学位論文
Doctoral Thesis
未熟B細胞のアポトーシスシグナル伝達経路の解析
(Ana lysis of apoptosis signal transduction pa thway in immature B cells)
田邊 香野 Kano Tanabe
指導教員
乾 誠治 教授
熊本大学大学院保健学教育部博士後期課程保健学専攻
2015 年 3 月
学位論文
Doctoral Thesis
論文題名 : 未熟B細胞のアポトーシスシグナル伝達経路の解析
( A n a ly si s o f a p o p to s i s s i gn a l t ran sd u ct io n p a th wa y i n i m ma tu r e B c e ll s )
著 者 名 : 田邊 香野 (単名) Kano Tanabe
指導教員名 : 熊本大学大学院保健学教育部博士後期課程保健学専攻 乾 誠治 教授
審査委員名 : 主 査 教授 大坪 和明
副 査 教授 乾 誠治
副 査 教授 大林 光念
副 査 准教授 畑 裕之
2015 年 3 月
[目次]
Ⅰ. 要旨 ... 1
Ⅱ. 学位論文の骨格となる公表主論文、参考論文、その他の論文リスト ... 5
Ⅲ. 謝辞 ... 6
Ⅳ. 略語一覧 ... 7
Ⅴ. 研究の背景と目的 ... 8
1. B 細胞レセプターを介したシグナル伝達応答 ...8
1.1 成熟 B 細胞における BCR 応答 ...10
1.2 未熟 B 細胞における BCR 応答 ...10
2. アポトーシス ...11
3. Bcl-xL ...14
4. ユビキチン/プロテアソーム系 ...15
5. リン酸化 ...17
6. Protein Kinase ...18
6.1 c-jun N-terminal kinase ...18
7. Protein phosphatase ...19
7.1 Protein phosphatase 6 ...20
8. α4 ...20
9. 本研究の目的 ...22
Ⅵ. 研究方法 ...23
1. 細胞株と細胞培養 ...23
2. 抗体と試薬 ...24
3. ウエスタンブロット ...24
4. 免疫沈降 (IP) ...25
5. トランスフェクション ...26
6. C-terminal α4 cDNA の作成 ...26
7. Cell Counting Kit-8 (CCK-8) を用いた生細胞数測定法による細胞生存率検出 ...26
8. siRNA 導入による内在性α4 のノックダウン ...27
9. 正常マウス未熟 B 細胞の精製 ...27
9.1 マウス骨髄から細胞の単離 ...27
9.2 B Cell Isolation Kit による B 細胞の単離 ...27
9.3 Streptavidin MicroBeads による未熟 B 細胞の単離 ...27
10. 細胞分画 ...28
Ⅶ. 研究結果 ...30
1. Bcl-xL Ser62 のリン酸化はマウス未熟 B 細胞株 WEHI-231 の BCR クロスリンクにより 誘導されるアポトーシスに重要 ...30
2. BCR クロスリンクにより誘導される Bcl-xL Ser62 のリン酸化は JNK により行われる ...31
3. PP6 は Bcl-xL Ser62 の脱リン酸化を行う ...33
4. JNK による Bcl-xL Ser62 のリン酸化を介したアポトーシスの制御は正常マウス B 細
胞でも認められる ...35
5. シグナル伝達分子 α4 の C-terminal 領域 mutant は JNK による Bcl-xL のリン酸化を
抑制する ...36
6. C-terminal α4 mutant は full-length α4 と Bcl-xL の結合を競合することでドミ ナントネガティブとして機能する。 ...37
7. α4 の C-terminal 領域 mutant を過剰発現させた WEHI-Bcl-xL 細胞では JNK による Bcl-xL のリン酸化が低下し、アポトーシス抵抗性を獲得する ...39
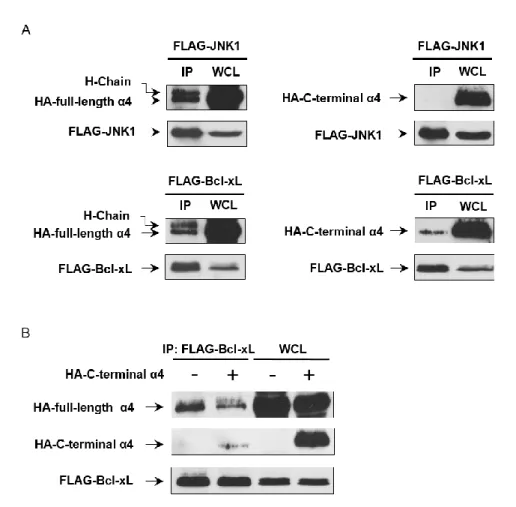
8. JNK による Bcl-xL Ser62 のリン酸化はα4 により正に制御される ...40
9. α4 は細胞質で JNK と Bcl-xL に結合しリン酸化に寄与する ...42
Ⅷ. 考察 ...43
1. 未熟 B 細胞のアポトーシスには JNK と PP6 により制御される Bcl-xL のセリン 62 番目 のリン酸化が重要である ...43
2. C-terminal α4 は JNK による Bcl-xL のリン酸化に対してドミナントネガティブとし て機能する。 ...44
3. JNK による Bcl-xL のリン酸化はα4 により正に制御されている ...45
Ⅸ. 結語 ...47
Ⅹ. 参考文献 ...48
1
[Ⅰ. 要旨]
[目的]
特異抗原が B 細胞表面に発現する B 細胞受容体 (BCR) に結合しクロスリンクを誘導すると、
細胞内にシグナルが伝えられる。BCR クロスリンクにより誘導される反応は成熟 B 細胞で は活性化であるのに対して未熟 B 細胞間では死という全く反対の結果を生じる。未熟 B 細 胞におけるアポトーシスによる細胞死は自己反応性 B 細胞の除去に重要であることが分か っている。しかし、このような反応性の違いを示す理由は解明されていない。これまでに 未熟 B 細胞のアポトーシスではミトコンドリアを介した系であること、Bcl-xL が重要であ ること以外は詳細なシグナル伝達経路は明らかとされていない。本研究では Bcl-xL のセリ ン 62 番目 (Ser62) のリン酸化に着目し、WEHI-231 細胞を用いて未熟 B 細胞のアポトーシ スシグナル伝達経路を解明することとした。さらに c-jun N-terminal kinase (JNK) によ る Bcl-xL Ser62 のリン酸化を制御する分子の機能解析を HEK293T 細胞を用いて行うことと した。
[方法]
Bcl-xL Ser62 のリン酸化は未熟 B 細胞株 WEHI-231 を 10μg/ml の 抗 IgM 抗体で刺激し、
リン酸化を特異的に認識する抗体によって検出した。Bcl-xL のリン酸化を制御する分子を 解析するために JNK 阻害剤 SP600125 およびホスファターゼ阻害剤 オカダ酸を用いた。未 熟 B 細胞の細胞生存率は Cell Counting Kit-8 を用いて検出した。未熟 B 細胞のアポトー シスシグナル伝達に対するα4 の機能を解析するため、HEK293T 細胞に Bcl-xL、JNK、α4 または C-terminal α4 の cDNA を HilyMax を用いてトランスフェクションした。α4 と他 分子の結合は免疫沈降とウエスタンブロットで検出した。さらに内在性のα4 をノックダ ウンするために RNAiMAX を用いて siRNA α4 を HEK293T 細胞にトランスフェクションした。
ラット大脳を細胞質、核、ミトコンドリアに分画したサンプルを準備し、ウエスタンブロ ットを用いて Bcl-xL、JNK、α4 の細胞内局在を決定した。
[結果/考察]
WEHI-231 細胞を抗 IgM 抗体で刺激すると Bcl-xL Ser62 のリン酸化が 15 分後に検出され、
6 時間以降から Bcl-xL のユビキチン化とタンパクの減少が認められた。この Bcl-xL Ser62
2
のリン酸化は JNK 阻害剤 SP600125 により抑制されたことから JNK が担っていることが示 唆された。次に Bcl-xL Ser62 のリン酸基を脱リン化するホスファターゼを検討したところ PP6 であることが明らかとなった。Bcl-xL Ser62 リン酸化は Bcl-xL のユビキチン化を誘導 し、分解に導くことでアポトーシスを引き起こしていることが示唆された。これらの反応 は正常マウス未熟 B 細胞でも同様に認められた。さらに JNK の Bcl-xL Ser62 のリン酸化を シグナル伝達分子 α4 が促進していることが HEK293T 細胞を用いた実験により明らかとな った。またα4 の 220 番目のアミノ酸から C 末端領域のミュータント (C-terminal α4) は JNK による Bcl-xL のリン酸化に対してドミナントネガティブとして機能することが分かっ た。Bcl-xL、JNK、α4 による反応は主に細胞質で行われていることが示唆された。
[結論]
BCR クロスリンクにより誘導される未熟 B 細胞のアポトーシスには Bcl-xL Ser62 のリン酸 化により誘導される Bcl-xL の分解が重要である。リン酸化と脱リン酸化はそれぞれ JNK と PP6 によって制御される。さらに JNK による Bcl-xL Ser62 のリン酸化はα4 が促進する ことが明らかとなった。また C-terminal α4 は Bcl-xL とは結合するが JNK とは結合せず、
ドミナントネガティブとして機能する。これらの反応は主に細胞質で行われていることが
示唆された。
3
Abstract of the Thesis
Purpose:
The crosslinking of B-cell receptor (BCR) by specific antigens on B cells induces intracellular signal transduction, which subsequently stimulates the activation of mature B cells and cell death among immature B cells. B cells with self-reactive BCRs are thus eliminated via apoptosis at the immature B cell stage. However, the mechanisms underlying this difference in BCR signaling remain to be determined.
Previous studies have shown that the apoptosis of immature B cells is induced via the mitochondrial pathway and that the expression of Bcl-xL is decreased upon BCR crosslinking. In this study, we explored the mechanisms regulating the apoptosis of immature B cells induced by BCR crosslinking and further determined the mechanisms underlying the degradation of Bcl-xL caused by Ser62 phosphorylation induced by JNK.
Methods:
WEHI-231 cells were stimulated with 10 μg/ml of anti-IgM antibodies (Ab), and the
phosphorylation of Ser62 (pSer62) associated with Bcl-xL was detected using a Western
blot analysis with anti-pSer62-specific Ab. In order to examine the roles of JNK and
PP2A subfamily phosphatases, a JNK inhibitor, SP600125, and phosphatase inhibitor,
okadaic acid, were employed. The relative viability of immature B cells was analyzed
using a Cell Counting kit-8. HilyMax transfection reagent was used for the transient
transfection of cDNAs of Bcl-xL, JNK, α4 and C-terminal α4 into the HEK293T cells,
and the associations between α4 and the other molecules were analyzed using
co-immunoprecipitation and a Western blot analysis. α4 siRNA was transfected in
order to suppress the expression of endogenous α4 in the HEK293T cells using RNAiMAX,
and the subcellular localization of Bcl-xL, JNK and α4 was analyzed using a Western
blot analysis with cytoplasmic, nuclear and mitochondrial fractions prepared from
the rat brain.
4
Results:
WEHI-231 cells were stimulated with anti-IgM Ab, and the BCR crosslinking subsequently induced the Ser62 phosphorylation of Bcl-xL within 15 minutes of the stimulation and degradation of Bcl-xL via ubiquitination after six hours of stimulation. The Ser62 phosphorylation induced by BCR crosslinking was effectively blocked by the application of the JNK inhibitor SP600125, suggesting that JNK is responsible for the Ser62 phosphorylation of Bcl-xL. The results further demonstrated that PP6 controls the Ser62 phosphorylation of Bcl-xL. Notably, Ser62 of Bcl-xL was phosphorylated by JNK, thus triggering the polyubiquitination and degradation in WEHI-231 cells. The degradation of Bcl-xL was also found to be regulated by JNK and PP6 in the normal murine immature B cells. We then prepared an assay system in which JNK phosphorylated Bcl-xL in the HEK293T cells. Using these cells, we found that α4 enhances the phosphorylation of Bcl-xL and that C-terminal α4 (220-340) functions as a dominant negative form. An analysis of the subcellular localization consequently revealed that α4 associates with both JNK and Bcl-xL in the cytoplasm.
Conclusions:
Ser62 phosphorylation and the subsequent degradation of Bcl-xL are important for the induction of apoptosis in immature B cells upon BCR crosslinking. These processes of phosphorylation and dephosphorylation are regulated by JNK and PP6, respectively.
Furthermore, we demonstrated that α4 enhances the Ser62 phosphorylation of Bcl-xL
induced by JNK. Moreover, C-terminal α4 functions as a dominant negative form in
the regulation of apoptosis of immature B cells and associates with only Bcl-xL, not
JNK. α4, thereby possibly regulating the phosphorylation of Bcl-xL induced by JNK
mainly in the cytoplasm.
5
[Ⅱ. 学位論文の骨格となる公表主論文、参考論文、その他の論文リスト]
1.公表主論文
Dominant negative form of alpha4 inhibits the BCR crosslinking-induced phosphorylation of Bcl-xL and apoptosis in an immature B cell line WEHI-231 Kano Tanabe and Seiji Inui
Biomedical Research, 36 (2015) in press
2.参考論文
1. Protein Phosphatase 6 Controls BCR-Induced Apoptosis of WEHI-231 Cells by Regulating Ubiquitination of Bcl-xL
Ryutaro Kajihara, Hitomi Sakamoto, Kano Tanabe, Kazuki Takemoto, Masayoshi Tasaki, Yukio Ando, and Seiji Inui
The Journal of Immunology, 192, 5720-5729 (2014)
3.その他の論文リスト
1. M12-Tet-Off pTRE2pur FLAG mTRAF3 細胞株の樹立 田邊 香野、坂本 瞳、乾 誠治
熊本大学医学部保健学科 紀要, 第 9 号, 27-38, (2013)
2. IL-4 による IgE へのクラススイッチと NF-κB シグナル伝達経路 田邊 香野、乾 誠治
臨床免疫・アレルギー科, 61, 504-512, 2014
6
[Ⅲ. 謝辞]
本研究を行うにあたり、全面的にご指導、ご支援下さいました熊本大学大学院 生命科学研 究部 (保健学系) 生体情報解析学分野 乾 誠治教授に深く感謝いたします。
論文作成にあたりましても、乾 誠治教授並びに熊本大学大学院 生 命 科 学 研 究 部 ( 保 健
学系) 生体情報解析学分野の皆様に深く感謝いたします。
7
[Ⅳ. 略語一覧]
Ab: 抗体
Bcl-xL: B-cell lymphoma-extra large BCR: B 細胞受容体
CD: Cluster Designation
IMDM: イスコフ改変ダルベッコ培地 IP: 免疫沈降
JNK: c-Jun N 末端キナーゼ KO: ノックアウト
MAPK: 分裂促進因子活性化タンパク質キナーゼ mIg: 膜型イムノグロブリン
OA: オカダ酸
PCR: ポリメラーゼ連鎖反応 PK: プロテインキナーゼ
PP: プロテインホスファターゼ (脱リン酸化酵素) PP6c: PP6 触媒サブユニット
siRNA: small interfering RNA
8
[Ⅴ. 研究の背景と目的]
1. B 細胞レセプターを介したシグナル伝達応答
B 細胞抗原受容体 (B cell receptor : BCR) は膜型 Immunoglobulin (mIg) M と Igα/Ig βヘテロ二量体から構成され、B 細胞表面に発現している。BCR は特異抗原を認識し、抗原 特異的に架橋される (BCR クロスリンク) ことでシグナル伝達経路の活性化を引き起こす。
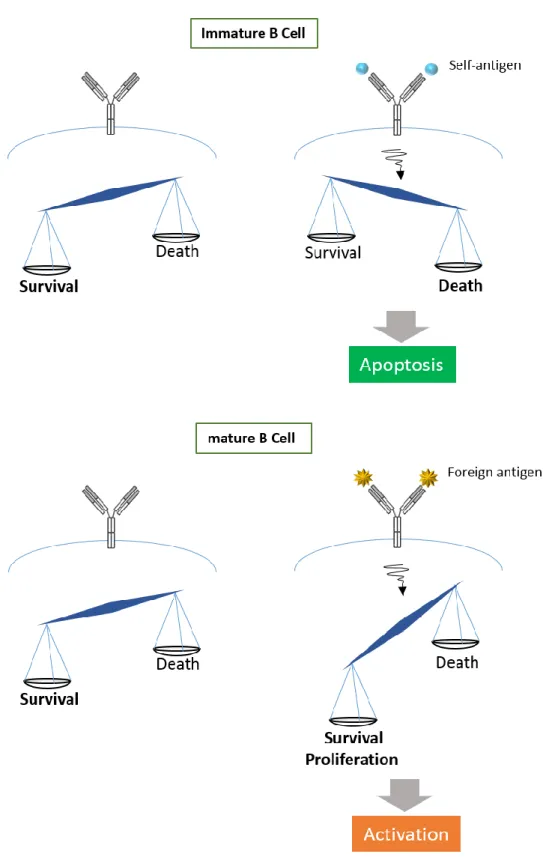
BCR から誘導されるシグナル伝達は B 細胞の成熟段階により異なった反応を示す。成熟 B 細胞では BCR が外来抗原を認識することで生存と増殖シグナルが伝達される。一方、未熟 B 細胞では抗原受容体の再編成 (レセプター編集) またはアポトーシス (クローン消失) が誘導される。これは未熟 B 細胞の分化段階で形成される BCR クロスリンクが自己抗原と 反応する自己反応性 B 細胞を除去するシステムとして機能しているためである。生体は自 己抗原と反応するような B 細胞の抗原認識部位の再編成を行うか、細胞自体を除去する (中枢性寛容) ことで成熟した自己反応性 B 細胞が生じないようにしている。このように同 一の BCR を介した刺激にもかかわらず成熟段階によって異なる反応を示すメカニズムの全 体は未だ解明されていない。
これまでの報告から、細胞内シグナル伝達を制御する分子の発現や局在の相違が同じ刺 激に対して異なる反応を示す理由であると推測されている。例えば、BCR が存在する脂質 ラフトは未熟 B 細胞では不完全であり、完全な成熟 B 細胞とでは下流に伝わるシグナルの 質が異なるということ、抗アポトーシス分子である Bcl-2 ファミリータンパクの発現が未 熟 B 細胞では成熟 B 細胞に比べて低いことが報告されている。また、B 細胞特異的に共刺 激を与え、生存に重要な AKT や SHP1 などの活性を誘導する CD19 や CD22 の発現は B 細胞が 成熟してから発現が増加する。さらに、未熟 B 細胞と成熟 B 細胞ではキナーゼやホスファ ターゼの発現や活性化レベルが異なることが下流のシグナル伝達に影響を与えているとも 推論されている (Niiro, et al.,2002)。すなわち異なる反応を呈する要因として、より強 く伝達されるシグナルが成熟 B 細胞では生存シグナルであり、未熟 B 細胞では死のシグナ ルであることが示唆されている (図 1)。
9
図 1:BCR クロスリンクによるシグナルは B 細胞の成熟段階により異なる反応を示す
10
1.1 成熟 B 細胞における BCR 応答
成熟 B 細胞は細胞膜表面に IgM と IgD を発現しており、BCR クロスリンクは細胞の生 存と増殖を誘導する。BCR の抗原認識部位に特異抗原が結合すると Igα (CD79A)/Igβ (CD79B) の immuno-receptor tyrosine-based activation motifs (ITAMs) が有する二 つのチロシン残基がリン酸化される。このリン酸化は Src ファミリーに属するチロシン キナーゼにより行われる。次にリン酸化された Igα/Igβヘテロ二量体を認識して結合 した Syk キナーゼが活性化され、B-cell linker protein (BLNK) をリン酸化する。リン 酸化された BLNK は BTK および PLCγ2 の SRC homology 2 (SH2)ドメインを介して結合す ることで、これらの分子をリクルートする。リクルートされた PLCγ2 は PIP
2の加水分 解を触媒し、DAG と IP
3が生成される。IP
3が増加することにより細胞内の Ca
2+濃度が上 昇し、NFAT 経路が活性化される。一方、DAG は PKCβを活性化し、CARD11 をリン酸化す る。リン酸化された CARD11 は最終的に NF-κB 経路を活性化する。さらに、BCR クロス リンクと同時に刺激される CD19 からは PI3K 経路が活性化される。このように複合的に 様々な経路 (NF-κB、PI3K、MAPK、NFAT、RAS 経路) が活性化され、細胞の生存や増殖 が誘導される (Niiro, et al., 2002; Ryan, et al., 2013)。
1.2 未熟 B 細胞における BCR 応答
B 細胞は骨髄で分化を開始し、Pro-B 細胞、Pre-B 細胞を経て、未熟 B 細胞へと分化す る。抗原を認識する膜型免疫グロブリンは分化に伴い遺伝子再編成を行いながら構築さ れる。まず Pro-B 細胞でカルネキシンと Igα/Igβが複合体を形成し Pro-BCR として細 胞膜に発現する。次に、Pre-B 細胞ではカルネキシンの代わりに VpreB とλ
5からなる代 替 L 鎖と、 VDJ 再編成を受けて合成された膜型免疫グロブリンの H 鎖が Igα/Igβと複合 体を形成し、Pre-BCR として発現する。Pre-BCR の抗原認識部位が正しく機能しているこ とが確認されると Pre-B 細胞は未熟 B 細胞へ分化する (ポジティブセレクション)。未熟 B 細胞では代替 L 鎖に代わり、膜型免疫グロブリンの L 鎖が再編成されて mIgM となる。
この未熟 B 細胞の分化段階では BCR クロスリンクはアポトーシスを誘導する (ネガティ
ブセレクション)。ネガティブセレクションを受ける骨髄では、未熟 B 細胞は多価の細胞
表面抗原および低抗原価の可溶性抗原に出会う。これらの抗原のうち多価抗原により
BCR クロスリンクが形成された未熟 B 細胞は自己反応性 B 細胞としてアポトーシスが誘
導されて除去される (Harverson, et al., 2004)。ネガティブセレクションを受けなか
11
った B 細胞は骨髄から出て血流に乗り、Transitional 1 (T1) B 細胞、T2 B 細胞を経て、
成熟 B 細胞へと分化する (図 2)。
B 細胞は BCR を介して無数とも言える抗原と一対一対応を行う (クローン選択説)。こ の抗原特異性は H 鎖と L 鎖に存在する抗原認識部位の構築の過程で獲得される。抗原認 識部位は V 領域、D 領域 (L 鎖は有さない)、J 領域の遺伝子断片がランダムに選ばれ、
不可逆的に再構成されることで H 鎖または L 鎖として合成される。このシステムにより 膨大なレパートリーを獲得する一方で、外来抗原のみならず自己抗原を認識してしまう ような mIgM を合成することがある。我々の生体はそのような自己反応性 B 細胞を成熟す る前に除去するために次のような機構を有している。①アポトーシスが誘導され細胞死 を起こす (BCR 誘導性細胞死)。②刺激に対して不応答性となる (アナジー) 。③再度、
VDJ 再編成を行い異なる抗原に特異性を示す BCR を形成する (レセプター再編成)。この メカニズムのうち、アポトーシスはミトコンドリアを介するカスパーゼ依存的な経路で あることが報告されている。また、JNK の関与が示唆されているが詳細なシグナル伝達 経路は明らかとなっていない。
このアポトーシスによる自己反応性 B 細胞の除去が正常に行われない場合、自己免疫 疾患や B 細胞腫、白血病などを引き起こす可能性がある。
図 2:B 細胞の分化過程
2. アポトーシス
細胞死には受動的細胞死であるネクローシスと計画的細胞死であるアポトーシスがある。
アポトーシスは電子顕微鏡観察に基づく形態学的な所見として 1972 年に報告された個体
12
をより良い状態に保つための積極的な細胞死である。アポトーシスは厳密に管理・調節さ れ発生段階での過剰な細胞の除去、自然発生する癌細胞や古くなった細胞の除去など様々 な場面で認められる。その他、放射線や熱、薬剤などの物理化学的ストレスや修復が困難 なほど DNA が傷ついた場合も同様にアポトーシスが誘導される。従って、アポトーシスが 正常に行われない場合、細胞の癌化や自己免疫疾患など様々な疾患の原因になると考えら れている。
アポトーシスは形態学的には、細胞表面の絨毛消失から始まり、細胞が萎縮して周辺細 胞から剥離する。次に核クロマチンが核膜周辺に凝集し、正常構造を失い核の断片化が起 きる。これらは顕微鏡下で観察され、その後、細胞膜は表面にブレブ (突起物) を形成し、
細胞膜構造を維持したまま断片化されてアポトーシス小体を形成する。このアポトーシス 小体はマクロファージなどにより貪食されて除去されるため、アポトーシスを起こした細 胞の内容物が周辺細胞に影響を与えることはなく、炎症は一般的に誘発されない。
アポトーシスの基本機構は線虫や昆虫から哺乳類まで極めてよく保存されており (Sin, 2001)、システインプロテアーゼであるカスパーゼとミトコンドリアが重要な働きをしてい る。アポトーシスが誘導されていない細胞では生存を維持するのために積極的にミトコン ドリア外膜透過性 (MOMP) を亢進さないようにミトコンドリア膜の安定性に努めている。
これはミトコンドリア膜間腔に局在するシトクロム c を放出させないためである。シトク ロム c の放出は下流のカスケード反応を誘導する要となる。そのためミトコンドリアでは 抗アポトーシス分子である Bcl-2 や Bcl-xL がミトコンドリア外膜に局在し、MOMP 亢進を 抑制している。この制御メカニズムはいくつかのモデルが提唱されているが統一的見解は 得られていない。これまでに Bcl-2 や Bcl-xL などが電位依存性陰イオンチャネル (VDAC) に影響を与えていることが報告されており、シトクロム c 放出制御の一端を担っていると 推論されている (Vander Heiden, et al., 2001; Malia, et al., 2007)。
カスパーゼはシトクロム c 放出前後のシグナル伝達に関与しており、カスパーゼファミ リーを形成している。カスパーゼファミリーは 3 つのサブファミリーに分けられているが、
イニシエーター・カスパーゼ:カスパーゼ-2, -8, -9, -10 とエフェクター・カスパーゼ:
カスパーゼ-3, -6, -7 がアポトーシスにおいて重要な機能を発揮する。
アポトーシス誘導刺激が細胞内に伝達されるとカスパーゼ-8, -10 が活性化される。活
性化したカスパーゼはアポトーシス促進性の Bcl-2 ファミリータンパクである Bid を切断
する。切断され短縮した tBid はミトコンドリアに移行し、Bax/Bak を活性化する。活性化
13
した Bax/Bak は MOMP を亢進させることによりシトクロム c 放出を誘導する。放出されたシ トクロム c は細胞質で Apaf1 と結合し、カスパーゼ-9 との活性化複合体 (Apoptosome) を 形成する。カスパーゼ-3 は活性化したカスパーゼ-9 により切断を受けて活性化され、細胞 が生存を保つために重要ないくつかの細胞内タンパクを切断・分解することで細胞のアポ トーシスを誘導する (Fan, et al., 2005) (図 3)。このようにカスパーゼの活性化はカス ケード状に進み、不可逆的な反応として細胞はアポトーシスを避けられなくなる。
図 3:ミトコンドリアを介したアポトーシス活性化経路。
14
3. Bcl-xL
B-cell lymphoma-extra large (Bcl-xL) は Bcl-2 ファミリーに属する抗アポトーシス 分子である。Bcl-2 ファミリータンパクは Bcl-2 homology (BH) domain という特徴的なア ミノ酸配列を有しており、最大 4 つの BH (BH1, BF2, BH3, BH4) domain を含む。Bcl-2 フ ァミリータンパクは次の 3 つのサブファミリーに分けられる。①Bcl-2 サブファミリー : すべての BH domain を有し (Mcl-l1 を除く)、抗アポトーシス作用を示す。Bcl-2, Bcl-xL, Bcl-w, A1, Mcl-1, Bcl-B が属する。②Bax サブファミリー : BH1 から 3 を有し (Bcl-xS および Bcl-GL を除く)、アポトーシス促進作用を示す。Bax, Bak, Bcl-xS, Bok, Bcl-GL が属する。③BH3-only サブファミリー : BH3 のみ有し、アポトーシス促進作用を示す。
Bad, Bid, Bim, Bik, Noxa, Puma, Bmf, Hrk が属する。これらのサブファミリーは緊密に 関わり合いながらアポトーシスの制御に寄与している (図 4)。
Bcl-xL は Bcl-x のスプライシングバリアントで、その他 Bcl-xS, Bcl-xβがバリアント として存在する。これらスプライシングバリアントはアポトーシスに対する作用が異なり、
Bcl-xL はアポトーシス抑制、Bcl-xS は促進、Bcl-xβは抑制または促進を行う。
Bcl-xL の発現量は細胞の生存に影響を与える。多くの腫瘍細胞では Bcl-xL の発現が亢 進しており、治療への抵抗性を示す要因であると推測されている。また、マウス未熟 B 細 胞株 WEHI-231 では Bcl-xL の過剰発現により BCR クロスリンクが誘導するアポトーシスに 対する抵抗性を獲得することが報告されている(Gottschalk, et al.,1994; Fang, et al., 1995; Wiesner, et al., 1997)。一方、紫外線照射によるアポトーシス誘導では Bcl-xL が 減少することが知られている (Zhang, et al., 2005; Park, et al., 2009; Ren, et al., 2011)。
Bcl-xL は 233 個のアミノ酸で構成されたタンパクで、N 端から BH4, BH3, BH1, BH2 domain と C 端に Transmembrane (TM) 領域を有する構造をしている。BH1 と BH2 は他の Bcl-2 ファ ミリーとヘテロダイマーを形成し、BH4 は抗アポトーシス活性を示す。Bcl-xL はホモダイ マーまたは Bak, Bax, Bcl-2 とヘテロダイマーを形成することができる。このうち Bax と ヘテロダイマーを形成した場合、抗アポトーシス作用は失われる。
Bcl-xL には多くのリン酸化部位が存在する。これまでにセリン 49 番目 (Ser49) のリン
酸化は細胞周期進行とチェックポイントに重要であると報告されている (Wang, et al.,
2011)。また、前立腺癌細胞株におけるセリン 62 番目 (Ser62) のリン酸化は微小管障害性
の薬剤であるタキソールや 2- methoxyestradiol (2-ME) が引き起こすアポトーシスに必
15
須である。 この Ser62 のリン酸化は JNK が行っていることが報告されている (Basu, et.al., 2003)。
図 4:Bcl-2 ファミリー
4. ユビキチン/プロテアソーム系
生体内のあらゆる場所に存在するタンパクは常に合成と分解が繰り返されている。生体
を構成するタンパクを入れ替え、ホメオスタシスの維持に必要な酵素を産生するとともに
余剰タンパクを除去するなど厳密に制御されている。タンパクの分解機序はプロテアソー
ム系とリソソーム系に分けられる。プロテアソーム系では主に細胞内に存在する不要にな
った転写因子やサイクリン、損傷を受けたタンパク、折り畳みに失敗したタンパクなどが
16
処理される。一方、リソソーム系ではエンドサイトーシスで取り込まれた細胞外タンパク やオートファゴソームで取り込まれたタンパクが処理される。
プロテアソーム系では分解するタンパクの目印としてユビキチン化が利用される。ユビ キチンは 76 個のアミノ酸からなるタンパクで 7 個のリジン残基 (K6, K11, K27, K29, K33, K48, K63) を有する。ユビキチン化はタンパク質翻訳後修飾の一種でいずれの残基もユビ キチン化が可能である。ユビキチン化が相互作用するタンパクに与える影響はユビキチン 化に利用される残基の種類と結合するユビキチンの数に依存する。このうち、プロテアソ ーム系の分解目印として用いられるのが K48 のポリユビキチン化である。K48 ユビキチン 化の検出はその後、タンパクがプロテアソームで分解されることを示唆する。
タンパクのユビキチン化は以下のプロセスで行われる。まずユビキチンの C 末端が ATP 依存的にユビキチン活性化酵素 (E1) のシステイン残基に結合する。E1 により活性化され たユビキチンはユビキチン結合酵素 (E2) のシステイン残基に渡される。すると E2 は基質 特異性の高いユビキチンリガーゼ (E3) と結合し、E3 と結合した基質タンパクのリジン残 基にユビキチンをイソペプチド結合する (ユビキチン化)。その後、基質タンパクに結合し たユビキチンの K48 を介したポリユビキチン鎖が形成されると、基質タンパクのユビキチ ン鎖を認識したプロテアソームがタンパクを内部に引き込み分解を行う (図 5)。
ユビキチン化が開始される前に基質タンパクのリン酸化が先行することが報告されてい る (Hunter, 2007)。 例えば PEST ドメインはプロリン (P)、グルタミン酸 (E)、セリン (S)、
スレオニン (T) に富んだ領域であり、このドメインを有するタンパクは PEST ドメインの
リン酸化により、E3 に認識されてユビキチン化が開始されることが明らかとなっている。
17
図 5:ユビキチン・プロテアソーム系
5. リン酸化
タンパクのリン酸化は極めて重要な翻訳後修飾であり、セリン (S)、スレオニン (T)、
チロシン (Y) の側鎖である水酸基をリン酸基に置換する反応である。リン酸化が関与する 反応はシグナル伝達、転写、細胞周期、分化、移動、増殖、アポトーシス、細胞間相互作 用、神経系および免疫系での機能制御など多岐にわたる。細胞内タンパクの約 30%は少な くとも一つの残基がリン酸化されていると言われており (Pinna, et al., 1996; Cohen, 2000)、その比率はリン酸化セリン(pSer) が 86.4%、リン酸化スレオニン (pThr) が 11.8%、
リン酸化チロシン (pThy) が 1.8%と報告されている (Olsen, et al., 2006)。
リン酸化は可逆的反応であり、リン酸化を行うプロテインキナーゼと脱リン酸化を行う
プロテインホスファターゼにより調節を受けている。リン酸基が付加または除去されると
18
タンパクは三次構造が変化し、タンパクの安定性や活性、他のタンパクとの相互作用、細 胞内局在などが変化する。
6. Protein Kinase
プロテインキナーゼ (PK) はタンパクのリン酸化を行う酵素で、基質タンパクの水酸 基に ATP のγ位のリン酸基をリン酸化エステル結合により共有結合させる。キナーゼはセ リン/スレオニンキナーゼとチロシンキナーゼに分けられる (Johnson, 2009)。セリン/ス レオニンキナーゼの種類が圧倒的に多く、約 80%を占める。
6.1 c-jun N-terminal kinase
c-Jun N-terminal kinase (JNK) は mitogen-activated protein kinase (MAPK) スー パーファミリーに属するセリン/スレオニンキナーゼで、c-Jun の N 端にある Ser63 と Ser73 をリン酸化するプロテインキナーゼとして 1991 年に同定された (Pulvere, et al., 1991)。
JNK は Tumor Necrosis Factor (TNF) や Interleukin-1 (IL-1) などのサイトカイン、
または浸透圧ストレスや酸化還元ストレス、放射線などの環境ストレス、TGF-βや EGF などの成長因子、病原体、毒素および薬剤などにより活性化されることが報告されてい る。そのため、ストレス応答性 MAPK (Stress-Activated Protein Kinase : SAPK) とも 呼ばれる。JNK の活性化は細胞死、増殖、生存、分化などと多岐にわたり (Ip, et al., 1998)、その機能は細胞の種類と刺激に依存して変化する。
JNK には JNK1, JNK2, JNK3 の 3 つのアイソフォームが存在する。JNK1/2 の発現は全般 性で広く分布している。一方、JNK3 は大部分が脳に発現し、一部神経系と精巣に発現し ている。JNK は悪性新生物との関わり合いが深く肝癌、肺癌、皮膚癌、脳腫瘍、白血病、
多発性骨髄腫、リンパ腫など様々ながんとの関係が報告されており、各アイソフォーム
により異なる作用を示すことも解析されている (Concetta, et al., 2014)。また JNK1
または JNK2 欠損 マウ スで は、 T 細 胞機 能の 喪失 によ る重 度の 免疫 不全を 呈す る
(Constant, et al., 2000)。JNK1/2 は未熟 T 細胞では高発現している一方、末梢のナイ
ーブ T 細胞ではその発現は低下している。しかし、免疫応答に伴い T 細胞が活性化する
と再び JNK1/2 の発現は増加することから、JNK1/2 は T 細胞の分化や活性段階に合わせ
て様々な機能を示すことが示唆されている。
19
JNK が関与する細胞死としてミトコンドリアを介したアポトーシスが報告されている。
JNK はミトコンドリアからシトクロム c の放出を誘導する (Hatai, et. al., 2000)。し かし JNK が標的とする分子など、そのメカニズムは明らかとされていない。制御分子の 候補としていくつかの研究より抗アポトーシス分子である Bcl-2 および Bcl-xL が挙げら れている (Maundrell, et al., 1997; Yamamoto, et al., 1999)。これらの分子を JNK がリン酸化することで抗アポトーシス機能を抑制すると推測されている。
JNK の活性化はカスケード状に生じる。まず種々の刺激に応じて、 MAP3K である MEKK1、
ASK1、TAK1、MLK などが活性化される。活性化した MAP3K は MAP2K である MKK4、MKK7 を 活性化する。活性化した MAP2K は JNK1 および JNK2 などの MAPK を活性化させることで、
標的分子となる p53、Bcl-2、Mcl-1、c-Jun、Fos、ATF などのリン酸化を誘導する。この JNK の活性化は JNK 阻害剤 SP600125 により阻害されることが分かっている (Benett, et al., 2001)。
7. Protein phosphatase
プロテインホスファターゼ (PP) は基質の水酸基に結合したリン酸基を加水分解するこ とで脱離させる、脱リン酸化酵素である。ホスファターゼはセリン/スレオニンホスファタ ーゼ (PSPs) とチロシンホスファターゼ (PTPs) に分けられる。
タンパクのリン酸化は可逆的な反応であるが、キナーゼとホスファターゼの種類には数
的差がありキナーゼがより多くの種類を有することがこれまでに明らかとなっている。キ
ナーゼもチロシンキナーゼ (PTKs) とセリン/スレオニンキナーゼ (PSKs) に分けることが
できるが、各々90 個と 428 個が確認されている。一方、ホスファターゼはチロシンホスフ
ァターゼが 107 個に対し、セリン/スレオニンホスファターゼは 30 個以下しか報告されて
いない (Johnson, et al., 2005; Lander, et al., 2001; Venter, et al., 2001)。しか
し、セリン/スレオニンホスファターゼはホロ酵素として機能することが分かっている。す
なわち、足場サブユニット、触媒サブユニット、調節サブユニットの各分子の組み合わせ
により多様な特異性を獲得することでキナーゼと同程度の多様性を確保していると考えら
れる (Shi, 2009)。
20
7.1 Protein phosphatase 6
Protein phosphatase 6 (PP6) は PP2A ファミリーに属する、セリン/スレオニンホス ファターゼである。
PP6 の触媒サブユニット (PP6c) は、足場サブユニットである ankyrin repeat-domain containing regulatory subunit (ARS)と調節サブユニットであると Sit4-associated protein (SAP) domain-containing subunit (PP6R) と共にヘテロ三量体を形成しホロ酵 素として機能する (Stefansson, et al., 2008)。調整サブユニットは複数報告されてお り、組み合わせにより PP6 は様々なタンパクを基質とすることが可能となる。一方、α4 は調節分子として作用するが PP6c とのみ結合することが知られており、α4 存在下では 足場サブユニットは PP6c とは結合できない。
PP6 は IL-2 刺激による細胞周期の制御に関与しており、がん細胞ではサイクリン D1 のコントロールを介して G1 期から S 期への移行を制限している。その他 NF-κB シグナ ル伝達を制御すること、および細胞のアポトーシスに関与しているという報告がある (Kajino, et al., 2006; Kajihara, et al., 2010; MacKeigan, et al., 2005)。PP6 の 阻害剤としてオカダ酸 (OA)、ミクロシスチン、サイクリン A が報告されている (Swingle, et al., 2007)。
8. α4
α4 (Immunoglobulin-binding protein 1 : Igbp1) は B 細胞受容体 (BCR) と複合体を 形成する Igα/Igβのうち、Igαと結合するシグナル伝達分子として 1995 年に同定された (Inui, et al., 1995)。
α4 は既存の典型的なシグナル伝達分子との相同性が認められず、明らかなキナーゼド メインも存在しない新規の分子である。一方、種間でのホモロジーは高く、アミノ酸配列 はよく保存されている。これまでにヒト、マウス、ラット、酵母などでα4 が報告されて いるが (酵母では Tap42 と呼ばれる)、ヒトとマウスではアミノ酸レベルのホモロジーは約 80%であり、特に C 端はよく保存されている (Maeda, et al., 1999)。
発現は全般性で、α4 ノックアウト (KO) マウスでは胎生致死となることから発生にも 重要な働きを担っていることが示唆された。また細胞株では Pre-B 細胞、B 細胞、骨髄腫、
T 細胞、単球、線維芽細胞でα4 の発現が確認されている (Inui, et al., 1995)。
α4 はこれまでに組織特異的 KO マウスの研究により、B 細胞の成熟や細胞増殖の分裂促
21
進的なシグナル伝達に重要であること、T 細胞では初期の T 細胞分化に重要であることが 示されている (Inui, et al., 2002; Ding, et al., 2003)。また神経系ではα4 はセリン /スレオニンホスファターゼである PP2A と Ca
2+/calmodulin-dependent protein kinase (CaMK) II と相互作用し学習記憶に対して重要な働きをしていることが明らかとなってい る (Yamashita, et al., 2006)。
α4 は PP2A ファミリーである PP4、PP6 とも相互作用することが知られており、Midline ring finger protein (MID) 1/2 などの機能調整にも関与していることが明らかとなって いる (Inui, et al., 1998; Prickett, et al., 2006; Du, et al., 2013)。
α4 は PP2Ac と直接結合し、 結合部位は 99-202AA と同定されている (Inui, et al., 1998;
Murata, et al., 1997; Nanahoshi, et al., 1998; Chen, et al., 1998)。α4 の機能は
細胞種や PP2A など酵素の基質に依存し変化する。ミエリン塩基タンパク (MBP) やヒスト
ンを基質とした場合、α4 は PP2A 活性を増強する (Inui, et al., 1998)。一方、B 細胞で
は LPS 刺激により活性化する S6 Kinase (S6K) を基質とした場合、α4 は PP2A 活性を抑制
することで S6K の活性化を促している (Yamashita, et al., 2005)。すなわちα4 は基質
に依存して PP2Ac の活性を正にも負にも調節することが明らかとなっている。
22
9. 本研究の目的
BCR クロスリンクは未熟 B 細胞のアポトーシスを誘導し、自己反応性 B 細胞の除去に重
要な役割を担っている。しかし、そのシグナル伝達経路はミトコンドリアを介した経路で
あることなど一部を除き明らかではない。そこで本研究では Bcl-xL のセリン 62 番目のリ
ン酸化に着目し、アポトーシスシグナル伝達経路を解析することで自己免疫疾患のみなら
ず発癌のメカニズムの解明の一端を担うことを目的とした。
23
[Ⅵ. 研究方法]
1. 細胞株と細胞培養
・WEHI-231
マウス未熟 B 細胞株 WEHI-231 では BCR クロスリンクによりアポトーシスが誘導され る。そのため未熟 B 細胞のアポトーシスモデル細胞株として WEHI-231 細胞が一般的に用 いられている。WEHI-231 細胞は BALB/c マウスと NZB マウスとの交雑 F1 マウスをミネラ ルオイル処理することで癌化を誘導させた B リンパ腫由来の細胞株であり、古典的な未 熟 B 細胞のように IgM
+/IgD
-ではなく IgM
+/IgD
+である。しかし、BCR クロスリンクにより 細胞周期の G0/G1 期に移行し刺激後 24-48 時間にアポトーシスを起こすことが知られて いる。
・WEHI-Bcl-xL
WEHI-231 細胞に FLAG-Bcl-xL を恒常的に過剰発現する細胞株。
・WEHI-Bcl-xL C-terminal α4
WEHI-Bcl-xL 細胞に FLAG-C-terminal α4 を恒常的に過剰発現する細胞株。
・WEHI-PP6
WEHI-231 細胞に FLAG-PP6 を恒常的に過剰発現する細胞株。
・HEK293T
ヒト胎児腎細胞株で一過性のタンパク発現に適した細胞株である。
・培養条件
37℃, 5% CO
2の条件に設定したインキュベータで培養を行った。
培養液は Iscove’s Modified Dulbecco’s Medium (IMDM) (GIBCO Laboratories, Grand
Island, NY) に 8% fetal bovine serum, 100 IU/ml penicillin, 100 μg/ml streptomycin,
50 μM 2-mercaptoethanol を添加したものを用いた。
24
2. 抗体と試薬
抗 FLAG M2 抗 体 (Stratagene, La Jolla, CA), 抗 HA 3F10 抗 体 (Roche, Basel, Switzerland), 抗 Bcl-xL 抗体 (Abcam, Cambridge, MA), 抗 Bcl-xL (pS62) phosphospecific polyclonal 抗体 (Millipore, Bedford, MA), 抗 K48 linkage-specific polyubiquitin 抗 体 (Millipore, Bedford, MA), 抗 PP2Ac 抗体 (Upstate Biotechnology, Lake Placid, NY), 抗 JNK 抗体 (Cell Signaling Technology, Danvers, MA), 抗β-tubulin III 抗体 (Sigma chemical company, St. Louis, MO), 抗 histone H3 抗体 (Active Motif, Carlsbad, CA) は ウエスタンブロットおよび免疫沈降に用いた。 抗α4 抗体は過去に精製したものを用いた (Inui, et al., 1998)。抗 PP6c 抗体はバージニア大学の Dr. Brautigan (Charlottesville, VA, USA) より譲渡された。ラットモノクローナル 11-26 ビオチン-抗マウス IgD delta chain 抗体 (abcam, Cambridge, UK), ビオチン抗マウス CD23 抗体 (BioLegend, San Diego, California) は正常マウス未熟 B 細胞の精製に用いた。細胞の刺激に用いた抗 IgM モノク ローナル抗体 M41 は バーゼル大学 (Basel, Switzerland) の Dr. Rolink より譲渡された。
また MG-132 (Millipore, Bedford, MA), SP600125 (Wako Chemical Industries, Osaka, Japan), オカダ酸 (Wako Chemical Industries, Osaka, Japan) は購入した。
3. ウエスタンブロット
Bcl-xL のリン酸化は 1% RIPA buffer [0.1% SDS, 0.5% deoxycholic acid, 1% NP40, 150mM NaCl, 50mM Tris-HCl (pH 8.0) ] にプロテアーゼ阻害 [200mM PMSF, 100mM Na
3VO
4, 1mg/ml Leupeptin, 100μM Pepstatin A, 10μg/ml Aprotinin] とホスファターゼ阻害剤 (nacalai tesque, Kyoto, Japan) を加えたものを用いて細胞の Pellet を Vortex により破砕した。
破砕後、氷上に 20 分間静置し、4℃, 13000rpm, 20 分間遠心したのちに可溶化分画を回収 した。この可溶化分画を 2×SDS-PAGE Sample buffer [0.125M Tris-HCl (pH6.8), 2% SDS, 20% glycerol, 0.002% BPB, 10% 2-mercapto ethanol] と等量混合した。SDS-PAGE ポリア クリルアミドゲルは Running Gel [0.375M Tris-HCl (pH8.8), 12% Acrylamide, 1% SDS, 0.5%
Ammonium persulfate (APS), 0.05% N,N,N’,N’-Tetramethyl ethylenediamid (TEMED) ] に Stacking Gel [0.125M Tris-HCl (pH6.8), 3.9% Acrylamide, 1% SDS, 0.5% APS, 0.05%
TEMED] を重層し作成した。SDS-PAGE 電気泳動は Running buffer [0.025M Tris, 0.2M
Glycine,1% SDS] 中で室温, 200V, 45 分間泳動した。SDS-PAGE により、サイズ依存的に分
離されたタンパクを電気ブロッティング装置を用いて PVDF 膜 (GE Healthcare, UK) にブ
25
ロットした。PVDF 膜は事前に親水化処理 (100% MtOH に 1 分間浸し、超純水に 5 分間浸水 した後、Transfer buffer [0.025M Tris, 0.2M Glycine, 10% Methanol] に 1 時間以上浸 した) を行った。ブロッティングは Transfer buffer 中で 4℃, 100V, 60 分間行い、トラ ンスファー後の PVDF 膜は Blocking 溶液 [5% スキムミルク, TBS-0.1% Tween20] で室温, 1 時間, 攪拌しながら反応させた。反応後、PVDF 膜は二回 Wash buffer [TBS-0.1% Tween20]
によりリンスを行った後に一次抗体に浸し、4℃, オーバーナイト (O/N) で反応させた。
室温にて Wash buffer を用いて、15 分 1 回, 5 分 2 回洗浄後、Horseradish peroxidase (HRP) 標識二次抗体に浸し、1 時間反応させた。Wash buffer により、15 分 1 回, 5 分 4 回洗浄後、
Pierce Western Blotting Substrate 試薬 (Thermo Scientific, CA, USA) を用いて化学 発光シグナルを誘導し、X 線フィル (FUJIFILM, Tokyo, Japan) に感光させて検出した。
4. 免疫沈降 (immunoprecipitation : IP)
0.5% CHAPS buffer [25mM Tris-HCl (pH 7.5), 150mM NaCl, 1mM DTT, 1mM EDTA, 15mM NaPPi, 100mM β-glycerophosphate, 25mM NaF, 0.05 % NaN
3, 100μM Na
3VO
4, 1mM PMSF, 10μg/ml aprotinin, 0.5% CHAPS] に プ ロ テ ア ー ゼ 阻 害 [200mM PMSF, 100mM Na
3VO
4, 1mg/ml Leupeptin, 100μM Pepstatin A, 10μg/ml Aprotinin] とホスファターゼ阻害剤を加えた ものを用いて細胞の Pellet を Vortex により破砕した。破砕後、氷上に 20 分間静置し、4℃, 13000rpm, 20 分間遠心したのちに可溶化分画を回収した。回収した可溶化分画のうち 20 μl を 2×SDS-PAGE Sample buffer と等量混合し、whole cell lysate (WCL) とした。残 りの可溶化分画には 50% w/v Protein-G セファロースまたは Protein-A セファロースを 60 μl 加え、4℃, Rotate, 1.5 時間反応させ、Pre-clear 操作とした。その後、4℃, 5000rpm, 1 分間遠心し、上清を回収した。回収した上清には目的タンパクに特異的に結合する抗体 を加え、4℃, Rotate, O/N で反応させた。沈降抗体に合わせ 50% w/v Protein-G セファロ ースまたは Protein-A セファロースを 100μl 加え、4℃, Rotate, 1.5 時間反応させた。
0.1% CHAPS buffer [25mM Tris-HCl (pH 7.5), 150mM NaCl, 1mM DTT, 1mM EDTA, 15mM NaPPi,
100mM β-glycerophosphate, 25mM NaF, 0.05 % NaN
3, 100μM Na
3VO
4, 1mM PMSF, 10μg/ml
aprotinin, 0.1% CHAPS] で 4 回洗浄し、上清を完全に除いた。2×SDS-PAGE Sample buffer
30μl をセファロースに重層し、混和後、5 分間煮沸することでセファロースと結合したタ
ンパクと沈降抗体の複合体を遊離させた。4℃, 10000rpm, 1 分間遠心したのち、上清を回
収して SDS-PAGE にて泳動後、ウエスタンブロットによりタンパクの検出を行った。
26
5. トランスフェクション
・HilyMax
HEK239T 細胞を 10cm Dish に 2×10
6個/10ml の状態に調整する。24 時間培養後に目的 の cDNA と無血清培地、HilyMax (Dojindo, Kumamoto, Japan) を混和し、室温, 15 分間 反応させた調整液を培養した細胞に添加する。添加した 4 時間後に培養液を新しいもの に変える。さらに 20 時間培養した細胞を 1×PBS EDTA (0.5mM EDTA in PBS) を用いて 回収する。
・Electroporation
WEHI-Bcl-xL 細胞を 1×10
7個に調節し、冷却した 1×KPBS (30mM NaCl, 8mM NaHPO
4, 1.5mM KH
2PO
4, 120mM KCl) で一回洗う。4℃, 1500rpm, 2 分間遠心後、冷却した 500μl の 1×KPBS に再懸濁する (1×10
7個/500μl)。WEHI-Bcl-xL 細胞 500μl と 40μg の pCMV-Tag2c C-terminal α4 および 15μg の pTRE2 pur の混和物を 1×KPBS で 100μl に なるよう調節したのち、Gene pulser Cuvette, 0.4cm (Bio-Rad, Hercules, CA) に移し、
950μF, 240V の条件で Gene Pulser II (Bio-Rad, Hercules, CA) を用いてエレクトロ ポレーションを実施した。反応液を培養液 20ml に再懸濁し、10cm Dish 2 枚に 10ml ず つ分注したのち、培養した。
6. C-terminal α4 cDNA の作成
α4 の 220 番目のアミノ酸から C 末端のアミノ酸を挟むようにプライマーを設計した。
プ ラ イ マ ー の 配 列 は 5’-GGGGATCCGAAAGAGACTCTTCAAGA-3’ お よ び 5’-GGGAATTCTCAG CCCATGTTCTGTCGGTT-3’である。pCMV TagII B α4 を NotI で直線化した Plasmid を鋳型と し、PCR を行った。回収したフラグメントを pCMV-HA vector へと組み込んだ。
7. Cell Counting Kit-8 (CCK-8) (Dojindo, Kumamoto, Japan) を用いた生細胞数測定法 による細胞生存率検出
対 数 増 殖 期 に あ る 細 胞 を 5 × 10
4個 /well に 調 節 し 、 96 well micro plate (TPP,
Trasadingen, Switzerland) に Duplication で準備した。各細胞は 10μg/ml の M41(抗マ
ウス IgM 抗体) で 48 時間刺激し、刺激終了の 3 時間前から CCK-8 溶液を添加した。生細胞
は NADH を介して添加された WST-8 を還元することでホルマザン色素を産生する。この色素
27
は 450nm の波長で計測できるため、吸光度を Enzyme-Linked Immuno-Sorbent Assay (ELISA) プレートリーダー (Bio-Rad, Hercules, CA) にて測定した。
8. siRNA 導入による内在性α4 のノックダウン
HEK293T 細胞を 24 well Plate 2 well に 0.75×10
5個/500μl ずつ調節する。このとき 培養液は 10% FCS Antibiotic free medium を用いる。24 時間培養後、無血清培地 50μl と RNAiMAX 3μl を混和した液、および無血清培地 50μl と 10μM のヒトα4 siRNA (Santa Cruz biotechnology, Dallas, TX) 2μl を緩和した液を 50μl ずつ等量混合し、室温で 10 分間反応させたものを添加する。24 時間培養後、細胞を播種しなおす。48 時間後に回収す る。
9. 正常マウス未熟 B 細胞の精製 9.1 マウス骨髄から細胞の単離
週齢 6~12 週の BALB/c マウス 6 匹から大腿骨を取り出し、中の骨髄を回収する。
骨髄は注射針を数回通し、細胞を単離させる。RBC Lysis buffer (0.83% NH
4Cl in 20mM Tis-HCl pH6.8) により、37℃, 5 分間反応させ赤血球を溶血させる。回収した細胞を MACS buffer (0.5% BSA and 2mM EDTA in PBS (pH7.2)) にサスペンドし 60μm ポアサ イズのナイロンメッシュを通したのち、カウントする。
9.2 B Cell Isolation Kit (Miltenyi Biotec, Bergisch Gladbach, Germany) による B 細胞の単離
6.5×10
7個の細胞に MACS buffer 260μl と Biotin Antibody Cocktail (Biotin-anti mouse CD43, Biotin-anti mouse CD4, Biotin-anti mouse Ter-119) 65μl 加え、4℃, 5 分間反応させる。MACS buffer 195μl と Anti-Biotin MicroBeads 130μl 加え、4℃, 10 分間反応させる。MS カラムを用いてカラムに吸着しなかった細胞を B 細胞として回 収する。
9.3 Streptavidin MicroBeads (Miltenyi Biotec, Bergisch Gladbach, Germany) によ る未熟 B 細胞の単離
2×10
7個の細胞に MACS buffer 150μl とビオチン化抗 IgD 抗体, ビオチン化抗マ
28
ウス CD23 抗体 3μl (1.5μg) 加え、4℃, 10 分間反応させる。MACS buffer 1ml 加 え、4℃, 1800rpm, 5 分間遠心したのち、上清を除く (Wash)。MACS buffer 180μl と Streptavidin MicroBeads 20μl 加え、4℃, 15 分間反応させる。再び Wash を行っ た後、MACS buffer 500μl にサスペンドする。MS カラムを用いてカラムに吸着しなか った細胞を未熟 B 細胞として回収する (6.8×10
6個)。
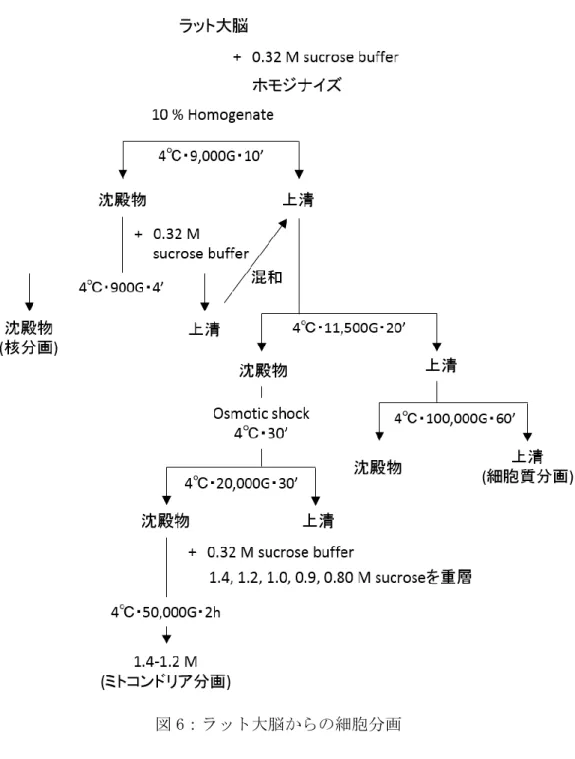
10. 細胞分画 (琉球大学 山本先生に供与頂いた)
断頭直後のラット大脳の重量を測定後、生理食塩水に浸しておく。重量に対して 9 倍量 の 0.32 M sucrose buffer で Potter homogenizer (0.3 mm クリアランス)を用いてホモジ ナイズする (1000-1200 rpm)。10% Homogenate 液を 4℃, 9000
g, 10 分間遠心し、上清と 沈殿物に分ける。沈殿物には初めに加えたのと同量の 0.32 M sucrose buffer に再懸濁し、
再びを 4℃, 9000
g, 10 分間遠心後、上清と沈殿物に分ける (核分画)。2 回に分けて回収
した上清をまとめた後、4℃, 11500
g, 20 分間遠心する。上清と沈殿物のうち、上清はさ
らに 4℃, 100000
g, 60 分間遠心したのち、その上清を細胞質分画として回収する。沈殿物
は 1g の大脳に対して 10ml の蒸留水を加え、ピペッティングしたのち 4℃, 30 分間静置す
る (Osmotic shock)。4℃, 20000
g, 30 分間遠心し、沈殿物を回収する。沈殿物を 0.32 M
sucrose buffer に再懸濁し、1.4, 1.2, 1.0, 0.9, 0.80 M sucrose を順に重層してから 4℃,
50000
g, 2 時間遠心する。上清のうち、1.4-1.2 M の層をミトコンドリア分画として回収す
る (図 6)。
29
図 6:ラット大脳からの細胞分画
30
[Ⅶ. 研究結果]
1. Bcl-xL Ser62 のリン酸化はマウス未熟 B 細胞株 WEHI-231 の BCR クロスリンクにより誘 導されるアポトーシスに重要。
未熟 B 細胞株 WEHI-231 は BCR クロスリンクによりアポトーシスが誘導される。以下の 実験では BCR クロスリンクを抗 IgM 抗体を用いてアポトーシスを誘導した。B 細胞表面に 発現している BCR は抗原特異性があり、特異的な抗原を用いない限り BCR クロスリンクは 成立しない。そこで抗原特異性がなく、殆どすべての BCR をクロスリンクさせることが可 能な抗 IgM 抗体を抗原の代わりに用いた。
これまでに、WEHI-231 細胞に抗アポトーシス分子 Bcl-xL を過剰発現させるとアポトー シス抵抗性が獲得されること、前立腺癌細胞では Bcl-xL のセリン 62 番目 (Ser62) のリン 酸化がアポトーシスに重要であることが報告されている。そこで未熟 B 細胞におけるアポ トーシスも同様に Bcl-xL Ser62 のリン酸化が重要ではないかと仮説を立て、実験を行った。
WEHI-231 細胞を 10μg/ml の抗 IgM 抗体 (M41) で刺激し、Bcl-xL Ser62 のリン酸化 (pSer62-Bcl-xL) をウエスタンブロットで解析した。すると刺激後 15 分から Bcl-xL Ser62 のリン酸化が見られた。さらに 6 時間以降ではタンパクの減少が検出された (図 7-A)。こ の減少はプロテアソーム阻害剤 MG-132 により阻害されたことから (Data not shown)、
Bcl-xL のタンパク量減少にユビキチン化が関与しているのではないかと着想し、解析を行
った。すると刺激後 6 時間から Bcl-xL のユビキチン化が検出された (図 7-B)。そこでア
ポトーシス誘導に伴う Bcl-xL のユビキチン化の上流にセリン 62 番目のリン酸化が必要な
のではないかと推測し以下の実験を行った。Bcl-xL の 62 番目のセリンをアラニンに置換
した S62A ミュータントを作成し、WEHI-231 細胞に 恒常的に 過剰発現させた細胞株
(WEHI-Bcl-xL-S62A) を作成した。この細胞株では WT-Bcl-xL を発現した WEHI-Bcl-xL 細胞
と比較しユビキチン化が低下することが分かった (図 7-C)。また WEHI-Bcl-xL-S62A 細胞
では WEHI-Bcl-xL 細胞と比較し、BCR クロスリンクによるアポトーシスに対して有意な抵
抗性を示した (図 7-D)。これらの結果から、BCR クロスリンクにより誘導されるアポトー
シスでは、先行する Bcl-xL のセリン 62 番目のリン酸化が Bcl-xL のユビキチン化に重要で
あることが明らかとなった。
31
図 7: BCR クロスリンクにより誘導されるアポトーシスには Bcl-xL のセリン 62 番目のリ ン酸化とユビキチン化による分解が重要である。
(A)WEHI-231 細胞を 10μg/ml の M41 で刺激し、pSer62-Bcl-xL をウエスタンブロットで検 出した。 (B) WEHI-231 細胞を 10μg/ml の M41 で 12 時間刺激し、回収 6 時間前から 10 μM の MG-132 で処理した。抗 Bcl-xL 抗体で IP を行ったのち、K48 ユビキチン化をウエス タンブロットで検出した。 (C)WEHI-Bcl-xL WT 細胞と WEHI-Bcl-xL S62A 細胞を 10μg/ml の M41 で 12 時間刺激し、回収 6 時間前から 10μM の MG-132 で処理した。FLAG-Bcl-xL を 抗 FLAG 抗体により IP 後、K48 ユビキチン化をウエスタンブロットで検出した。 (D) WEHI-231 細胞, WEHI-Bcl-xL WT 細胞, WEHI-Bcl-xL S62A 細胞を 10μg/ml の M41 で 48 時 間刺激したのち、細胞生存率を CCK-8 を用いて検出した。
2. BCR クロスリンクにより誘導される Bcl-xL Ser62 のリン酸化は JNK により行われる。
これまで未熟 B 細胞では JNK の活性化が BCR クロスリンクによるアポトーシス誘導に重
要であることが報告されている (Takada, et al., 2001; Takada, et al., 2006)。さらに
前立腺癌細胞では JNK による Bcl-xL のセリン 62 番目のリン酸化がアポトーシスに重要で
あることが示されている。そこで未熟 B 細胞の BCR クロスリンクにより誘導されるアポト
ーシスに重要な Bcl-xL Ser62 のリン酸化を JNK が行っているのではないかと推測し、JNK
32
阻害剤 SP600125 を用いて解析した。 WEHI-231 細胞を 10μg/ml の抗 IgM 抗体で 1 時間刺激 すると同時に SP600125 で処理を行い Bcl-xL のセリン 62 番目のリン酸化を調べた。すると Bcl-xL のリン酸化は SP600125 存在下では低下した (図 8-A)。この結果より Bcl-xL Ser62 のリン酸化を JNK が行っていることが示唆された。さらに SP600125 存在下では Bcl-xL の 分解が抑制された (図 8-B)。そこで Bcl-xL の K48 ユビキチン化を解析すると SP600125 処 理により抑制されることが分かった (図 8-C)。このことから、未熟 B 細胞におけるアポト ーシス誘導では JNK による Bcl-xL のセリン 62 番目のリン酸化が先行し、その後 Bcl-xL のユビキチン化が生じることで Bcl-xL が分解されることを明らかとした。
図 8:JNK は BCR クロスリンクにより Bcl-xL Ser62 をリン酸化する。
(A)WEHI-231 細胞を 10μg/ml の M41 で 1 時間刺激した。このとき同時に 10μM の SP600125 により処理を行った。刺激した細胞の pSer62-Bcl-xL をウエスタンブロットで検出した。
(B)WEHI-231 細胞を 10μg/ml の M41 で 12 時間刺激した。このとき同時に 10μM の SP600125
により処理を行った。刺激した細胞の Bcl-xL をウエスタンブロットで検出した。 (C)
WEHI-231 細胞を 10μg/ml の M41 で 12 時間刺激し、回収 6 時間前から 10μM の MG-132 で
処理した。このとき同時に 10μM の SP600125 により処理を行った。抗 Bcl-xL 抗体で IP
を行ったのち、K48 ユビキチン化をウエスタンブロットで検出した。
33
3. PP6 は Bcl-xL Ser62 の脱リン酸化を行う。
リン酸化はキナーゼとホスファターゼが制御する可逆的な反応である。Bcl-xL Ser62 のリン酸化は JNK が行うことが示されたため、脱リン酸化を行うホスファターゼの検討 を行った。
WEHI-231 細胞を抗 IgM 抗体で種々の時間刺激すると同時に、PP1 および PP2A ファミリ ー阻害剤であるオカダ酸 (OA)で処理を行い Bcl-xL Ser62 のリン酸化変化を検討した。
すると OA 処理を行った細胞では pSer62 Bcl-xL の増強が認められた (図 9-A)。さらに BCR クロスリンクにより誘導されるアポトーシスも、OA 処理により亢進することが明ら かとなった (図 9-B)。OA に強い感受性を示すホスファターゼには PP1、PP2A、PP4、PP6 の 4 種類がある。これらのなかから Bcl-xL Ser62 の脱リン酸化を行っているホスファタ ーゼを特定するために次の実験を行った。まず Bcl-xL と各ホスファターゼの結合を検討 した。すると PP6c のみ Bcl-xL と直接結合した (図 9-C)。このことより、Bcl-xL の脱リ ン酸化を行っているホスファターゼは PP6 の可能性が示唆された。そこで PP6 を恒常的 に過剰発現させた WEHI-231 細胞 (WEHI-PP6) を作成し、 PP6 の機能を検討した。すると、
WEHI-PP6 細胞では WEHI-231 細胞と比べ、抗 IgM 抗体により誘導される Bcl-xL の分解が
抑制された (図 9-D)。さらに、WEHI-PP6 細胞では Bcl-xL のユビキチン化も抑制されて
いることが分かった (図 9-E)。このことより、未熟 B 細胞株 WEHI-231 では Bcl-xL のセ
リン 62 番目のリン酸化を PP6 が脱リン酸化することでアポトーシス抵抗性を示すことを
明らかとした。
34
図 9:PP6 は Bcl-xL のセリン 62 番目のリン酸基を脱リン酸化することによりアポトーシス 抵抗性を示す。
(A)WEHI-231 細胞を 10μg/ml の M41 で各時間刺激した。同時に 50nM の OA で処理を行い
pSer62-Bcl-xL の変化をウエスタンブロットで検出した。 (B)WEHI-231 細胞を 10μg/ml
の M41 で 24 時間刺激したのちに細胞生存率を CCK-8 を用いて検出した。同時に 50nM の OA
で処理を行い生存率の変化を検討した。 (C)HEK293T 細胞に FLAG-Bcl-xL および FLAG-PP1c,
-PP2Ac, -PP4c, -PP6c をそれぞれ HilyMax で一過性に過剰発現させた。抗 Bcl-xL 抗体で
IP 後、抗 FLAG 抗体を用いてウエスタンブロットを行った。 (D) WEHI-231 細胞と WEHI-PP6
細胞を 10μg/ml の M41 で 12 時間刺激後、Bcl-xL をウエスタンブロットで検出した。 (E)
WEHI-231 細胞と WEHI-PP6 細胞を 10μg/ml の M41 で 12 時間刺し、回収 6 時間前から 10μM
の MG-132 で処理した。抗 Bcl-xL 抗体で IP を行ったのち、K48 ユビキチン化をウエスタン
ブロットで検出した。
35