This document is downloaded at: 2016-09-29T02:37:33Z
Title ホシガレイの全雌種苗生産技術の開発に関する研究
Author(s) 築山, 陽介
Citation Nagasaki University (長崎大学), 博士(水産学) (2013-02-20)
Issue Date 2013-02-20
URL http://hdl.handle.net/10069/31464
Right
ホシガレイの全雌種苗生産技術の開発に関する研究
Development of the Technique for All-Female
Seedling Production in the Spotted Halibut
Verasper variegatus
2012 年 12 月
長崎大学大学院生産科学研究科
築山 陽介
目 次
第1章 緒 言 1 第2章 雌性発生二倍体の作出 7 材料および方法 8 結 果 10 考 察 17 第3章 人為的な性制御 20 第1節 ホシガレイの性分化 20 材料および方法 20 結 果 21 考 察 24 第2節 高水温飼育による性転換 25 材料および方法 25 結 果 30 第3節 メチルテストステロン投与による性転換 43 材料および方法 44 結 果 48 考 察 58 第4節 総合考察 63 謝 辞 67 文 献 68第 1 章 緒 言 ホシガレイVerasper variegatus は,カレイ科マツカワ属に分類され,本州 中部以南,朝鮮半島西岸,渤海,黄海に分布している(山田1986)(Fig. 1)。 本種は長崎県内では橘湾及び有明海において底曳網,刺網により12 月~5 月に かけて主に漁獲され(森川ら2000),市場では 3,000~5,000 円/kg で取引され る高級魚である。 本種は岩手県,宮城県,福島県,大阪府および長崎県において種苗生産また は種苗放流が行われている(水産庁・独立行政法人水産総合研究センター・(社) 全国豊かな海づくり推進協会2012)。ただし,東北地方での種苗生産は 2011 年 の東日本大震災の影響を受け,現在は休止している。 長崎県では魚介類種苗量産技術開発研究事業(平成9-13 年度)および第 2 期 魚介類種苗量産技術開発研究事業(平成14-22 年度)により総合水産試験場で 本種の種苗生産技術開発に取り組み,10 万尾単位での種苗生産技術を確立させ, 長崎県漁業公社へ技術移管している。また,生産した種苗を用いて,魚類養殖 多様化推進事業により養殖試験を実施し,平成22 年度には養殖マニュアルを作 成している。 ホシガレイの養殖は,長崎県の陸上養殖施設においてヒラメParalichthys olivaceus に代わる魚種として普及を行っているが,ヒラメよりも成長が遅いた め(木本・佐藤2003),それを補う方法が必要になる。ホシガレイは,ヒラメ やマツカワVerasper moseri などの他の異体類と同様に雌が雄よりも成長が良 く,雌は満3 歳で体重 1 kg に達するが,雄は 0.5 kg 未満であること(佐久間 2001)から,雌種苗のみを飼育することが有利とされている(八木 2002)。 通常,本種の性比は1:1であるが,成熟するまで外見による雌雄の判別は困
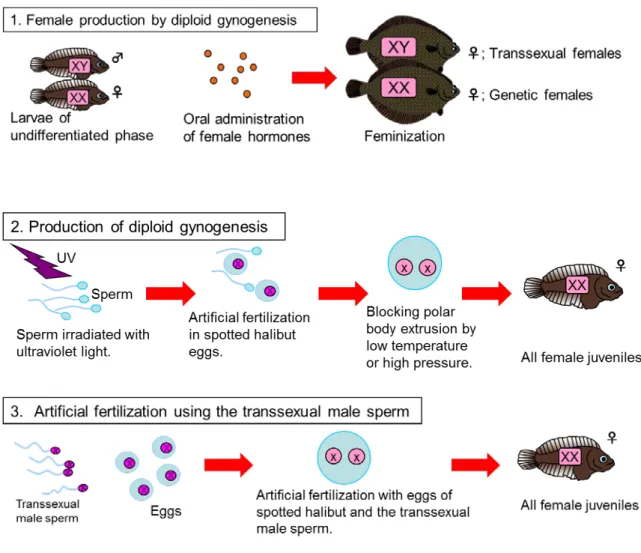
難で,成長の雌雄差も孵化後1年半までは顕著でないため(佐久間2001),養 殖現場において雌雄選別を行うことは難しい。ホシガレイ養殖では雌のみを飼 育することが効率的であるが,前述の理由から早期の雌雄選別が行えないため, 種苗の全雌化が効率の良い養殖を実現するための方策と考える。 全雌種苗を得るためには,1)雌性ホルモンの投与,2)染色体操作による雌 性発生,3)遺伝的雌を雄へ性転換させた偽雄の精子を用いた人工授精等の方法 がある(Fig. 2)。雌性ホルモン投与による雌化は,仔稚魚へのエストラジオー ル-17β(以下,E2)の浸漬または経口投与によって行われ,異体類ではヒラメ (田中1988;山本ら 1987)およびマツカワ(森ら 1995)で実施されている。 浸漬法では,ヒラメの性的未分化期(11~70 日齢)に E2 10 μg/l で処理するこ とにより,雌比率が対照区の35.7%に対して 48%となったが,全雌化には至っ ていない。一方,経口投与法による雌化はヒラメとマツカワで実施されており, ヒラメでは56~90 日齢,マツカワでは 73~103 日齢にかけて E2 1 μg/g・diet を毎日投与することにより,全雌種苗が得られている(田中1988;森ら 1995)。 染色体操作による雌性発生はホシガレイを対象として行われており,第二極 体放出阻止型の雌性発生の条件について検討したところ,八木ら(2004)は授 精3~5 分後に-1.5~-1℃で 60 分間処理することにより,Ji et al.(2010)は授 精5 分後に-1℃で 40~75 分間処理することにより雌化を誘導している。 遺伝的雌を雄へ性転換させた偽雄の精子を用いた人工授精による全雌化では, まず合成雄性ホルモンのメチルテストステロン(以下,MT)を性分化期の個体 に投与し,遺伝的雌を雄に性転換させなければならない。ホシガレイの遺伝的
性決定様式はヒラメと同様にXX/XY 型であり(Ji et al. 2010),性転換した遺
伝的雌の精子はX 染色体のみを持つことから,これを卵子と受精させれば全て
されている(山本1992)。しかし,Ma et al.(2010)はホシガレイの性決定様 式がZZ/ZW 型である可能性を論じており,この性決定様式が正しいとすると, 雌はZW 遺伝子を持つことから,全雌作出には WW 型の遺伝子を持つ偽雄を作 出しなければならない。全雌作出にいたる作業過程に多少の違いはあるものの, MT 処理による偽雄の作出は,全雌作出に有効な方法である。本研究では,ホシ ガレイの性決定様式はXX/XY 型の性決定様式として議論する。雌性ホルモンを 投与する方法は大量の種苗を少ない労力で性転換させることが可能であるが, 食品安全上,養殖魚には適さない。染色体操作による雌性発生魚の作出では, 孵化率が通常発生魚と比較してかなり低く,大量の種苗生産には解決しなけれ ばならない課題が多い。これに対して偽雄の精子を用いた種苗生産は,食品と して安全な大量の雌種苗が得られることから,実用性の高い技術として注目さ れている。 本研究では,このような背景のもと,ホシガレイの全雌種苗を生産するため に,まず雌性発生二倍体の作出に関する実験を行った。この実験では低温処理 による第二極体放出阻止型の雌性発生二倍体の作出条件である低温処理の開始 時間,処理水温および継続時間について検討した(第2 章)。続いて,魚類の遺 伝的な性を人為的に操作するためには,性的未分化期に性操作を継続して行う 必要があることから(Nakamura and Takahashi 1973),性分化の過程を明ら
かにする試験を行った(第3 章第 1 節)。また,偽雄を作出するために必要な, 遺伝的雌を雄化する技術の開発に向け,ヒラメおよびマツカワが水温等の環境 要因により性転換すること(山本1995;Goto et al. 1999)から,ホシガレイに ついても性転換する可能性が高いと考えられたため,ホシガレイ仔稚魚の飼育 水を加温または冷却し,性分化に及ぼす影響について試験を行った(第3 章第 2 節)。最後に,MT の経口投与試験を行い,ホルモンの添加濃度および投与期間
による性分化および生残に及ぼす影響について調べた(第3 章第 3 節)。これら の成果をもとに,ホシガレイの全雌化技術の最適化について考えるとともに, 長崎県におけるホシガレイの種苗生産戦略についても議論する。
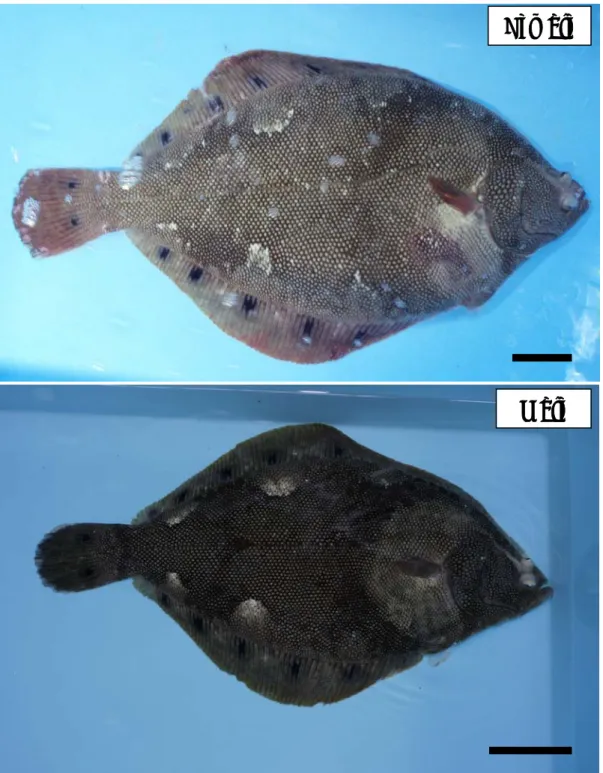
Fig. 1. Spotted halibut Verasper variegatus. Bars = 50 mm.
Male
Female
Fig. 2. Possible techniques for all-female seedling production in the spotted halibut Verasper variegatus.
第 2 章 雌性発生二倍体の作出 ホシガレイの全雌種苗生産に有効な方法には,緒言で述べたように1) 雌性ホ ルモンの投与による雌の作出,2) 染色体操作による雌性発生,3)遺伝的雌を雄 へ性転換させた偽雄の精子を用いた人工授精による雌の作出の 3 つの方法があ る。しかし,食品としての安全面から実用可能な方法は,後者の 2 つの方法, すなわち染色体操作による雌性発生および遺伝的雌を雄へ性転換させた偽雄の 精子を用いた人工授精による雌魚の生産である。そこで本研究では,まず染色 体操作によるホシガレイの雌性発生二倍体の作出を試みた。作出した雌性発生 二倍体をそのまま種苗生産に用いることができれば,大量の全雌を比較的簡単 に生産することができる。しかし,雌性発生処理により孵化率が通常魚よりも 低下することから,魚種によってはこの方法を用いて全雌生産の産業化に必要 な尾数を確保するのは極めて困難である。しかし,雌性発生二倍体は,遺伝的 雌のホルモンによる偽雄化にも活用できる。偽雄を作出するホルモン処理には 性的に未分化の孵化後間もない個体を用いることが多いが,天然の個体を用い た場合,遺伝的雄と雌が1:1 で混在することから,作出した雄の半分が遺伝的雄 となる。そのため,本来の性がいずれであったかを検討し,偽雄のみを選別し なければならない。偽雄と遺伝的雄を判別するにはホシガレイが成熟する 2 歳 まで飼育し,採取した精子を用いて後代検定を行う方法があるが,そのために は飼育の労力や場所および親魚の個体識別も必要であり,非効率的である。一 方,雄化処理を施す魚が遺伝的雌のみであれば,雄化処理後に出現した雄は間 違いなく偽雄であり,後代検定の必要がない。このように雌性発生二倍体の作 出は偽雄作出に活用可能である。この場合,雌性発生二倍体を直接全雌生産用 の種苗として利用するのとは異なり,人工授精に用いる精液は数個体の偽雄か ら必要量を十分に得られることから,雌性発生処理によって生残率が低下し,
わずかの個体しか生産できなかったとしても十分利用可能である。そこで本研 究では,ホシガレイの雌性発生二倍体を作出条件を確認するための試験を行っ た。 一般に,雌性発生二倍体の作出において,対象種の卵発生を誘起でき,かつ 雑種を生じても非生存性となる異種精子を用いれば,作出個体が雌性発生二倍 体であるか否かの判断が容易になること(山川ら1987)から,これまでホシガ
レイの雌性発生にはマダイPagrus major(八木ら 2004),マコガレイ Limanda yokohamae(八木ら 2005)およびスズキ Lateolabrax japonicus(Ji et al. 2010) の精子が用いられている。本研究では,これまでの事例をもとにマダイ精子を 用いるとともに,ホシガレイ精子を用いて雌性発生二倍体の作出を試みた。 材料および方法 供試魚 長崎県橘湾で漁獲されたホシガレイ天然親魚を長崎県総合水産試験場へ搬入 し実験に用いた。配偶子の確保にあたり,山田ら(2006)に従い,背筋部へヒ ト絨毛性生殖腺刺激ホルモン(HCG)を 100 IU/kg・BW となるように投与した。 その後 24 時間以上経過した雌親魚より採卵し,雄からは採精を行った。また, 採精用マダイ親魚には,長崎市内の養殖業者から入手した養殖魚を長崎県総合 水産試験場において養成した 4 歳魚を用いた。雌性発生用の精子の確保にあた り,マダイ親魚を12 月に海面生簀から陸上水槽に収容し,飼育水温を徐々に昇 温して 20℃で成熟を促し,採精した。紫外線照射試験用の配偶子は,5 月に海 面生簀内で成熟した親魚から採卵および採精した。
精子への紫外線照射による遺伝的不活化 精子を不活化する適切な紫外線照射量を確認するため,マダイおよびホシガ レイの精子を用いて実験を行った。親魚から採取したマダイおよびホシガレイ の精液をクロダイ人工精漿(Morisawa 1985)で 100 倍に希釈した。希釈精液 5 ml を直径 14 cm のプラスチックシャーレ上に薄く延ばし,ロータリーシェー カーで振とうしながら,殺菌灯 1 本(GL-15,東芝)により紫外線を照射した。 紫外線照射量は,シャーレ上の5 箇所の紫外線強度を紫外線強度計(UVC-254, カスタム)を用いて測定し,その平均値に照射時間を乗じて算出した。照射紫 外線量は,マダイ精子では1,2,5,10,20,30,40,50,60 および 70 mJ/cm2, ホシガレイ精子では0.5,1,2,5,10,20,30,40 および 50 mJ/cm2とした。 精子の不活性化の程度を調べる実験では,紫外線照射した精子を同種の成熟 卵と人工授精させ,その後の発生の変化を観察した。成熟卵を試験区毎にボウ ルへ分け,紫外線を照射した精子を媒精した。対照区には,紫外線を照射して いない精子を媒精した。浮上卵 100 粒以上を 500 ml ビーカーに収容し,イン キュベーターで卵管理した。孵化までの期間中は,1 日 1 回,沈卵の除去および 海水交換を行い,実体顕微鏡を用いて卵または孵化仔魚を観察した。受精率, 胚体形成率および孵化率は,次の式で算出した。 受 精 率 (%) = (受精卵数/ビーカー収容卵数) ×100 胚体形成率 (%) = (胚体形成卵数/ビーカー収容卵数) ×100 孵 化 率 (%) = (孵化仔魚数/ビーカー収容卵数) ×100 雌性発生処理条件の検討 低温処理によりホシガレイの第二極体放出阻止型雌性発生二倍体の作出を試 みた。卵発生の誘起には,30 mJ/cm2 の紫外線照射により不活化したマダイ精
子を用いた。低温処理の各条件を検討するため,1) 授精から低温処理開始まで の時間,2) 低温処理の継続時間,および 3) 処理水温の3 項目について実験を 行った。 低温処理開始時間の検討は,開始時間を授精後 1.5,3,5 および 7 分とし, この4 区について-1℃の低温処理を 60 分間継続することにより行った。開始時 間の影響は,受精率および孵化率によって評価した。低温処理の継続時間の検 討は,処理時間を30,60 および 90 分間とし,この 3 区について,受精 3 分後 から0.5℃の低温処理を施して,受精率および孵化率を算出した。低温処理水温 の検討は,処理水温を-1,1,3 および 5℃とし,この 4 区について受精 3 分後 に60 分間の低温処理により行った。この実験では,不活化マダイ精子に加えて, 不活化していないホシガレイとマダイの精子を用いた低温処理を施さない試験 区も設け(自然水温,14℃),授精率および孵化率を求めた。低温処理には,チ ラー(CL-80R,タイテック)または海水氷により冷却した海水を用いた。 上記の全ての試験には,異なる 3 個体の親魚より得られた卵を用いて同一の 試験を繰り返し行った。 統計処理 授精率および孵化率の有意差は一元配置の ANOVA の後,Tukey-Kramer 法 による多重比較により求めた。 結 果 精子の遺伝的不活化 紫外線照射により不活化した精子を用いた受精試験の結果をFig. 3 に示す。
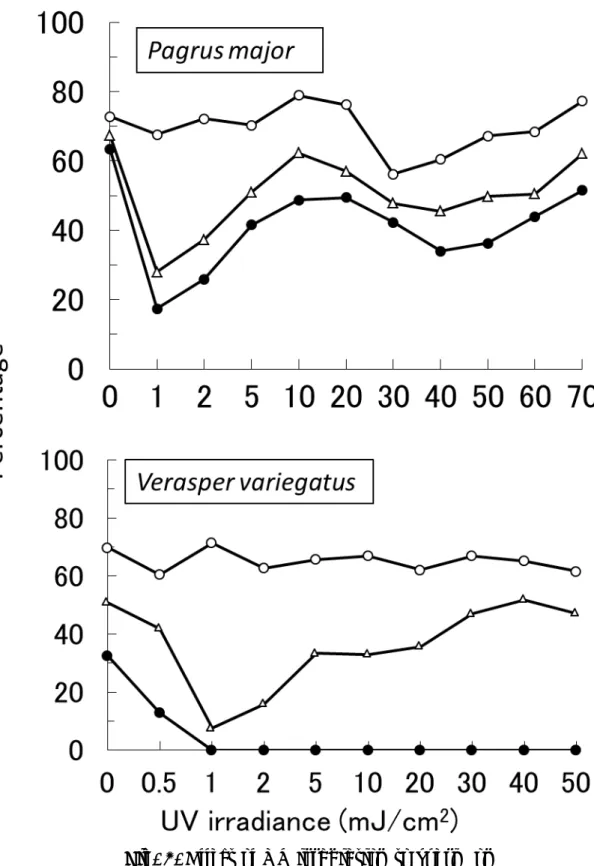
マダイの精子では,紫外線を照射しない対照区の授精率は 72.8%,胚体形成率 および孵化率は67.3%および 63.4%であった。1 mJ/cm2の紫外線を照射した精 子を用いると,胚体形成率および孵化率が大幅に低下し,28.0%および 17.4% と最低値となった。紫外線照射量の増加に伴い,10 mJ/cm2で胚体形成率およ び孵化率が62.3%および 48.7%に回復した。紫外線照射区の孵化仔魚は全て体 が湾曲した奇形魚であった。 ホシガレイ精子では,紫外線を照射しない対照区の授精率は 69.8%と比較的 高く,胚体形成率および孵化率は 50.8%および 32.5%であった。紫外線 0.5 mJ/cm2の照射では,胚体形成率および孵化率の低下が確認され,1 mJ/cm2の 照射によりそれらは大幅に低下した。1 mJ/cm2での胚体形成率および孵化率は 最低値の 7.6%および 0%を示した。2 mJ/cm2 以上の照射では胚体形成率が 15.9%となり,照射量の増加とともに胚体形成率も増加し,30 mJ/cm2以上で横 ばいとなり,40 mJ/cm2において51.8%と最高値を示した。また,1 mJ/cm2以 上の照射区では生存した孵化仔魚はみられなかった。1 mJ/cm2以上の紫外線を 照射した区の胚体は,対照区のそれと比較して体が太く短い,半数体の特徴を 示した。
Fig. 3. Effect of UV irradiation to sperm on fertilization (○), embryo forming (△) and hatching (●).
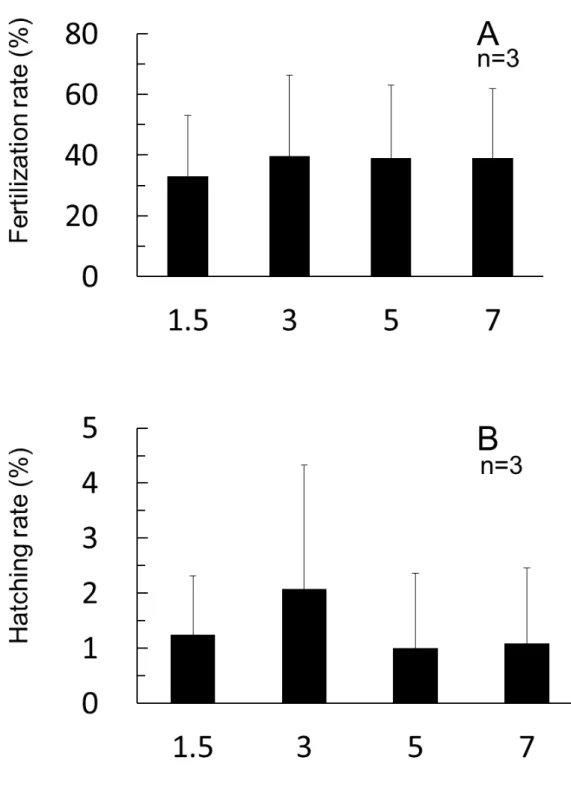
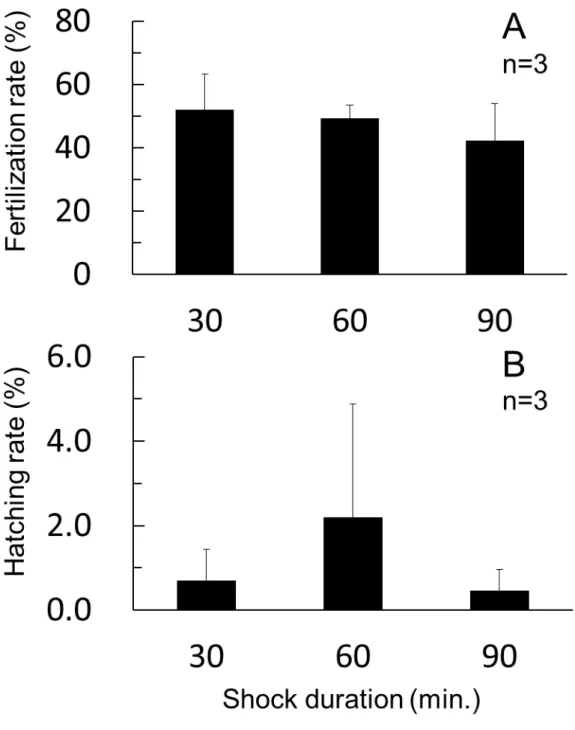
雌性発生処理条件 低温処理開始時間を検討した試験の結果をFig. 4 に示す。受精率は 1.5~7 分 後の各区で有意差がみられなかったものの,1.5 分後区が他の区と比較して,や や低い値を示した。孵化率は受精 3 分後の値が他の区と比較して高かったが, 有意差はなかった。 低温処理の継続時間を検討した試験の結果をFig. 5 に示す。授精率は処理時 間が長くなるに従って低下する傾向がみられたが,各区間に有意差はなかった。 また,孵化率は60 分間処理区において,30 および 90 分間処理よりも高い値を 示したが,有意差はなかった。 低温処理水温の影響を調べた試験の結果をFig. 6 に示す。ホシガレイ精子, マダイ精子および紫外線照射により不活化したマダイ精子の受精率は 57.4~ 60.1%となり,ほぼ同じ値を示し,有意差はみられなかった。低温処理水温の 影響を調べたところ,低温処理を施さなかった卵の授精率と比べ,低温処理を 施した卵の授精率は 39.4~49.4%となり,低値を示したが,有意差はみられな かった。また,この受精率は処理水温が高くなるに従い,増加する傾向がみら れた。孵化率は,ホシガレイ精子を用い低温処理を施さない卵では43.6±19.3% であったが,-1~3℃の低温処理区では 7.1~7.6%と低い値を示し (P<0.05),5℃ 処理区では10.7%となり,有意差は認められなかった。
Fig. 4. Effects of starting time after fertilization of cold shock treatment on fertilization (A) and hatching (B).
Fig. 5. Effect of cold shock duration on fertilization (A) and hatching (B). Bars indicate SD.
Fig. 6. Effect of cold water temperature on fertilization (A) and hatching (B).
Bars indicate SD. Different letters show significantly different mean values (P<0.05).
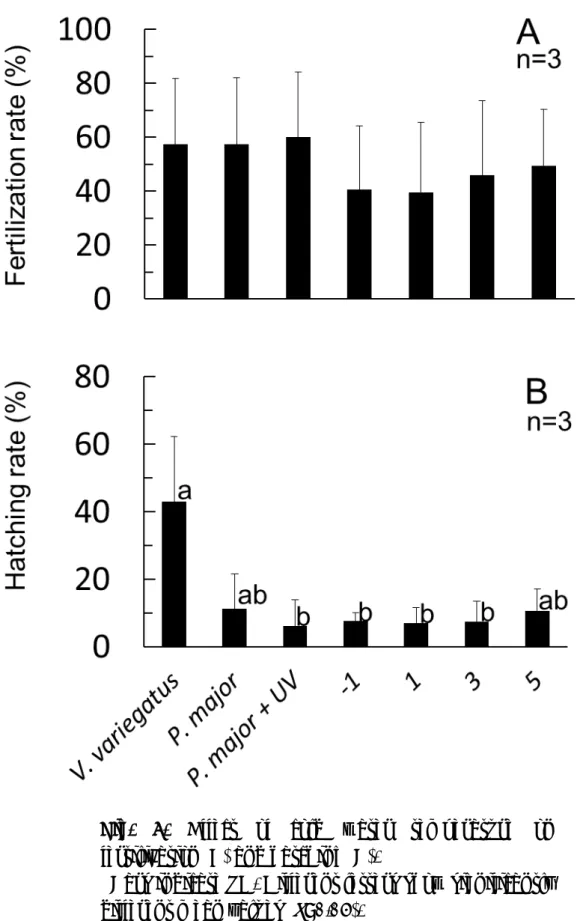
考 察 紫外線を照射したマダイおよびホシガレイの精子を人工受精に用いたホシガ レイ卵の胚体形成率は紫外線照射により減少し,1 mJ/cm2において最低値を示 した。さらに紫外線照射量の増加に伴い胚体形成率は徐々に増加し,マダイ精 子では10 mJ/cm2,ホシガレイ精子では30 mJ/cm2以上の照射量で対照区とほ ぼ同等まで胚体形成率が回復した。孵化率は胚体形成率と同様に1 mJ/cm2で最 低値を示し,マダイ精子では紫外線量の増加に伴い増加した。ホシガレイ精子 では1 mJ/cm2以上の照射区では孵化仔魚はみられなかった。これは,紫外線照 射によるHertwig 効果であると推察される。Hertwig 効果とは, 紫外線等の照 射により精子の染色体が部分的に切断されることによって胚発生に対して生じ た有害作用が,ある線量を超すと精子の染色体が消失または動原体が損失し, これが卵割に参加しなくなることにより,逆に減少していくことである(小野 里1982)。このことから,精子への紫外線照射はマダイでは 10 mJ/cm2以上, ホシガレイでは30 mJ/cm2以上行うことで,精子の遺伝子が完全に不活化され ると推察された。既往の報告による孵化率が低下する紫外線量は,ヒラメ精子 では1.1~2.3 mJ/cm2(田畑ら 1986),マツカワ精子では 2~5 mJ/cm2(森ら 2004)であり,本研究の結果とほぼ同じであった。マダイ精子では 3~10 mJ/cm2 (山川ら1987)と本研究の結果より照射線量が高いが,これは使用しているシ ャーレの大きさ(直径9 cm)と処理液量(3 ml)による液量の深さおよび精子 の希釈濃度(50 倍)による差と推察される。 雌性発生においては,紫外線照射および倍化操作の成否により雌性発生二倍体 の他に,同種の精子を用いた場合には 3 倍体,通常の 2 倍体および半数体が作 出され,異種の精子では異質 3 倍体,交雑魚および半数体が生じる。ホシガレ
イ卵およびマダイ精子を用いて雌性発生魚の作出を行った場合,異質 3 倍体お よび交雑魚が僅かに孵化するが,こちらは開口までに全てへい死し,半数体は 孵化しないことから,生残魚は雌性発生二倍体と推察される(八木ら 2004)。 したがって,ホシガレイの雌性発生二倍体作出には異種精子であるマダイ精子 が有用であると判断した。しかし,マダイはホシガレイの産卵期である冬季に は,成熟促進用の加温水槽が必要なことから,将来は八木ら(2005)が検討し ているマコガレイのようなハンドリングが容易でホシガレイと成熟期が同時期 である異種魚種の精子を用いることが望ましいと思われる。 低温処理による雌性発生魚の作出条件として,低温処理の開始時間,処理の継 続時間および処理水温の 3 項目について実験を行った。低温処理の開始時間を 検討したところ,受精1.5~7 分後の全ての区において孵化仔魚が得られた。雌 親魚により傾向は異なるものの,授精 3 分後の処理開始が最も高い孵化率を示 した。マツカワの雌性発生二倍体作出では,受精 3~11 分後の処理開始により 孵化仔魚が得られ,9 分後の孵化率が有意に高くなっている(森ら 2004)。この ように,マツカワではホシガレイより処理開始時間を遅くしても高い孵化率が 得られている。これは森ら(2004)によるマツカワの卵管理水温が 8~10℃と 本試験の卵管理水温12~14℃より 2~6℃低く,卵発生速度が遅いことが原因の 一つと考えられた。 低温処理の継続時間の試験では,30~90 分間の全ての試 験区で孵化仔魚が得られた。卵のロットにより傾向は異なるが,60 分間の低温 処理の孵化率が高く,最も適していると考えられた。他の異体類の低温処理継 続時間は,マツカワでは60~90 分間(森ら 2004),ヒラメでは 45 分間(田畑 ら 1986),マコガレイでは 60 分間(柿本ら 1994),ヌマガレイ Platichthys stellatus では 60 分間,スナガレイ Limanda punctatissima では 60 分間であ
処理水温の試験では,-1~5℃の全ての区で孵化仔魚が得られたが,ほとんどが 奇形魚であった。この比較試験では最適な処理水温を明らかにすることはでき なかったが,これまでに行った雌性発生二倍体の作出では,-1.2~1.4℃の低温 処理により雌性発生二倍体魚が得られていることから,この範囲内に低温処理 に適した水温があると推察された。これらの結果から,受精3 分後に-1.2~1.4℃ の低温処理を60 分間継続することで雌性発生二倍体が作出できると考えられた。 これは八木ら(2004)が授精 5 分後に,-1℃の低温処理を 60 分間継続すること で雌性発生二倍体が得られると報告した事例とほぼ一致していることから,ホ シガレイに適した方法であると考えられる。
第 3 章 人為的な性制御
第 1 節 ホシガレイの性分化
魚類の遺伝的な性を人為的に性ホルモン等の処理により制御するには,性分化 期間の初期から終了までの全期間を通して性制御の処理を継続する必要がある (Nakamura and Takahashi 1973)とされることから,性制御を行うにあたり
対象種の性分化の時期および期間を確認しておくことが必要である(森ら1995) そこで,ホシガレイにおいても,性分化に関する知見を得ることを試みた。 材料および方法 供試魚 長崎県橘湾において漁獲されたホシガレイ天然親魚から第 2 章と同様に, HCG を用いて採卵し,人工授精により得られた仔稚魚を供試魚に用いた。実験 に用いた仔魚の発育ステージの分類・表記は,有瀧ら(2001)の定義に従った。 全長6.1±0.1 mm(4 日齢,ステージ C)の仔魚 1,000 尾および全長 18.7±1.5 mm(46 日齢,ステージ H)の仔魚 1,000 尾をそれぞれ 200 l および 500 l 水槽 に収容し,飼育した。仔魚への餌料系列には,宮木ら(2005)の方法に従い, シオミズツボワムシL 型Brachionus plicatilis を 5 日齢から,配合飼料(ラブ ラァバ,林兼産業)を30 日齢から,アルテミアArtemia sp. のノープリウス幼 生を37 日齢から給餌した。シオミズツボワムシとアルテミアは,栄養強化剤(バ イオクロミスリキッド,クロレラ工業)により給餌前に栄養強化した。飼育水 には紫外線殺菌海水を用い,自然水温で飼育した。シオミズツボワムシの給餌 期間中はナンノクロロプシスNannochloropsis oculata を 50 cells/ml になるよ う飼育水へ添加した。
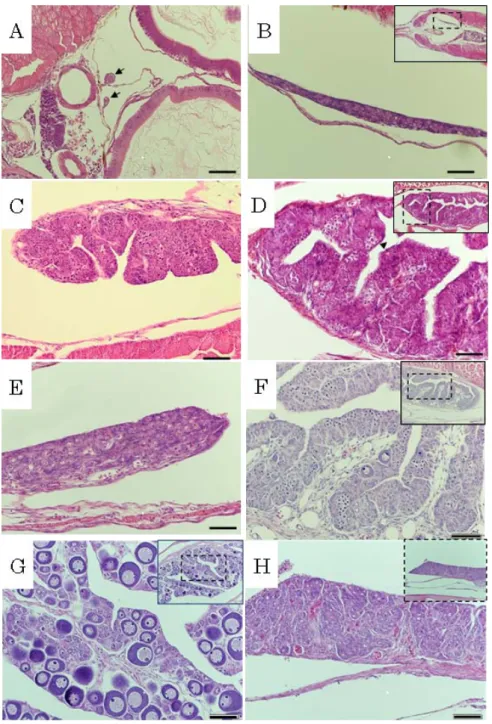
サンプリング 200l 水槽からは 15,25 および 30~100 日齢まで 10 日毎に各 10 個体をサン プリングし,500l 水槽からは 80~180 日齢まで 10 日毎に各 10 個体をサンプリ ングした。採取した仔稚魚は,万能投影機を用いて全長を測定した。 生殖腺の組織学的観察 サンプリングした全ての供試魚は,ブアン氏液で固定後,70%エタノールで 保存した。固定した生殖腺を定法により5 μm のパラフィン切片にし,ヘマトキ シリン・エオシンの二重染色を施し,光学顕微鏡下で観察した。仔魚の発育ス テージを有瀧ら(2001)に従って判定した。 結 果 生殖腺の組織像をFig. 7 に示す。30 日齢の仔魚は全長 12.7±0.9 mm(n=10), ステージ E であった。組織観察により,副腎輸管と消化管の間に一対の生殖隆 起が認められた。生殖隆起内部には,少数の始原生殖細胞が認められる未分化 の状態であった(Fig. 7-A)。70 日齢の仔魚は全長 20.5±1.0 mm(n=10),ス テージH でほぼ変態を完了しており,着底していた。これらの個体の生殖腺は 大きくなり,厚みも50 μm ほどを示し,副腎輸管から肛門方向へ伸長した。ま た,生殖細胞数も増加したが,生殖腺において性分化を示す形態的特徴は確認 されなかった(Fig. 7-B)。90 日齢の稚魚は全長 28.4±4.9 mm(n=10),ステ ージI で変態が完了していた。生殖腺の組織観察において,一部の個体で生殖腺 に卵巣腔の形成が初めて確認され,雌への性分化が確認された。減数分裂中の 生殖細胞が多く観察され,生殖腺内の細胞数がさらに増加していた(Fig. 7-C)。
100 日齢の稚魚は全長 35.4±5.2 mm(n=10)であった。生殖腺の外部形態によ り 2 つの型に分類できるようになった。一方は腹腔最後部から尾部の筋肉に沿 って生殖腺が後方へ伸長し,この生殖腺組織には卵巣腔の形成が確認され,雌 への分化が確認された(Fig. 7-D)。他方の生殖腺では後方への伸長が認められ ず,腹腔後部に付着しているだけであった。組織観察では結合組織が多く認め られ,生殖細胞は卵巣へ分化した個体と比較すると少なく,将来は雄へ分化す ると考えられた(Fig. 7-E)。120 日齢の稚魚は全長 59.4±14.1 mm(n=10)で あった。卵巣では,卵巣腔がさらに伸長し,卵巣薄板が形成され,減数分裂前 期の卵母細胞が多数確認された。一部に染色仁期の卵母細胞が観察され,雌へ の分化が進んでいた(Fig. 2-F)。160 日齢の稚魚は全長 93.2±12.9 mm(n=10) に成長し,卵巣内には周辺仁期の卵母細胞が多数観察された(Fig. 7-G)。一方, 精巣に分化すると考えられる生殖腺では精原細胞がみられ,包嚢を形成した (Fig. 7-H)。
Fig. 7. Cross sections of gonad in spotted halibut Verasper variegatus. A, 30 days after hatching (DAH). Arrows show the sexually undifferentiated gonad; B, Sexually indifferent gonad at 70 DAH; C, Ovary at 90 DAH. Arrow head shows the ovarian cavity; D, Ovary at 100 DAH. Arrow head shows the ovarian cavity; E, Gonad at 100 DAH; F, Ovary including perinucleolus oocytes at 120 DAH; G, Ovary including perinucleolus oocytes at 160 DAH; H, Testis including spermatogonia at 160 DAH; Bars=100 μm.
考 察 本研究によって,ホシガレイの性分化過程が明らかになった。自然水温で飼 育したホシガレイは,70 日齢(全長 21 mm,ステージ H)までは性的未分化の 状態であったが,90 日齢(全長 28 mm,ステージ I)の稚魚では,一部の個体 で生殖腺に卵巣腔の形成が初めて確認され,雌への性分化が確認された。100 日齢(全長35 mm,ステージ I)では生殖腺の外部形態が 2 つの型に分類され た。すなわち,一方の生殖腺は形態的に未分化のままであったが,他方のそれ では卵巣腔の形成が観察されており,卵巣への分化が確認された。このことか ら,ホシガレイの雌への性分化は70~100 日齢(全長 21~35 mm,ステージ H ~I)の間に行われると推察された。また,多くの魚種と同様に,雌における卵 母細胞の出現が雄における精母細胞の出現よりも早期に観察された。ヒラメで は,75~88 日齢(全長 30 mm 前後)の雌において卵巣腔の形成がみられ,体 の後方へ生殖腺が伸長し,88 日齢(全長 55 mm)の稚魚で減数分裂初期の卵母 細胞が初めて確認されている(田中1987)。また,マツカワでは,73 日齢(全 長30 mm)の個体の生殖腺は未分化で雌雄判別できないが,99 日齢(全長 55 mm)では生殖腺に2つの型がみられ,一方に卵巣腔の形成が確認され,雌への 分化が確認されている(森ら1995)。ホシガレイは,ヒラメと比較して雌への 性分化が若干遅く,マツカワとほぼ同じであることが分かった。これは,ホシ ガレイの変態完了が70~80 日齢前後(全長 25 mm 前後)とヒラメの変態完了 (36~50 日齢,全長 9.0~19.4 mm,原田ら 1966)よりも遅れることから,魚種 による発生および発達速度の違いと関連していると考えられる。
第 2 節 高水温飼育による性転換 本種の近縁種であるマツカワでは,性分化期の仔魚を高水温で飼育すると性 比が雄に偏ることが報告されており(Goto et al. 1999;森 2006),ヒラメでは 性分化期に高水温飼育することにより雄化される(山本 1992;岩田ら 1998)。 このように仔魚の高水温処理が遺伝的な雌個体の性を雄に誘導することは,ヒ ラメ科やカレイ科の魚類においては一般的な現象と考えられ,ホシガレイも飼 育水温の調節によって雄化を誘導できる可能性が考えられる。本種においても マツカワおよびヒラメと同様に飼育水温の調節により人為的に「偽雄」を作出 することにより,全雌種苗の生産が可能となる。しかし,ホシガレイ仔稚魚は 高水温により生残率が低下することが知られており(村田ら1998;佐久間 2001), 雄化を促す高水温処理は,生残に悪影響を及ぼすことが予想される。したがっ て,本種において高水温処理によって産業的に利用できる雄化を行うことが可 能か否かを明らかにするためには,高水温の性分化に及ぼす影響に加えて,生 残に及ぼす影響についても検討する必要がある。 そこで,本研究では異なる水温でホシガレイの仔稚魚を飼育し,生残および 性分化に及ぼす水温の影響を調べた。 材料および方法 供試魚 平成21~23 年 1 月に長崎県橘湾で漁獲された天然魚から,人工授精により得 られた受精卵をアルテミア孵化槽で卵管理し,孵化仔魚を飼育水槽へ収容し, 後述する条件で飼育した。仔魚への餌料系列には,シオミズツボワムシL 型を 5 日齢から,配合飼料(ラブラァバ,林兼産業)を30 日齢から,アルテミアのノ
ープリウス幼生を 37 日齢から給餌した。シオミズツボワムシとアルテミアは, 栄養強化剤(バイオクロミスリキッド,クロレラ工業)により栄養強化した。 飼育水には紫外線殺菌海水を用い,シオミズツボワムシの給餌期間中はナンノ クロロプシスを50 cells/ml になるよう添加した。 試験方法 本実験における飼育条件をTable 1 に,水温処理方法を Fig. 8 に示す。また, 実験に用いた仔魚の発育ステージの分類・表記は,有瀧ら(2001)に従った。 試験1 では全長 6.1±0.1 mm(4 日齢,ステージ C)の仔魚 1,000 尾を 200 l 水 槽に収容し,10℃,20℃,23℃,26℃,28℃および自然水温(12.9-26.7℃)の 6 つの異なる条件下で飼育した。試験 2 では全長 15.0±1.3 mm(40 日齢,ステ ージG~H)の仔魚 1,000 尾を 1 kl 水槽に収容し,26℃,28℃および自然水温 (13.6-27.9℃)で飼育した。試験 3 では全長 18.7±1.5 mm(46 日齢,ステー ジH)の仔魚 1,000 尾を 500 l 水槽に収容し,23℃および自然水温(14.2-29.0℃) で飼育した。試験4 では全長 34.6±3.9 mm(95 日齢,ステージ I)の稚魚 300 尾を300 l 水槽に収容し,28℃および自然水温(14.8-24.9℃)で飼育した。28℃ での飼育は130 日齢(28℃-a 区)および 146 日齢(28℃-b 区)までとし,以降 は自然水温とした。各試験において,自然水温で飼育した水槽を対照区とした。 試験区における水温設定は,設定水温になるまで 1℃/日の割合で飼育水を昇温 または冷却した。また,自然水温が設定水温を超えた場合には,自然水温で飼 育を継続した。各水槽の水温はデジタル温度計(SK-250WPⅡ-K,佐藤計量器 製作所)を用い,毎日午前8~9 時頃に測定した。
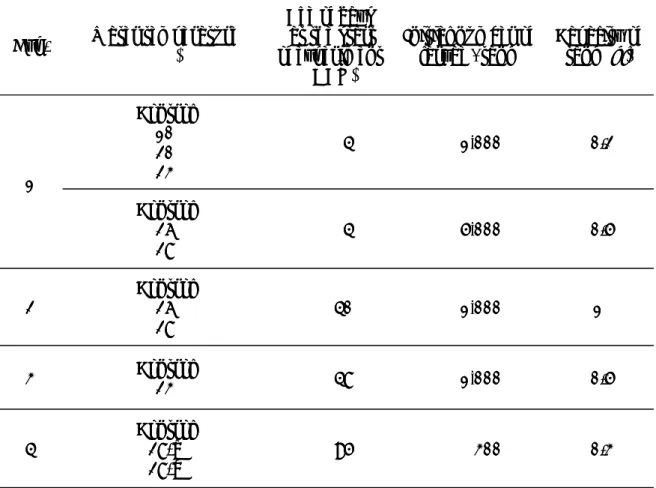
Table 1. Outline of the rearing experiments
Exp. Water temperature(℃)
Age of days at the start of experiment
(DAH)
Initial number of
larvae / tank Capacity of tank (kl)
1 Control 10 20 23 4 1,000 0.2 Control 26 28 4 5,000 0.5 2 Control26 28 40 1,000 1 3 Control23 48 1,000 0.5 4 Control28-a 28-b 95 300 0.3
Table 1. Outline of the rearing experiments
Exp. Water temperature(℃)
Age of days at the start of experiment
(DAH)
Initial number of
larvae / tank Capacity of tank (kl)
1 Control 10 20 23 4 1,000 0.2 Control 26 28 4 5,000 0.5 2 Control26 28 40 1,000 1 3 Control23 48 1,000 0.5 4 Control28-a 28-b 95 300 0.3
20 DAH 17 DAH
200
A
B
C
D
Cont. 28℃ 26℃ 10℃ 23℃0
20℃ 26℃ 28℃ Cont. 23℃ Cont. Cont. 28℃-a 28℃-bDAH
W
ater
te
m
per
a
tu
re
(℃
)
4 DAH 50 DAH 65 DAH 62 DAH 15 DAH 22 DAH 17 DAH 59 DAH 49 DAH 95 DAH 110 DAH 131 DAH 146 DAH 138 DAH 150 DAH30
0
30
0
30
0
30
0
122 DAH 142 DAH 192 DAH 140 DAH 141 DAH 20 DAH 17 DAH200
A
B
C
D
Cont. 28℃ 26℃ 10℃ 23℃0
20℃ 26℃ 28℃ Cont. 23℃ Cont. Cont. 28℃-a 28℃-bDAH
W
ater
te
m
per
a
tu
re
(℃
)
4 DAH 50 DAH 65 DAH 62 DAH 15 DAH 22 DAH 17 DAH 59 DAH 49 DAH 95 DAH 110 DAH 131 DAH 146 DAH 138 DAH 150 DAH30
0
30
0
30
0
30
0
30
0
30
0
30
0
30
0
30
0
30
0
30
0
30
0
30
0
30
0
30
0
30
0
122 DAH 142 DAH 192 DAH 140 DAH 141 DAHFig.8. Experimental design for study on temperature effects in spotted halibut Verasper variegatus.
A, Experiment 1; B, Experiment 2; C, Experiment 3; D, Experiment 4. Control fish was reared on natural water temperature.
サンプリング 試験1 では 15,25 および 30~100 日齢まで 10 日毎に各試験区から 10 個体 ずつサンプリングし,試験終了時(200 日齢)に対照区,10℃区および 20℃区 からそれぞれ 120 個体,23℃区から 25 個体をサンプリングした。なお,26℃ および28℃区ではへい死魚が多かったため,7 日齢において各 10 個体をサンプ リングした。また,26℃区では 40 日齢に 25 個体を採集し試験を終了したが, 28℃区はそれまでに全てがへい死した。試験 2 では 40~100 日齢の期間,10 日 毎に各試験区から10 個体ずつサンプリングし,試験終了時(200 日齢)に対照 区から 180 個体,26℃区から 161 個体をそれぞれサンプリングした。ただし, 28℃区では飼育個体が大量へい死したため,140 日齢で実験を終了した。試験 3 では80~180 日齢の期間,10 日毎に各試験区から 10 個体ずつサンプリングし, 試験終了時(200 日齢)に各区から 120 尾ずつをサンプリングした。試験 4 で は 134 日齢および 174 日齢に各試験区から 10 尾ずつサンプリングし,試験終 了時(200 日齢)に対照区から 177 個体,28℃-a 区から 158 個体および 28℃-b 区から75 個体をそれぞれサンプリングした。 生残率の算出 試験開始時から50 日齢,51~100 日齢,101~150 日齢,151~200 日齢まで の生残率と試験期間内での生残率をそれぞれ求めた。生残率は以下の式により 算出した。 生残率 (%) = (生残個体数/収容個体数または各期間の初日の生残個体数) ×100
生殖腺の組織学的観察および雌雄比の調査 サンプリングした全ての供試魚は全長を測定した後,ブアン氏液で固定し, 70%エタノールで保存した。固定した生殖腺を定法により 5 μm のパラフィン 切片にし,ヘマトキシリン・エオシンの二重染色を施し,光学顕微鏡下で観察 した。また,飼育水温と雌雄比の関係を明らかにするため観察にはすべての実 験区の仔稚魚を用いた。雌雄の判別には,生殖腺内に形成される卵巣腔と精小 嚢,および卵母細胞と精原細胞の有無を用いた。雌雄比は,雌に対する雄の割 合で表した。 生殖腺体指数(GSI)の算出 生殖腺体指数(GSI)は以下の式により算出した。 GSI=(生殖腺重量/体重)×100 統計処理 性比の偏りおよび生残率の有意差はχ2検定により行った。全長およびGSI の 有意差は一元配置の ANOVA の後,Tukey-Kramer 法による多重比較により求 めた。 結 果 水温の実測値 各試験における水槽の水温変化をFig. 9 に示す。試験 1 の自然水温は試験開 始時に14.0℃であったが,122 日齢には 20℃,142 日齢には 23℃に達し,試験 終了時には 25.6℃に達した(対照区)。20℃および 23℃区の加温は自然水温
が加温区の設定水温と同等になった時点で終了した。20℃区における設定水温 到達後の平均水温は19.9±0.18℃,23℃区におけるそれは 22.9±1.06℃,26℃区 では26.4±0.27℃,28℃区では 28.8±0.05℃であった。試験 2 の自然水温は試験 開始時に 15.9℃であり,試験終了時には 26.3℃であった(対照区)。26℃区の 加温は192 日齢には 26℃を超えたため終了した。28℃区は 140 日齢までにほと んどの個体がへい死したことから,140 日齢で飼育を中止した。26℃区におけ る 設 定 水 温 到 達 後 の 平 均 水 温 は 25.9±0.24 ℃ , 28 ℃ 区 に お け る そ れ は 27.8±0.20℃であった。試験 3 の自然水温は試験開始時に 15.7℃であったが,141 日齢において 23.0℃に達し,試験終了時には 27.8℃となった(対照区)。23℃ 区の加温は,自然水温が加温区の設定水温と同等となった時点で終了した。23℃ 区における設定水温到達後の平均水温は,23.0±0.62℃であった。試験 4 の対照 区における自然水温は試験開始時に16.2℃,試験終了時に 24.8℃であった。28℃ -a 区および 28℃-b 区は,それぞれ 130 日齢および 146 日齢に 28℃での飼育を 終了し,徐々に水温を低下させた後,138 日齢および 150 日齢に自然水温に戻 した。28℃-a 区における設定水温到達後の平均水温は 28.4±0.92℃,28-b 区に おけるそれは28.7±0.83℃であった。 生残および成長 試験1 における 10℃,20℃および対照区の試験終了時の生残率は,それぞれ 26.5,24.1 および 34.8%であり,対照区のそれが最も高くなった (χ2-test, P<0.05) (Table 2)。10℃および 20℃区間を除く各区の生残率には有意差が認 められた (P<0.05)。加温した試験区のうち,23℃,26℃および 28℃区では 50 日齢までに大量減耗し,生残率が著しく低下した。特に,28℃区では 20 日齢ま でに全ての仔魚がへい死し,26℃区では 40 日齢までに生残率が 0.5%となった
Fig. 10. Changes in mean total length of spotted halibut Verasper variegatus during the experiments.
Vertical bars show the standard errors. Different letters show significantly different mean values (P<0.05).
ことから飼育を中止した。試験終了まで飼育できた 23℃区の生残率は 2.5%で あった。 この試験における仔稚魚の全長は,対照区および 20℃区で 35.4±5.2 mm お よび37.7±3.0 mm であったのに対し,23℃区ではそれより劣り (P<0.05),10℃ 区では最も成長が停滞した(Fig. 10-A)。 試験2 における 26℃および 28℃区の生残率は,それぞれ 16.2%および 0.4% となり,対照区の30.7%より低い値を示した (P<0.05)。また,成長は対照区お よび 26℃区と比較して 28℃区が遅く (P<0.05),試験終了時の平均全長は対照 区,26℃区,28℃区の順に小さくなった(Fig. 10-B)。 試験 3 における対照区および 23℃区の生残率は,それぞれ 92.1%および 85.9%と高かったが,23℃区の値は対照区のそれより低かった (P<0.05)。23℃ 区および対照区の全長はほぼ同じ値を示し,試験1 のように 23℃区と対照区の 成長差はみられなかった(Fig. 10-C)。 試験4 における対照区および 130 日齢まで加温を継続した 28℃-a 区の生残率 は62.0%および 55.7%となり,ほぼ同じであったが,146 日齢まで加温を継続 した 28℃-b 区では生残率が 28.3%と低くなった (P<0.05)。平均全長は対照区 と比較して,28℃-a 区および 28℃-b 区のいずれにおいても加温期間中には小さ かったが (P<0.05),加温終了後には対照区とほぼ同じ全長を示した(Fig. 10-D)。 性分化 各試験区の100 日齢の個体の生殖腺を観察したところ,10℃区(全長 21 mm, ステージI)のそれは未分化の状態であった。20℃区(全長 38 mm,ステージ I)の生殖腺は卵巣腔および卵母細胞が発達し,雌への分化が認められた。23℃ 区(全長28 mm,ステージ I)の生殖腺には,卵巣腔および染色仁期の卵母細
胞が認められた。26℃区(全長 39 mm,ステージ I)の生殖腺に卵巣腔が観察 され,雌への性分化が認められたが,卵母細胞は染色仁期であった。28℃区(全 長23 mm,ステージ H~I)の生殖腺は未分化であった。28℃区では全長 25 mm (140 日齢)でも性分化は始まっておらず,生殖腺には生殖原細胞が数個観察さ れるにとどまった。 性比 試験1 では 23℃区において雄/雌比は 2.28 であり,雄への偏りがみられたが (χ2-test,P<0.05),10℃,20℃および対照区の性比は 1:1(雄/雌比 0.94~1.04) であった(Table 3)。26℃区および 28℃区は生残率が低く,途中で飼育を中止 したため,性比の算出に至らなかった。試験2 では 26℃区において雄が雌より も多かったが(雄/雌比 1.33),有意差は認められなかった。28℃区では生残尾 数が少なく,性比の算出には至らなかった。試験3 では 23℃区および対照区の 性比は1:1 であった。試験 4 では 131 日齢まで 28℃で飼育した後,自然水温に した28℃-a 区と対照区の性比は 1:1 であったが,146 日齢まで 28℃で飼育した 28℃-b 区は雄への偏りが認められた (P<0.05)。 GSI 試験 4 の実験終了時に全供試魚から生殖腺を摘出し,GSI を算出した (Fig. 11)。雌の GSI は 28℃-a 区および 28℃-b 区が対照区と比較して小さく (P<0.01), 28℃-b 区は 28℃-a 区と比較しても小さかった (P<0.01)。GSI は対照区,28℃ -a 区,28℃-b 区の順に小さくなり,加温期間が長くなるに従い GSI も小さくな った。雄の GSI は 28℃-a 区および 28℃-b 区が対照区と比較して小さく (P<0.05),28℃-a 区と 28℃-b 区の間では有意差は認められなかった。
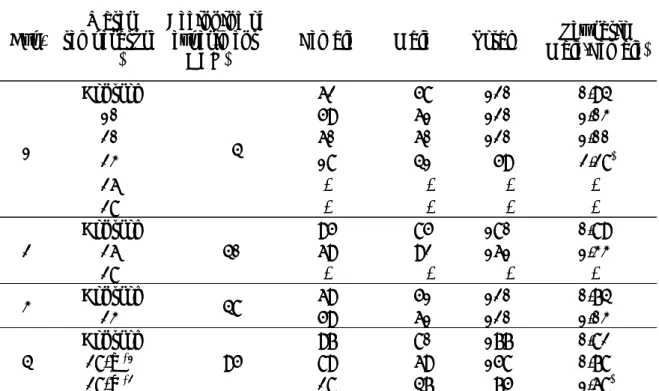
Table 3. Sex ratio of spotted halibut in each experiment Exp. temperatureWater
(℃)
Beginning of experiment
(DAH) Female Male Total
Sex ratio (Male/Female) 1 Control 4 62 58 120 0.94 10 59 61 120 1.03 20 60 60 120 1.00 23 18 41 59 2.28* 26 - - - -28 - - - -2 Control26 40 9569 8592 180161 0.89 1.33 28 - - - -3 Control23 48 6959 5161 120120 0.74 1.03 4 Control28-a *1 95 9789 8069 177158 0.82 0.78 28-b *2 28 47 75 1.68*
Control fish was reared natural water temperature. Asterisks show significant difference from theoretical 1:1 sex ratio (P<0.05) .
*1, The group started heating-up to 28℃ at 95 DAH and kept 28℃ until 131 DAH; *2, The group started heating-up to 28℃ at 95 DAH and kept 28℃ until 146 DAH. Table 3. Sex ratio of spotted halibut in each experiment
Exp. temperatureWater (℃)
Beginning of experiment
(DAH) Female Male Total
Sex ratio (Male/Female) 1 Control 4 62 58 120 0.94 10 59 61 120 1.03 20 60 60 120 1.00 23 18 41 59 2.28* 26 - - - -28 - - - -2 Control26 40 9569 8592 180161 0.89 1.33 28 - - - -3 Control23 48 6959 5161 120120 0.74 1.03 4 Control28-a *1 95 9789 8069 177158 0.82 0.78 28-b *2 28 47 75 1.68*
Control fish was reared natural water temperature. Asterisks show significant difference from theoretical 1:1 sex ratio (P<0.05) .
*1, The group started heating-up to 28℃ at 95 DAH and kept 28℃ until 131 DAH; *2, The group started heating-up to 28℃ at 95 DAH and kept 28℃ until 146 DAH.
Fig. 11. Gonadosomatic index (GSI) of female and male in experiment 4.
Vertical bars show the standard errors. Different letters show significantly different mean values (P<0.05).
考 察 試験1 においてホシガレイを 23,26 および 28℃で飼育したところ,これら の試験区における50 日齢までの生残率は対照区(水温 12.4~16.4℃)のそれと 比較して低い結果となった。これは,23℃以上の高水温が本種仔魚の生残に悪 影響を与えていることを示している。これまでにも,ホシガレイ仔稚魚の生残 率は12~21℃の範囲において水温が低いほど高い傾向を示すが(佐久間 2001), 27℃では生残率が低下することが報告されている(村田 1998)。九州西岸にお いて本種の天然仔魚が採集された際の海水温は11.1~15.7℃である(乃一ら 2006;Wada et al. 2006,2012)ことから,23℃以上の水温はホシガレイ仔魚 の好適水温から大きく外れており,それが生残率を低下させる原因であると推 察される。このような高水温による生残率の低下は,同じ異体類のヒラメ仔魚 でも報告されており,10~22 日齢のヒラメ仔魚は 20~24℃において高い生残 率を示すが,26℃以上になると生残率が低下し,27℃が半数致死温度であると される(嶋田ら2006)。日本沿岸の天然海域においてヒラメ仔稚魚が確認され た場所の水温は12~25℃であることから(下茂ら 2000),26℃以上の水温はヒ ラメの好適水温より高いと考えられる。このように本種で確認された高水温に よる生残率低下は,ヒラメ仔魚でも観察される現象であるが,本種の生残にお ける適正水温の上限はヒラメのそれよりもやや低いと考えられる。 本研究で実施した26℃および 28℃飼育において,4 日齢から加温した試験 1 では仔魚が大きく減耗し,40 日齢までに飼育を中止したが,40 日齢以降に加温 した試験2 では対照区と比較して生残率が低くなったものの試験終了まで生残 した。これは仔魚の高水温耐性が40 日齢以降に向上していることを示唆してい る。有瀧ら(2001)による仔魚の発育ステージの分類では,30 日齢には上屈前
仔魚期および上屈仔魚期であるステージD および E,40 日齢には眼の移動が開 始したステージG および有眼側にほぼ移動を終えたステージ H に達していた。 ステージG は底棲生活への移行期で,着底生活に必要な機能はほぼ完成してい ることから(有瀧ら2001),高水温耐性の向上はこのような生活様式の変化と 深く関わっている可能性が高い。また本研究において,稚魚への変態完了後の 95 日齢以降に 28℃へ加温した試験 4 の生残率は,上述した 4 日齢および 40 か ら28℃へ加温した試験 1 および試験 2 の結果よりも高くなった。これは,仔稚 魚の高温耐性が変態後の成長に伴い,上昇することを示唆している。変態時の 異体類の仔魚では,急激な生理学的・形態学的変化が引き起こされるが(堀田 ら2001),このような時期は外部環境の変動に敏感である可能性が高い。マガ レイPleuronectes herzensteini では生息水温が 0~23℃とされるものの,仔魚 飼育においては水温21℃以上では安定した飼育が行えないことから,この水温 はマガレイ仔魚にとっての適水温から外れていると述べられており(有瀧1996), 本種やマガレイの着底前の変態期仔魚にとって高水温は生残に対する負の要因 として作用すると考えられる。 本研究におけるホシガレイ仔魚の成長は,自然水温で飼育した対照区と比較 して20℃では促進されるものの,それ以外の試験区では同等もしくは劣ってお り,特に10℃および 28℃では成長が著しく停滞した。ホシガレイ仔魚の成長は, 水温9~21℃の範囲において水温が高いほど促進される(有瀧ら 2004)。また, 稚魚の飼育適水温は18~24℃であり,最適飼育水温は 21℃付近にあるとされて いる(村田ら1998)。これらの報告と本研究の結果はよく一致しており,ホシ ガレイの仔稚魚の成長促進を目的とした飼育水の加温は,20~21℃が良いと考 えられる。 ホシガレイの理論上の性比は雌雄1:1 であり(Ji et al. 2010),本研究におい
ても,10~20℃による飼育試験では 1:1 の性比が得られた。しかし,ヒラメ科 およびカレイ科魚類では性的未分化期に高水温で飼育すると性比が偏ることが 知られている。例えば,ヒラメでは40 日齢から飼育水温を 25.0~27.5℃として 飼育すると性比が雄に偏り,27.5℃では全ての個体が雄になる(山本 1992)。ま たマツカワでは,32~79 日齢(全長 10.1~29.7 mm)の個体を 14℃から 18℃ に昇温して飼育を続けると性比が雄に偏り,10.1 mm から昇温した個体では全 てが雄となる(Goto et al. 1999)。本実験ではヒラメ科およびカレイ科魚類の特 徴として考えられる性的未分化期の性決定に関するゆらぎの大きさに注目し, 高水温飼育を行うことよって雄化の誘導を試みた。その結果,4 日齢から 23℃ で飼育した個体のうち69.5%が,また 95 日齢から 28℃で飼育した個体のうち 62.7%がそれぞれ雄となったことから,他のヒラメ科およびカレイ科魚類と同 様にホシガレイの性分化には水温が影響することが明らかとなった。これは, 23℃よりも高水温で本種仔魚を飼育すれば,完全に雄化が誘導される可能性を 示している。しかし,本実験で実施した高水温飼育では生残率が大幅に低下し, ヒラメおよびマツカワのように,すべての種苗の雄化誘導には至らなかった。 本研究で用いた23~28℃の飼育に限らず,本種の仔魚は 21℃の飼育でも生残率 が低下するという(有瀧ら2004)。これは,同じ異体類であっても,性分化に 影響を与える水温域と生存に影響を与える水温域とが大きく異なることを示唆 している。さらに興味深いことに,試験2 で実施した 40 日齢からの 28℃飼育 群の140 日齢の稚魚において,生殖腺内に始原生殖細胞のみが存在する性的未 分化個体のみが観察された。このように,28℃処理は雄化を誘導する可能性を 持つものの,長期に亘る高温処理は成長および生残だけでなく生殖腺の発達そ のものを強く阻害することを示唆している。 マツカワを用いた温度処理による雄化誘導試験では,性分化に影響を及ぼす
温度感受性の獲得と消失が論じられており,全長約10 mm(32 日齢)から温度 感受性が発現した後,成長に従い感受性が低くなり,全長約41 mm(95 日齢) では昇温飼育しても性比は1:1 と変わらないことから,この時期に温度感受性は 消失すると考えられる(Goto et al. 1999;萱場 2005)。本研究では,全長 6 mm (4 日齢,B ステージ)の仔魚を 23℃で飼育すると雄化が誘導されたことから, 変態前の仔魚において性分化に影響を与える温度感受性が備わっていることが 判った。また,全長35 mm(95 日齢,I ステージ)のホシガレイ稚魚を 28℃で 全長70 mm(150 日齢)まで飼育したところ性比が雄へ偏った(雄/雌比 1.68) ことから,少なくとも全長70 mm まで温度感受性が残っていると考えられる。 雄化を誘導する温度に対する反応性は,マツカワでは14℃から 18℃への 4℃の 昇温飼育で雄化することから比較的高い。一方,ホシガレイでは15℃から約 5℃ の昇温飼育では雄化せず,8~13℃の昇温でようやく性比の雄への偏りがみられ たことから,マツカワと比べ雄化を誘導する温度に対する反応性が低いと考え られる。このように,同属のマツカワとホシガレイでも温度感受性や反応性に 違いがあることが判った。 本研究の結果は,本種が水温操作によって性転換を誘導できる魚種ではある ものの,他の異体類と比較すると,人為的な雄への性転換は容易ではない魚種 であることを示している。それは本種の性決定のゆらぎがヒラメやマツカワな どよりも少ないことに起因するのかも知れない。このような,性決定に及ぼす 遺伝的支配の強さや環境影響の感受性などを今後明らかにする必要がある。
第 3 節 メチルテストステロン投与による性転換 これまで述べてきたとおり,全雌種苗生産には遺伝的雌を雄化し偽雄を作出 することが重要である。ヒラメやマツカワ等の異体類が持つ水温依存性の性決 定を利用した雄化は本種においても可能であったが,高水温による仔稚魚の飼 育は生残にも悪影響を及ぼしたことから,効率的に雄へ誘導することができな かった。 全雌種苗の生産においては,大量の雌種苗を同時に得られることから,遺伝 的雌を雄化した偽雄の精子を用いた種苗生産が実用性の高い技術と考えられて いる。偽雄の作出には,水温調節または雄性ホルモン投与等によって遺伝的雌 を人為的に雄化することが必要である。性ホルモンによる魚類の性転換(性分 化誘導)は古くから試みられており(隆島・会田 1984),同属のヒラメではメ チルテストステロン(以下,MT)投与により雄化した偽雄を用いた全雌種苗生 産が実証されている(山本1992)。本種においても,MT 処理により作出した偽 雄の精子を用いて全雌種苗生産が可能と考えられる。しかし,MT を用いた雄へ の誘導においては,その投与量によっては正常に雄化されないことや,過剰投 与すると逆に雌化が促されたり(Nakamura 1975;Tabata 1991;山本 1992; 藤岡1993;山本 1995),生残率が低下すること(安藤ら 2008)が知られている。 従って,MT 処理を本種の雄化誘導に利用する場合には,その投与量を十分に検 討する必要がある。そこで本研究では,異なる量の MT をホシガレイ仔稚魚へ 経口投与し,生残および性分化に及ぼす MT の影響を調べた。また,併せて投 与する期間の検討も行った。
材料および方法
供試魚
平成20~23 年の 1~2 月に長崎県橘湾で漁獲された天然親魚からヒト絨毛性
生殖腺刺激ホルモン(HCG)を用いて採卵し,人工授精により得られた仔稚魚
を後述する条件で飼育して供試魚に用いた。仔魚には,シオミズツボワムシ L
型Brachionus plicatilis を 5~50 日齢,アルテミア Artemia sp.の幼生を 37~ 50 日齢,配合飼料(ラブラァバ,林兼産業)を 30 日齢から給餌し,50 日齢以 降は配合飼料のみを与えた。シオミズツボワムシとアルテミアは,栄養強化剤 (バイオクロミスリキッド,クロレラ工業)により栄養強化した。シオミズツ ボワムシの給餌期間中はナンノクロロプシス Nannochloropsis oculata を 50 cells/ml になるよう飼育水に添加した。 試験方法 MT 投与濃度の検討を試験 1 および 2 において,効果的投与期間の検討を試 験3 および 4 においてそれぞれ実施した。MT 投与試験区の配合飼料には,MT (メチルテストステロン,和光純薬工業)を溶かした 99.5%エチルアルコール を均一に添加した後,アルコールを除去するため十分に乾燥させた。対照区の 配合飼料には,試験区と同量の 99.5%エチルアルコールのみを添加し,同様の 処理を行った。これを毎日,6~8 回飽食給餌した。MT 添加濃度,投与期間を 含む飼育条件をTable 4 に示す。各試験における MT の投与開始時の仔魚の発 育段階は,有瀧ら(2001)の分類でステージ G であった。試験 1 では,MT を 0.1, 1,10 μg/g・diet で添加した区および無添加(対照区)区の計 4 区を設けた。 35 日齢の仔魚を 500 尾ずつ 0.5 kl 水槽へ収容し,40~300 日齢の期間,MT 添
加飼料を給餌した。その後,一部の稚魚はMT 無添加飼料を用いて満 2 歳まで 継続飼育した。試験2 では,MT を 0.001,0.01,0.1 μg/g・diet で添加した区 および無添加区の計4 区を設けた。7 日齢の仔魚を 5,000 尾ずつ 0.5 kl 水槽へ 収容し,40~240 日齢の期間,MT 添加飼料を給餌した。その後,一部の稚魚は MT 無添加飼料を用いて満 2 歳まで継続飼育した。試験 3 では,MT を 0.5 μ g/g・diet で添加した飼料を 46~110,130,170 および 300 日齢の期間に給餌し た区および無添加区の計5 区を設けた。40 日齢の仔魚を 1,000 尾ずつ 0.5 kl 水 槽へ収容し,MT 添加飼料を上記の期間給餌した。その後,無添加の飼料を給餌 し継続飼育した。試験4 では,MT を 0.5 μg/g・diet で添加した飼料を 42~130, 140,150,160,170,180,190 および 200 日齢の期間に給餌した区および無 添加区の計 9 区を設けた。37 日齢の仔魚を 500 尾ずつ 0.5 kl 水槽へ収容し, MT 添加飼料を上記の期間給餌した。その後,無添加の飼料を給餌し継続飼育し た。 生残率の算出 実験1 では,MT 処理終了時の 300 日齢において,実験 2 では,MT 処理終 了時の240 日齢において,実験 3 および 4 では,それぞれ試験終了時の 300 お よび200 日齢において,生残率を求めた。生残率は以下の式により算出した。 生残率 (%) = (生残個体数/収容個体数) ×100
生殖腺の組織学的観察および雌雄比の調査 各試験区の供試魚を定期的にサンプリングし,生殖腺発達を組織学的に観察 した。生殖腺の採取にあたり,仔稚魚の生殖腺のみを摘出するのは困難である ことから,個体全体をブアン氏液で固定し,その後70%エタノールで保存した。 固定した仔稚魚の生殖腺のある部位を体ごとパラフィンに包埋し,切片とした。 パラフィンへの包埋,切片の作成は常法に従った。また,切片は 5 μm とし, ヘマトキシリン・エオシンの二重染色を施し,光学顕微鏡下で観察した。雌雄 の判別は,生殖腺の外部形態および生殖腺内に形成される卵巣腔と精小嚢,お よび卵母細胞と精原細胞の有無を指標として行い,生殖腺内に卵巣および精巣 の組織がみられた個体は間性とした。 生殖腺体指数(GSI)の算出 試験1 および 2 の個体を MT 投与が終了した後もさらに継続飼育し,生殖腺 の摘出が可能となった個体を用いて,生殖腺体指数(GSI)を以下の式により算 出した。 GSI=(生殖腺重量/体重)×100 統計処理 性比の偏りおよび生残率の有意差はχ2 検定により行った。全長および GSI の有意差は一元配置の ANOVA の後,Tukey-Kramer 法の多重比較検定により 行った。
結 果 成長および生残 試験1 における生残率は,対照区,MT 0.1,1 および 10 μg 区ではそれぞれ 57.0%,58.6%,61.0%および 51.6%となり,MT 10 μg 区のそれは対照区と比 べて有意に低かった (P<0.05) (Table 5)。この試験終了時における全長は,対 照区では154.9±11.2 mm,MT 0.1 μg 区では 145.3±10.4 mm,MT 1 μg 区 では136.1±9.7 mm および MT 10 μg 区では 132.0±10.9 mm となり,MT 1 および10 μg 区は対照区および MT 0.1 μg 区より有意に小さかった (P<0.05) (Fig. 12-A)。 試験2 における生残率は,対照区,MT 0.001,0.01 および 0.1 μg 区ではそ れぞれ88.2%,85.0%,79.8%および 86.5%となり,MT 0.01 μg 区は対照区よ り有意に低かった (P<0.05) (Table 6)。200 日齢における各区の全長は,対照区 では88.1±5.5 mm,MT 0.001 μg 区では 72.0±4.6 mm,MT 0.01 μg 区で は61.8±5.7 mm,MT 0.1 μg 区では 76.7±5.6 mm となり,MT 投与区は対 照区よりも全長が有意に小さかった (P<0.05) (Fig. 12-B)。 試験3 における生残率は,対照区,MT 0.5 μg/g・diet 投与終了 110 日,130 日,170 日,300 日区では,それぞれ 80.2%,90.5%,91.6%,91.8%および 93.3% となり,対照区のそれが MT 区に比べて有意に低かった (P<0.05) (Table 7)。 MT 投与を終了した際の各区の全長は,110 日齢では 48.3±13.0 mm,130 日齢 では72.2±5.9 mm,170 日齢では 98.1±9.7 mm,300 日齢では 148.0±15.4 mm であった。300 日齢における各区の全長は,対照区では 175.7±1.73,MT 投与 終了110 日区では 160.2±15.8 mm,130 日区では 161.6±18.3 mm,170 日区 では154.3±18.6 mm,300 日区では 147.8±15.4 mm となり,MT 投与区は対
照区と比較して有意に小さかった (P<0.05) (Fig. 12-C)。 試験4 における生残率は 47.0~75.4%の範囲で,130 日齢まで MT 投与した 130 日区が最も低くなり,180 日区が最も高かった (P<0.05) (Table 8)。MT 投与期 間と生残率の間に相関は認められなかった。200 日齢における平均全長は, 113.2~127.6 mm の範囲であった。MT 投与期間と全長との間には相関は認めら れなかった (Fig. 12-D)。 MT 投与による生殖腺の変化 本試験では,正常に分化した卵巣 (Fig. 13-A),卵巣と精巣の組織が混在する
間性生殖腺 (Fig. 13. B-D),正常に分化した精巣 (Fig. 13-E)が観察された。試 験 4 においては,間性生殖腺をさらに詳細に観察したところ,以下の 3 つに分 類された。ⅰ)卵巣腔が形成されているものの,卵巣と精巣の組織が混在する 生殖腺 (Fig. 13-B),ⅱ)卵巣腔の形成が認められず,卵巣と精巣の組織が混在 する生殖腺 (Fig. 13-C),ⅲ)精巣細胞に少数の周辺仁期の卵母細胞が散在する 生殖腺 (Fig. 13-D)。 性比 試験1 における 300 日齢の結果を Table 5 に示す。MT 0.1 μg および 1 μg 区では雄が 76.2%および 79.2%出現し,雄の割合が有意に高かった (P<0.01)。 また,間性個体もそれぞれ7.4%および 2.6%出現した。しかし,10 μg 区では 間性個体3.9%を除いた雌雄比はほぼ 1:1 であり,対照区との間に有意な差は認 められなかった。試験2 の結果を Table 6 に示す。各区の性比に偏りは認められ なかった。また,この試験において間性個体は出現しなかった。試験 3 の結果 をTable 7 に示す。MT 0.5 μg/g・diet を 110 日齢まで投与した区および対照区
の雌雄比はほぼ1:1 であった。130,170 および 300 日齢まで MT を投与した区 では,雄の割合がそれぞれ67.0%,74.3%および 94.0%となり,雄への性比の 偏りが認められた (P<0.05)。試験 4 の結果を Table 8 に示す。MT を投与した 全ての区で,雄への性比の偏りが認められた (P<0.05)。MT 0.5 μg/g・diet を 130~150 日齢まで投与した区では雄が 55.4~78.7%出現した。雌もわずかに存 在したが,その他はⅰ~ⅲ段階の間性個体であった (Fig. 13-B, C, D)。160~200 日齢まで MT を投与した区では雌が出現せず,雄性生殖細胞で占められる雄個 体と間性個体のみとなった。特に180 日齢以上まで MT 投与した区では,出現 した間性個体はⅲ段階の生殖腺のみであった。 GSI 試験 1 および 2 の供試魚を継続飼育し,生殖腺を摘出して GSI を算出した (Fig. 14)。試験 1 において,MT 投与区の雌の GSI は対照区と比較して小さか った (P<0.05)。雄の GSI には各区間で有意差はなかった。また,MT 投与区に おいて間性個体が2.6~7.4%みられたが,間性個体および雄の GSI はほぼ同じ 値であり,有意差は認められなかった。 試験2 において,MT 0.001 μg および 0.01 μg 区の雌の GSI に有意差は認 められなかった。MT 0.1 μg 区の雌の GSI は,他の試験区より有意に小さかっ た (P<0.05)。
Fig. 12. Changes in mean total length of spotted halibut during the experiments.
Vertical bars show the standard errors. Different letters show significantly different mean values (P<0.05).