目 次
小特 集 825 841 849 865 873 883 889 895 901 ∼ 916 人や動物の暮らしに関わる環境研究 ―日本畜産環境学会第 16 回大会について― ………中井 裕 温泉熱利用小型メタンシステムによる コミュニティ再生………多田千佳 ベトナム国ハノイ市バヴィの酪農………森山浩光 中国亜熱帯・広東省山岳地の貧栄養土壌・ 灌木林における山羊放牧 1. 山羊放牧をする農家………塩見正衛 哺乳類の卵子と初期発生(4) ―真獣類と有袋類の特性―………菅原七郎 実践飼料学の失敗と成功(26) ―品質管理半世紀の軌跡から―………本澤清治 飼料学(150)―飼料中の有害物質 有毒植物 (poisonous plants)(1)― ………山中典子・祐森誠司・石橋 晃 Dr. Ossy の畜産・知ったかぶり(76) 希少家畜について①………押田敏雄 エクアドルにおける活動(その 3) ―インバブーラ県における酸性・塩類土壌別 化学的特性の解析と地域村民の鉄欠乏貧血 との関係(一仮説)―………冨田健太郎 畜産界ニュース………913 産業 動物畜産
の
研 究
Sustainable Livestock Production and Human Welfare
2017
株式会社
養 賢 堂
840 畜産の研究 第71巻 第10号 (2017年) 【おしらせ】
温 泉 熱 利 用 小 型 メ タ ン シ ス テ ム に よ る コ ミ ュ ニ テ ィ 再 生
多 田 千 佳
1 1 東北大学大学院農学研究科(Chika Tada)1 . は じ め に
持続可能な社会の構築には様々な産業および家庭からの環境負荷を軽減する必要がある。食 品廃棄物全体では,食品廃棄物全体では約 2,200 万 t 発生し,その約 80%は焼却・埋め立て処 理されていると推計されており 1),再生利用が求められている。食品廃棄物は含水率が高く, 焼却処理では,水分を飛ばすための熱量も使用されることになる。また,食品の原料である食 物は,地球で人間によって使用される水の 92%を農業生産で使用している2)ことからも,食品 の焼却は,生産過程で使用された莫大な水も,エネルギーをかけて蒸気にし,無駄に大気に返 すことにもなる。 水分を多く含んだ生ゴミ廃棄物の有効な活用方法としてメタン発酵がある 3)。メタン発酵は 密閉型の嫌気発酵を行うため,発酵過程でのアンモニア,メタン,および,窒素酸化物等の揮 散がなく,環境への負荷を低減できる 4)。また,発酵により発生したメタンガスはエネルギー 利用でき,さらに,残渣物には窒素,リンなどの肥料成分も含まれることから,資源循環が可 能であり,環境負荷をゴミ処理の面で削減できるだけでなく,農業生産の面でも化学肥料削減 や水の有効活用によって環境負荷削減が可能な手法である。 これまで,メタン発酵システムは,発酵槽の加温に必要なエネルギー,原料運搬や発酵残渣 の処理に必要なシステム稼働運転維持の消費エネルギーに見合ったエネルギー生産を行うため に大規模化することが一般的であった。 本研究では,より地域に身近で,誰でもハンドリング可能な分散型小型メタン発酵システム を設置することで,分散型エネルギー生産やコミュニティにおける資源循環を可能にすること を目的とした。そのために,加温にかかる熱源確保と原料運搬エネルギーの軽減,消化液の再 利用が必要である。 そこで,熱源については,温泉熱を活用することにした。日本は世界でも第 3 位の地熱資源 国であり,平成 21 年度には 3,170 ヶ所の温泉地,1 万 4,294 件の宿泊施設がある5)が,特に, 温泉が豊富に湧き出る宮城県大崎市の鳴子温泉では,かけながし湯として温泉の湯を流し続け ており,風呂から溢れ出る湯は,そのまま「どぶ」に捨てられている。本研究では,かけなが した後の湯をメタン発酵タンクの加温に活用することにした。 原料運搬については,地域住民や観光客参加型で行うことにし,自分で生ゴミを持参してタ ンクに投入できる仕組みをつくることにした。 消化液は,液肥として無料配布し,地域の方に野菜栽培や園芸に利用してもらうことにした。 本研究では,これらを宮城県大崎市鳴子温泉の温泉街に実験サイトをつくり,エネカフェメ タンというカフェを開いて地域参加型システムの小型メタン発酵システムと資源循環の実証研 究を行った。842 畜産の研究 第71巻 第10号 (2017年)
2 . 小 型 メ タ ン 発 酵 シ ス テ ム と エ ネ カ フ ェ メ タ ン
(1)温泉熱活用型小型メタン発酵システム 図 1 に小型メタン発酵システムの概略図を示す。小型メタン発酵システムは,メタン発酵タ ンク,消化液タンク,脱硫槽,脱臭槽,ガス貯留バルーン,ガス昇圧機,温泉熱交換用タンク からなる。メタン発酵システムは FRP 製でタンク容量 1.4 m3であり,煙突のように伸びた部 分が,生ゴミ投入口になっている。タンク内は 3 槽構造で,1 槽で溶解した生ゴミ液が次の槽 へ越流する仕組みである。撹拌の代わりに,第 3 槽から第 1 槽へ液体を返送するポンプが設置 されている。生ゴミ投入口には,脱臭槽へつながる排気口があり,生ゴミ投入時の臭いを極力 抑制するようになっている。本タンクは,機器の故障等を極力防ぐため,生ゴミ粉砕機は設置 していない。生ゴミは通常の料理の際に排出される大きさのまま投入され,微生物分解のみで 可溶化される。第 2 槽,第 3 槽には,微生物担体としてポリウレタン担体と炭素担体が充填さ れている。担体を入れることで,水素資化性メタン菌の付着が増え,発酵槽内の反応が安定し ている。 タンクの加温には,タンク周辺にお湯が流れるように,タンク周辺をホースでぐるぐると覆っ ている。ホース内に流れる湯は,鳴子温泉のかけ流し湯を貯留したタンクで熱交換によって加 温された湯が回っており,メタン発酵槽は 35℃前後になっている。 第 3 槽からでた消化液は消化液タンクに移動する。これらは,希望者にペットボトルやポリ タンクに入れて持って行ってもらうようにした。 脱硫槽には酸化鉄ペレットを充填し,脱臭槽には活性炭を充填した。ガス貯留バルーンは 1m3 のバイオガス用バルーン (TOYO ゴム)である。ガス昇圧機は,ドラム缶を 2 つ組み合わせて 作った。下のドラム缶 300 L に,上には 200L のドラム缶をひっくり返している。上のタンク は滑車によって,上下に動くことが可能になっている。下のドラム缶に水が入っており,バルー ンからバイオガスが供給される。上に乗ったドラム缶の重みでガス圧ができ,カフェの建物内 にあるコンロにバイオガスが供給される仕組みである。 図 1 温泉熱活用型小型メタン発酵システムの概略図ラット 2 細胞期卵子は mHECM-1 培地(Schini & Bavister 1988)で培養すると 96 時間目ま でに 97%以上が胚盤胞期に達するが TLMC では 8 細胞期から培養して胚盤胞形成,拡張期, ZP 脱出までを記録した。 表 5 初期発生時間,子宮到達,着床と妊娠期間(原著) 1細胞 2細胞 4細胞 8細胞 16細胞 桑実期 胚盤胞期 子宮到達 時期 着床 妊娠期間 hr hr hr (日) (日) 単口類 アルマジロ - - - 32 - ~62 有袋類 フクロネズミ ~40 48 66 ~72 - 84 96 24 前核期 胎子初期 12.5 着床遅延 (DI) チャアンテキ ~24 36 ~108 ~120 132 144 150 〃 〃 胎子初期 27 ヌス マドリシミント ~24 24 48 72~80 85 120 196 〃 〃 〃後期 10.7 プシス イイロジオズミ ~24 35 55 74 96~100 132 144 〃 〃 体節期 13.5 オポサム 初期 ダマヤブワラ 24~36 67.5 77.6 80 - 144 168 〃 〃 胎子 27 哺乳中着 ビー 床遅延 食中類 トガリネズミ ~50 60~64 70~81 - - ~96 96~109 96~109 胚盤胞 <10 19~20 交尾後 翼手類 コウモリ 24~43 48~72 72~120 120~140 142~192 192~216 216~240 11~12日 〃 13~14 150> 〃 吸血コウモリ ~48 48~72 96~144 144~192 ~264 264~312 ~360 ~<16日 〃後期 <18 <84 〃 短尾コウモリ >48 ~72 - ~130 ~168 ~216 240~288 ~13~16日 〃 <16 霊長類 ヒト 25 29~40 31~55 79 85 92~120 109~169 72 16~32細胞 7~12 280 赤毛サル 24 24~36 36~48 48~72 72~96 96~100 96 16〃 9 150~180 排卵後 ヒヒ ~24 >24 ~48 48~72 72~120 120~144 144~168 120~144 〃 <13 173~193 齧歯類 マウス ~24 24~38 38~50 50~60 60~70 68~80 74~89 72 桑実期 5 20~21 哺乳中DI シロアシマウス ~18 18~20 38~50 60~74 80~90 96 100 72 8~16細胞 6 22~27 ハタネズミ ~12 ~24 36> 50> 60> ~70 70> 70 8〃 5~6 21 トビネズミ ~22 22~24 50> 70~74 90~94 106 106~130 106 16~32〃 5.3/4 24~26 ハムスター ~16 16~18 24~26 36~42 48~60 60~90 ~86 60~66 8~16 5 16~17 ラット 12~20 37~61 57~65 64~87 84~92 96~120 105~120 90~98 桑実期 5~5.3/4 21~23 哺乳中DI モルモット ~30 30~35 30~75 80 - 100~115 115~140 80~85 8細胞期 5.3/4 65~71 ビスカチャ ~24 ~96 ~144 ~240 - - ~360 144 4〃 <18 145~155 食肉類 イタチ 31~50 54> 64~72 64~116 74~120 120~146 146~264 120~140 32~桑実 13~14 60~69 ミンク ~48 72 96 - 120~144 - 144~168 192 8~16細胞 25 46~70 DI ネコ ~48 72~76 76~90 - 90~96 >150 150~190 96~192 32~桑実 13~14 60~69 イヌ ~48 - - - 48~72 - 96~120 96~120 15~18 15~18 58~71 有蹄類 ウマ - 24 27~33 53~60 96~99 98~100 120~144 144~156 桑実後期 8.9w 329~345 ウシ ~30 40~50 44~65 46~90 70~141 144~ 192~216 96 8~16細胞 30~35 280 ヒツジ ~36 36~50 50~67 - 67~72 - 144~168 48~96 16〃 17~18 144~152 ヤギ ~30 30~48 60 85 98 120~140 158 98 16~32細胞 13~18 146~151 ブタ ~51 51~65 66~72 90~110 - 110~114 114 75 4~6細胞 11 112~116 ラクダ ~30 35~40 48 48~70 72 96 120 92~96 16~桑実期 14 380~401 交尾排卵 アルパカ、ラマ 30 ~72 - - - 96~120 120> 72~96 〃 12~14 344 シカ 28 28~32 - - - 120 148 160~250 DI ウ サギ類 ウサギ 12~24 24~30 26~32 32~46 40~47 47~68 68~76 72~75 桑実、胚盤 7 30~32 初期 備考
876 畜産の研究 第71巻 第10号 (2017年)
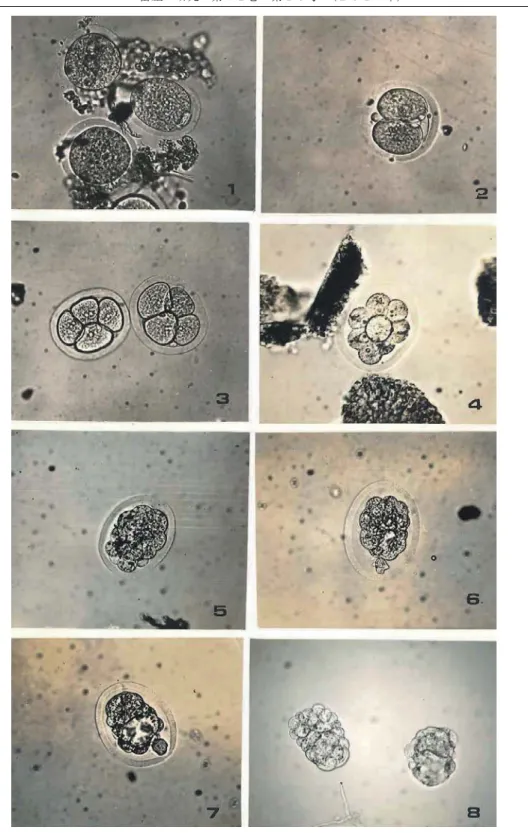
図 14 ラットの初期発生胚(光学顕微鏡)
図 15 ハムスターの初期発生(光学 LM と走査 SEM 像)(原著) 1 a,b,c:接合子 1a:透明帯表面に精子尾部 *第一極体 1b:透明帯除去した SEM 像 1C:卵細胞膜(微絨毛)にトラップされた精子頭部 2 a,b,c:2 細胞期 微絨毛は長いが割球間の連結構造は微絨毛がない 3 a,b,c:4 細胞期 各割球間の結合は微絨毛による。卵割腔も形成 4 a,b,c:8 細胞期 5 a,b,c:胚盤胞期中期 微絨毛が少ない
898 畜産の研究 第71巻 第10号 (2017年)
タヒ(図 7):タヒあるいはタキはモンゴル語で,モウコノウマ(蒙古野馬,英語: Przewalski's Wild Horse,学名: Equus ferus przewalskii)はシマウマ,ノロバを除いた唯一の現存する野生 馬とされていて,かつてアジア中央部,特にモンゴル周辺(アルタイ山脈周辺)に多数生息して いたようですが,野生では一度絶滅し,飼育個体の子孫を野生に戻す試みがモンゴル(図 8)各 地で続けられています。英語圏での別名は Asian Wild Horse または Mongolian Wild Horse とさ れています。タヒの数は最新のデータでは 300 頭程度の生息が確認されています。
この報文の最初に述べましたが,2017 年 8 月末に全日本鹿協会(事務局長は日本大学の小林信一教 授)のイベントとしてのモンゴル研修に参加(総員は 11 名)させて戴きました。ツアー自体は 4 泊 5 日 で,前後の 2 泊は首都のウランバートル市内のホテルに,中 2 泊は Hustai National Park ホスタイ(ホ スタイとはモンゴル語で白樺の意)国立公園(図 9)に隣接した草原のゲルキャンプに宿泊しました。 この国立公園はモンゴル中央部にあり,ウランバートルの西方約 100km の海抜 1,842m の草原地 帯に位置します。タヒをオランダの動物園の飼育種から 1980 年代に再導入し,保護増殖が続けられています。公 園自体は 1992 年に設立され 1998 年に国立公園になりま した。 国立公園の入口にもゲルキャンプがあり,講義室 や売店もあり,講義室ではタヒの歴史やビデオが見られ ます。ビデオは日本語上映でしたが,残念ながら売店に はビデオや CD の取扱いは無く,英語版の動物解説書が ありました。また,レンジャーの青年(図 10)が,我々の バスに乗り込んでポイントで案内をして下さいました。 図 7 群行動をするタヒ(筆者撮影) 図 8 モンゴルとホスタイ国立公園の位置 図 9 国立公園入口の案内板 図 10 若いレンジャーと著者 (中央の後ろ姿は小林教授)
また,貧血調査地域において,被験者の高貧血割合が認められた地域と先の類別塩類土壌類 別地帯とは必ずしも一致せず,類別酸性地帯にも認められたことである。したがって,酸性土 壌地帯における Fe 含有量の多様性について調査することが必要となり,以下,検討した結果を 述べていく。 5)塩基類に関しては,交換性の他水溶性も共存 これらの結果から考えられることは,図 9 にも示したが,通常,土壌コロイドのマイナス荷 電に電気的に結合している塩基類を『交換性塩基』と称し,乾燥地帯かつ低降水量地帯では, コスタ地域と異なり,この交換性の他,水溶性の塩基類も共存していることが有力的である。 交換性の塩基類は土壌 pH を増大させ,一方,水溶性は EC を増大させることで知られている。 なお,コスタ地域では,高降水量により,基本的に水溶性塩類は溶脱されてしまうので,一般 的には,EC は測定されない。 つまり,とくに,酸性土壌において,土壌 pH ならびに EC と塩基類との間に相関関係が認め られたものが存在したことから,交換性と水溶性が共存しているといえよう。とくに,K は EC と顕著な相関関係にあったことから,酸性ならびに塩類土壌において,水溶性が有力であろう (塩類土壌では pH とも 5%による相関関係より,交換性も共存)。 6)土壌 pH, EC および Fe との関係 pH と Fe に関しては,酸性土壌では相関関係がなく,塩類土壌において負の顕著な相関が認 められた。つまり,酸性土壌では pH 値に関係なく,Fe のバラツキが多く,塩類土壌では pH 値の増大に応じて Fe の有効度が減少していくということである(図 10)。 他方,Fe と EC では,酸性土壌において負の相関関係(5%)が認められ(図 11),これは, 水溶性塩基類の増大も Fe の有効度を低下させる要因であることが考えられる。 7)Fe と塩基類との関係 Fe と塩基類について見ると,酸性土壌において,Fe と K,Fe と Mg に顕著な負の相関関係 が認められ,さらに,塩類土壌では Fe と Mg にも顕著な負の相関関係があった(図 12 および 図 14)。他方,Ca と Fe に関しては,両土壌とも相関関係は認められなかった(図 13)。 Ca2+ Ca2+ Ca2+ Mg2+ Mg2+ Ca2+ Mg2+ Ca2+ Mg2+ Mg2+ Ca2+ Mg2+ Ca2+ Mg2+ Ca2+ Ca2+ Mg2+ Mg2+ コスタ地域 シエラ地域 図 9 コスタおよびシエラ地域における土壌コロイドに存在する塩基類のモデル
908 畜産の研究 第71巻 第10号 (2017年) これらのことより,Fe の沈殿は,水溶性ならびに交換性塩基の共存が大きく関係し,とくに, Mg の影響が大であると推察している。 酸性土壌 塩類土壌 図 10 土壌タイプ別における pH (H2O)と Fe との相関関係 注釈: *相関係数において 5%による有意差あり。** 1%による有意差あり。 酸性土壌 塩類土壌 図 11 土壌タイプ別における EC と Fe との相関関係 注釈: *相関係数において 5%による有意差あり。** 1%による有意差あり。 酸性土壌 塩類土壌 図 12 土壌タイプ別における交換性 K と Fe との相関関係 注釈: *相関係数において 5%による有意差あり。** 1%による有意差あり。