した Caco-2 細胞 sheet を、HBSS
溶液にて 2 回洗浄した。洗浄後、Transwell の apical side に 1.5 mL、basal side に 2.6 mL の HBSS 溶液を加え、37°C で 60 分間プレインキュベートした。インキ
ュベーション後に測定した値を初期 TEER とした。なお、TEER の測定は、 Millicell®-ERS (Millipore, MA, U.S.A) を用いて測定し (Fig. 4)、Caco-2 細胞 sheet
の TEER を (3) 式より算出した。測定は、3 回ずつ行い、その平均値をその時間 の TEER とした。また、透過実験に用いた Caco-2 細胞 sheet の初期 TEER は、 500 Ω・cm2以上とした。
TEER (Ω・cm2
) = {(実測値)-(polycarbonate 膜の抵抗値)}× (膜の表面積) ・・・(3)
初期の TEER を測定した後、apical side に種々濃度の PLA を適用し、120 分ま で TEER を測定した。
PLA 除去後の TEER の回復の推移を調べるため、120 分経過後、HBSS 溶液で
PLA を washout し、apical side に fresh な HBSS 溶液を、basal side に DMEM 培地
を加え、37°C で 1 時間インキュベートした。インキュベーション後、通常の培
Apical side
Basal side
Polycarbonate membrane Electrode
25
地の培養状態に戻した。24 時間経過後に TEER を測定した後に、培地換えを行 い、さらに 24 時間後に TEER を測定した。
FD-4 のみかけの透過係数 (Papp) と TEER の関係を評価するために、TEER か
ら(4)式により膜コンダクタンス(Gt)を算出した。
Gt (mS/cm2) = 1/TEER (Ω・cm2) ・・・ (4)
2-2-4. FD-4 透過実験
Transwell にて 21-28 日間培養し、TJ を形成した Caco-2 細胞 sheet を、HBSS 溶液にて 2 回洗浄した。洗浄後、Transwell の apical side に 1.5 mL、basal side に 2.6 mL の HBSS 溶液を加え、37°C で 60 分間プレインキュベートした。インキ
ュベーション後、apical side に種々濃度の PLA と 2.5 mg/mL の FD-4 を適用し、 apical side より basal side へと透過した FD-4 量を定量した。120 分まで 15 分おき
に basal side の溶液を 0.2 mL サンプリングし、その後直ちに fresh な HBSS 溶液
を同量添加した。得られたサンプルは、ホウ砂リン酸緩衝液(0.2 M Na2B4O7, 0.2 M KH2PO4, pH 8.5)を用いて 100 倍希釈し、分光蛍光光度計を用いて FD-4 の蛍 光強度を測定した。測定波長は、励起波長(Ex)495 nm、蛍光波長(Em)515 nm で行った。FD-4 の Papp を(5)式より算出した。 ・・・ (5) ここで、dQ/dt (µg/sec) は FD-4 の定常状態フラックスであり、C0 (µg/mL) は apical
side の FD-4 初濃度、A (cm2) は、Caco-2 細胞 sheet の有効表面積である。本実験
時間内では sink 条件は保たれている。
Papp of FD-4 (cm/sec) = dQ/dt (A×C0)
26 2-2-5. FD-4 透過経路の観察
FD-4 透過実験終了後の Caco-2 細胞を HBSS で 2 回洗浄し、細胞を 4% paraformaldehyde-PBS を用いて、室温で 10 分間固定した。固定した細胞をカバ
ーガラスに載せ、退色防止剤である 1,4-diazabicyclo[2.2.2] octane を含む mowiol で封入した。FD-4 の蛍光像を共焦点レーザー走査型顕微鏡 (FV1000, Olympus, Tokyo) を用いて観察した。
2-2-6. MTT assay
種々濃度の PLA を Caco-2 細胞 sheet に 2 時間適用した。その後、細胞シート を HBSS で 2 回洗浄し、MTT 試薬 (1.5 mg/mL) を含む DMEM に置き換え、3 時間インキュベートした。生成したホルマザン色素を dimethyl sulfoxid (DMSO)
を用いて溶解した。得られたサンプルの吸光度を GENios microplate reader (Tecan,
Mannedorf, Switzerland) を用いて波長 535 nm で測定した。得られた吸光度から
(6) 式より Cell viability を算出した。
2-2-7. Immunofluorescence microscopy
PLA を濃度及び適用時間を変えて処理した Caco-2 細胞を acetone : methanol (1:1) を用いて、4°C で 10 分間固定した。その後、ブロッキング溶液として、3%
スキムミルク及び 0.1% Tween-20 を含む Tris buffered saline (10 mM Tris-HCl pH 7.2, 150 mM NaCl, TBS) で 1 時間インキュベートしてから、ブロッキング溶液で
希釈した一次抗体の occludin (dilution, 1:250), ZO-1(dilution, 1:125), claudin-4 (dilution, 1:250) 及び tricellulin (dilution, 1:125) を用いて、4°C で一晩インキュベ
Cell Viability (%) =
Absorbance
PLA27
ートした。TBS-T で 15 分間の洗浄を 3 回行い、ブロッキング溶液で希釈した二 次抗体 (Alexa fluor 488, 594, dilution, 1:500) で 1 時間インキュベートした後、細 胞を再び TBS-T で 15 分間の洗浄を 3 回行った。サンプルをスライドガラスに載 せ、退色防止剤を含む mowiol で封入した。サンプルの蛍光を FV1000 を用いて 観察した。さらに、TJ 関連タンパク質の蛍光密度を Gopalakrishnan らの方法を
用いて定量した42)。蛍光画像をランダムに選択し、バックグランド値を Image J
software program の threshold 機能を用いて補正した。このイメージを反転させ、
gray value 値を測定した。得られた値からバックグランドの値を引いたものを TJ
タンパク質の蛍光密度とした。
2-2-8. Immunoblot analysis
Caco-2 細胞を HBSS で 2 回洗浄し、RIPA buffer (25 mM Tris-HCl pH 7.5, 150 mM NaCl, 1.0% NP-40, 1.0% sodium deoxycholate, 0.1% SDS, protease inhibitor cocktail)
を用いてライセートを調製した。サンプル間のタンパク質量を補正するために、 BCA protein assay を用いてタンパク質量を測定した。それぞれのサンプルを 2 ×
SDS sample buffer と同量混合し、95°C で 10 分間加熱した。その後、12%
SDS-polyacrylamide gel を用いてタンパク質を分離し(200 V, 40 min)、PVDF 膜へ
転写した (70 V, 120 min)。この PVDF 膜を 3%スキムミルクを含む PBS-T で 1 時 間インキュベートし、洗浄後、一次抗体である occludin (dilution, 1:1000), ZO-1 (dilution, 1:500), claudin-4 (dilution, 1:1000), tricellulin (dilution, 1:1000) 及 び
GAPDH (dilution, 1:1000) を用いて、4°C で一晩インキュベートした。PVDF 膜を
再び PBS-T で 15 分間洗浄し、これを 3 回行った。洗浄後、これらの一次抗体に 対する HRP-conjugated 二次抗体 (dilution, 1:1000) で 1 時間インキュベートした。
28
Signal West Dura を塗布し、生じた化学発光を LAS-1000 を用いて検出した。TJ
関連タンパク質及び GAPDH のバンド強度を Image J software program を用いて 定量し、GAPDH で補正を行った。
2-2-9. 統計解析
29
第 3 節 結果
2-3-1 Caco-2 細胞 sheet における TEER 及び FD-4 透過性に対する PLA の影響
Figure 4A 及び B に 10、25、50 及び 100 µg/mL PLA を Caco-2 細胞の apical side に適用した際の TEER 及び FD-4 透過量を示す。10、25、50 及び 100 µg/mL PLA を Caco-2 細胞 sheet に適用すると、TEER は適用直後より急激に低下し、適用後 45 分までにそれぞれコントロールの 90、58、45 及び 39%になり、PLA の濃度 に依存して低下した。一方、種々濃度の PLA を適用したときの FD-4 の透過量 は、PLA 濃度の増加に伴い増大した。しかし、10 µg/mL PLA では、FD-4 透過量 はほとんどコントロールと変わらなかった。Figure 4C に Fig. 4B の FD-4 透過量 から算出した見かけの透過係数 (Papp) を示す。10、25、50 及び 100 µg/mL PLA の適用で、Papp は Control に比べ、それぞれ 1.2、4.0、17 及び 31 倍と高い値を 示し、25 µg/mL 以上の PLA 適用で、有意に増加した (p < 0.05) 。加えて、FD-4 の Papp と Gt の間には有意な直線関係が認められた (Fig. 4D, p < 0.05) 。
30
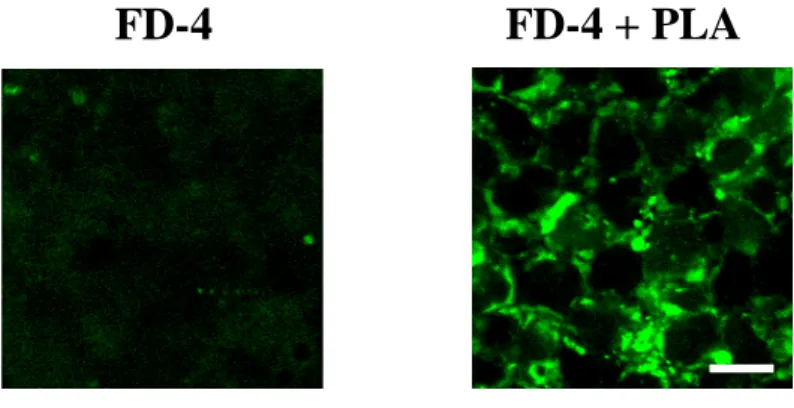
Figure 5 に FD-4 の透過経路を観察した結果を示す。PLA を適用すると、細胞
間隙部位に FD-4 の蛍光の濃染像が観察されたが、FD-4 単独適用ではほとんど 観察されなかった。
Fig. 4 Effects of PLA on the TEER and FD-4 permeability in the Caco-2 cell monolayer.
The change in the TEER (A), amount of FD-4 that crossed the cell layer (B) and the Papp of FD-4 (C) after treatment of Caco-2 monolayers with PLA (, 0 μg/mL, , 10 μg/mL, , 25 μg/mL, , 50 μg/mL, , 100 μg/mL). Relationship between Papp of FD-4 and Gt (D). Each data point represents the mean ± S.E. (n=4). * p < 0.05 vs. Control.
0 20 40 60 80 100 120 0 15 30 45 60 75 90 105 120 TEER (% o f in iti al v al u e ) Time (min) (A) (B) (C) Pa p p o f FD -4 ( × 10 7c m /s ) Time (min) FD -4 P e r m e a te d (μ g/cm 2)
*
*
*
0 5 10 15 20 0 15 30 45 60 75 90 105 120 0 2.5 5 7.5 10 12.5 Concentration (µg/mL) 0 10 25 50 100 Pa p p o f FD -4 ( × 10 7c m /s e c ) 0 1 2 3 4 0 2 4 6 8 10 12 14 (D) 1/TEER r2= 0.926 p<0.05 Gt (mS/cm2)31 Concentration (µg/mL) C ell v ia b ilit y ( % o f co n tro l)
0
10
25
50
100
0
20
40
60
80
100
120
Figure 6 に種々濃度の PLA を 2 時間適用したときの細胞生存率を示す。いず れの濃度の PLA の適用においても、Caco-2 細胞の生存率はほぼ 100%であり、 Caco-2 細胞に対して PLA は少なくとも 100 µg/mL まで細胞傷害性を示さないこ とが明らかとなった。これらの 結果から、Caco-2 細胞 sheet にお いて、PLA は細胞傷害性を示す ことなく、細胞間隙経路の開口 を介して、FD-4 の透過を促進す ることが示唆された。
Fig. 5 Confocal laser scanning microscopic observation of Caco-2 cell monolayers after administration of FD-4 with 100 μg/mL PLA. Bar = 5 μm
Fig. 6 Effect of PLA on cell viability.
Caco-2 cell monolayers were treated with PLA for 2h Each data represents the mean ± S.E. (n = 4)
FD-4 + PLA
FD-4
32
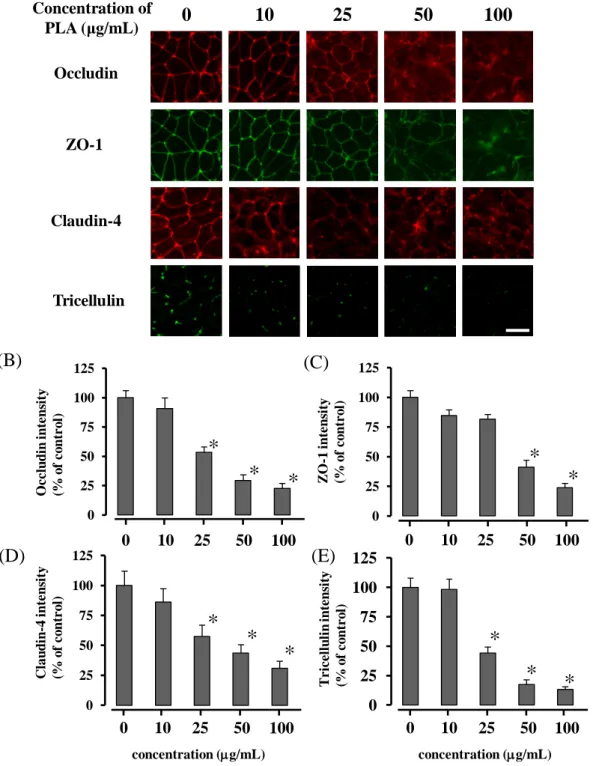
2-3-2 Caco-2 細胞 sheet の TJ 関連タンパク質の局在性に対する PLA の影響
Figure 7A に種々濃度の PLA を 2 時間適用した後の occludin、 ZO-1、claudin-4
及び tricellulin の免疫蛍光染色画像を、Figs. 7B-E にそれぞれの TJ 関連タンパク
質の細胞間隙における蛍光強度を示す。PLA を適用していない (PLA 0 μg/mL)
Caco-2 細胞において、occludin、ZO-1 及び claudin-4 は細胞間隙に局在し、tricellulin
は三細胞間に局在していた。一方、種々濃度の PLA の適用により、細胞間隙で の TJ 関連タンパク質の局在性は低下した。その効果は PLA 濃度依存的であり、 ZO-1 においては PLA 50 μg/mL以上の適用で、その他の TJ 関連タンパク質では、 PLA 25 μg/mL 以上の適用で有意に低下した (p < 0.05) 。特に、100 µg/mL PLA において、いずれの TJ 関連タンパク質の蛍光は細胞間隙でほとんど観察されな かった。また、FD-4 の透過が有意に増大する PLA 濃度 (25 μg/mL) において、
occludin 及び tricellulin は、claudin-4 や ZO-1 と比較して細胞間隙からの消失が大
きかった。
Figure 8A に 50 µg/mL PLA 適用後の occludin、 ZO-1、claudin-4 及び tricellulin の局在性の経時的変化を、Figs. 8B-E にそれぞれの TJ 関連タンパク質の細胞間 隙における蛍光強度を示す。いずれの TJ 関連タンパク質も PLA の処理時間の延 長に伴い細胞間隙部位への局在性が低下し、適用後 30 分以上で有意に低下した (p < 0.05) 。特に occludin と tricellulin の低下が著しく、occludin においては、適
用後 15 分以上で有意に低下した (p < 0.05) 。Figure 4B の FD-4 の透過プロファ イルの定常状態の直線から X 切片に外挿して得た lag time は約 26 分であり、 occludin と tricellulin の蛍光強度が著しく低下したときの時間に近かった。
33 0 10 25 50 100 ZO-1 Occludin Concentration of PLA (μg/mL) Tricellulin Claudin-4 (A) 0 25 50 75 100 125 O c c lu di n int e nsi ty (% of c on tr ol )
*
*
*
0 10 25 50 100 ZO -1 i nt e nsi ty (% of c on tr ol ) 0 25 50 75 100 125*
*
0 10 25 50 100 0 25 50 75 100 125*
*
*
0 10 25 50 100 Cl a u di n -4 i nt e nsi ty (% of c on tr ol ) concentration (µg/mL)*
*
*
0 10 25 50 100 concentration (µg/mL) T ricellu lin in ten sit y (% of c on tr ol ) (B) (C) (D) (E) 0 25 50 75 100 125Fig. 7 Effects of the PLA concentration on the distribution of tight junction proteins in Caco-2 cell monolayers 120 min after treatment with PLA .
The tight junction proteins disappeared from the cell–cell contacts in a PLA dose-dependent manner (A). The intensities in all junction proteins were decreased with increasing PLA concentrations (B–E).
34
Fig. 8 Time-dependent alterations in the distribution of tight junction proteins after the application of 50 μg/mL PLA to Caco-2 cell monolayers.
The tight junction proteins disappeared from cell–cell contacts in a time-dependent manner following the treatment with PLA (A). The intensities in all junction proteins were decreased with increasing application times (B–E). Occludin and tricellulin were decreased more than the other proteins (B and E).
Each data point represents the mean ± S.E. (n=4). * p < 0.05 vs. Control.
0 15 30 45 60 120
Exposure time with 50 μg/mL PLA (min) ZO-1 Occludin Tricellulin Claudin-4 0 25 50 75 100 125
*
*
*
*
*
O c c lu di n int e nsi ty (% of c on tr ol ) 0 15 30 45 60 120 0 25 50 75 100 125*
*
*
*
0 15 30 45 60 120 ZO -1 i nt e nsi ty (% of c on tr ol ) 0 25 50 75 100 125*
*
*
*
0 15 30 45 60 120 Cl a u di n -4 i nt e nsi ty (% of c on tr ol ) 0 25 50 75 100 125*
*
*
*
0 15 30 45 60 120 T ricellu lin in ten sit y (% of c on tr ol )Time (min) Time (min)
(A)
(B) (C)
35
2-3-3 Caco-2 細胞 sheet における AJ 関連タンパク質の局在性に対する PLA の
影響
Figure 9A に 50 及び 100 μg/mL PLA 適用後の AJ 関連タンパク質である
E-cadherin 及び β-catenin の免疫蛍光染色画像を、Figs. 9B 及び C にそれぞれの
AJ 関連タンパク質の細胞間隙での蛍光強度を示す。PLA を適用していない (PLA
0 μg/mL) Caco-2 細胞において、E-cadherin 及び β-catenin は細胞間隙に局在して
いたが、PLA の適用により、細胞間隙への AJ 関連タンパク質の局在性は PLA の濃度の増加に伴い有意に低下した (p < 0.05) 。また、PLA による E-cadherin
とβ-catenin の細胞間隙部位への局在性の低下は同程度であった。
Fig. 9 Effects of the PLA concentration on the distribution of adherens junction proteins 120 min after treatment of Caco-2 cell monolayers with PLA .
Adherens junction proteins disappeared from the cell–cell contacts in a PLA dose-dependent manner (A). The intensities of the expression of all junction proteins were decreased with increasing PLA concentrations (B and C).
Each data point represents the mean ± S.E. (n=4). * p < 0.05 vs. Control.
E-cadherin β-catenin 50 100 Concentration of PLA (μg/mL) 0 0 25 50 75 100 125 * * 0 25 50 75 100 125 * * 0 50 100 0 50 100 Concentration (µg/mL) E -ca d h eri n int ens it y (% o f co n tr o l) β-ca ten in int ens it y (% o f co n tr o l) (A) (B) (C) Concentration (µg/mL)
36 0 25 50 75 100 125 0 25 50 75 100 125 ZO -1 (A r b itr ar y u n it) * * * 0 30 60 120 Occlu d in (A r b itr ar y u n it) Cl a u di n -4 (A r b itr ar y u n it) 0 25 50 75 100 125
(B)
ZO-1 Occludin Claudin-4 GAPDH 0 30 60 120(A)
Tricellulin Time (min) 0 25 50 75 100 125 T ricellu lin (A r b itr ar y u n it) Time (min) 0 30 60 120 Time (min)2-3-4 Caco-2 細胞 sheet における TJ 関連タンパク質の発現量に対する PLA の
影響
Figure 10A に 100 μg/mL PLA 適用後の occludin、 ZO-1、claudin-4 及び tricellulin のイムノブロット解析の結果を、Fig. 8B にそれぞれの TJ 関連タンパク質のシグ ナル強度を示す。ZO-1、claudin-4 及び tricellulin においては PLA を適用しても、 これら TJ 関連タンパク質の発現量はコントロールと比較して有意な変化は認め られなかった。一方、occludin については PLA 適用後 30 分以上で、その発現量 は有意に低下したが ( p < 0.05) 、その強度は 75%程度であった。
Fig. 10 Changes in the tight junction protein levels after treatment with 100 μg/mL PLA.
An immunoblot analysis of the tight junction proteins in Caco-2 cell monolayers was assessed after treatment of the cells with PLA for various times (A), and the density of the protein spots was also analyzed (B).
Each data point represents the mean ± S.E. (n=4). * p < 0.05 vs. Control.
37
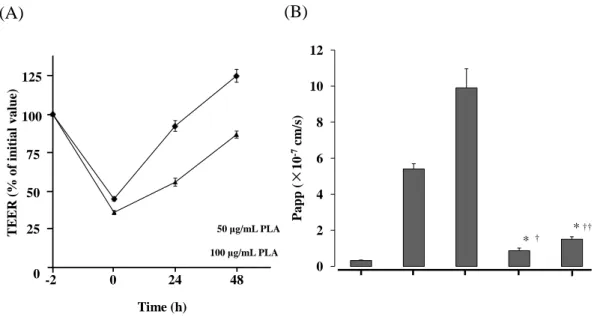
2-3-5 PLA による透過促進効果の可逆性の検討
PLA による TEER の低下や FD-4 の透過促進、TJ 関連タンパク質の局在性の
変化は PLA の除去により、可逆性を示すかどうかを、50 及び 100 μg/mL の PLA
を Caco-2 細胞シートに 2 時間適用後、HBSS で洗浄・除去することにより評価 した。Figs. 11A 及び B にそれぞれ PLA 除去後の TEER の変化及び FD-4 の Papp
を示す。PLA の除去により、TEER は徐々に回復した。特に 50 μg/mL PLA 適用
では除去 24 時間でほぼ完全に回復した。一方、100 μg/mL PLA 適用では 48 時間
で Control の 80%程度であった。PLA 除去後 24 時間での FD-4 の Papp は、50 及
び 100 μg/mL PLA 適用のいずれにおいても、適用時と比較して著しく低下する
ことが示された。
Fig. 11 Recovery of the TEER and FD-4 permeation.
After removal of PLA from the Caco-2 cell monolayers, the TEER values increased (A), and FD-4 permeation was significantly decreased
Each data point represents the mean ± S.E. (n=4). * p < 0.05 vs. Control.
0 2 4 6 8 10 12 P a pp ( × 10 -7 cm /s ) * †† * † T E E R ( % o f init ia l v a lue) 0 125 75 50 25 100 -2 0 24 48 Time (hr) 50 μg/mL PLA 100 μg/mL PLA Time (h) (A) (B)
38
Figure 12A 及び B-E にそれぞれ PLA 除去後の occludin、ZO-1、claudin-4 及び
tricellulin の局在性及び細胞間隙でのそれら TJ 関連タンパク質の蛍光強度の変化 を示す。いずれの TJ 関連タンパク質においても、PLA を除去することにより、 細胞間隙部位に戻ることが明らかとなった。特に細胞間隙への ZO-1 の再局在化 が速かった。Claudin-4 は 48 時間後、ほぼ 100%近くまで回復した。一方、occludin 及び tricellulin は 48 時間においても、完全には回復しなかった。 100 μg/mL PLA treatment Time after washout of PLA 0 25 50 75 100 125 0 25 50 75 100 125 0 25 50 75 100 125 0 25 50 75 100 125 O cc ludi n i nt ens it y (% o f co n tr o l) * * ZO -1 i nt ens it y (% o f co n tr o l) * C la udi n -4 nt ens it y (% o f co n tr o l) * * T ric ellu in int ens it y (% o f co n tr o l) * * * * 2 24 48 2 24 48 2 24 48 2 24 48 Control Control Control Control (B) (C) (D) (E)
Fig. 12 Recovery of redistribution of tight junction proteinsafter removal of PLA from Caco-2 cell monolayers.
Tight junction proteins were redistributed to the cell-cell contact (A), and the expression level of the tight junction proteins recovered (B-E).
Each data point represents the mean ± S.E. (n=4). * p < 0.05 vs. Control.
ZO-1 Occludin Tricellulin Claudin-4 100 μg/mL PLA treatment Control 2 24 48 Time (h) Time after washout of PLA (A) (B) (C) (D) (E) Time (h)
39
Figure 13 に PLA 除去後の occludin の発現量の変化を示す。PLA の 2 時間の適
用により減少した occludin の発現量は、PLA 除去後 24 時間で完全に PLA 適用 前の発現量にまで回復していた。
Fig. 13 Recovery of the occludin expression level after removal of PLA from Caco-2 cell monolayers.
An increased occludin level was detected by the immunoblot and densitometric analyses after the removal of PLA .
Each data point represents the mean ± S.E. (n=4). * p < 0.05 vs. Control.
Occludin GAPDH 100 μg/mL PLA treatment Time after washout of PLA O cc ludi n (A rbi tr a ry uni t) 0 25 50 75 100 125
*
Control 2 24 48 Time (h)40
第4節 小括
本章では、Transwell で Caco-2 細胞を培養し、コンフルエントに達した細胞 sheet
を用いて、細胞間隙経路の開口マーカーである TEER 及び FD-4 の透過性、さら には TJ 関連タンパク質の occludin 及び ZO-1 に加えて、これまでに検討されな かった claudin-4 及び tricellulin の局在性や発現量に対する PLA の影響を評価し た。
Caco-2 細胞 sheet においても、家兎摘出鼻粘膜やラット鼻粘膜と同様に、PLA
は FD-4 の透過量を増大し、そして TEER は低下した。さらに、これらの効果は PLA の適用濃度に依存した。さらに、種々濃度の PLA を適用後に得られた Papp
と Gt に良好な相関が得られたことに加え、FD-4 の蛍光画像から得られた透過経 路は細胞間隙であることが示された。これらの結果は、Caco-2 細胞においても PLA は細胞間隙経路を開口して、FD-4 の透過を促進することを示唆している。
さらに、TJ 関連タンパク質の局在性を評価した結果、PLA が家兎摘出鼻粘膜 の occludin 及び ZO-1 に対して影響を与えたように、Caco-2 細胞シートにおいて も同様な効果を示し、その効果は濃度依存的かつ時間依存的であった。さらに、 claudin-4 及び tricellulin に関しても、PLA は濃度依存的かつ時間依存的にそれら
の局在性に影響を与え、細胞間隙部から消失させた。Figures 7 及び 8 の結果、 並びに lag time の評価から、occludin 及び tricellulin は ZO-1 及び claudin-4 と比較 して TJ 部からの消失量が大きいこと、FD-4 の透過速度が増大する時間と、 occludin 及び tricellulin の消失速度が有意に早くなる時間が一致することから、
occludin 及び tricellulin の TJ 部からの経時変化が水溶性高分子薬物の透過促進に
おいて極めて重要であると思われる。
一方、PLA は AJ 関連タンパク質の E-cadherin 及びβ-catenin に対しても、その
41
TJ 関連タンパク質のうち、claudin-4、tricellulin 及び ZO-1 の発現量は、PLA
の適用でほとんど変化せず、occludin もわずかに減少した程度であった。これら の結果は、評価した TJ 関連タンパク質が、PLA の適用により細胞間隙部から消 失するのは、それらの分解によるものではなく、細胞内への内在化によること を強く推察させる。
PLA の効果の可逆性を評価した結果、PLA 除去後、TEER の上昇、FD-4 透過
量の減少が認められた。加えて、TJ 関連タンパク質の局在性や発現量が適用前 の状態に回復することが観察された。このことは、PLA の作用が可逆的である ことを示唆している。興味深いことに、PLA 除去後において TJ 関連タンパク質 の ZO-1 が最も速く細胞間隙部に再局在し、これは ZO-1 が TJ の再構築・再形成 において重要な役割を果たしている可能性が考えられる。
42
第 2 編 PLA による TJ 関連タンパク質の細胞間隙部位からの消失
機構
第1節 小緒言
前編の結果から、細胞間隙部位に分布している TJ 関連タンパク質の occludin, ZO-1, claudin-4 及び tricellulin は、PLA を適用することにより細胞間隙部位から
消失し、それは主に内在化によると推察された。しかし、PLA による TJ 関連タ ンパク質の消失がどのようなメカニズムによるのかはこれまでに報告されてい ない。従って、この消失機構を解明することは、PLA による透過促進メカニズ ムの極めて有用な情報を提供できる。 TJ 関連タンパク質の細胞間隙部位からの消失に関して、少なくとも 3 つの経 路の寄与を考えることができる。1 つ目として、TJ 関連タンパク質の mRNA レ ベルの減少を引き起こすことである。TJ 関連タンパク質の mRNA レベルの減少 に伴い、合成される TJ 関連タンパク質が減少し、TJ へリクルートされるタンパ ク質量が低下し、結果として、細胞間隙部での TJ の形成が不完全になる経路で ある。このような機構による TJ の破綻は短時間では引き起こされず、24~48 時 間かけて徐々に TJ が破綻する30)。しかし、PLA は TJ 関連タンパク質を減少さ せないことに加えて、細胞間隙からの迅速な TJ 関連タンパク質の消失を引き起 こすために、mRNA の減少を介した TJ の破綻を導かないことが容易に想像でき る。 2 つ目として、TJ 関連タンパク質が細胞間隙部位において分解される経路で ある。この TJ の分解経路として代表的なものに、タンパク質分解酵素の matrix metalloprotinases (MMPs) が挙げられる。特に、MMP-2 や MMP-9 が TJ の破綻に