Instructions for use
Title
活性酸素によるグリオキサールの生成とその変異誘発能に関する研究Author(s)
紙谷, 尚子Citation
北海道大学. 博士(薬学) 乙第5217号Issue Date
1997-09-30DOI
10.11501/3130611Doc URL
http://hdl.handle.net/2115/32599Type
theses (doctoral)File Information
3963.pdfHokkaido University Collection of Scholarly and Academic Papers : HUSCAP
活性酸素によるグリオキサールの生成と その変異誘発能に関する研究
紙 谷 尚 子
A 3'‑AMP 5
にAMP BPB C
5‑CHO‑dU 5
にCMP c y c l o ‑ dA c y c l o
刷dG dA
5 ' ‑ dA MP dC
5'‑dCMP ddNTP dG 5'‑dGMP dGTP DMEM DMSO DNA dNMP dNTP dT 5
にdTMP DTT dUTP dUTPase ECD EDTA
略語表
Adenine
Adenosine 3 ' ‑ m o n o p h o s p h a t e Adenosine 5 ' ‑ m o n o p h o s p h a t e Bromophenol b l u e
C y t o s i n e
5
・F o r m y l ‑ 2 ' ‑ d e o
勾 ほr i d i n e C y t i d i n e 5 ' ‑ m o n o p h o s p h a t e 8 , 5
にc y c l o
・2 '‑ d e o x y a d e n o s i n e 8 , 5
にc y c l o ‑ 2 ' ‑ d e o
勾r
思l a n o s l n e 2
にDeoxyadenosine
2
にDeoxyadenosine5 ' ‑ m o n o p h o s p h a t e 2 ' ‑ D e o x y c y t i d i n e
2 ' ‑ D e o x y c y t i d i n e 5 ' ‑ m o n o p h o s p h a t e
2 ' , 3 ' ‑ D i d e o x y r i b o n u c l e o s i d e 5
にt r i p h o s p h a t e 2 ' ‑ D e o x y g u a n o s i n e
2 ' ‑ D e o x y g u a n o s i n e 5
にmonophosphate
z
幽Deoxyguanosine5 ' ‑ t r i p h o s p h a t e D u l b e c c o ' s m o d i f i e d E a g l e medium Dimethyl s u l f o x i d e
D e o x y r i b o n u c l e i c a c i d
2
にDeoxyribon u c l e o s i d e 5 ' ‑monophosphate 2 '
幽D e o x y r i b o n u c l e o s i d e5
にt r i p h o s p h a t e 2 ' ‑ D e o x y t h y m i d i n e
2
にDeo
幻r t h y m i d i n e5 ' ‑ m o n o p h o s p h a t e D i t h i o t h r e i t o l
2 ' ‑ D e o x y u r i d i n e 5 ' ‑ t r i p h o s p h a t e 2
三D e o x y u r l
必ne5 ' ‑ t r i p h o s p h a t a s e E l e c t r o c h e m i c a l d e t e c t o r
Ethy l e n e d i a m i n e t e t r a a c e t i c a c i d
FCS
G 3'‑GMP 5'‑GMP HPLC I P I ' G LB M F
MOPS
NADH NMR NTA O
狂2‑0H‑ dA 2
・OH‑ dA TP 8‑0H 欄dA ιOH
胎dC 8‑0H‑dG
OPD
PCR P ‑ g a 1 RNA RNase T T r i s tRNA 5
にUMP UV X ‑ g a 1
F e t a 1 c a l f serum Guanine
Guanosine 3'‑monophosphate Guanosine 5 '
引lOnophosphate
H i g h ‑ p e r f o r m a n c e l i q u i d chromatography I s o p r o p y l ‑ s
・D ‑t h i o g a l a c t o p y r a n o s i d e L u r i a ‑B e r t a n i
Mutation 金 equency
3
・(N‑ morpholino ) p r o p a n e s u l f o n i c a c i d Reduced n i c o t i n a m i d e adenine d i n u c l e o t i d e Nuclear magnetic r e s o n a n c e
N i t r i l o t r i a c e t i c a c i d Hydro
勾r 1
2
・H y d r o x y ‑ 2 ' ‑ d e o x y a d e n o s i n e
2 ‑ H y d r o x y ‑ 2 ' ‑ d e o x y a d e n o s i n e 5 ' ‑ t r i p h o s p h a t e 8
・Hydroxy
齢2 ' ‑ d e o x y a d e n o s i n e
5 ‑ H y d r o x y ‑ 2 ' ‑ d e o x y c y t i d i n e 8
・Hydroxy‑2
乙d ω e o
ほxyg
郡u
泊a n o ω S 1
出ne e
0‑♂ .
PO
札1ymerasec h a i n r e a c t i o n P h e n y 1 ‑ s
・D ‑ g a 1 a c t o p y r a n o s i d e R i b o n u c l e i c a c i d
R i b o n u c l e a s e Thymime
T r i s ( h ydroxymethy 1 ) aminoethane Tr a n s f e r RN A
U r i d i n e 5'‑monophosphate I D t r a v i o 1 e t
5
・Bromo
・4
・c h l o r o
鴫3
・i n d o l y 1
・0
・D ‑ g a 1 a c t o p y r a n o s i d e
目次
序論 1
第 1 章 DNA 及びその関連物質(ヌクレオシド、ヌクレオチド) からのグリオキサール生成
第 1 節 DNA 及びその関連物質(ヌクレオシド、ヌクレオチド)
の Fe 2 + ‑ E D T A 処理 6
第 2 節 OPD 反応によるグリオキサールの検出 7 6
第 2 章種々の活性酸素産生系において生成する修飾ヌクレオシド 13 第 l 節 ヌクレオシドの活性酸素処理 13
第 3 章復帰変異試験を用いたグリオキサールの誘発する変異
スペクトルの検索 28
第 1 節 Ames 試験 E による夕、、リオキサールの変異原性の検出 28
第 4 章大腸菌におけるグリオキサールの変異誘発能 32 第 1 節 変 異 体 選 別 の 原 理 32
第 2節大腸菌のグリオキサール処理 34
第 3 節 変 異 体 の l a c I 遺伝子の配列解析 37
第 5 章 鴎乳動物細胞におけるグリオキサールの変異誘発能 47 第 1 節 変 異 体 選 別 の 原 理 49 第 2 節 プラスミド pMY189 のグリオキサール処理と
COS‑7 細胞へのトランスフェクション 第 3 節 supF 変異体の選別
第 4 節 変 異 体 の supF 遺伝子の配列解析
ー ム の ム
A吐
w h u p n v p
円U
第 6 章 プラスミドを用いた大腸菌におけるグリオキサールの 変異誘発能の解析についての検討
第 1節 グリオキサール処理したプラスミドを大腸菌に トランスフェクションした実験
結 語
謝辞
実験の部
引用文献
63
63
68
69
70
95
l
序論
我々は日常生活において様々な化学物質や放射線にさらされており、その中 には生命体の遺伝情報を司る DNA に作用し、突然変異を誘発するものも少なく ない。 DNA 中に生じる突然変異は、生物の進化という観点からは重要な役割を 担っていると考えられるが、現存の生物にとってはむしろ有害な現象であると 考えられる。突然変異を誘発する要因の中で代表的なものとしては、 ( i )DNA
複製時における誤った塩基対形成、 ( i i )活性酸素、放射線、アルキル化剤、あ るいは種々の変異原性物質による DNA の修飾、あるいは ( i i i ) DNA 合成時に
DNA 鎖に取り込まれる DNA 前駆体の修飾等が知られている o
近年、生体への影響という観点から、活性酸素に対する関心が高まりつつあ る。活性酸素は癌に代表されるような加齢に伴って増加する疾患に関与してい ると考えられている 1 。活性酸素とは一般にスーパーオキシド(一0 2 ) 、過酸化 水素 ( H 2 0 2 )、ヒドロキシルラジカル(・ OH)及び一重項酸素 ( 1 0 2 ) などの酸 素種を指す。広義には脂質過酸化物 (LOOH 、 LOO' )やハロゲン化酸素 (C10
‑)、さらには内皮由来弛緩因子として同定された一酸化窒素ラジカル (NO.)
など生理学的作用の重要性が指摘されるものも含まれる。これらの活性酸素は 好気的条件下での細胞呼吸によって生じるのみならず、ある種の環境変異原 (種々の化学物質や放射線等)によっても生成する。さらに、活性酸素は炎症 時に好中球やマクロファージから放出されることが知られている。
これまでに、活性酸素によって生じる DNA 損傷は数多く報告されている 20
その例を図 1 に示す。また、それらの DNA 損傷の変異誘発能について現在まで に得られている結果を表 1に示した。例えば 5 ・ヒドロキシシトシンは主として C → T トランジッションを誘発し 3
,4 、 8 ‑ ヒドロキシグアニンは主として G → T ト
ランスバージョンを誘発することが報告されているら凡また、 8 ・ヒドロキシア
デニン 1 1
,13
,14 及びデオキシリボース(以下 a b a s i cs i t e の呼称を用いる)は実験
系によって異なった変異を誘発することが知られている 1
5‑2 1 0 また、表中には示
していないが、 2 ‑ ヒドロキシアデニンの誘発する変異についても明らかにされて
おり、 2 ‑ 0 H ‑ dA TP は DNA 中の C 残基に対して誤って取り込まれることにより
2
f':JH
2
~
̲OHr ' i f
0‑.....、N'
5
・hydroxycytosine 凸
Mo n
z n
︑ f f e HN ト
4 N
相
i
側
! 戸 川
ι L y
同
¥
│
│ / は
N J
ー ︑
N
出hH
n o
0 0 i
抑
o k i
耐
間
i φ
thymine g l y c o l 5
・f o r m y l u r a c i l
C
M H a
o r
2 U
H i
c w
︒ 工 吋
NI AY
配
H 4 r 0 .
卯
hU
FO
2b O 。 ト 争O~OH
deoxyribose
H
ぐ X ト
︑ 司 J''''eB
M山
a
q d N l m m
O M A U Nィ
m︒
l︽
x a n t h i n e α‑dA
図 1 活性酸素によって生じる DNA 損傷の例。 dRf はデオキシリボース残基を 示している。
G:C → A:T トランジッションを誘発する 220 これまでに活性酸素によって誘発さ
れる変異に関して数多くの報告がなされているが、活性酸素による突然変異の
中には、未だ原因となる損傷が同定されていないものもある D 既知の DNA 損傷
によって誘発される未知のメカニズムによる変異であるのか、あるいは未知の
DNA 損傷による変異であるのか非常に興味深い。
3
表 l 活性酸素によって生じる DNA 損傷が誘発する変異
お 一
e o 一 m
. H
計 四︽ J v
e
一 ︒A
一
n d
凶一
W 7
向
一 一
dn v
VJ
‑ L H H
戸 ︑
i v
ふhydroxyadenine
thymine glycol 5‑formyluracil
ふhydroxymethyluracil ふhydroxyguanine
abasic site
hypoxanthine
xanthine
α
幽dAExperimental system
i n v i t r o
ι c o l i
的
v i t r o i n v i t
れO
E.
c o l i
NIH3T3 cellsι c o l i i n
νi t r o
E.c o l i
的
v l t r o
E.c o l i
NIH3T3 cells CO$‑7 cells E. co l i
yeast NIH3T3 cellsi n v i t r o
ι c o l i
NIH3T3 cellsi n v i t r o i n v i t r o
NIH3T3 cells 的v i t r o
Mutation
C → T , C → G
C → T
A→
C,
A→
TND a )
ND
A → G , A → T , ( A → C ) T → C
b )
T → C
ND
G
→
T G→
TG
→
T,
G→
A,
5'ーflankingmutation G→
T,
5'‑flanking mutation abasic site→
Tabasic site
→
Gabasic site
→
A,
flanking mutationA → G
A → G A → G G → A
b )
G " ‑ 7 A
G " ‑ 7 A
A→
T,
A→
Gb ) a )
Not detected.b )
Insertion of a single nucleotide was observed.4
著者は活性酸素によって生ずる変異原物質であるグリオキサールに着目したむ グリオキサールは、種々の食品(醤油、味噌、コーと一等)中に存在する
23,
24のみならず、煙草の煙中にも存在するお。また、グルコースの加熱処理 2 6 ある いは生理的条件下におけるグルコースの自動酸化
27によっても生成する o さら に、脂質過酸化の過程においてもグリオキサールの生成が報告されているお。す なわち、生体の構成成分である糖及び脂質からグリオキサールが生成すること が見い出されている。
グリオキサールの毒性及び変異原性に関する報告は数多くなされているお。
性毒性は、ラットでは LD50
コ2 . 0 2g l k g 、ハムスターでは LD50 = 0 . 7 6 g l k g と報 告されているお。サルモネラ菌 TAI00 、 TAI02 、 TAI04 株を用いた復帰変異試 験 (Ames 試験)でグリオキサールが変異原性を有すること 3 1
・3 4 、また、過酸化 水素の共存による変異原性の増強が確認されているお,お。さらに、グリオキサー ルが発癌フ。ロモーター活性を有することが報告されており 36 , 37 、発嬉のイニシエ ーション、プロモーションの両過程にグリオキサールが関与する可能性がある と思われる。
H‑ +
OH OH
図 2 dGに対するグリオキサールの付加反応
グリオキサールはジカルボニル化合物であることから、非常に反応'住に富ん
でいると考えられる。例えば、グリオキサールは生体の構成成分である蛋白質
に結合したり、蛋白質問のクロスリンクを引き起こすことが知られている
27,
38,
390
5
さらに、サリチル駿ヒドロキシラーゼのグリオキサール処理により、本来の酵 素活性が失われ、新たに NADH デヒドロゲナーゼ活性を示すとの報告もあるお。
この場合には活性に関与するアルギニン残基がグリオキサールにより修飾され たためであると考えられる o また、グリオキサールと DNA との反応では、 DNA 鎖の切断 40 や 、 DNA 中のグアニン (G) 残基に対する特異的な結合が知られて おり 4 1
,4 2 、グリオキサール ‑dG 付加体の構造は図 2 に示したような 3 環性の構造 をとっていることが NMR により明らかにされている 43
,44 0
本研究は、活性酸素の作用により DNA からグリオキサールが生成することを 見い出したことに端を発した。 DNA よりグリオキサ‑}レが生成した場合、容易 に DNA に作用し DNA 付加体を形成し変異を誘発し得ると考えられる o そこで、
グリオキサールが誘発する変異を解析し、活性酸素による突然変異にグリオキ サールが関与している可能性について検討した。 a b a s i cs i t e 等に代表されるよ うに、ある DNA 損傷の誘発する変異は実験系によって相違が見られる。このこ とから、 DNA 損傷による変異を解析する擦には、幾つかの異なった系における 変異の解析を検討するべきであると考えられ、 3 種の系を用いて実験を行った。
第 1 章では DNA 及びその関連物質(ヌクレオシド、ヌクレオチド)からのグ
リオキサール生成について述べる。第 2 章では種々の活性酸素産生系において
生成する修飾塩基の量を比較する。第 3 翠ではサ
lレモネラ菌を用いた Ames 試
験立におけるグリオキサ‑}レの変異原性について述べる。第 4 章及び第 5 輩で
は、大湯菌及び晴乳動物細胞におけるグリオキサールの変異誘発能について 1 1 買
に述べる D
6
第
1
章DNA
及びその関連物質(ヌクレオシド、ヌクレオチド)からのグリオキサール生成
グリオキサールは、序論で述べた様に、我々の生活環境中に広く存在する変 異原物質であり、また、活性酸素の作用により生体の構成成分である糖及び脂 質からも生成することが知られている o グリオキサールは DNA中のグアニン (G) 残基に対して特異的に結合することから、その変異誘発能についても非常 に興味がもたれる D
本章では、活性酸素 ( F e 2 +‑EDTA‑02) の作用により DNA 及びその関連物質 (ヌクレオシド、ヌクレオチド)からグリオキサールが生成することを見い出 した実験について述べるお。
第 1 節 DNA 及びその関連物質(ヌクレオシド、ヌクレオチド)の Fe 2 九 EDTA 処翠
EDTA は 2 価金属イオンとモル比 1 対 l のキレート型化合物を形成する。 Fe 2 + をキレートした Fe 2 ヘ EDTA 錯体はフェントン反応試薬として知られており活性 酸素産生系として一般に用いられている o フェントン反応とは過酸化水素存在 下で Fe2 + イオンの作用により活性酸素を生成する反応である (Fe2 + + H202 + H+
→ Fe 3 + + ・ OH + H20)
ベ本実験のおおまかなフロトコールを図 1 ‑ 1 に示した。基質として、仔ウシ絢
線 DNA 、 10 種のモノヌクレオチド (5'‑AMP 、5'‑GMP 、5 '
・CMP 、5'‑UMP 、
5 ' ‑ dA M P 、5'‑dGMP 、5'‑dCMP 、5'‑dTMP 、3'‑AMP 、3'‑GMP) あるいはデオ
キシグアノシン (dG) を用いた。緩衝液中 (pH7 . 4 ) 、 37
0C 、 60 分間、酸素
存在下で撹持しながら基質を Fe2 七日 DTA 処理した。続いて、次節で述べるよう
に、生じたグリオキサールを定量し、また、必要に応じて 8‑0H‑dG を定量した。
Phosphate Buffer (pH 7.4)
長 e ‑ = EDTA DNA 句 、 胃 y
FeS04
̲ Add
l たて‑‑ OPD
shaking
D e t e c t i o n 0 1 G l y o x a l A n a l y z e w i t h HPLC 入 ( 3 1 5 n m )
↑
九 日
OH pp t .
D e t e c t i o n 0 1 8
回OH
田dG A n a l y z e w i t h ECD
寸i P L C
図 1 ‑ 1 実験方法
:
之
+ : x
〉→く))Glyoxal
OPD
図 1
・2 OPD 反応
第 2 節 OPD 反応によるグリオキサールの検出
Quinoxaline 入max
315 nm
7
グリオキサールは 0 ・フェニレンジアミン ( O P D ) に結合し、 3 1 5
n mに吸収 極大を有するキノキサリンを形成することが報告されている(図 1 ・ 2) 2 4 0 そこ で 、 F e 2 + ‑ E D T A 処理により生成したグリオキサールをキノキサリンとして検出 することにした。前節で述べた F e 2 ヘ EDTA 処理後、直ちに OPD を添加して反 応させ、この反応で形成されたキノキサリンを酢酸エチルにより抽出した後、
3 1 5 n m の吸収極大を利用して逆相 HPLC を用いてキノキサリンを検出した。
8
ところで、 8 ‑ 0 H . d G は代表的な酸化的 DNA 損傷のーっとして知られており 必、活性酸素による発癌過程にも関与している可能性があると考えられている。
従って、本実験におけるグリオキサールの生成量を 8
・OH‑dG の生成量と比較す ることは非常に興味深い。そこで、基質として仔ウシ絢腺 DNA を用いた実験で は 、 Fe 2 : ヘ EDTA 処理により生成した 8 ‑ 0 H ‑ d G を同時に定量した。 DNA 中に生 じた 8 ‑ 0 H ‑ d G の検出は、電気化学検出器 ( E C D ) を接続した HPLC を用いて 常法 4 7 ,必により行った。
2 5 0 3 0 0 3 5 0 4 0 0 Wavelength ( n m )
1.
5
守A315 1
~0
.5 "10
1:O 2 4 6
y n u
・
m m
︒ ︒ 戸 し
V
‑ m T
r
叫 ー 岬...1 2 1 4
図 1
・3 逆相 HPLC によるキノキサリンの検出。グリオキサール (10μg) と
OPD との反応で形成されたキノキサリンを紫外線 ( 3 1 5nm) の吸光度 により検出した。カラムは、 BeckmanU l t r a s p h e r e ODS 5μ(4.6 x 250
mm) を用い、溶出は 50% メタノールにより行った。流速は 0 . 8 m l ! m i n
とした。キノキサリンの吸収スペクトルを上部に示した。
図 1
・3 に逆相 HPLC によるキノキサリンの検出例を示した。カラム保持時間
7 . 5分の位置に独特の吸収スペクトルを有するキノキサリンのピークが観察さ
れた。また、カラム保持時間 4分の位置のピークは、グリオキサールに対して
過剰に存在する OPD に由来するものであると考えられる o 尚、キノキサリンの
9
ピーク面積よりグリオキサール生成量を算出する際、既知量のグリオキサール と OPD との反応により予め作成した標準曲線を用いた。 DNA の Fe 2 ヘ EDTA 処 理によるグリオキサール及びら OH‑dG の生成量を表 1
・1 に示した口グリオキサ ール生成量は 8
・OH‑dG の約 17 倍であった。 DNA の Fe 2
ヘEDTA 処理における Fe2 +‑EDTA 濃度によるグリオキサール生成量の変化を図 1
・4 に示した口 Fe2 ヘ
EDTA 処理濃度の上昇に伴いグリオキサール生成量が増加した。但し、グリオ キサール生成量は Fe 2 九 EDTA 処理濃度に比例しなかった。本実験条件下では、
反応、に供した全量の Fe 2 + がフェントン反応に利用されるのではなく、幾分かは 速やかに酸化されて Fe 3 + となるためであると考えられる。さらに、 Fe2 +‑EDTA 処理時間について調べた(図 1
・5 ) 0 Fe 2
ヘEDTA 処理濃度が 5 m M の条件では、
反応開始後 5 分間でグリオキサール生成量がほぼ最大値に達した。酸素存在下 の条件では、 Fe 2
+が速やかに酸化されて Fe 3
+となるためであると考えられる O
表 1
・1 DNA の Fe2 ヘ EDTA 処理によるグリオキサール及び 8ωOH‑dG の 生成量 a
︑ ︑ ︐ ノ 剛
A 一 N 一 D 一 g
一5 1
同2 2
m
一助一 土 丘 町 一 ゆ
] 一 和 一 日
間 一
︑G
一
o ‑ 8
m 一 円
J
VU一 ハ
u
m
一 十 一
m
一切
8‑OH‑dG
(nmollmg DNA) (mmolfmol d G ) 9 . 8 士 0 . 6 1 5 . 3
土0 . 8 7
a 平均値(ロコ 3 )
土SD 値を示した。 緩衝液中 (pH7 . 4 ) 、 37
0C 、 60 分間、酸素
存在下で撹持しながら仔ウシ胸腺 DNA c l mg)のお 2+‑EDTA 処理 (5mM) を
行い、本文中で述べた方法により、生成したグリオキサール及び 8
・OH‑dG を
定量した。
1 0
@
1 5
‑ 、 自 国
c> ユ,
戸 10
6 x o
ho
申 園
。
c コ
。
霞
〈
5
6 2 3 4 5
[Fe 2 + ] (mM)
。 。
DNA の F e 2 + ‑ E D T A 処理におけるグリオキサール生成の F e 2 ヘ EDTA 濃 度依存性。緩衝液中 ( p H7 . 4 ) 、 3 7 0 C 、 6 0 分間、酸素存在下で撹弁し ながら仔ウシ胸線 DNA( 1 m g ) の F e 2 ヘ EDTA 処理を行い、本文中で述 べた方法によりグリオキサー
jレ生成量を算出した。
図 1 ‑ 4
@
8
6
4
{ 窃 ユ } 戸
c x o h r o
問
︒
Z30Ed日
2
。 o 20 40 60 80
R e a c t i o n T i m e ( m i n )
DNA の F e 2 九日 DTA 処理におけるグリオキサール生成のタイムコース o
緩衝液中 ( p H7 . 4 ) 、 3 7 0 C 、酸素存在下で撹持しながら仔ウシ胸線 DNA ( 1 m g ) の F e 2 ヘ EDTA 処理 ( 5mM) を行い、本文中で述べた方法に よりグリオキサール生成量を算出した。
図 1 る
1 1
1 0 種のモノヌクレオチド及び dG の F e 2 + ‑ E D T A 処理により生成したグリオキ サールの量を表 1 ・ 2 にまとめた。グリオキサー
lレ生成量は 5 ' ‑ dA MP において若 干高値を示したが、 2 ' ‑ 位の OH基の有無による影響はみられなかった。また、
デオキシリボヌクレオチド (dNMP) のリン酸基が S ・位に存在する場合( 5 ' ‑ dNMP) と 3 ' ‑ 位に存在する場合 ( 3 ' ‑ d N M P ) とでグリオキサール生成量に差は みられなかった。さらに、 A 、G 、C 、 T の塩基の違いによる影響は、 5 ' ‑ dA MP 以外は認められなかった。また、ヌクレオシドである dG の F e 2 + ‑ E D T A 処理に おいてもグリオキサールが生成した。これらのことから、 DNA 中のグリオキサ
表 1 ・ 2 ヌクレオチド及びヌクレオシドの F e 2 +
但DTA 処理により生成した グリオキサールの量 a
G l y o x a
l/n u c l e o t i d e o r n u c l e o s i d e (nmo
l/mg) (mmo
l/m o l ) 5 '
働AMP 1 1 3
土1 6 . 7 39+5.6 5'‑GMP 88
士1.8 32+
1.2 5 ' ‑ C l ¥ 在 P 1 1 6
土3 . 5 3 7
土1.2 5'‑UMP 1 0 1
土1 4 . 7 33+4.5 5 ¥ dAM P 172+6
1.6 57+20.6 5'‑dGMP 116+17.7 40+6.5 5'‑dCMP 9 7
土1 2 . 7 30+3.6 5'‑dTMP 1 0 7 や 2
1.2 34+7.0 3 ' ‑ dAM P 107+15.0 35+4.9 3'‑dGMP 111+16 . 4 39+6.0 dG 107+4 . 4 29+
1.0
a 平均値 ( n
コ3 )
土SD 億を示した。 基質としてヌ クレオチドあるいはヌクレオシドを用いた。緩衝 液中 (pH7 . 4 ) 、 37 " C、 6 0 分間、酸素存在下で 撹持しながら基質(1 mg) のおか担 DTA 処理
( 5 mM) を行い、本文中で述べた方法により、
生成したグリオキサール及び8 ・ OH‑dG を定量した。
12
ール生成部位は塩基部ではなく糖の部分であると予想される G
F e 2 ヘ EDTA 錯体による DNA 切断反応、にヒドロキシルラジカル(・ OH) が関 与していることが知られている 49
,50 。ヒドロキシルラジカルは DNA の糖部 Y 幽
位あるいは 4 ' . 位の水素を引き抜くことにより DNA 鎖を切断する。 1 ' . 位の水素 を引き抜いた場合には、塩基部が脱離した後 3 ' . 及び 5 ' . モノリン酸エステル断片 が生成する。一方、 4 ' . 位の水素を引き抜いた場合には 3 ヒホスホグリコレートが 生成する 5 1 。この 3 ' . ホスホグリコレートは、その一部分にグリオキサールに類 似した構造を有しており、さらなる反応によりグリオキサールが生成する可能 性が考えられる。 P o g o z e l s k i らは過酸化水素及びアスコルビン酸存在下での
F e 2 + . E D T A による DNA 鎖の切断反応に糖部 4 ' . 位あるいは 5 ' . 位の水素の引き抜 きが関与していると報告している民また、計算によれば、 B 型 DNA 中では糖 部の炭素幽水素簡の結合のうち糖部 4 ' . 位の結合が最も不安定であるとされてい る 49 0 以上のことから、 F e 2
ヘEDTA 反応系内で生成したヒドロキシルラジカ
Jレ が糖部 4 ' . 位の水素を引き抜くことによりグリオキサールが生成すると予想され
る 。
ところで、グリオキサール同様のジカルボニル化合物としてケトキサール
( R . C O . C H O ) がある。ケトキサールもまた種々の食品中に存在する変異原物 質であり、 DNA 中のグアニン (G)残基に結合することが報告されているお。
なかでもメチルグリオキサー
lレ ( C H 3 . C O . C H O ) は、種々の食品中に最も多量 に含まれるケトキサールであり、最も強い変異原性を示すことが知られている。
サルモネラ菌 TA100 株を用いた復帰変異試験 ( A m e s 試験)では、メチルグリ オキサー
lレの変異原性の強さはグリオキサールの約 5 倍であると報告されてい るへまた、メチルグリオキサールも OPD に結合しキノキサリン誘導体を形成 することが報告されているヘしかし、本実験でメチルグリオキサールは検出さ れなかった。序論でも述べたが、 DNA に結合する物質が DNA より生成した場
合、容易に DNA 付加体を形成し変異を誘発し得ると考えられる。従って、グリ
オキサールが活性酸素の作用により DNA 及びその関連物質(ヌクレオシド、ヌ
クレオチド)から生成することは、その変異誘発能において極めて重要な意味
をもっと考えられる。
1 3
第 2章 種 々 の 活 性 酸 素 産 生 系 に お い て 生 成 す る修飾ヌクレオシド
前章で述べた実験で、活性酸素の作用により DNA 及びその関連物質(ヌクレ オシド、ヌクレオチド)からグリオキサールが生成することを見い出した。そ の生成量は代表的な酸化的 DNA 損傷のーっとして知られるらOH‑dG に比べて はるかに高いものであった。従って、活性酸素の生体への影響にグリオキサー ルが関与している可能性が示された。
さて、序論でも述べたように、活性酸素によって様々な DNA 損傷が生じるこ とが報告されている。そこで、グリオキサ‑)レ及び他の様々な修飾塩基の生成 量を比較することは、活性酸素による突然変異の機構を解明する上で重要であ ると考えられる。本軍では、種々の活性酸素産生系を用いてヌクレオシドの活 性酸素処理を行った実験について述べる日。
第 1
節ヌクレオシドの活件酸素処理
活性酸素産生系としては第 1 章で述べた実験で用いた Fe2 ヘ EDTA 処理の他に、
Fe2 ヘ NTA 処理 5 5 、及び、 y 線照射の系を用いた。 EDTAと NTAは共に Fe イオ
ンのキレート剤であり、 i n v i t r o において仔ウシ胸腺 DNA に対して Fe2 +‑EDTA 処理あるいは Fe2 ヘ NTA 処理した実験では、いずれも Fe イオン単独処理に比べ てより多くの DNA 損傷が検出されている 5 6 。また、 NTA は腎発癌物質であり、
動物に Fe
・NTA 錯体を注射すると高確率で腎臓癌を誘発し 5 7 、その標的臓器で ある腎臓には 8
・OH‑dG が蓄積することが知られている弱点 9 。この錯体は 2 分子 の Fe と 2 分子の NTA により形成されたキレート型化合物(図 2
・1 )であるが、
一般に NTA は Fe とモル比 4 対 1 の混合比で用いられている。過剰の NTAに より Fe イオンを完全にキレートし溶解させるためである。
本実験では、 4 種のヌクレオシド(dA、 dG 、dC 、dT) を基質として用いた。
緩衝液中で 3 穏の活性酸素産生系 ( 5 m M Fe 2
ヘEDTA-02~ 5 m M Fe2 ヘ 20m M
NTA
・02 、及び 2900Gy の γ 線照射)による処理を行った。
A
H H
o o o o
p u p u
︒
F h n ζU H U H
く
pu
H O O C H
司4
14
図 2
・1 NTA ( A ) 及び F e ‑ N T A 錯体 ( B ) の構造
M W
OH OH
2
・OH‑dA( 1 ) g l y o x a l ‑ d G ( 4 )
OH OH OH
ふOH.dA( 2 ) 8
・OH‑dG( 5 ) 5
・OH‑dC( 7 )
OH OH OH
c y c l o
四dA( 3 ) c y c l o
・dG( 6 ) 5‑CHO‑dU ( 8 )
図 2 ‑ 2 本実験において検出した修飾ヌクレオシド。 Glyoxal‑dG、c y c l o ‑ dA及
び c y c l o
・dG の構造式中の *"は不斉炭素原子を表わしている o 右上の
枠内にグリオキサールの構造式を示した。
1 5
ヌクレオシドの活性酸素処理反応後、直ちに逆椙 HPLC による分析を行った。
前章ではグリオキサールをキノキサリンとして検出する方法を用いたが、本実 験ではグリオキサールの検出と他の修飾ヌクレオシドの検出を向時に行うため に、グリオキサールをグリオキサ
‑Jレ ‑dG 付加体(図 2 )として検出する方法を 用いた。本実験で検出された修飾ヌクレオシドを図 2 ・ 2 に示した。各々の修飾 ヌクレオシドは、各々に特有な吸収スペクトルの確認及び各々の修飾ヌクレオ シドの標準試料と保持時間が一致することにより同定した。また、既知量の標 準試料のピーク面積を基に生成量を算出した(その際に各化合物の吸収極大を 考慮し、それぞれの化合物について適当な波長を選択した) 0 尚、本実験では 図 2 ‑ 2 に示した修飾ヌクレオシド以外のものについては向定していないが、各々 の解析で検出された全てのピークについて吸収スペクトルを確認したところ、
その大部分がヌクレオシド由来のピークで、はなかった。
まず、 Fe 2 +‑EDTA 処理実験の結果について述べる。dAの Fe 2 +‑EDTA 処理で は c y c l o ・dA及び 2 心 H‑ dAの 2 種の修飾ヌクレオシドが生成した(図 2 ・ 3A)。
基質として dGを用いた場合には、グリオキサ
‑Jレ ‑dG 付加体、 c y c l o ‑ d G 及び
8‑0H‑dG の 3 種の修飾ヌクレオシドが生成した(図 2 ・ 3B) 0 また、基質とし
て dC を用いた場合には 5 ・ OH‑dC の生成、基質として dT を用いた場合にはふ
CHO‑dU の生成が観察された(図 2 ‑ 4 )。尚、 Fe 2 +‑EDTA 処理実験においては
8 ・ OH‑ dAは検出されなかった(図 2 ・ 3A) 0 c y c l o ‑ dAは不斉炭素原子 ( 5 ' ‑ C )の
存在により 2 種の立体異性体が存在するが(図 2 ・ 2 ) 、本条件においては単一の
ピークとして検出された(図 2 ・ 3A)。向様に c y c l o ‑ d G は 2 種の立体異性体、グ
リオキサ
‑ Jレ ‑dG 付加体は 4種の立体異性体が存在するが、それぞれ単一のピ
ーク及び 2 本のピークとして検出された(図 2
・・3B) 0 尚、グリオキサー
lレ ‑dG
付加体のカラム保持時間の長い方の異性体のピークと c y c l o ‑ d G のピークが、用
いた条件において同一の位置に検出された。著者は、様々な溶出条件を用いて
荷ピークの分離を試みたが、いずれの条件においても分離を行うことができな
かった。従って、グリオキサール‑dG 付加体の 2本のピークのピーク面積比が
常に一定であることを利用して、グリオキサール ‑dG 付加体及び c y c l o ・ dG の生
成量を算出した。
j こど¥ 5
¥ 一 四
ぺ ¥
//
/ /
3 4
/今、
/ ¥ / ¥
¥ ノ ¥ / ¥
¥一一一
T
十..‑,‑.280
臼320 340
(i)
v
→dA
γ
ノ¥一‑
240 260 260
A
10
の a OF
×
ωぉ ︿
240
O
一一戸一「一~一寸一-r--r一一「←吋ーーγ一一『ー→一一←「コ
20 30
40 mlnよー
10
頃ー
5 dG
。
10
の a OF
×む お︿
B
O
2 b E ‑ F
ゐ mlnC l A (A)
及びdG(B)の Fe 2
九EDTA 処現実験における逆i/f: 1 HPLC による解析。紫外線 (256nm) のl 政光度によ る解析結果を示している。検出された修飾ヌクレオシドのピークの数字は、図 2
・2 に対応している o ピーク 1 、 3 、 4 、5 については、吸収スペクトルを k 部に示している。図 A中の矢印は 8
心H‑ clAの溶 1 1 1 位躍を示している。尚、
ピーク 4 ' の吸収スペクトルはピーク 4 と同一で、あるため省略した。特に表記のないピークについては向定していな
し=。o 10
図 2
・3
8 ¥ 一 ⁝
ハ [ 一 一
︑ ¥
一 ︒
一人
j¥
ノ¥一7
ト
F→
dC
10
の' OF
×
ω ω N
︿
A
J ¥ ¥
一回、司‑‑‑‑‑‑./'
ー-T一→ー→ー守一一戸一~ー?ー「ア
1 0 2 0 3 0 4 0 η 1
1nd T
8 1
す 斗
。
10 り OF
×
ω ω N
︿
自
O
一千四一←
一一 1 一「一日「一一叩~ー~.,.1 0 2 0 30 4 0 mln
dC ( A )
及びdT( B )
のF e 2
七EDTA
処理実験における逆相HPLC
による解析。紫外線( 2 5 6nm)
の吸光度によ る解析結果を示している。検出された修飾ヌクレオシドのピークの数字は、関2
・2
に対応しているO
ピーク7
、8
に ついては、吸収スペクトルを上部に示して、いる。尚、特に表記のないピークについては同定していない。。
j 玄 1 2
・4
表 2
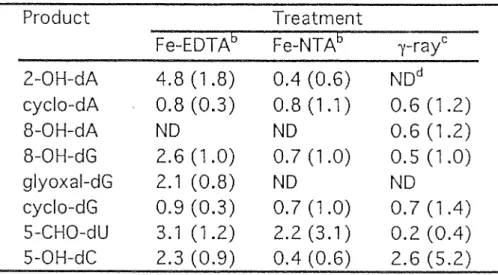
・1 ヌクレオシドの活性酸素処理実験において検出された 修飾ヌクレオシドの生成率 a
Product Treatment
F e
山EDTAb Fe‑NTAb ) ' ‑ r a y c 2‑0H‑dA 4 . 8 ( 1 . 8 ) 0 . 4 ( 0 . 6 ) NDd
一cyclo‑dA 0 . 8 ( 0
鴎3 ) 0 . 8 ( 1 . 1 ) 0.6 ( 1 . 2 ) 8
叩OH‑dA ND ND 0.6 ( 1 . 2 ) 8‑0H
同dG 2 . 6 ( 1 . 0 ) 0 . 7 ( 1 . 0 ) 0.5 ( 1 . 0 ) g l y o x a l ‑ d G 2 . 1 ( 0 . 8 ) ND ND cyclo‑dG 0 . 9 ( 0 . 3 ) 0 . 7 ( 1 . 0 ) 0.7(1 . 4 ) 5‑CHO‑dU 3 . 1 ( 1 . 2 ) 2.2(3.1) 0.2 (0 . 4 ) 5
世OH‑dC 2 . 3 ( 0 . 9 ) 0 . 4 ( 0 . 6 ) 2.6 ( 5 . 2 )
自
Y i e l d sshown a r e the r a t i o s (%) o f a m o d i f i e d n u c l e o s i d e t o t h e sum o f the m o d i f i e d and unmodified n u c l e o s i d e s . F o r m o d i f i e d dA and dG , a l l m o d i f i e d products a r e
i n 4 1 u d e d i n the c a l c u l a t i o n s . The number i n parentheses r e p r e s e n t s t h e y i e l d s r e l a t i v e t o 8‑0H‑dG.
b A s i n g l e d e o x y r i b o n u c l e o s i d e was t r e a t e d .
CA mixture o f e q u a l l amounts o f t h e f o u r n u c l e o s i d e s was t r e a t e d .
dNot d e t e c t e d .
18
表 2
・1に Fe 2 ヘ EDTA処理実験 ( 5m M Fe 2 +‑EDTA、37
0C、30分)で検出さ
れた修飾ヌクレオシドの生成率を示した。生成率は、各々のヌクレオシドに由 来する全体のヌクレオシドに対する修飾ヌクレオシドの割合として算出した。
ここで検出された修飾ヌクレオシドの生成率は、 Fe 2 +‑EDTA処理濃度が 1‑5 m Mの条件では Fe 2 +‑EDTA処理濃度に依存して上昇した。 2‑0H‑dAは最も生 成量の多い修飾ヌクレオシドであり、その生成量は代表的な酸化的 DNA 損傷の ーっとして知られる 8
・OH‑dGの1.8倍であったむ 5‑CHO‑dU、5
・OH‑dC、及び
グリオキサ~)レ -dG 付加体の生成率は 8・ OH-dG と同等であった。以上の点より、
Fe 2 九 EDTA処理による修飾ヌクレオシドの生成率は 2 心 H‑dA> 5‑CHO‑dU >
19
8
・OH‑dG>5
・OH‑dC>グリオキサール‑dG 付 加 体 >c y c l o ‑ dAであった(表 2
・. 1 ) 0 但し、グリオキサー
lレ ‑dG付加体の場合には、 Fe 2
七.EDTA処理により dG より生成したグリオキサールのなかで、未修飾で存在する dG と反応しグリ オキサール‑dG 付加体を形成したもののみが検出可能となる o 従って、グリオキ サール単独の生成量は 8
鴨OH‑dG に比べてはるかに多いものと予想される口ここ で、既知量のグリオキサールと dG との反応から作成した標準曲線より平衡定数 を算出し、本実験におけるグリオキサール生成量を概算した(図 2
・5 ) 0 その結 果、グリオキサー
jレ単独の生成震は 8
・OH‑dG の約 3 . 2 倍となった。計算上、反 応に供した dG の 8% が分解してグリオキサール形成に至ったと考えられる。前 章においてヌクレオチドの Fe 2 +‑EDTA 処理実験におけるグリオキサール生成量 は塩基の違いによる影響はほとんど認められなかったことを考慮すると、 4 種の ヌクレオシドからグリオキサールが生成した場合には、その生成率は 8
・OH‑dG の約 13 倍と予想される。
dG
+g l y o x a l
ごglyoxal‑dG
[glyoxal‑dG]
標準曲線より
平 衡 定 数K
出= 0.29 mM‑l [dG] [glyoxa I ]
Fe 2 +‑EDTA 反応において
.4
8‑0H‑dG
乙伊
cy c 1 o‑dG
dG 本 ご ケ
¥ご喰ll..
glyoxal ‑il> glyoxal‑dG
" dG (remained)
2.6%
0.9%
2.1%
94.5%
ここで、 glyoxal が a m M 生成したとすると
K
z (1‑a)X0.021
= 0.29 mM‑l : . a
出0.077m M { ( 1 ‑ a ) x 0 . 9 4 5 } x a
4 種ヌクレオシドからの生成を考慮に入れると
glyoxal 生 成 量 = 4 x a = 0.32 m M
図 2 ‑ 5 グリオキサール生成量の算出方法
20
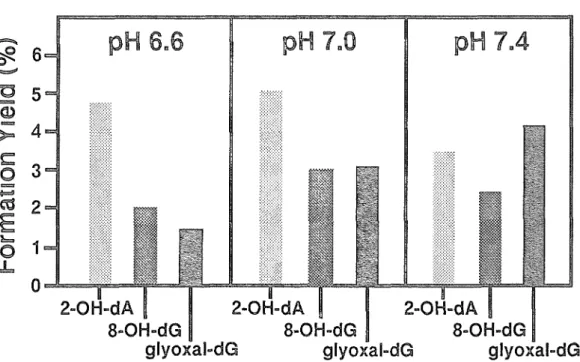
ところで、通常、生体内の pH は中性に保たれているが、様々な状況に応じて その pH は微妙に変化している。活性酸素の作用による修飾ヌクレオシドの生成 に、この pH のわずかな変化が影響を及ぼすかについて興味がもたれる o そこで、
2‑0H‑ d A 、 8‑0H‑dG 及びグリオキサー
lレ ‑dG 付加体の 3 種の修飾ヌクレオシド について Fe 2
+担DTA 処理時の pH の影響を検討した(図乙 6 ) 。中'往付近の pH のわずかな違いにより、各々の修飾ヌクレオシドの生成率に差が生じた。これ
らの修飾ヌクレオシドが生成する過程で形成される反応中間体の安定性が、 pH の違いにより異なるためであると予想される。グリオキサール‑dG 付加体の生成 率は
p狂7 . 4 で 4.2% と最も高値であったが、いずれの条件においてもグリオキサ ール単独の生成量として算出した場合には 8
・OH‑dG の 10 倍以上と考えられる。
~ 6~ pH 6 . 6 pH 7 . 0 pH 7 0 4
匂 " "
"a 5
(1)
芸 ご
4
‑ 53
一 時間S C
活2
冨
も国 1
LO L 。
ιOH‑dA I
8
脚OH‑dG
g l y o x a l ‑ d G
図 2
・6 Fe 2 ヘ EDTA 処理時の pH が 2
・OH‑ d A 、 8
・OH‑dG 、グリオキサール‑dG
付加体の生成率に及ぼす影響
A . G
6
dC
1 4
1
﹂i d l J i ‑
‑ } d i
ハ
uqペJ F→ひ
3
dT
3
川
7 1 V
川川町
20
10
の
' O F X S
︿ N
w
‑山」人ぺJ川
O
o 10 20 30 40
minヌクレオシドの F e 2 + ‑ N T A 処理実験における逆相 HPLC による解析。紫外線 ( 2 5 6nm) の吸光度による解析結果 を示している。検出された修部 j f ヌクレオシドのピークの数字は、図 2
・2 に対応している。ピーク 6 の吸収スペクト ルを図中に示している。図 A 中の矢印は 8
・OH‑dAの溶
j11位訟を示している O 尚、特に表記のないピークについて は伺定していない。
図
2‑7
2
CI‑‑
A U 1 1 1 1 1
7 1 1 V i
c"l c"l
「 寸 一 一 ? 一 ‑
れ
1
1ndT
O
Fーマー~一寸一「