2.6.1 緒言 2.6.1.1 血管内皮増殖因子 血管内皮増殖因子(VEGF)は、血管内皮細胞にとって不可欠の増殖因子であり1)、内皮細胞の 分裂促進因子として機能するのみならず、既存の血管からの新たな血管形成(血管新生)や血管 系の初期形成(脈管形成)のプロセスである内皮細胞の遊走、生存や管腔形成を促進する。また、 VEGF は血管透過性を亢進させる。さらに、VEGF は正常な血管形成や生理的な血管新生において 重要な役割を果たすほか、様々な疾患においても、病的な血管新生や血管透過性の亢進に深く関 わっていることが知られている。 VEGF (「VEGF-A」として記される場合もある)には、一連の関連たん白質の存在が知られてい る。ヒトや哺乳類の VEGF ファミリーには、胎盤増殖因子(PlGF)、VEGF-B、VEGF-C や VEGF-D がある。これらの VEGF ファミリーはいずれも二量体を形成し、3 種の受容体型チロシンキナー ゼ[VEGFR-1(flt-1)、VEGFR-2(flk-1)及び VEGFR-3]のいずれか又は複数に結合する1,2)。こ れら受容体へのリガンドの結合は、受容体の二量体化、自己リン酸化及び細胞内シグナル伝達を 誘導し、細胞機能を調節する。
VEGFR-2 は主として血管内皮細胞に発現しているが、VEGFR-1 は血管内皮細胞のほかに、好中 球、単球、マクロファージや周細胞などにも発現している。VEGFR-1 は、VEGFR-2 よりも VEGF に 対する高い結合親和性を有しているものの、VEGF による二量体化で生じる内皮細胞の VEGFR-1 活性化は弱い。したがって、すべての VEGF(VEGF-A)アイソフォームの作用は、主に VEGFR-2 の活性化を介して生じると考えられる。VEGFR-1 は関連分子である PlGF や VEGF-B にも結合する。 VEGF と異なり、PlGF や VEGF-B は正常の血管形成や成熟動物の生理的な血管新生に必須ではない。 しかしながら、VEGF と同様に PlGF 及び VEGF-B は病的な血管のリモデリングに関与している1)。 その他の VEGF ファミリーである VEGF-C や VEGF-D は VEGFR-3 に高親和性で結合する。VEGFR-3 はリンパ管内皮細胞に主として認められ、VEGF-C や VEGF-D は主にリンパ管新生の調節に関与し ていると考えられる2)。 VEGF は単一遺伝子によってコードされているが、選択的スプライシングやたん白質分解切断 により生じるいくつかの異なるアイソフォームが存在する1)。それぞれの VEGF アイソフォームは いずれも二量体として機能するものの、その大きさ並びにヘパリンやニューロピリンと呼ばれる アクセサリーたん白質(非シグナル性の結合たん白質)への結合能には違いがある。例えば、ヒ トで最も主要なアイソフォームである VEGF165 は、ヘパリンやニューロピリンに結合する。 VEGF121も多くの組織や病的な状態で高レベルに発現するが、ヘパリンやニューロピリンに結合す るドメインを欠いている1,2)。いずれの VEGF アイソフォームも主として VEGFR-2 への結合とその 活性化を介して作用を発現するものの、血管パターンの制御過程において異なる役割を担ってい る。VEGF と同様に、PlGF も複数のアイソフォームが選択的スプライシングによって生じ、ヘパ リンやニューロピリンへの結合能に違いが認められる。 臨床データ及び疫学データあるいは非臨床試験結果から、VEGF が、滲出型(血管新生)加齢 黄斑変性(AMD)、糖尿病網膜症及び虚血性網膜症などの眼性血管疾患の特徴である血管透過性 亢進、浮腫及び血管新生において重要な役割を果たしていることが示された。例えば、動物の眼 内への VEGF 投与、あるいはトランスジェニック法により局所的 VEGF 過剰発現を誘導すると、異 常な血管漏出及び眼血管新生が生じる3,4)。また、脈絡膜新生血管(CNV)の動物モデルでは、 VEGF シグナルの阻害により CNV の発症が強力に抑制されたことから、CNV 発症に VEGF 刺激が必
要であることが示唆されている5,6)。同様に、虚血性網膜症及び糖尿病網膜症の動物モデルにお いて、VEGF を薬理学的に中和すると、血管新生及び血管漏出の発現抑制あるいは回復が認めら れる7,8)。 VEGF は病的な眼血管新生や網膜浮腫に至るプロセスに関わる基本的因子であることから、 VEGF の阻害はこれらの病態を特徴とする疾患に対する有望な治療法の一つとなることが期待さ れる。また、VEGF に加え、関連の血管新生たん白質である PlGF もまた眼血管新生及び血管漏出 に関与していると考えられている9,10) 2.6.1.2 VEGF Trap-Eye
VEGF Trap(アフリベルセプト)は、ヒト VEGFR-1 の第 2 Ig ドメインとヒト VEGFR-2 の第 3 Ig ドメインをヒト IgG1 の Fc ドメインに直列に融合した新規の組換えヒト融合たん白質である (図 2.6.1- 1)。組換え VEGF Trap たん白質(ホモ二量体糖たん白質)は、チャイニーズハム スター卵巣(CHO)K1 細胞で発現させ、ろ過及びクロマトグラフィー法で精製する。
図 2.6.1- 1 VEGF Trap の構造
VEGFR-1 及び VEGFR-2 は互いに近縁の受容体であり、それぞれ細胞外に 7 個の Ig ドメインと細胞内に 1 個のチロ シンキナーゼドメインを有する。VEGF Trap には、融合した VEGFR-1 の第 2 Ig ドメイン及び VEGFR-2 の第 3 Ig ドメイン、並びにそれらと融合した IgG1 の Fc 領域が含まれている。
VEGFR-1 と VEGFR-2 それぞれのリガンド結合に重要なドメインを取込むことによって、VEGF Trap はすべての VEGF アイソフォームに結合するようにデザインされた11)。実際に、VEGF Trap
は VEGF に結合し、その活性を阻害することが様々な試験において確認されている。また、VEGF Trap は VEGF のヘパリン結合アイソフォーム及びヘパリン非結合アイソフォームいずれにも同等 に結合した[解離定数(KD)は、VEGF165:0.5pM、VEGF121:0.36pM]。VEGF に対する高い結合親 和性により、VEGF Trap は VEGF と複合体を形成し、VEGF とその細胞表面受容体との相互作用を 阻害する。VEGF Trap は、VEGFR-1 のリガンドである PlGF-2 及び VEGF-B にも高い結合親和性 (KD:それぞれ 39pM 及び 1.92pM)を有している。VEGF Trap の VEGF 及び PlGF に対する結合は、
in vivo においてそれらリガンドとその内因性 VEGF 受容体との相互作用を阻害し、異常な血管
新生及び血管漏出を抑制すると考えられる。
VEGF Trap-Eye は、硝子体内投与用として高純度に精製された VEGF Trap の等浸透圧性製剤で あり、滲出型 AMD、糖尿病黄斑浮腫及び網膜血管閉塞性疾患を有する患者を対象に臨床試験が実 施されている。これらの疾患は先進国における中途失明の主な原因となっている。臨床試験では、 VEGF Trap-Eye (VEGF Trap として)0.5mg 又は 2.0mg を月 1 回又はそれ以下の頻度で罹患眼に 硝子体内投与した。本概要に記載の非臨床試験プログラムは、これら疾患の適応に VEGF Trap-Eye を使用する根拠及び安全性を裏付けるために計画された。
VEGF Trap は、ヒトの VEGF 及び PlGF と同程度の親和性でマウス、ラット、ウサギ及びサルの VEGF 及び PlGF に結合することから、これらの動物種において非臨床の薬理試験、薬物動態試験 及び毒性試験を実施した。本概要中で示した VEGF Trap の非臨床試験のほか、公表されている非 臨床試験4,12,13)において、滲出型 AMD、糖尿病網膜症及び虚血性網膜症の動物モデルにおける異 常な血管透過性及び血管新生に対する VEGF Trap の抑制効果が検討された。いずれのモデルにお いても、VEGF Trap は全身又は硝子体内投与で異常な血管透過性及び血管新生を効果的に抑制し た。非臨床安全性試験の評価を裏付け、VEGF Trap-Eye の臨床使用を支持する知見を得るために、 硝子体内投与による薬物動態試験を実施し、VEGF Trap の in vivo における分布及び消失につい て全身曝露も含めて評価した。VEGF Trap の毒性プロファイルは、マウス、ラット、ウサギ及び サルを含む複数の動物種を用いた単回及び反復投与毒性試験によって評価した。また、VEGF Trap の安全性プロファイルを硝子体内、皮下及び静脈内を含む複数の投与経路により評価した。 さらに、硝子体内投与による VEGF Trap-Eye の眼に対する慢性毒性をサルを用いて評価した。サ ルの網膜は構造的及び機能的にヒトの網膜と類似しているため、サルをこれらの試験に用いる動 物種として選択した。また、サルの硝子体は比較的容量が大きいため、ヒト臨床用量よりも高い 投与量で VEGF Trap-Eye を反復硝子体内投与したときの影響が評価可能である。本概要に示す一 連の非臨床試験により、硝子体内投与した VEGF Trap は、網膜及び脈絡膜に効果的に曝露すると ともに、全身曝露を最小限にし、かつ安全性プロファイルが良好であることが確認された。 VEGF Trap-Eye の申請効能・効果は、「中心窩下脈絡膜新生血管を伴う加齢黄斑変性」であり、 申請用法・用量は、「アフリベルセプト(遺伝子組換え)として 2mg(0.05mL)を 1 ヵ月ごとに 1 回、連続 3 回(導入期)硝子体内投与する。その後の維持期においては、通常、2 ヵ月ごとに 1 回、硝子体内投与する。なお、症状により投与間隔を適宜調節する。」である。
略語一覧
略 語 英 語 名 称 日 本 語 名 称
ADA Anti-drug antibody 抗薬物(VEGF Trap)抗体 ADCC Antibody dependent cell-mediated
cytotoxicity
抗体依存性細胞傷害(作用) AMD Age-related macular degeneration 加齢黄斑変性
ALP Alkaline phosphatase アルカリホスファターゼ
AST Aspartate aminotransferase アスパラギン酸アミノトランスフェラーゼ ALT Alanine aminotransferase アラニンアミノトランスフェラーゼ AUC Area under the concentration-time
curve
濃度-時間曲線下面積 AUC0-∞ Area under the concentration-time
curve (from time 0 to infinity)
濃度-時間曲線下面積(0 から無限大時 間)
AUC(0-x) Area under the concentration-time curve over x h
濃度-時間曲線下面積(0 から x 時間) bFGF Basic fibroblast growth factor 塩基性線維芽細胞増殖因子
BSA Bovine serum albumin ウシ血清アルブミン
CDC Complement dependent cytotoxicity 補体依存性細胞傷害(作用) CHO Chinese hamster ovary チャイニーズハムスター卵巣
CL Clearance クリアランス
Cmax Maximum serum or plasma (drug) concentration
最高血清/血漿中(薬物)濃度 CNV Choroidal neovascularization 脈絡膜新生血管
CTAD Sodium citrate, theophyline, adenosine, dipyridamole
クエン酸ナトリウム、テオフィリン、アデ ノシン、ジピリダモール
ECG Electrocardiogram 心電図
EDTA Ethylenediamine tetraacetic acid エチレンジアミン四酢酸 ELISA Enzyme-linked immunosorbent assay 酵素免疫測定法
ERG Electroretinogram 網膜電図
F Bioavailability バイオアベイラビリティ
FA Fluorescein angiography フルオレセイン蛍光眼底造影 FSH Follicle stimulating hormone 卵胞刺激ホルモン
GD Gestation day 妊娠日
GGT Gamma-glutamyl transferase ガンマーグルタミルトランスフェラーゼ GLP Good laboratory practice 医薬品の安全性に関する非臨床試験の実施
の基準
hRF Human rheumatic factor ヒトリウマチ因子
HRP Horse radish peroxidase 西洋ワサビペルオキシダーゼ HUVEC Human umbilical vein endothelial
cells
ヒト臍帯静脈内皮細胞 IC50 Concentration inducing 50% of
inhibition
略 語 英 語 名 称 日 本 語 名 称 ICH International conference on
harmonization of technical requirements for registration of pharmaceuticals for human use
日米 EU 医薬品規制調和国際会議
Ig Immunoglobulin 免疫グロブリン
IM Intramuscular 筋肉内
IOP Intraocular pressure 眼圧
IP Intraperitoneal 腹腔内 IU International unit 国際単位 IVT Intravitreal 硝子体内 IV Intravenous 静脈内 KD (Equilibrium) Dissociation constant 解離定数 LH Luteinizing hormone 黄体形成(化)ホルモン LLOQ Lower limit of quantitation 定量下限
LOAEL Lowest observable adverse effect level
最小毒性量
LYO lyophilized 凍結乾燥剤
MRT Mean residence time 平均滞留時間 NOAEL No observed adverse effect level 無毒性量
OIR Oxygen-induced retinopathy 酸素誘発虚血性網膜症 PEG Polyethylene glycol ポリエチレングリコール PDGF Platelet-derived growth factor 血小板由来増殖因子 PlGF Placental growth factor 胎盤増殖因子 PS20 Polysorbate 20 ポリソルベート 20
SC Subcutaneous 皮下
SCID Severe combined immunodeficiency 重症複合免疫不全
SD Sprague Dawley -
S.D. Standard deviation 標準偏差
t1/2 Terminal elimination half-Life 終末相における消失半減期
TK Toxicokinetics トキシコキネティクス
Tmax Time to reach maximum (serum/plasma) concentration
最高(血清/血漿中)濃度到達時間 VEGF Vascular endothelial growth factor 血管内皮増殖因子
VEGFR Vascular endothelial growth factor receptor
血管内皮増殖因子受容体 Vss Volume of distribution at steady
state
参考文献一覧
1) Takahashi H et al., Clin Sci 2005:109:227-241 2) Alitalo K et al., Cancer Cell 2002:1:219-227 3) Miller JW et al., Diabetes Metab Rev 1997:13:37-50
4) Saishin Y et al., J Cell Physiol 2003:195:241-248 ([参考]4.2.1.1-5 参照) 5) Kwak N et al., Invest Ophthalmol Vis Sci 2000:41:3158-3164
6) Krzystolik MG et al., Arch Ophthalmol 2002:120:338-346 7) Aiello LP et al., Proc Natl Acad Sci USA 1995:92:10457-10461 8) Qaum T et al., Invest Ophthalmol Vis Sci 2001:42:2408-2413 9) Carmeliet P et al., Nat Med 2001:7:575-583
10) Rakic JM et al., Invest Ophthalmol Vis Sci 2003:44:3186-3193 11) Holash J et al., Proc Natl Acad Sci USA 2002:99:11393-11398
12) Cursiefen C et al., J Clin Invest 2004:113:1040-1050 ([参考]4.2.1.1-10 参照) 13) Cao J et al., Invest Ophthalmol Vis Sci 2010:51:6009-6017 ([参考]4.2.1.1-6 参照)
2.6.2 薬理試験の概要文の目次
2.6.2.1 まとめ ... 3
2.6.2.2 効力を裏付ける試験 ... 5
2.6.2.2.1 In vitroにおける作用 ... 6
2.6.2.2.1.1 VEGF Trap と VEGF ファミリー関連リガンドの結合... 6
2.6.2.2.1.2 HUVEC における VEGF-A165依存性受容体リン酸化の阻害 作用 ... 7 2.6.2.2.1.3 VEGF 誘発カルシウム動員の阻害作用... 8 2.6.2.2.1.4 VEGF Trap の補体依存性細胞傷害及び抗体依存性細胞傷 害作用 ... 8 2.6.2.2.2 In vivoにおける作用 ... 9 2.6.2.2.2.1 マウスにおける脈絡膜新生血管及び VEGF 誘発血液網膜 関門破綻に対する VEGF Trap の抑制作用 ... 9 2.6.2.2.2.2 網膜下マトリゲルモデルラットにおける VEGF Trap によ る CNV の形成抑制、CNV の退行、並びに CNV に付随する 炎症及び線維化の抑制 ... 13 2.6.2.2.2.3 サルにおける VEGF Trap による実験的 CNV の形成抑制並 びに既存の活動性病変の血管漏出の速やかな抑制 ... 14 2.6.2.2.2.4 VEGF Trap 投与による糖尿病ラットにおける網膜血管漏 出の改善 ... 16 2.6.2.2.2.5 VEGF Trap による網膜虚血マウスにおける病的な血管新 生の抑制 ... 17 2.6.2.2.2.6 VEGF Trap による角膜傷害マウスにおける炎症及び血管 新生の抑制 ... 19 2.6.2.3 副次的薬理試験 ... 21
2.6.2.3.1 正常及び担癌マウスにおける遊離型 VEGF Trap 及び VEGF Trap 複合体の濃度、並びに抗腫瘍効果との相関性... 21 2.6.2.4 安全性薬理試験 ... 24 2.6.2.4.1 中枢神経系 ... 24 2.6.2.4.2 心血管機能 ... 24 2.6.2.4.2.1 げっ歯類における血圧に及ぼす VEGF Trap の影響:遊離 型 VEGF Trap の血中濃度との相関性 ... 24 2.6.2.4.2.2 カニクイザルにおける心血管機能に及ぼす VEGF Trap の 影響 ... 25 2.6.2.4.3 血栓形成 ... 26 2.6.2.4.4 呼吸機能 ... 26
2.6.2.4.5 創傷治癒 ... 26
2.6.2.5 薬力学的薬物相互作用 ... 27
2.6.2.6 考察及び結論 ... 28
2.6.2.7 図表 ... 31
2.6.2 薬理試験の概要文 2.6.2.1 まとめ 血管内皮増殖因子(VEGF)は、眼血管新生など血管新生を特徴とする様々な疾患を初めとして、 生理的及び病的な新生血管の形成に重要な役割を果たしている1)。また、病的新生血管において VEGF は血管透過性の亢進や浮腫を誘発することも知られている2)。 VEGF Trap(アフリベルセプト)は、血管内皮増殖因子受容体-1(VEGFR-1)の第 2 Ig ドメイ ンと血管内皮増殖因子受容体-2(VEGFR-2)の第 3 Ig ドメインを、ヒト免疫グロブリン G1 (IgG1)の定常領域(Fc)に融合させた組換えたん白質である。VEGF Trap は分子量 97 キロダ ルトン(kDa)のたん白質に %程度の糖鎖が付加された、総分子量 115kDa の二量体糖たん白 質である。チャイニーズハムスター卵巣(CHO)K1 細胞で発現させた組換え VEGF Trap を、ろ過 及びクロマトグラフィーで精製し、硝子体内投与(IVT)用に製剤化した VEGF Trap-Eye を製造 する。
VEGF Trap はヒト VEGF に対し高い親和性を有しており、VEGF-A165及び VEGF-A121各アイソ フォームに対する解離定数(KD)はそれぞれ 0.5pM 及び 0.36pM である。VEGF Trap は、VEGF に 結合して複合体を形成することにより、VEGF とその細胞表面受容体との相互作用を阻害する。 ヒト胎盤増殖因子-2(2)及び VEGF-B も VEGFR-1 のリガンドであるが、VEGF Trap は PlGF-2(KD:39pM)及び VEGF-B(VEGF-B(10-108)の KD:1.92pM)に対しても高親和性で結合する(表 2.6.2- 1、2.6.3.2、4.2.1.1-1 VGFT-MX-08022、4.2.1.1-11 VGFT-MX-11016 参照)。
VEGF Trap は、VEGF による血管透過性亢進及び血管新生を阻害することが非臨床薬理試験にお いて示されている。In vitro において VEGF Trap はヒト、ウサギ、ラット及びマウスの VEGF-A に高親和性で結合し、ヒト臍帯静脈内皮細胞(HUVEC)の VEGF 誘発による受容体リン酸化及びカ ルシウム動員を阻害する。In vivo においては、既報の成績3,4,5)や後述のとおり、VEGF Trap の 全身又は硝子体内(IVT)投与で、異常な血管漏出及び病的な血管新生が効果的に抑制され、脈 絡膜新生血管(CNV)、糖尿病網膜症、虚血性網膜症及び角膜傷害などの眼性血管疾患の動物モ デルに付随する炎症性反応が回復した。特に、サルのレーザー誘発 CNV モデルにおいて既に形成 されている脈絡膜病変部位の血管漏出が、VEGF Trap 単回硝子体内投与により、速やかに回復し た。また、マトリゲル網膜下投与により誘発した CNV ラットモデルにおいて、VEGF Trap は新生 血管の退行を促進することが示された。
VEGF Trap は全身投与でも、病的血管新生や血管漏出を伴う他の in vitro 及び in vivo モデ ル、例えば、乾癬6)、角膜移植拒絶7)及び腫瘍増殖8)などのモデルにおいて、薬理効果を発揮す ることが確認されている。腫瘍の同種又は異種移植マウスモデルを用いた用量反応性試験におい て、腫瘍増殖を著明に抑制するのに必要な VEGF Trap の皮下投与量は 2.5mg/kg(週 2 回)以上 であった8)。効果が認められた投与量における血中「遊離型」(非結合型)VEGF Trap 濃度は、 投与期間中 VEGF Trap 複合体の最大濃度(1~2μg/mL)を超えるレベルで維持されていた。 VEGF は、血管の様々な病態に関わるほかに、血圧調節にも関与することが知られており、 VEGF 阻害作用を有する薬剤を、その薬理効果を示す用量で全身投与すると、VEGF 阻害剤のクラ スエフェクトとして血圧上昇がみられることが知られている9,10)。VEGF Trap も同様に、げっ歯 類への単回皮下投与で、収縮期血圧と拡張期血圧に用量に相関した上昇が認められた。ラットで は、0.5mg/kg 以上で血圧の用量比例的な上昇がみられ、10mg/kg 以上で最大上昇値 15~20mmHg に達した。血圧上昇の持続期間は作用用量範囲(0.5~25mg/kg)で用量に応じて延長し、また遊
離型 VEGF Trap の血漿中濃度と相関していた。血中の遊離型 VEGF Trap 濃度がおよそ 1μg/mL を 下回ると血圧はベースラインレベルに回復した。 サルにおける VEGF Trap の反復皮下投与又は静脈内投与時の安全性薬理エンドポイントを評価 した結果、中枢神経系又は心電図(ECG)パラメータに影響は認められなかった。ウサギを用い た電気的傷害誘発血栓症モデルにおいて、VEGF Trap は 30mg/kg までの反復静脈内投与で静脈及 び動脈血栓形成に影響を及ぼさなかった。ラットにおける VEGF Trap 250mg/kg までの単回静脈 内持続投与により、呼吸機能パラメータに影響は認められなかった。しかしながら、ウサギの切 開創及び切除創モデルにおいて、VEGF Trap は反復静脈内投与により、検討したすべての用量 (0.3~30mg/kg)で創傷治癒を抑制した。いずれの動物モデルでも、創傷治癒パラメータに対す る VEGF Trap の影響はおおむね用量依存的で、0.3mg/kg 投与群では投与初期の観察時点で軽微 な抑制がみられた。一方、3 及び 30mg/kg 投与群では創傷治癒の抑制はより顕著で、試験期間中 持続した。このように、高用量投与群で創傷治癒のより強くかつ持続的な抑制がみられたが、こ れは遊離型 VEGF Trap の平均血漿中濃度と相関しており、投与後すべての評価時点で遊離型 VEGF Trap 濃度が VEGF Trap 複合体濃度よりも高かった。これに対して、0.3mg/kg 投与群では遊 離型 VEGF Trap の平均血漿中濃度は一時的に VEGF Trap 複合体濃度を上回ったのみであった。
上記のように、VEGF Trap が血圧、創傷治癒及び腫瘍増殖に及ぼす影響は、眼内の病的な血管 新生及び血管漏出を阻害するのに必要な VEGF Trap 硝子体内投与量を大きく上回る投与量を全身 投与したときに認められる。そのような影響が生じ得る投与量(全身投与)における血中遊離型 VEGF Trap 濃度は、臨床試験で硝子体内投与後に予想される濃度、あるいはサルの VEGF Trap 硝 子体内投与(体重補正)後の濃度(表 2.4.3-2)をはるかに上回る濃度(1~2μg/mL 以上)に達 している。したがって、VEGF Trap 硝子体内投与は、眼外組織において VEGF が介在する生物学 的作用のプロセスには意味のある影響を及ぼさないと考えられる。
2.6.2.2 効力を裏付ける試験
VEGF-A(単に「VEGF」として記される場合もある)は VEGF/PDGF(血小板由来増殖因子)遺 伝子ファミリーの一つであり、正常な血管形成や様々な病態において、血管透過性亢進及び血管 新生を誘導する。その他の VEGF ファミリーには VEGF-B、VEGF-C、VEGF-D、VEGF-E 及び PlGF など がある。VEGF-A は主に、2 つの膜貫通型の受容体型チロシンキナーゼ VEGFR-1 及び VEGFR-2 に結 合し、活性化することで作用を発揮する。3 つめの関連する受容体 VEGFR-3 は主にリンパ管形成 の調節に関与しているが、この受容体には主に VEGF-C 及び VEGF-D が結合し、VEGF-A の結合は弱 い11)。 VEGF-A は主として VEGFR-2 を介したシグナル伝達により、血管内皮に作用を及ぼすと考えられ ている1)。VEGFR-2 がリガンドを介して二量体を形成することにより受容体が自己リン酸化され、 ホスファチジルイノシトール 3'キナーゼ/Akt などの経路を介してシグナルが下流に伝達される。 VEGFR-2 は主に血管内皮細胞の細胞表面に存在しており、細胞の生存、遊走や増殖に重要な役割 を果たしている。VEGF-A は当初、内皮のバリア機能を低下させ、様々な病態において水や高分子 を血管外へ滲出させる血管透過性因子として発見された2)。VEGFR-1 は、VEGF-A やその類縁分子 である PlGF 及び VEGF-B にも結合する。VEGFR-1 の VEGF-A に対する親和性は VEGFR-2 よりも高い が、同様の条件下で比較すると、VEGF-A を介した VEGFR-1 の自己リン酸化は VEGFR-2 よりも弱い。 VEGFR-1 活性化が血管内皮で弱いことは、VEGFR-1 の直接的な血管透過性亢進作用及び血管新生 作用が弱いことと一致する12)。しかし、VEGFR-2 とは異なり、VEGFR-1 は内皮前駆細胞のほか、 ある種の周細胞、平滑筋細胞や白血球など内皮細胞以外の様々な細胞にも発現している。特に、 VEGF 及び PlGF は、様々な病態で白血球、特に単球の強力な化学走化性因子及び活性化因子とし て機能する。したがって、内皮細胞への直接作用による血管新生と血管透過性亢進に加え、VEGF と PlGF は血管の構造及び機能の調節に関与する様々な細胞に対して、VEGFR-1 を介して作用を 及ぼすことが可能である12)。PlGF は VEGF と異なり、正常な血管形成には重要な役割を担ってい ないと考えられているが、様々な病態において病的な血管新生や血管漏出を惹起することが知ら れている13,14)。また、VEGF と同様、PlGF はヒトの CNV 膜中に存在し、動物での実験的 CNV 形成 に寄与していることが確認されている15)。さらに、VEGF-B も動物モデルにおいて、網膜及び脈 絡膜での血管新生や血液網膜関門の破綻に関与することが報告されている16)。
VEGF Trap は、ヒト VEGFR-1 の第 2 Ig ドメインとヒト VEGFR-2 の第 3 Ig ドメインを、ヒト IgG1 の定常領域に融合した組換えたん白質である。VEGF Trap は VEGF-A に強く結合することで VEGF-A とその受容体との相互作用を阻害し、VEGF-A の生物学的作用を抑制する。また、VEGF Trap は他の VEGFR-1 リガンドである PlGF にも結合する。
VEGF Trap のリガンド結合特性を明らかにし、VEGF 受容体を発現しているヒト血管内皮細胞を 用いて、VEGFR-2 の VEGF 依存性リン酸化に及ぼす VEGF Trap の影響を検討した。また、眼性血管 疾患の様々な動物モデルを用いて、病的な血管漏出及び血管新生に対する VEGF Trap の効果を検 討した。
2.6.2.2.1 In vitro における作用
2.6.2.2.1.1 VEGF Trap と VEGF ファミリー関連リガンドの結合
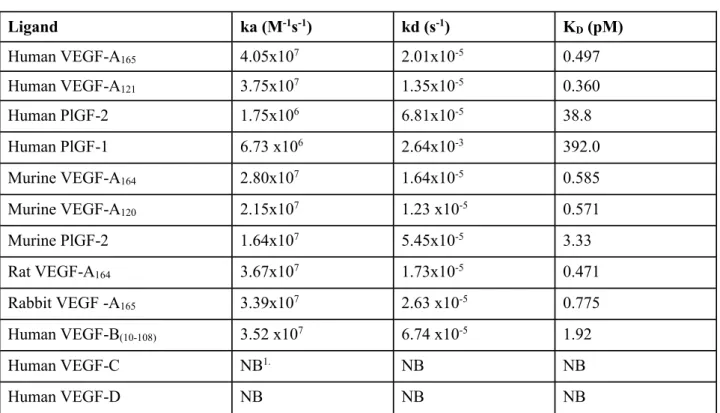
参照項目:4.2.1.1-1 VGFT-MX-08022 4.2.1.1-2 VGFT-MX-08021 4.2.1.1-11 VGFT-MX-11016 [参考]4.2.1.1-12 Papadopoulos manuscript Oct 31 2011 VEGF Trap はヒト VEGF-A165及びヒト PlGF-2 と高親和性で結合する組換え融合たん白質である。 ヒト由来の VEGFR-1 に結合し得るヒト VEGF/PlGF リガンドファミリーの他のアイソフォーム (例えばヒト VEGF-A121、ヒト PlGF-1 など)も VEGF Trap に結合することが確認されている。さ らに、VEGF Trap はマウス、ラット、ウサギなどのヒト以外の動物種の VEGF 及び PlGF に対して も同等の親和性で結合することから、これらの動物種を用いて非臨床薬理試験及び毒性試験を実 施することが可能である。なお、カニクイザル VEGF-A に対する VEGF Trap の結合親和性につい ては検討していないが、サルの VEGF-A のアミノ酸配列はヒトと同一であることから、ヒト及び サルの VEGF-A と VEGF Trap の結合作用に差はほとんどないと考えられる。
VEGF Trap と VEGF ファミリーの 12 のリガンドとの結合について検討するため、表面プラスモ ン共鳴法(BiaCore 法)を用いて解離定数(KD)を求めた。リガンドには、ヒト VEGF-A165、ヒト VEGF-A121、ヒト PlGF-1、ヒト PlGF-2、ヒト VEGF-B(10-108)、ヒト VEGF-C、ヒト VEGF-D、マウス VEGF-A164、マウス VEGF-A120、マウス PlGF-2、ラット VEGF-A164及びウサギ VEGF-A165を用いた。
BiaCore 法による測定は、デキストラン被覆チップ(CM-5)を使用した BiaCore 3000 又は 2000 装置を用いて実施した。アミンカップリング法を用いて、Protein A を CM-5 チップにカッ プリングさせた。VEGF Trap の結合親和性を BiaCore 法を用いて測定するため、まずアミンカッ プリングさせた Protein A の表面に VEGF Trap(0.625μg/mL)を注入した(100μL/分)後、さ らに VEGF Trap の表面に様々な濃度の VEGF ファミリーのリガンドを注入した(50 又は 100μL/ 分)。得られたデータは Scrubber 2 software を用いて処理し、Biaevaluation 4.1 software により解析した。
VEGF Trap は、検討した 12 リガンド中 10 のリガンドに対し特異的に結合した。一方、ヒト VEGF-C 及びヒト VEGF-D に対しては測定可能なレベルの結合を示さなかった。また、VEGF Trap は ヒト、マウス、ラット、ウサギ由来の VEGF-A に対して、高親和性(KD値は 1pM 未満)で結合し た。また、ヒト及びマウス PlGF に対しても VEGF Trap は高親和性を示した。リガンドと VEGF Trap の結合に対する KD値を表 2.6.2- 1に示す(2.6.3.2 参照)。
表 2.6.2- 1 VEGF Trap と VEGF ファミリー関連リガンドの結合(BiaCore 法)に関する パラメータ Ligand ka (M-1s-1) kd (s-1) K D(pM) Human VEGF-A165 4.05x107 2.01x10-5 0.497 Human VEGF-A121 3.75x107 1.35x10-5 0.360 Human PlGF-2 1.75x106 6.81x10-5 38.8 Human PlGF-1 6.73 x106 2.64x10-3 392.0 Murine VEGF-A164 2.80x107 1.64x10-5 0.585 Murine VEGF-A120 2.15x107 1.23 x10-5 0.571 Murine PlGF-2 1.64x107 5.45x10-5 3.33 Rat VEGF-A164 3.67x107 1.73x10-5 0.471 Rabbit VEGF -A165 3.39x107 2.63 x10-5 0.775 Human VEGF-B(10-108) 3.52 x107 6.74 x10-5 1.92 Human VEGF-C NB1. NB NB Human VEGF-D NB NB NB
Abbreviations used: ka = Association rate constant, kd = Dissociation rate constant, KD= Equilibrium
dissociation constant.
1.NB= no detectable binding
2.6.2.2.1.2 HUVEC における VEGF-A165依存性受容体リン酸化の阻害作用
参照項目: 4.2.1.1-3 VGFT-MX-08016 ヒト臍帯静脈内皮細胞(HUVEC)は VEGF 受容体を発現していることから、外因性 VEGF の添加
により in vitro にて培養し増殖を誘導することができる。HUVEC を用いて VEGF Trap の
VEGF-A165依存性 VEGFR-2 リン酸化阻害能を評価した。
ヒト VEGF-A165(1nM)を様々な濃度の VEGF Trap(0.5-3.0nM)とインキュベートした後、 HUVEC に添加して VEGFR-2 のチロシンリン酸化を誘導した。
HUVEC に添加してインキュベートした後、HUVEC を溶解し、抗 VEGFR-2 抗体を用いて免疫沈降 した。得られた抗体-VEGFR-2 複合体を用いて SDS ゲル電気泳動を実施した。電気泳動後、たん 白を PVDF 膜に転写し、マウスの抗リン酸化チロシンモノクローナル抗体を添加した。次に HRP 標識したヤギの抗マウス抗体とインキュベートした後、リン酸化 VEGFR2 を ECL(enhanced chemiluminescence)検出システムにより可視化した。
VEGF Trap の濃度が VEGF-A165濃度に対し化学量論比以下(モル比 1:1 以下)では、遊離型 (非結合型)VEGF-A165により受容体リン酸化が誘導されるが、モル比が 1:1 以上では、受容体 リン酸化の阻害が認められた(2.6.3.2 参照)。
2.6.2.2.1.3 VEGF 誘発カルシウム動員の阻害作用
参照項目:4.2.1.1-3 VGFT-MX-08016 HUVEC を用いて、ヒト VEGF-A165と VEGFR-2 の結合及びその後のカルシウム動員の活性化に対す る VEGF Trap の抑制能について検討した。独立した 3 試験において、50pM の VEGF-A165により誘 導されるカルシウム動員に対する VEGF Trap の抑制能について検討した。
Fluo-4(カルシウムインジケーター)で標識した HUVEC に、VEGF Trap(54.9pM-40nM)を添加 し、さらに VEGF165(50pM)を添加しカルシウム動員を刺激した。細胞における蛍光強度変化は FlexStation(Molecular Devices 社)を用いて発光波長 525nm で記録し、データは各種濃度の VEGF Trap の平均ピーク蛍光強度を用いて解析した。 VEGF Trap はカルシウム動員を効果的に抑制し、細胞内カルシウム動員を 50%抑制する濃度 (IC50)は 1.2~1.7nM であった(2.6.3.2 参照)。 2.6.2.2.1.4 VEGF Trap の補体依存性細胞傷害及び抗体依存性細胞傷害作用 参照項目: 4.2.1.1-4 VGFT-MX-07014 VEGF Trap の作用における Fc 介在性エフェクター機能の役割を検討するために、数種の細胞 アッセイを用い、補体依存性細胞傷害(CDC)作用及び抗体依存性細胞傷害(ADCC)作用を検討 した。
ADCC:初代培養 HUVEC 及び腫瘍細胞株(肺癌 Calu6、大腸癌 DLD1、線維肉腫 HT1080、類表 皮癌 A431 細胞株)と様々な濃度の VEGF Trap(最終濃度 0.85pM~50nM)[±ヒト VEGF-A165 (10nM)]をインキュベートした。さらに、エフェクター細胞としてヒト末梢血リンパ球を添加 し 、 イ ン キ ュ ベ ー ト し た 。 細 胞 溶 解 性 は 、 培 養 液 中 に 放 出 さ れ た LDH ( lactate dehydrogenase)を測定することにより評価した。
CDC:様々な濃度に VEGF Trap を希釈し(最終濃度 0.85pM~50nM)、ADCC アッセイと同様の標 的細胞に添加した[±ヒト VEGF-A165(10nM)]。さらに、補体を含むヒト正常血清を添加し、イ ンキュベートした。細胞の生存を alamarBlue 蛍光色素により検出した。
VEGF Trap(0.85pM~50nM)[±VEGF-A165(10nM)]は、初代培養 HUVEC 及び腫瘍細胞株(線維 肉腫 HT1080、肺癌 Calu-6、大腸癌 DLD-1、類表皮癌 A431 細胞株)のいずれにおいても ADCC 作用 を示さなかった。同様に、VEGF Trap(0.85pM~50nM)[±VEGF-A165(10nM)]は、初代培養 HUVEC 及び腫瘍細胞株(線維肉腫 HT1080、肺癌 Calu-6、大腸癌 DLD-1、類表皮癌 A431 細胞株)の いずれにおいても、本アッセイの陽性対照であるリツキシマブ(抗 CD20 IgG1 抗体)が、CD20 抗原を発現するバーキットリンパ腫由来 Daudi 細胞に殺細胞作用を示す条件下で、CDC 作用を誘 発しなかった(2.6.3.2 参照)。
以上の結果より、VEGF Trap は in vivo において CDC 作用及び ADCC 作用のいずれも発揮しな いと考えられることから、in vivoにおける VEGF Trap の作用は、VEGF Trap が VEGF 又はその他 の VEGFR リガンドに結合し、リガンドの受容体への結合を妨げることにより発揮されていると考 えられる。
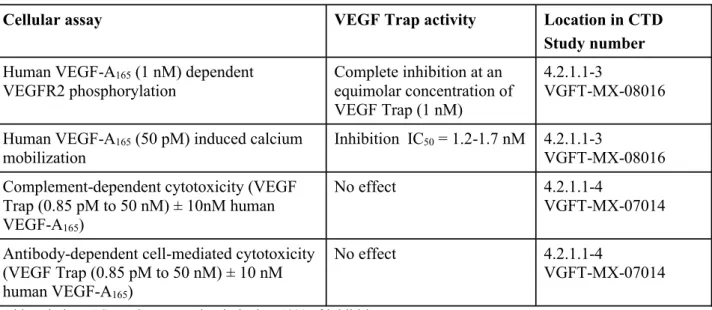
表 2.6.2- 2 In vitroにおける VEGF Trap の作用
Cellular assay VEGF Trap activity Location in CTD
Study number Human VEGF-A165(1 nM) dependent
VEGFR2 phosphorylation Complete inhibition at an equimolar concentration of VEGF Trap (1 nM) 4.2.1.1-3 VGFT-MX-08016
Human VEGF-A165(50 pM) induced calcium
mobilization
Inhibition IC50= 1.2-1.7 nM 4.2.1.1-3
VGFT-MX-08016 Complement-dependent cytotoxicity (VEGF
Trap (0.85 pM to 50 nM) ± 10nM human VEGF-A165)
No effect 4.2.1.1-4
VGFT-MX-07014
Antibody-dependent cell-mediated cytotoxicity (VEGF Trap (0.85 pM to 50 nM) ± 10 nM human VEGF-A165)
No effect 4.2.1.1-4
VGFT-MX-07014
Abbreviation: IC50= Concentration inducing 50% of inhibition.
2.6.2.2.2 In vivo における作用 非臨床試験に加えて、臨床試験や疫学的データからも VEGF Trap-Eye が滲出型(新生血管)加 齢黄斑変性(AMD)及び糖尿病網膜症やその他の虚血性網膜症などの眼性血管疾患の治療薬にな り得ることを支持する根拠が示されている。このような眼性血管疾患では 2 つの共通する特徴、 すなわち、血管漏出に伴って二次的に生じる網膜浮腫と異常血管の増殖がみられる。 異常な血管透過性亢進及び血管新生に対する VEGF Trap の抑制効果を様々な眼性血管疾患動物 モデルを用いて検討した。VEGF Trap は、遺伝子導入による VEGF の過剰発現マウス、又は外因 性 VEGF 硝子体内投与マウスにおける網膜の血管漏出を抑制し、糖尿病モデルラットにおいて網 膜血管透過性を正常化することが確認された。また、VEGF Trap はげっ歯類及びサルの脈絡膜新 生血管(CNV)モデル、マウス酸素誘発虚血性網膜症モデル及び縫合糸誘発角膜血管新生モデル で血管新生を抑制した。特に、サルのモデルでは、レーザー傷害の 15 日後に VEGF Trap を単回 硝子体内(IVT)投与した結果、活動性 CNV 病変における血管漏出が速やかに回復した。これら すべての試験において、VEGF Trap の全身あるいは硝子体内投与により、血管透過性及び血管新 生が効果的に抑制され、評価部位に付随する炎症も著明に改善した。 2.6.2.2.2.1 マウスにおける脈絡膜新生血管及び VEGF 誘発血液網膜関門破綻に対する VEGF Trap の抑制作用 参照項目:[参考]4.2.1.1-5 J Cell Physiol 2003-195-241 動物は AMD そのものは発症しないが、レーザー照射によりブルッフ膜とその上に重なる網膜色 素上皮に局所的な損傷を起こすことによって、滲出型 AMD でみられるような脈絡膜新生血管病変 を形成させることができる。この損傷は、網膜色素上皮層と網膜下腔へ向かう脈絡毛細血管の異 常増殖を刺激する。ブルッフ膜の損傷は、滲出型 AMD を含むすべての CNV の病態に共通してみら れる特徴である。
レーザー傷害誘発 CNV マウスモデルにおいて、C57BL/6 マウス(成熟)の片眼に VEGF Trap (4.92μg)を、もう一方の眼に対照のたん白質としてヒト Fc(hFc、4.92μg)をレーザー傷害 後直ちに単回硝子体内投与した。投与後 14 日目にマウスを麻酔し、フルオレセイン標識デキス トランで灌流した後、両眼を摘出して組織学的な評価を行った。CNV 病変の大きさは脈絡膜フ ラットマウントの画像解析により測定した。 VEGF Trap を硝子体内投与したマウスでは、hFc を硝子体内投与したマウスと比較して CNV 病 変面積が小さかった(図 2.6.2- 1)。高用量の VEGF Trap(25mg/kg)を皮下投与した試験にお いても、hFc を皮下投与した対照群と比較して、レーザー傷害マウスモデルの CNV 病変の面積は 小さかった(図 2.6.2- 2、2.6.3.2 参照)。 図 2.6.2- 1 レーザー傷害誘発 CNV マウスモデルにおける VEGF Trap 単回硝子体内投与後の CNV 病変に対する抑制作用 C57BL/6 マウス(成熟)の片眼に VEGF Trap(4.92μg)を、もう一方の眼に hFc(4.92μg)をレーザー照射後直 ちに硝子体内投与した。投与 14 日目に、フルオレセイン標識デキストランで灌流した後、CNV 面積を測定し た。VEGF Trap を投与した眼(54 傷害部位/19 眼)では、hFc を投与した眼(44 傷害部位/19 眼)と比較し て、画像解析により測定した CNV 面積が有意に小さかった(P<0.0001、Student’s two-tailed t-test)。
図 2.6.2- 2 レーザー傷害誘発 CNV マウスモデルにおける VEGF Trap 皮下投与後の CNV 病変に 対する抑制作用
C57BL/6 マウス(成熟)に、レーザー照射により片眼当たり 3 ヵ所にブルッフ膜の損傷を誘発した。レーザー 照射前、レーザー照射後 2、5、8 及び 11 日目に、マウスに 25mg/kg の VEGF Trap 又は hFc を皮下投与した。 VEGF Trap を投与したマウス(52 傷害部位/20 眼)では、hFc を投与したマウス(57 傷害部位/20 眼)と比 較して、画像解析により測定した CNV の平均面積が有意に小さかった(P<0.0001、Student’s two-tailed t-test)。
VEGF 誘発血液網膜関門破綻に対する VEGF Trap の抑制効果を評価するため、2 種類のマウスモ デルを用いて追加試験を実施した。最初の試験では、マウスに VEGF Trap(25mg/kg)又は hFc を皮下投与し、翌日 10-6M の組換えヒト VEGF 1μL を硝子体内投与した。投与後 6 時間に網膜の 血管透過性を[3H]マンニトールをトレーサーとして評価した。VEGF Trap を投与したマウスでは、 対照と比べて網膜対肺の漏出比が有意に減少し、血液網膜関門機能の改善が示された(図 2.6.2- 3、2.6.3.2 参照)。続く試験では、ドキシサイクリンで誘導可能なロドプシンプロ モーターの制御下で、光受容体に導入した VEGF 遺伝子が発現されるよう設計したトランスジェ ニックマウスを用いた(rhoVEGF マウス)。VEGF の過剰発現を誘導するため、rhoVEGF マウスの 飲水にドキシサイクリンを加え、ドキシサイクリン投与の 1 日前及び 2 日後に VEGF Trap 又は対 照のたん白質である hFc を同様に皮下投与した。2 回目の VEGF Trap 投与 1 日後に[3H]マンニ トールをトレーサーとして網膜血管透過性を測定した。本試験においても、同量の hFc を投与し た対照群のマウスと比較して、VEGF Trap 投与マウスでは VEGF 誘発血液網膜関門の破綻が抑制 されることが示された(図 2.6.2- 4、2.6.3.2 参照)。
図 2.6.2- 3 マウスにおける VEGF Trap 皮下投与後の VEGF 誘発血液網膜関門破綻に対する 抑制作用
C57BL/6 マウス(成熟)に、25mg/kg の VEGF Trap 又は hFc を皮下投与し、翌日 1μg(10-6M)の VEGF を硝
子体内投与した。投与後 6 時間に網膜の血管透過性を[3H]マンニトールをトレーサーとして評価した。VEGF
Trap を投与したマウス(9 匹、18 眼)では、hFc を投与したマウス(9 匹、18 眼)と比較して網膜対肺の漏出 比が有意に減少し、血液網膜関門機能の改善が示された。
図 2.6.2- 4 VEGF 過剰発現マウスにおける VEGF Trap 皮下投与後の VEGF 誘発血液網膜関門破 綻に対する抑制作用
ドキシサイクリンで網膜での VEGF の発現を誘発したトランスジェニックマウスに、25mg/kg の VEGF Trap 又 は hFc をドキシサイクリン投与の 1 日前及び 2 日後に皮下投与し、[3H]マンニトールを指標に血管透過性を測
定した。VEGF Trap を投与したマウス(10 匹、20 眼)では、hFc を投与したマウス(10 匹、20 眼)と比較し て網膜対肺の漏出比が有意に減少した。
2.6.2.2.2.2 網膜下マトリゲルモデルラットにおける VEGF Trap による CNV の形成抑制、 CNV の退行、並びに CNV に付随する炎症及び線維化の抑制
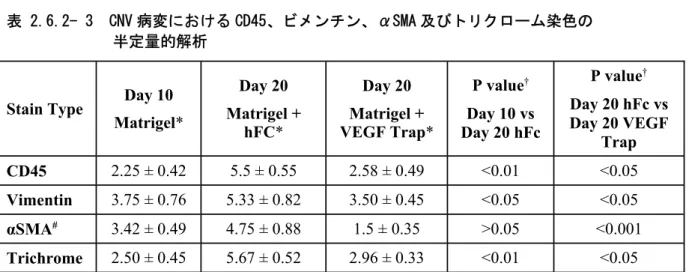
参照項目:[参考]4.2.1.1-6 Invest Ophthalmol Vis Sci 2010-51-6009 Sprague-Dawley 系成熟ラットに 33G 針を用いて 75%のマトリゲル溶液を網膜下に投与し、CNV を誘発した。本モデルでは、マトリゲル投与後 4 日目に新生血管の伸展がみられ、10 日目まで にはマトリゲル投与領域に脈絡膜毛細血管から伸長した血管網が広範に形成される。VEGF Trap が本モデルの CNV 形成を抑制するかどうかを検討するために、VEGF Trap(12.5mg/kg)をマトリ ゲル投与後 2 日目及び 6 日目に皮下投与した。対照群の動物には、ほぼ等モルの hFc(分子量 58kDa)6.25mg/kg を対照たん白質として皮下投与した。10 日目に動物を安楽死させ、蛍光色素 (DiI)を投与して血管を視覚化し、CNV 範囲を定量した。 対照群ではすべての眼に高度の CNV 形成が認められた(CNV の面積:211.35±146.00 × 103μ m2、平均値±標準偏差、n=12)が、VEGF Trap 投与群では CNV 形成は全く認められなかった (CNV の面積:0±0 × 103μm2、n=12)(2.6.3.2 参照)。 また、マトリゲル投与後 10 日間 CNV を形成させ、一方の群の眼を摘出し治療前の CNV のベー スラインとした。残りの動物に VEGF Trap(12.5mg/kg)又は等モル濃度の hFc(6.25mg/kg)を 10、13 及び 16 日目に皮下投与した。マトリゲル投与後 20 日目に眼を摘出し、CNV を評価した。 マトリゲル投与後 10 日目ですべての眼に CNV が認められ、20 日目では hFc 投与群の CNV の形 成範囲は拡大した。一方、VEGF Trap 投与群の CNV の面積(37.10±45.87 × 103μm2、n=14) は、10 日目の治療前対照群の面積(244.50±225.21 × 103μm2、n=16)の 15%にとどまり、14 例中 6 例の眼には CNV が全く認められなかった。以上の結果から、VEGF Trap は 10 日目から 20 日目の間に新たに生じる CNV を抑制するのみでなく、既に形成された CNV 病変も退行させること が示された。同じ処置を施した動物から作製した眼の切片をヘマトキシリン・エオジン染色した 結果、20 日目の網膜下病変部位の総容積(血管及び血管以外のすべての構成要素)は、hFc 投与 群(7.5±0.9 × 108μm3、n=8)と比較して、VEGF Trap 投与群(2.0±0.2 × 108μm3、n= 10)で明らかに小さく、ヘマトキシリン・エオジン染色された細胞含有量も少なかった。一連の 切片におけるビメンチン、平滑筋アクチン及び CD45 の免疫染色でも、hFc 投与群と比較し、 VEGF Trap 投与群の病変部位では、筋線維芽細胞及び白血球は少なく、また網膜下病変のコラー ゲン沈着も著明に減少していることがマッソントリクローム染色により確認された(表 2.6.2-3)。これらの結果は、VEGF Trap がマトリゲルの網膜下沈着による CNV 形成を抑制するのみで なく、異所性の新生脈絡膜血管の退行も誘導することを示している。さらに、VEGF Trap は筋線 維芽細胞及び炎症性細胞の網膜下病変部位への浸潤も抑制した(2.6.3.2 参照)。
表 2.6.2- 3 CNV 病変における CD45、ビメンチン、αSMA 及びトリクローム染色の 半定量的解析
Stain Type Day 10
Matrigel* Day 20 Matrigel + hFC* Day 20 Matrigel + VEGF Trap* P value† Day 10 vs Day 20 hFc P value† Day 20 hFc vs Day 20 VEGF Trap CD45 2.25 ± 0.42 5.5 ± 0.55 2.58 ± 0.49 <0.01 <0.05 Vimentin 3.75 ± 0.76 5.33 ± 0.82 3.50 ± 0.45 <0.05 <0.05 αSMA# 3.42 ± 0.49 4.75 ± 0.88 1.5 ± 0.35 >0.05 <0.001 Trichrome 2.50 ± 0.45 5.67 ± 0.52 2.96 ± 0.33 <0.01 <0.05 半定量的解析:各染色の範囲及び強度について、独立した 2 名の観察者がスコア(代表的な切片において、ス コア 0 を「染色なし」、スコア 6 を「最も広範囲で濃い染色」とみなした、スケール 0-6 のスコア)により盲検 下で評価した。 * Mean ± SD (n=6)
†Kruskal-Wallis test and Dunn test. #αSMA = α smooth muscle actin
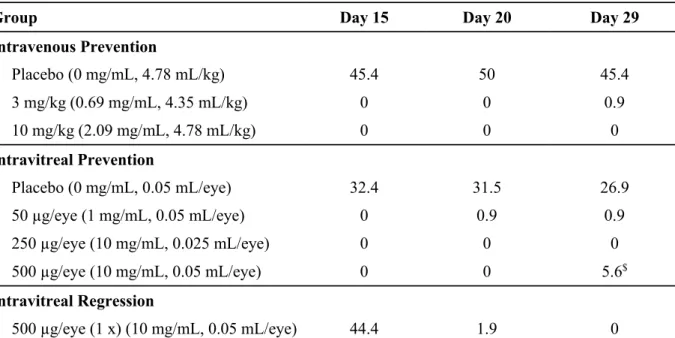
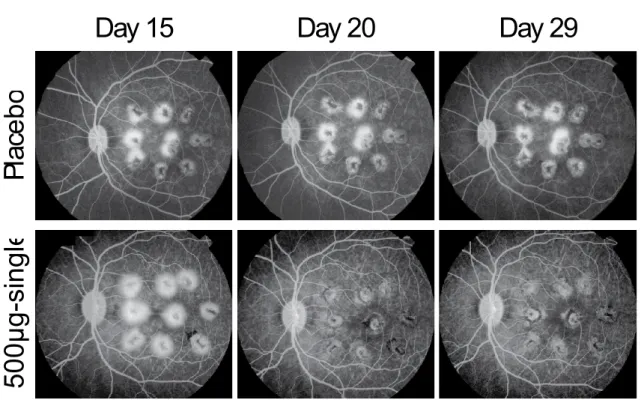
2.6.2.2.2.3 サルにおける VEGF Trap による実験的 CNV の形成抑制並びに既存の活動性 病変の血管漏出の速やかな抑制 参照項目:4.2.1.1-7 VGFT-TX-03027 成熟カニクイザルを用いて、レーザー誘発 CNV に対する VEGF Trap 投与の効果について検討し た。サルの網膜に細く強力なレーザーを 9 ヵ所照射し、形成される活動性 CNV 病変をフルオレセ イン蛍光眼底造影(FA)により、照射前 1 回、並びに照射後 15、20 及び 29 日目に評価した。予 防効果試験では、VEGF Trap をレーザー傷害の 1 週間前から静脈内投与(3 又は 10mg/kg、週 1 回)又は硝子体内投与(50、250 又は 500μg、2 週間に 1 回)した。治療効果試験では、活動性 CNV が既に形成されているレーザー傷害後 2 週間目に、VEGF Trap を単回硝子体内投与(500μ g)した。対照群のサルにはプラセボをレーザー傷害の 1 週間前から週 1 回静脈内投与又は 2 週 間に 1 回硝子体内投与した。各群には、それぞれ 6 匹(雄 3 匹/雌 3 匹)の動物を割り当てた。 CNV 病変を FA により視覚化し、盲検下で評価者によりグレード 1(過蛍光なし)からグレード 4 (レーザー照射部位の境界を越えて広がる後期漏出と鮮明な過蛍光)に分類した。 プラセボを硝子体内又は静脈内投与した群では、レーザー傷害後 15 日目に活動性 CNV 病変 (グレード 4)がレーザー傷害部位のそれぞれ 32.4%及び 45.4%に認められた(表 2.6.2- 4)。 一方、VEGF Trap を静脈内又は硝子体内投与した群では、グレード 4 の病変は全くあるいはほと んど形成されなかった。さらに、活動性 CNV 病変形成後に VEGF Trap(500μg)を単回硝子体内 投与すると、投与後 2 週間でグレード 4 の病変の割合が 44%から 0%に低下した(表 2.6.2- 4 及び図 2.6.2- 5)。 VEGF Trap 硝子体内投与の忍容性は良好であり、前眼部及び硝子体に炎症 性細胞は全く認めないか、又は認めたとしても微量(trace~1+)であった(2.6.3.2 参照)。
表 2.6.2- 4 サルのレーザー傷害後 15、20 及び 29 日における各群のグレード 4 の割合
Group Day 15 Day 20 Day 29
Intravenous Prevention Placebo (0 mg/mL, 4.78 mL/kg) 45.4 50 45.4 3 mg/kg (0.69 mg/mL, 4.35 mL/kg) 0 0 0.9 10 mg/kg (2.09 mg/mL, 4.78 mL/kg) 0 0 0 Intravitreal Prevention Placebo (0 mg/mL, 0.05 mL/eye) 32.4 31.5 26.9 50 µg/eye (1 mg/mL, 0.05 mL/eye) 0 0.9 0.9 250 µg/eye (10 mg/mL, 0.025 mL/eye) 0 0 0 500 µg/eye (10 mg/mL, 0.05 mL/eye) 0 0 5.6$ Intravitreal Regression 500 µg/eye (1 x) (10 mg/mL, 0.05 mL/eye) 44.4 1.9 0 予防効果試験では、VEGF Trap をレーザー傷害の 1 週間前から静脈内投与(3 又は 10mg/kg、週 l 回)又は、硝 子体内投与(50、250 又は 500μg/eye、2 週間に 1 回)した。治療効果試験では、VEGF Trap をレーザー傷害の 2 週間後に単回硝子体内投与(500μg)した。両試験の対照群の動物には、レーザー傷害の 1 週間前からプラ セボを週 1 回静脈内投与又は 2 週間に 1 回硝子体内投与した。投与群及び対照群には、それぞれ 6 匹(雄 3 匹 /雌 3 匹)の動物を割り当てた。フルオレセイン蛍光眼底造影(FA)を用いて CNV 病変を視覚化し、盲検下 で評価者によりグレード 1(過蛍光なし)からグレード 4(レーザー照射部位の境界を越えて広がる後期漏出 と鮮明な過蛍光)に分類した。上表は、評価の各時点(レーザー傷害後 15、20 及び 29 日目)における臨床的 に重度(グレード 4)と判断された投与群及び対照群の病変の割合(平均値)を示す。 $この増加は投与群 1 匹の両眼のみにみられたグレード 4 の病変に起因した。なお、当該動物でみられた異常 反応の原因は不明であった。
Day 15
Day 20
Day 29
P
la
cebo
500µg-single
図 2.6.2- 5 形成されたグレード 4 病変に対する VEGF Trap 硝子体内投与の治療効果IVT 治療効果試験:プラセボ(反復)又は VEGF Trap(単回 500μg)投与群におけるレーザー傷害 15, 20 及び 29 日後の後期フルオレセイン蛍光眼底造影画像を示す。プラセボ群(上段)の動物では、いずれの観察時点に おいても 9 つの傷害処置部位のほとんどでグレード 4 の血管漏出が認められた。VEGF Trap 群(下段)では、 VEGF Trap 投与前である傷害 15 日後では 9 つの傷害部位すべてでグレード 4 の漏出がみられたが、20 日後 (VEGF Trap 投与 5 日後)ではグレード 4 は認められなかった。傷害 29 日後でも血管漏出の再発はみられな かった。 サルにおいて、硝子体内投与後の血漿中遊離型 VEGF Trap 濃度は、静脈内投与後に比べ明らか に低かった(2.6.6.2.2.1 表 2.6.6- 3 参照)。50μg/eye を硝子体内投与後の血漿中遊離型 VEGF Trap 濃度は、定量下限未満であった。250 及び 500μg/eye 硝子体内投与後の血漿中濃度は 用量依存的であり、硝子体内投与後 24 時間で最大濃度に達し、次回投与前には検出限界付近又 はそれ以下に低下した。一方、3 及び 10mg/kg 静脈内投与後の血漿中遊離型 VEGF Trap 濃度は、 すべての測定時点において 500μg/eye 投与後よりも高かった。(2.6.6.2.2.1 表 2.6.6- 3 参 照) 2.6.2.2.2.4 VEGF Trap 投与による糖尿病ラットにおける網膜血管漏出の改善 参照項目: 4.2.1.1-8 VGT-NC-007 VEGF の過度の発現が、糖尿病における網膜血管新生と黄斑浮腫の発症に中心的な役割を果た すと考えられている。糖尿病の網膜内で起こり得る変化を検討するため、2 種の動物モデル、す なわち糖尿病(ストレプトゾトシン処置)ラットにおける網膜浮腫、並びにマウス虚血誘発網膜 血管新生(2.6.2.2.2.5 参照)に対する VEGF Trap の効果を評価した。 Sprague Dawley 系雄性成熟ラットに、10mM クエン酸緩衝液(pH4.5)に溶解したストレプトゾ トシン(60mg/kg)を単回腹腔内投与することにより糖尿病を誘発した。24 時間後に高血糖が認 められない動物は試験から除外した。ストレプトゾトシン処置 1 週間後に血糖値を再測定し、糖
尿病の状態を確認した。糖尿病の発症が確認された動物の片眼に、VEGF Trap(3μg/3μL)を単 回硝子体内投与し、もう一方の眼に対照たん白質の hFc を投与した。投与後 48 時間に、エバン スブルー色素(45mg/kg)を静脈内投与し、2 時間後に網膜の色素濃度を測定して網膜血管透過 性を評価した。 網膜におけるエバンスブルー色素の平均濃度は、非糖尿病ラットに比較して、糖尿病ラットで 約 3 倍上昇した(図 2.6.2- 6)。一方、VEGF Trap を硝子体内投与した糖尿病ラットでは、hFc を硝子体内投与した糖尿病ラットに比べてエバンスブルー色素濃度が有意に減少し、非糖尿病 ラットと同程度まで低下した。VEGF Trap 硝子体内投与 48 時間後の血中に遊離型 VEGF Trap は 検出されなかった。また、VEGF Trap 硝子体内投与 1 週間後の血管漏出の評価においても、網膜 血管透過性の正常化が認められた(2.6.3.2 参照)。 図 2.6.2- 6 VEGF Trap 硝子体内投与によるストレプトゾトシン誘発糖尿病ラットの 網膜におけるエバンスブルー色素漏出の抑制 雄性成熟ラットにストレプトゾトシンを投与し糖尿病を誘発した。非糖尿病群のラットにはストレプトゾトシ ンの代わりにクエン酸緩衝液を投与した。糖尿病誘発1週間後、片眼に VEGF Trap(3μg/3μL)を単回硝子 体内投与し、もう一方の眼に対照のたん白質 hFc(3μg/3μL)を投与した。48 時間後にエバンスブルー色素 の血管外漏出を指標として網膜血管透過性を評価した。VEGF Trap は、糖尿病モデルにおける網膜血管透過性 亢進を有意に改善した(unpaired t-test)。図中の値は平均値±標準誤差。 2.6.2.2.2.5 VEGF Trap による網膜虚血マウスにおける病的な血管新生の抑制 参照項目: 4.2.1.1-9 VGT-NC-013 VEGF Trap の抗血管新生作用を、酸素誘発虚血性網膜症(OIR)のマウスモデルを用いて検討 した。OIR は、幼若マウスを生後 7 日目から高酸素環境に一時的に曝露することで生じさせた (高酸素症)。高酸素環境下では、中心網膜内の微小血管系の発達が低下する。幼若マウスを生
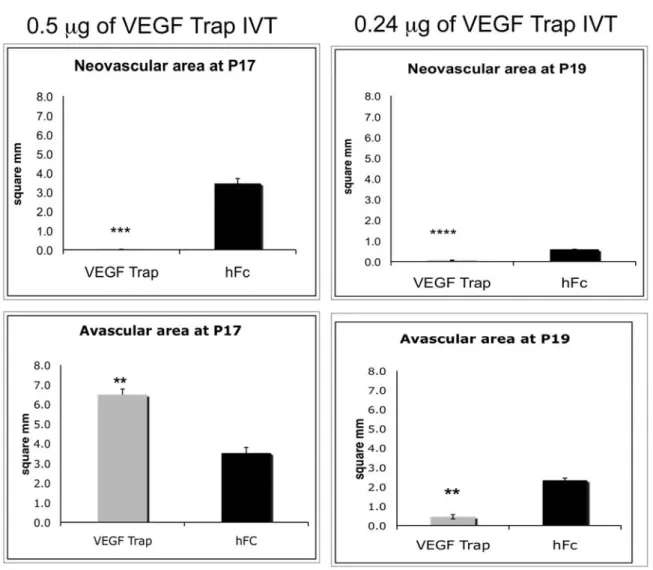
後 12 日目に高酸素環境下から室内の通常酸素環境下に戻すと、網膜が低酸素状態となり、増殖 糖尿病網膜症や未熟児網膜症などの虚血性網膜症と同様に血管新生が刺激される。 生後 14 日目(室内酸素に戻した 2 日後)に VEGF Trap(0.5μg、5mg/mL、100nL/eye)又は同 量の対照たん白質(hFc)を幼若マウスに単回硝子体内投与し、3 日後(生後 17 日目)に網膜を 採取した。高酸素に曝露した対照群のマウスの網膜では、内境界膜を貫通する血管のふさ状分岐 と、特に視神経乳頭周辺の網膜表面における無秩序な血管の伸長を特徴とする病的な新生血管が みられた。これに対して VEGF Trap を投与した群のマウスでは、網膜の病的な新生血管の形成が 抑制されていた(図 2.6.2- 7)。この用量(0.5μg)の VEGF Trap は、中心網膜の無血管領域 における血管再生範囲も減少させた。一方、生後 15 日目に低用量(0.24μg、11.82mg/mL、 20nL/eye)の VEGF Trap を硝子体内投与した群では、病的な網膜血管新生は抑制されたが、正常 に血管再生された中心網膜の範囲は生後 19 日目までに拡大した(図 2.6.2- 7)(2.6.3.2 参 照)。結論として、高用量の VEGF Trap による強力な VEGF の阻害は、病的な血管新生を完全に 抑制するが、正常な網膜血管の再生も若干遅延させた。これに対して、低用量の VEGF Trap は、 正常な網膜血管の再生を遅らせることなく、むしろ促進する一方、病的な血管新生は抑制した。 この結果は、病的に形成される血管は正常の血管よりも VEGF 阻害に対して感受性が高いことを 示唆している。
図 2.6.2- 7 OIR マウスにおける VEGF Trap 硝子体内投与後の網膜の病的な血管新生及び 血管再生
レクチン染色の網膜フラットマウントの画像より、異常な血管を含む網膜の大きさを測定して病的な血管新生 の範囲を定量した(上段)。フラットマウントの画像は、Nikon Eclipse 顕微鏡、Adobe Photoshop 及び Scion Image を使用して取り込んだ。中心網膜の無血管領域も同様の方法で測定した(下段)。VEGF Trap 又は hFc 0.5μg 又は 0.24μg の硝子体内投与の効果をそれぞれ左列及び右列に示す。投与条件盲検下で、評価者がすべ ての画像を撮影及び解析した(n=4、試験ごとに各治療群で検討した網膜数)。データは平均値±標準誤差で 示し、unpaired t-test を用いて解析した(**p<0.01、***p<0.001、****p<0.0001)。P=生後日数。 2.6.2.2.2.6 VEGF Trap による角膜傷害マウスにおける炎症及び血管新生の抑制 参照項目:[参考]4.2.1.1-10 J Clin Invest 2004-113-1040 マウスの角膜実質内に留置した縫合糸により誘発した角膜傷害により、高度の血管新生反応が みられる。本モデルでは、VEGF-C と VEGF-D による VEGFR-3 の活性化を介して生じるリンパ管新 生 が、血 管 新 生 と同時に 生じる 。最初 の試験で は、 Balb/C マ ウ ス(成 熟)に VEGF Trap (12.5mg/kg)又は同量の対照たん白質(hFc)を、縫合糸傷害時に単回腹腔内投与し、7 日後に 角膜を評価した。
その結果、VEGF Trap を投与したマウスでは、hFc 投与マウスとは異なり血管新生のみならず 角膜リンパ管新生も完全に抑制されることが明らかになった4)(血管+リンパ管面積:VEGF Trap 投与群 2.3±1.5%、hFc 投与群 49±12%)(2.6.3.2 参照)。 本モデルでは、血管新生及びリンパ管新生に先行して、血管炎症並びに傷害された角膜への白 血球(主にマクロファージと好中球)の遊走が生じる。VEGF-A や関連リガンドの PlGF は、白血 球の細胞表面に存在する VEGFR-1 を介して、白血球の強力な化学走化性因子として機能すること が知られている。活性化した白血球は、VEGF-C 及び VEGF-D を発現する。したがって、局所的に 発現が上昇した VEGF-A や PlGF による白血球の傷害部位への遊走を VEGF Trap が抑制することに より、VEGF Trap は間接的にリンパ管新生を抑制する可能性が考えられた。これを確かめるため に、角膜傷害後の白血球浸潤に対する VEGF Trap の阻害能を評価する試験を再度実施した。 VEGF Trap 投与群(188±14)では、対照の hFc 投与群(909±167)と比較して角膜切片中の 炎症性細胞数が減少した。さらに、角膜傷害前の放射線照射による全骨髄由来細胞の全身的な枯 渇[血管面積:放射線照射群 18.4±4%、対照(非照射)群 49.6±10.4%、リンパ管面積:放射 線照射群 16.4±3.2%、対照群(非照射)38±12.23%]、あるいはリポソーム封入クロドロ ネートによるマクロファージの局所的な枯渇[血管面積:リポソーム封入クロドロネート群 11.3±5.8%、対照(リン酸緩衝液)群 42.3±11.3%、リンパ管面積:リポソーム封入クロドロ ネート群 10.8±2.5%、対照(リン酸緩衝液)群 38.8±4.7%]により、血管新生及びリンパ管 新生が有意に抑制されることも示された。これらの結果から、VEGF-A 及び PlGF は、血管新生や リンパ管新生に必要なシグナルを供給及び増幅させる白血球/マクロファージの動員を介して、 角膜傷害の病態生理学的反応に重要な役割を果たしていると考えられる。なお、最近の報告によ ると、VEGF Trap は bFGF による角膜の血管新生を抑制し17)、またマウス角膜移植後の VEGF Trap 投与により、血管新生、リンパ管新生及び炎症を抑制するのみならず、同種移植角膜の生着も改 善することが明らかにされている7,18,19)。
2.6.2.3 副次的薬理試験
2.6.2.3.1 正常及び担癌マウスにおける遊離型 VEGF Trap 及び VEGF Trap 複合体の濃度、
並びに抗腫瘍効果との相関性
参照項目:[参考]4.2.1.2-1 Proc Natl Acad Sci USA 2007-104-18363 乾癬6)、角膜移植拒絶反応7)及び腫瘍異種移植8)などの動物モデルを用いた非臨床試験におい て、VEGF Trap 全身投与による病的な血管新生の治療及び他の生物学的作用について評価した。 腫瘍の同種又は異種移植モデルでは、用量反応性試験を実施し、VEGF を完全に中和するのに必 要な VEGF Trap の用量及び血中濃度を検討した。それらの試験の一つでは、担癌マウスと正常マ ウスの両方で、遊離型 VEGF Trap 血中濃度と VEGF に結合した VEGF Trap(VEGF Trap 複合体)の 濃度を比較し、腫瘍増殖に対する効果との相関性を検討した(2.6.3.3 参照)。
B16F1 マウス黒色腫細胞株、A673 ヒト横紋筋肉腫細胞株又は MMT マウス乳癌細胞株を皮下に移 植した SCID マウスに、VEGF Trap 0、0.5、1、2.5、10 及び 25mg/kg を週 2 回皮下投与(1~2 週 間)し、血漿中遊離型 VEGF Trap 及び VEGF Trap 複合体の濃度を ELISA 法で測定した。
その結果、腫瘍の種類によらず遊離型 VEGF Trap 濃度は、検討した用量範囲において用量に応 じて上昇し、2.5mg/kg でおよそ 10μg/mL のレベルに、25mg/kg ではおよそ 100μg/mL に達した。 一方、VEGF Trap 複合体濃度は低用量域で漸増し、図 2.6.2- 8に示すとおり、2.5mg/kg 以上の 用量で最大濃度である 1~2μg/mL に達した。一方、VEGF Trap は 2.5mg/kg 以上の用量で腫瘍の 増殖を抑制した。すなわち、抗腫瘍効果のみられる用量では、遊離型 VEGF Trap 濃度は VEGF Trap 複合体の最大血漿中濃度を超えていた。
図 2.6.2- 8 VEGF Trap の用量と遊離型 VEGF Trap 及び VEGF Trap 複合体の血漿中濃度の関連 並びに腫瘍増殖に対する効果との相関
マウスの皮下(脇腹)に B16F1 マウス黒色腫細胞株(A)、A673 ヒト横紋筋肉腫細胞株(B)又は MMT マウ ス乳癌細胞株(C)を移植し、腫瘍がおよそ 100 mm3に増殖した後、VEGF Trap 0、0.5、1、2.5、10 及び
25mg/kg を週 2 回 1~2 週間皮下投与した。血漿中の遊離型 VEGF Trap(Free Trap、図中青線)と VEGF Trap 複 合体(VEGF Trap Complex、図中緑線)を測定した。腫瘍の種類に関係なく遊離型 VEGF Trap レベルは検討範 囲内の用量で、用量に応じて増加し、2.5mg/kg で 10μg/mL に達し、25mg/kg でおよそ 100μg/mL に上昇した。 一方、VEGF Trap 複合体は 2.5mg/kg 以上の用量で最大濃度 1~2μg/ml に達した。腫瘍容積(図中赤線)は、 2.5mg/kg 以上の用量では一貫して減少し、この用量(2.5mg/kg)は、遊離型 VEGF Trap の血漿中濃度が VEGF Trap 複合体濃度を超える最低用量である(n=6/群)。
図 2.6.2- 9 VEGF Trap の用量と遊離型 VEGF Trap 及び VEGF Trap 複合体の血漿中濃度の関連 マウスに移植した腫瘍の重量が体重の 3%未満である場合、腫瘍から産生される VEGF はマウスの内因性の VEGF と比較して少量であることが示唆される。(A 及び B)マウス(A)又はヒト(B)腫瘍がおよそ 100 mm3に増殖した後、VEGF Trap 0.5、1、2.5、10 及び 25mg/kg を週 2 回 1~2 週間投与した。血中の遊離型
VEGF Trap と VEGF Trap 複合体を試験終了時に測定した。マウス由来腫瘍の移植モデルでは、いずれの場合も 最終腫瘍容積によらず、血中の VEGF Trap 複合体濃度はおよそ 1μg/mL であった。一方、ヒト由来腫瘍の移植 モデルでは VEGF Trap 複合体濃度はおよそ 0.1μg/mL であった。遊離型 VEGF Trap 濃度は徐々に増加し、 2.5mg/kg の用量で VEGF Trap 複合体濃度を超え、25mg/kg でおよそ 100μg/mL に達した(n=6/用量)。 (C)マウス及びヒトの腫瘍型を示す。
マウス(Lewis 肺癌、B16F10 黒色腫)又はヒト(SK-NEP Wilm 腫瘍及び HT1080 線維芽肉腫) 由来の腫瘍を移植したマウスでも同様の結果が得られた。VEGF Trap 複合体濃度は VEGF Trap の 投与量の増加に伴い上昇し、2.5mg/kg 以上で最大濃度の約 1~2μg/mL に達した。この用量レベ ルでは、遊離型 VEGF Trap の血中濃度は VEGF Trap 複合体濃度よりも高かった(図 2.6.2- 9)。
VEGF Trap を同投与スケジュールで正常なマウスに投与したときの遊離型及び結合型の VEGF Trap 濃度は担癌マウスでの濃度と同等であった。したがって、予想以上に多くの内因性かつ生 体内利用可能な VEGF が通常でも正常組織により産生されており(約 0.006μg/g 組織/日)、腫 瘍異種移植由来の VEGF は比較的少量にとどまることが示唆された。
2.6.2.4 安全性薬理試験
ICH ガイドライン S7A「安全性薬理試験ガイドライン」(平成 13 年 6 月 21 日付 医薬審発第 902 号)で定義されている in vitro 安全性薬理試験により融合たん白質などの高分子量化合物 を評価する必要性は低いと考えられる。したがって、GLP に準拠した in vivo試験及び GLP 非準
拠のin vivo試験(げっ歯類の血圧に関する 1 試験)により VEGF Trap の安全性薬理評価を行っ
た(2.6.3.4 参照)。 2.6.2.4.1 中枢神経系 参照項目: 4.2.3.2-17 VGFT-TX-02037 4.2.3.2-15 VGFT-TX-05009 VEGF Trap の中枢神経系に及ぼす影響については、毒性試験で得られた成績から一般症状、直 腸温など安全性薬理のエンドポイントを評価した。雌雄カニクイザルに、VEGF Trap 1.5、5、15 及び 30mg/kg を週に 2 回、3 ヵ月間皮下投与した試験において、カニクイザルの一般症状及び直 腸温への影響は VEGF Trap のいずれの群においても認められず、VEGF Trap の投与による中枢神 経系への影響は認められなかった(表 2.6.2.7- 1、2.6.7.7L、4.2.3.2-17 VGFT-TX-02037 参 照)。同様に、雌雄カニクイザルに VEGF Trap 3、10 及び 30 mg/kg を週 1 回、15 週間、その後 27 週まで 1 週おきに静脈内投与した試験(表 2.6.2.7- 2、2.6.7.7J、4.2.3.2-15 VGFT-TX-05009 参照)においても中枢神経系への影響は認められなかった。 2.6.2.4.2 心血管機能 2.6.2.4.2.1 げっ歯類における血圧に及ぼす VEGF Trap の影響: 遊離型 VEGF Trap の血中濃度との相関性 参照項目:[参考]4.2.1.3-9 VGFT-MX-08018 C57BL/6 マウスに 2.5 及び 25mg/kg の VEGF Trap を、Wistar-Kyoto ラットに 0.05、0.15、0.5、 1、2.5、5、10 及び 25mg/kg の VEGF Trap 又は 5mg/kg の hFc を単回皮下投与したときの血圧に 及ぼす影響をテレメトリー法で検討した(非 GLP 試験)。投与前 48 時間以上の測定記録を血圧 と心拍数のベースライン値とし、測定は投与後およそ 3~4 週間まで継続した(2.6.3.4 参照)。 VEGF Trap 2.5 及び 25mg/kg を C57BL/6 マウスに単回皮下投与した結果、収縮期血圧及び拡張 期血圧いずれも統計学的に有意な上昇を示し、最大上昇は投与後 24~48 時間以内に認められた。 2.5 及び 25mg/kg を投与したマウスにおける収縮期血圧の各動物での最大上昇の平均値は、それ ぞれ 14.3±0.7mmHg 及び 17.3±1.1mmHg であった。ベースライン値を上回る収縮期血圧の上昇の 持続期間は 2.5mg/kg 投与群のマウスで約 7 日であったのに対し、25mg/kg 投与群ではおよそ 21 日間持続した。同様の用量依存的な影響は拡張期血圧にも認められたが、最大上昇幅はわずかに 小さかった。血圧上昇の持続時間は血中の遊離型 VEGF Trap の存在と密接に関係しており、血中 の遊離型 VEGF Trap 濃度がおよそ 1μg/mL を下回るまで、収縮期血圧と拡張期血圧は投与前ベー スライン値を上回る上昇を持続した。また、VEGF Trap 投与マウスにおいて一時的かつ用量依存 的な心拍数の減少が認められたが、その作用は統計学的に有意ではなかった(図 2.6.2.7- 1A、 図 2.6.2.7- 1B 参照)。