前立腺癌における新規アンドロゲン応答遺伝子
G3BP2 の同定及び機能解析
日本大学大学院医学研究科博士課程 外科系泌尿器科学専攻
芦苅 大作 2014 年
指導教員 髙橋 悟
前立腺癌における新規アンドロゲン応答遺伝子
G3BP2 の同定及び機能解析
日本大学大学院医学研究科博士課程 外科系泌尿器科学専攻
芦苅 大作 2014 年
指導教員 髙橋 悟
【目次】
【概要・緒言】
... 1
Ⅰ 前立腺癌と疫学
... 1
Ⅱ 治療
... 2
Ⅲ 前立腺癌におけるアンドロゲンの分子作用機序
... 9
Ⅳ ゲノム医学的手法を用いた
AR
下流シグナルの探索... 10
【目的】
... 13
【対象と方法】
... 14
Ⅰ 細胞及び細胞培養液(培地)
... 14
Ⅱ
Luciferase assay ... 14
Ⅲ
Quantitative reverse transcription-PCR (qRT-PCR) ... 15
Ⅳ
Western Blot analysis ... 16
Ⅴ
Electrophoresis Mobility Shift Assay (EMSA) ... 17
Ⅵ
Immunofluorescence (
蛍光免疫染色) ... 18
Ⅶ
G3BP2
安定過剰発現細胞株(Stable
細胞)
の作製... 18
Ⅷ
small interfering RNA (siRNA) ... 19
Ⅸ
Cell proliferation assay ... 19
Ⅹ
Cell migration assay ... 20
Ⅺ
Immunohistochemistry (
免疫組織染色) ... 20
【結果】
... 22
Ⅰ アンドロゲン応答遺伝子
G3BP2
の同定... 22
Ⅱ
G3BP2 ARE
の同定及びアンドロゲン応答性検討... 22
Ⅲ
G3BP2
のアンドロゲン応答性の検討... 24
Ⅳ
G3BP2
安定過剰発現細胞株を用いた前立腺癌細胞における機能解析... 25
Ⅴ
G3BP2
発現抑制による前立腺癌細胞への影響の検討... 25
Ⅵ 前立腺癌臨床検体における
G3BP2
発現解析... 26
【考察】
... 28
Ⅰ
ChIP-chip
法及びChIP-sequence
法による新規アンドロゲン応答遺伝子の同定28
ⅡG3BP2
の分子機能... 29
Ⅲ
G3BP2
の臨床病理に関する考察... 31
Ⅳ 今後の展望
(
前立腺癌治療標的としてのG3BP2
の応用) ... 32
【まとめ】
... 34
【謝辞】
... 35
【表・図・図説】
... 36
【引用文献】
... 57
【研究業績】
... 63
1
【概要・諸言】
Ⅰ 前立腺癌と疫学
前立腺癌(
Prostate Cancer: PCa
)とは、前立腺(外腺)に発生する病気、癌の一つである。様々な組織型の悪性腫瘍が生じうるが、その殆どは腺癌
である。
前立腺癌は他の癌に比べ高齢者に多く、年齢階級別罹患数は高齢者ほど多くな
っている。
2005
~2009
年の5
年間の日本人男性の年齢調整罹患率(昭和60
年 の年齢構成を基準人口として調整:/10
万人/
年)の推計値の平均は41.3
であり、胃癌、大腸(結腸・直腸)癌、肺癌に次いで
4
番目に高く、全男性悪性新生物 全部位の11.3
%を占めている1。しかし、近年では急増傾向にあり、将来の前立 腺癌罹患数の増加に関しては、2025
~2029
年の5
年間の年間平均罹患数は118,200
人と男性癌の第1
番目となり、胃癌の103,000
人、肺癌の86,700
人を超えると予測されている1 。
また日本での
2005
~2009
年の男性年齢調整癌死亡率は8.2
であり、肺癌、胃 癌、大腸癌、肝臓癌、膵臓癌、食道癌に次いで7
番目に高く、同時期の前立腺 癌死亡数は、男性における悪性新生物死亡原因の4.8
%を占めている2 。年間前立腺癌死亡数は,上昇傾向にあり、
2000
年には4,072
人(2000
~2004
年の5
年間の平均)であるが、将来予測では、2025
~2029
年の5
年間の年平均2
死亡数は
15,700
人になると推定されており、2000
~2004
年の1.9
倍にあたり、男性癌のなかで第
5
番目の死亡数になると予測されている2 。2008
年の世界の前立腺癌罹患数は899,000
人で、死亡数は258,000
人と予測されている。また、
2030
年には年間の新規発生前立腺癌数は約170
万人となり、約
50
万人もの人が前立腺癌で死亡するとの予測もある3 。前立腺癌は、欧米の先進国に発生率の高い癌で、
2000
~2006
年の世界の国別前 立腺癌罹患率では、年齢調整罹患率(世界人口で調整:/10
万人/
年)で米国118.2
が最も高く欧州諸国も軒並み高い。一方、アジア諸国の罹患率は低く、日本は15.1
と低い4 。しかし、近年の食生活の欧米化や高齢化社会の到来などにより患者数の急
速な増加傾向などを受け、患者自身の認識も高まり、今や医療上の大きな
問題となっている。
Ⅱ 治療
前立腺癌は診断時の病期を基にして治療法を選択する。前立腺癌は治療の
選択肢が非常に多く、また選択する際は生存期間や性機能温存の問題など
患者本人の考えも重要視される。
3
1
.
手術療法(根治的前立腺全摘除術)前立腺癌が前立腺内に留まっている場合(限局性前立腺癌)では手術によ
り癌細胞が完全切除できるため、最も根治性の高い治療法として位置づけ
られている。しかし、前立腺癌の治療法には様々な選択肢があり実際に前
立腺全摘除術の適応と考えられるのは次のように考えられている 5 。
⑴
10
年以上の期待余命がある⑵ 完全切除が可能な病態であること
⑶ 全身状態や合併症などに問題がない
期待余命が
10
年以上という点では日本では75
歳くらいまでがその適応と 考えられている。またそのためにも全身状態や合併症に問題がないとされている。
手術で切除するのは前立腺、精嚢、精管の一部、膀胱頸部の一部などであ
る。また手術時に同時に所属リンパ節(閉鎖リンパ節等)郭清も行われる。
手術法としては開腹手術、体腔鏡手術(腹腔鏡手術)、内視鏡下ミニマム
創手術、ロボット支援下手術などがあり、手術経路として経恥骨後式、経
会陰式等がある。さらに勃起に関わる神経を温存する神経血管束温存手術
と温存しない手術に分かれる。
4
2
.
放射線療法(外部照射、密封小線源療法)前立腺癌の根治療法には手術療法以外に放射線療法などがあるが治療成績
に大きな差がないことや体への負担が軽いという期待から近年では放射線
治療の役割が注目されている 6 。また、進行癌で骨転移などを有する症例
に対して局所疼痛コントロール目的に行う対症的放射線治療も重要な役割
を担っている。
放射線治療技術は画像診断技術の発展とともに飛躍的に進歩してきた。
外照射では近年、三次元原体照射(
three-dimensional conformal radiation
therapy : 3D-CRT
)やコンピュータを用いて自動的に放射線照射装置を制御 す る 強 度 変 調 放 射 線 治 療 (
intensity modulated radiation therapy : IMRT
)を用いて70Gy
以上の局所への十分な高線量投与と直腸への線量抑制を両立することが可能になってきている。
他の選択肢として密封小線源療法(
brachytherapy
)がある。最も良い適応 はGleason Score
やprostate specific antigen (PSA)
値 を 合 わ せ たD’Amico
のリスク分類で低~中リスク分類の症例である。密封小線源療法は麻酔下に砕石位にて
transrectal ultrasonography (TRUS)
を用いて前立 腺を画像モニターで把握しながら会陰部よりヨウ素125
シード線源を刺入 する方法である 7 。5
3
.
ホルモン療法1940
年代にHuggins
らがホルモン療法の有用性を提唱しノーベル賞を受賞して以来今日まで、進行性前立腺癌治療の第一選択である 8 。
一般的に前立腺癌はほぼ全ての癌細胞内においてアンドロゲン受容体(
AR
) の発現が認められおり、アンドロゲン依存性を示し、アンドロゲン刺激により癌細胞が増殖することが知られている 9-10 。そして、アンドロゲン作
用を抑制するホルモン療法を施行すると、アポトーシスが誘導され癌細胞
が死滅し腫瘍は縮小していく。
ホルモン療法の方法としては、血中アンドロゲンの除去・抑制、前立腺に
おけるアンドロゲン作用の拮抗、前立腺癌細胞への直接作用などがある。
血中テストステロンを去勢域に低下させる方法として、両側精巣摘除術に
よ る 外 科 的 去 勢 術 や 黄 体 化 ホ ル モ ン 刺 激 ホ ル モ ン (
luteinizing hormone-releasing hormone
:LHRH
)アゴニストやLHRH
アンタゴニストによる内科的去勢が施行される。
LHRH
アゴニストは一過性に黄体化ホ ルモン(luteinizing hormone
:LH
)及びテストステロンを上昇させる(フ レアアップ現象)が認められ、その後LHRH
受容体のダウンレギュレーシ ョンを引き起こしてLH
及びテストステロンが著明に抑制される。このフレ アアップの予防及び去勢だけでは治療効果が不十分なことよりアンチアン6
ドロゲン剤を併用する
MAB
(maximum androgen blockade
)療法が主流 となっている。初回内分泌療法に不応となった場合はアンチアンドロゲン剤の中止(アン
チアンドロゲン除去症候群)、別のアンチアンドロゲン製剤への変更(ア
ンチアンドロゲン交代療法)を行う。さらに不応となった場合などはグル
ココルチコイド療法やエストロゲン療法などが有効な場合がある。
近年、数種類の新しいホルモン療法治療薬が開発中である。
LHRH
を直接 抑制するLHRH
アンタゴニストであるデガレリクスは、投与後のフレアア ップを起こさない。またステロイド代謝酵素(CYP17
)阻害薬であるアビ ラテロンやTAK-700
は、コレステロールからのアンドロゲン合成を阻害す ることで治療効果を示す。また、第2
世代のアンチアンドロゲン剤であるMDV3100
は、強力なアンドロゲン受容体阻害作用を有し、治療効果が期待されている。
4
.PSA
監視療法前立腺癌と診断されてもすぐに治療せずに経過を観察するという選択肢も
ある。これは転移巣がなく
Gleason Score
が低い早期癌を臨床上意義のな い癌と判断してPSA
測定で経過をみるPSA
監視療法(active surveillance
)7
という方法である。
5
.
抗癌化学療法前立腺癌はアンドロゲン感受性に増殖することから、手術療法や放射線療
法などの根治療法が適応にならない転移症例であっても、ホルモン療法に
より病状は改善し
PSA
は低下する。しかしその作用は長く続かず多くは数 年 以 内 に 治 療 抵 抗 性 と な り 、 去 勢 抵 抗 性 前 立 腺 癌 (castration resistant prostate cancer
:CRPC
)となる。現在、日本国内で
CRPC
に対する確立した治療法はほとんどないが、タキ サン系抗癌剤であるドセタキセル(docetaxel
:DTX
)療法はCRPC
に対す る治療法のひとつである。ドセタキセルはtubulin
に結合し微小管の脱重合 を阻害することにより細胞分裂を阻害する抗癌剤である。2004
年にドセタ キセルとプレドニゾロンあるいはリン酸エストラムスチンとの併用療法が有意に生存期間を延長したと報告された11-12 。
6
.
アンドロゲン受容体(AR
)を標的とした新規治療薬AR
の解析が進むのとともに癌組織中のステロイドについての研究が進んだ。
CRPC
の骨転移巣では組織中のテストステロン濃度が上昇しており、8
アンドロゲン合成に必要な酵素活性の亢進が原因であることが明らかとさ
れた13 。
また、
CRPC
ではbackdoor pathway
と言われるステロイド合成経路を獲得 することにより癌組織中のアンドロゲン濃度が上昇することも提唱された14 。
これらより
CRPC
に至る過程においてAR
の活性化が重要でありこれらを 標的とした治療が注目されている。①
CYP17
阻害薬コレステロールからアンドロゲンが合成される経路において、
CYP17
は重 要な酵素である。これを阻害することにより癌組織中のアンドロゲン濃度が低下し
AR
の活性化を阻害することが可能となる。Abiraterone
(アビラテロン)はCYP17
を阻害する経口薬であるが、ドセタキセル抵抗性
CRPC
に対してアビラテロン及びステロイド投与療法とス テロイド単独投与療法の大規模第Ⅲ相試験が行われ全生存期間が有意に延長されたことから
CRPC
に対する内分泌療法として注目されている。日本 で も 臨 床 試 験 が 実 施 さ れ て い る 。 他 にTAK700
も ア ビ ラ テ ロ ン 同 様 にCYP17
阻害薬だが、阻害する酵素活性の選択性に相違がありアビラテロンに比べコルチゾール合成がある程度維持されるといわれている。
9
②
MDV3100
(enzalutamide
)非ステロイド性アンチアンドロゲン薬として広く日本でも使用されている
ビカルタミドやフルタミドは
AR
に点突然変異などがある場合に作動性に 働くことがあることが知られている。そこで第二世代アンチアンドロゲン剤として注目されている
MDV3100
はAR
への親和性が高く、作動性もな く、AR
の核内への移行も阻害することが示されている 15 。大規模第Ⅲ相 試験の中間発表ではAFFIRM
試験(ドセタキセル後)において生存期間の 予測中央値が有意に延長したとの報告があり期待されている 16-17 。Ⅲ 前立腺癌におけるアンドロゲンの分子作用機序
前立腺癌はアンドロゲンによりその増殖の制御を受けている。アンドロゲ
ンのうち、血液中で最も高濃度に存在し重要な役割を果たしているのがテ
ストステロンであり、その約
90
%は精巣由来である。視床下部からLHRH
が分泌され、下垂体でのLH
の合成分泌を刺激する。そして、LH
は精巣に 働いてテストステロンの合成分泌を促進する。テストステロンは標的臓器である前立腺細胞内に取り込まれると、
5α
還元酵素の働きによってジヒド ロテストステロン(dihydrotestosterone
:DHT
)に代謝される。DHT
はテ ストステロンよりも活性が強く標的細胞内のAR
と結合して複合体を形成10
する。このように
DHT
がリガンドとして核内受容体であるAR
と複合体を 形 成 し 、 核 内 に 移 行 す る と ア ン ド ロ ゲ ン 応 答 配 列 (androgen response element : ARE
)に結合して、他の転写活性化因子(coactivator
)とともに標的遺伝子のプロモーター周辺のヒストン修飾を変化させることにより転
写活性を促進、
mRNA
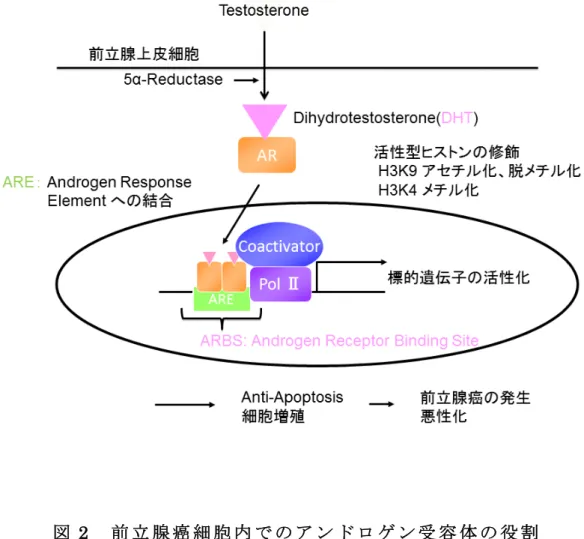
及び蛋白の発現を介してアンドロゲン作用を現す。アンドロゲン応答遺伝子の中には過剰発現することにより、
anti-apoptosis
、 細胞増殖に働き前立腺癌の発生、悪性化を促すものがある18-21(
図2)
。その ためAR
を阻害する抗アンドロゲン療法は前立腺癌に対する確立された治 療法の一つであるが、しばしば治療中に癌が再燃しアンドロゲン非依存的に増殖する治療抵抗性の獲得が臨床上の大きな課題となっている 22-23 。そ
の機序については不明な点が多いが、前立腺癌細胞内の
AR
発現強度に関係 なく、AR
自身の活性型変異や増幅、癌組織中でのステロイド合成経路の獲 得など、アンドロゲンシグナルが活性化されている状態となっているとの報告がある 24-25 。そのためアンドロゲン応答遺伝子がホルモン療法不応性
癌の増殖に働いている可能性が示唆されている。
Ⅳ ゲノム医学的手法を用いた
AR
下流シグナルの探索前立腺癌の発症及び進展に重要な役割を果たしているアンドロゲンの標的
11
遺伝子に着目した研究が盛んに行われいくつかの報告がある。東京大学大
学院医学系研究科抗加齢医学講座では、より直接的な
AR
の標的遺伝子を同 定するために、AR
陽性前立腺癌細胞株にアンドロゲンを付加しゲノムDNA
上 に 結 合 し たAR
を 特 異 抗 体 に よ り 同 定 す る ク ロ マ チ ン 免 疫 沈 降 法(
Chromatin immunoprecipitation : ChIP assay
)に、標識したChIP DNA
をゲノムタイリングアレイにハイブリダイゼーションさせ、有意なシグナルを得られる領域を検出する
ChIP-chip
法や、次世代シークエンサーによ るシークエンスを組み合わせたChIP-sequence
法などを施行してきた。こ れ ら に よ り ゲ ノ ム ワ イ ド にAR
結 合 領 域 を 含 むAndrogen Receptor Binding Sites
(ARBSs
)を約500bp
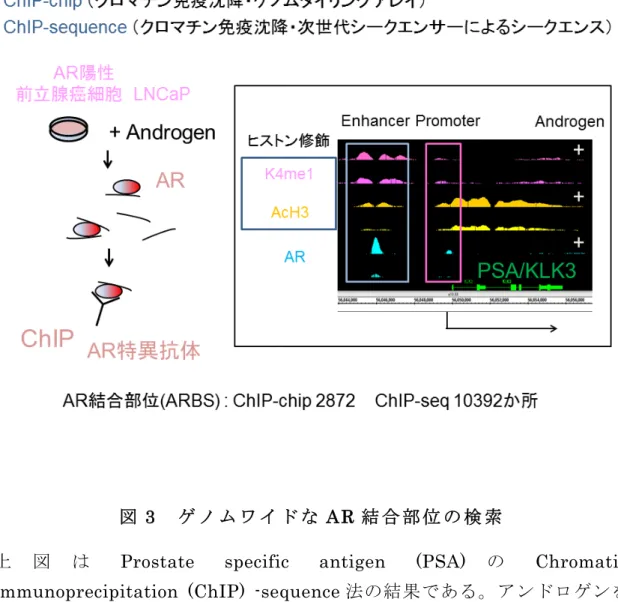
単位で同定できるようになった。我々は
ChIP-chip
法により2872
箇所(p-value < 10
-5)、ChIP-sequence
法に より10392
箇所(> 10
倍)のARBS
を同定した 26-27 (図3
)。これらの方法により転写因子の結合部位をゲノムワイドに同定可能となっ
た。また、これらを用いてより直接的な
AR
のターゲット遺伝子である新規 アンドロゲン応答遺伝子を同定し、機能解析することで前立腺癌細胞内におけるアンドロゲンの分子作用機序が徐々に明らかとなってきている 28-31 。
このように
ChIP-chip
法及びChIP-sequence
法により同定されたAR
結合 部位の中から、より直接的なAR
応答遺伝子を同定し発現制御解析をするた12
めに、全ゲノムの遺伝子転写開始点より
3kb
以内にAR
結合部位(ARBS
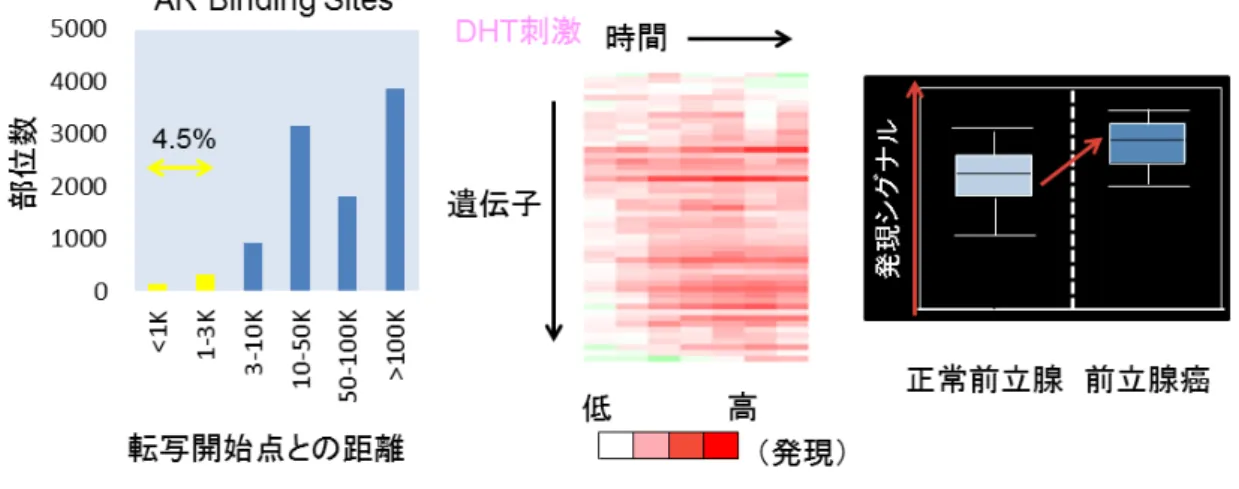
) を有する遺伝子を抽出してきた。得られたARBS
のうち転写開始点3kb
以 内のものは約4.5
%で、これらについてCap Analysis of Gene Expression
(
CAGE
), RNA-sequence
法,
マイクロアレイ法などによりアンドロゲン 応答性発現誘導解析を行い、さらに臨床サンプルによるマイクロアレイ発現解析(
Oncomine
)で正常前立腺組織に比べ前立腺癌での発現が有意に上昇しているものに絞り込みを行った(図
4
)。以上の方法により全
9
遺伝子が抽出されたが、アンドロゲン応答遺伝子と して既報の遺伝子以外のものについて、さらなる機能解析を行いG3BP2
に 絞り混んだ。13
【目的】
前述の方法により同定された新規の
AR
標的遺伝子群の中で我々はG3BP2
(
Ras GTPase activating protein (SH3 domain) binding protein2
)に着目 した。本研究では新たな前立腺癌の治療標的や診断マーカーの発見へとつながることを目指し、
G3BP2
のアンドロゲン応答性の検討及び前立腺癌細 胞内での機能解析を行い検討した。14
【対象と方法】
Ⅰ 細胞及び細胞培養液(培地)
前立腺癌細胞は、
AR
陽性ヒト前立腺癌細胞株LNCaP
細胞(the American Type Culture Collection, Manassas, VA, USA
)及び別のAR
陽性前立腺癌細胞株
VCaP
細胞(the American Type Culture Collection, Rockville, MD
) を使用した。LNCaP
細胞の細胞培養にはRPMI1640
に10
%ウシ胎児血清(
fetal bovine serum; FBS, JRH Bioscience)
を混合した培地にて行い、VCaP
細胞の細胞培養にはDulbecco’s modified Eagle’s medium
(DMEM
)に
15
%FBS
を混合した培地にて37
℃、5
%CO
2条件下で培養を行った。また、アンドロゲン(
DHT
)刺激前には約72
時間フェノールレッドフリー 培養液にチャコール処理された5
%FBS
を混合した培地にて培養を行い、ホルモン除去を行った。
Ⅱ
Luciferase assay
pGL3-basic Luciferase vector
(Promega, Madison, WI, USA
)を用いて、アンドロゲン応答遺伝子候補
G3BP2
のTSS
近傍のARBS
を含むPromoter/
Enhancer
領域(Chromosome 4: 76816078-76818629
(hg18
),
全2551bp
)15
を
insert
としてG3BP2 promoter-Luc construct
を作成した。また、その
ARBS
中に含まれるARE
領域(Transfac
により同定)の中央3 bp
よりそれぞれ上流及び下流に2 bp
の位置の塩基に変異を入れた(それぞれ
C→A
及びG→C
)G3BP2 mutation-Luc construct
を作成した。それぞ れのconstruct
はLNCaP
細胞にXP DNA Transfection Reagent
(Roche Applied Science, Mannheim, Germany
)を使用してtransfection
し48
時間培養後、リガンド刺激を行い検討した。
Tk-PRL
プラスミド10 ng
を一緒 に遺伝子導入し、遺伝子導入の効率で標準化するための内部コントロールとして使用した。
Ⅲ
Quantitative reverse transcription-PCR
(qRT-PCR
)細胞より
RNA
をISOGEN
(Nippon Gene, Tokyo, Japan
)を用いて溶解、抽出した。一本鎖
cDNA
はPrimescript RT reagent kit
(TAKARA, Kyoto, Japan
)を用いてmRNA
より逆転写合成した。qRT-PCR
はABI step one
(
Life technologies, Carlsbad, CA, USA
)を用いて行い遺伝子発現量を定 量化した。用いた
Primer sequences
は以下の通りである。GAPDH
forward: 5’-GGTGGTCTCCTCTGACTTCAACA- 3’
16
reverse: 5’-GTGGTCGTTGAGGGCAATG- 3’
G3BP2
forward: 5’-TGTGGAACTTCGCATCAATACC- 3’
reverse: 5’-AAACGTACTTCCCCTCGAAACA- 3’
PSA
forward: 5’-CAGGAACAAAAGCGTGATCTTG- 3’
reverse: 5’-GCTGTGGCTGACCTGAAATACC- 3’
TMPRSS2
forward: 5’-TCAACCCCTCTAACTGGTGTGA- 3’
reverse: 5’-AGGCGAACACACCGATTCTC- 3’
また、抗アンドロゲン剤として
Bicalutamide
(Sigma, St Louis, MO, USA
) を使用した。Ⅳ
Western blot analysis
Whole cell lysate
をLysis buffer
(NP40 buffer : 50mM Tris-HCl ph8.0, 150mM NaCl, 1.0% NP40
) に て 溶 解 し て 回 収 、 蛋 白 濃 度 測 定 はBCA protein assay kit
(Pierce Biotechnology, Rockford, IL, USA
)を用いた。約
30 µg
の蛋白を8
%SDS-polyacrylamide gel
で電気泳動しImmobilon-P Transfer Membrane
(Millipore, Billerica, MA, USA
)に転写させた。メンブレンはそれぞれ後述の
1
次抗体及び2
次抗体と反応させthe Western
Blotting Chemiluminescence Luminol Reagent
(Santa Cruz
17
Biotechnology, Santa Cruz, CA, USA
)を用いて発色させた。抗体はそれぞれ
Anti-G3BP2 antibody (Abcam, Cambridge, MA, USA)
、Anti-Flag antibody (Sigma, St Louis, MO, USA)
、Anti-β-actin monoclonal antibody (Sigma, St Louis, MO, USA)
を用いた。Ⅴ
Electrophoresis Mobility Shift Assay
(EMSA
)LNCaP
細胞を15cm dish
で培養後、Vehicle
処理とDHT100 nM
処理を48
時 間 行 っ た 後 に 回 収 、 核 内 タ ン パ ク 質 の 分 離 を 施 行 し た 。 核 抽 出 は
Hypotonic Buffer
(20mM HEPES pH7.9, 10mM KCl, 1mM EDTA, 1mM EGTA, 0.65% NP-40, 1mM DTT
) に て 施 行 し 核 内 タ ン パ ク 質 の 分 離 はRIPA Buffer
(25 mM Tris-HCl pH 7.6, 150 mM NaCl, 1.0% NP- 40, 1.0%
sodium deoxycholate
、0.1% SDS
)を用いて抽出した。ARE
及びARE mutation
のそれぞれのオリゴ核酸の配列は以下の通りである。
ARE
:CTCTGCGGAGCACGCGCAGTACGCTGC
ARE mutation
:CTCTGCGGAGCAAGCGCACTACGCTGC
また核内タンパク質とオリゴ核酸は室温で
15
分間incubation
を行い、Gel
shift assay
にはDIG Gel Shift Kit, 2
ndGeneration
(Roche Applied Science,
18
Mannheim, Germany
)を用いて検討した。抗
AR
抗体とのincubation
時間及び濃度は図に記載の通りである。ま た 、 抗 体 は
Anti-AR rabbit monoclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA)
を用いてassay
を施行した。Ⅵ
Immunofluorescence
(蛍光免疫染色)ホルモン除去した
LNCaP
細胞にVehicle
及びDHT100 nM
処理48
時間後 にそれぞれ4
%パラフォルムアルデヒド、0.1
%Triton X-100
、2
%BSA
処理 を順次行い、G3BP2
抗体(1
:200
希釈)処理24
時間後にAlexa-488
ヤギanti-Rabbit IgG
(1
:1000
希釈)処理1
時間施行した。核の染色にはDAPI
(
Nacarai tesque, Kyoto, Japan
) を 使 用 し た 。 撮 影 はconfocal laser scanning microscope
(model FLUOVIEW FV10i; Olympus, Tokyo, Japan
)にて行った。
Ⅶ
G3BP2
安定過剰発現細胞株(Stable
細胞)の作製pcDNA3.1-FLAG
に前述のLuciferase construct
と同様にしてG3BP2
をinsert
しLNCaP
細胞にtransfection
した。G418 500 µg/ml
処理しN
末端に
Flag
付きのG3BP2
安定過剰発現細胞株(Flag-G3BP2 #1, #2
)を作製19
した。
Ⅷ
Small interfering RNA
(siRNA
)以 下 の
Stealth RNAi
及 び コ ン ト ロ ー ルRNAi
(Life technologies, Carlsbad, CA, USA
)を購入し使用した。siG3BP2 #1
(Cat.No. G3BP2 HSS114989
)siG3BP2 #2
(Cat.No. G3BP2 HSS114990
)上記の
siRNA
は全て10 nM
にて使用した。Transfection
はLipofectamine RNAiMAX Reagent
(Life technologies, Carlsbad, CA, USA
)を用いた。Ⅸ
Cell proliferation assay
細 胞 増 殖 率 を
MTS proliferation assay
(Cell Titer 96 Aqueous One Solution Cell proliferation assay, Promega, Madison, WI, USA
)を用いて定量的に検討した。
Stable
細胞(G3BP2 #1, #2
)及びpcDNA3.1-FLAG
空Vector transfection
細胞(Vector #1, #2
)をそれぞれ96 well dish
の各well
に
3000
個の細胞を4 well
ずつ入れ0, 24, 48, 72
時間培養後に増殖率を測 定した。20
また、
siRNA
を用いたassay
では各well
に3×10
3個の細胞を入れ24
時間 後(day 1
)にsiRNA
をtransfection
しday 6
まで測定した。Ⅹ
Cell migration assay
細 胞 の 遊 走 能 を 検 討 す る た め フ ィ ブ ロ ネ ク チ ン 処 理 し た
Cell Culture Insert with an 8.0-µm pore size PET filter
(Becton Dickinson
)を用いた。Stable
細胞及びsiRNA
を用いた実験系において、5×10
4個の細胞を各well
に入れ
24
時間培養しメタノール固定後ギムザ染色しpore
を通過した細胞 数を蛍光顕微鏡(Nikon eclipse TE 2000-U; Nikon, Tokyo, Japan)×400
視 野にてランダム5
視野で撮影検討した。Ⅺ
Immunohistochemistry
(免疫組織染色)日本大学医学部附属板橋病院にて前立腺癌の診断のもと前立腺全摘除術を
施行した
101
例について、anti-G3BP2 antibody
(1
:100
希釈)を用いた 免疫組織染色をperoxidase catalyzed signal amplification
(CSA
)system
(
DAKO, Carpinteria, CA, USA
)を用いて施行した。評価法は光学顕微鏡(
×200
)で全細胞数のなかで陽性細胞数が何%存在するか(score 0,
陽性 細胞なし; score 1, < 1/100; score 2, 1/100 to 1/10; score 3, 1/10 to 1/3;
21
score 4, 1/3 to 2/3; score5, > 2/3
)、及び染色強度(score 0,
染色されず; score 1,
弱; score 2,
中; score 3,
強)を足し合わせてIR score; 0-8
までで評価した。既報論文を参考にして
IR score
≧7
を強発現、IR score
≦6
を弱発現とした 21,30 。ま た 、 全 て の 統 計 処 理 の 結 果 は 平 均 値
±
標 準 偏 差(SD)
に て 表 し た 。Student’s unpaired two-tailed t test
は2
パラメータ間の比較で用い、one-way ANOVA
は2
群間比較の際用い、p < 0.05
の場合統計的有意差有りとした。また、臨床サンプルでの統計学的有意差は
χ
2検定または相関解 析で検討し、統計ソフトはGraphpad Prism ver 5.0 software
(GraphPad Software Inc, CA, USA
)及びJMP. Pro 9 software
(SAS Institute, Cary, NC
)を使用した。なお、本研究は日本大学医学部附属板橋病院倫理委員会の承認を得て施行した。
22
【結果】
Ⅰ アンドロゲン応答遺伝子
G3BP2
の同定前述のごとく
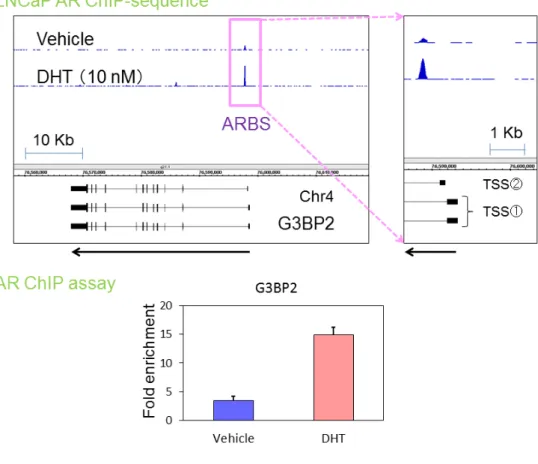
AR
陽性ヒト前立腺癌細胞株LNCaP
細胞を用いてゲノムワイ ドにアンドロゲン応答遺伝子を検索した。ChIP-sequence
法によりG3BP2
のTSS
近傍(3kb
以内)にリガンド刺激によって多くのAR
の結合が認め られる領域を同定した(図5
上)。また、AR ChIP assay
によりリガンド 付加での有意な結合の増加を確認できた(図5
下)。G3BP2
はChromosome 4: 76786990-76817629 (hg18), reverse strand
に位置しており、核蛋白やRNA
輸送の足場蛋白としての機能や様々なシグナリングとの関与が報告されている 32-35 。また、癌との関連性として、乳癌細胞において過剰発現し
ていることや、大腸癌細胞での細胞増殖への関与、
p53
及びMDM2
と結合 しubiquitin
化やdegradation
を阻害している等の報告がある 37-39 。Ⅱ
G3BP2 ARE
の同定及びアンドロゲン応答性の検討まず
G3BP2
のアンドロゲンやAR
による発現制御メカニズムを解析した。G3BP2 TSS
近傍のARBS
内及び周囲のモチーフ検索を公共データベース(
BIOBASE Biological Databases
内にあるTransfac
)を用いて検索した23
ところ、
ARBS
内に1
つのAndrogen responsive element
(ARE
)を認め、また周囲にはいくつかの
AR
関連転写因子群(Oct1, FOXA1/ HNF3
)など の結合配列が認められた(図6
)。次に、実際にこの
ARE
を介した結合が起きているのか、またこのAR
の結 合がアンドロゲン応答性に重要かどうかを解析した。まず初めに前述のごとく
Luciferase vector
にG3BP2 ARBS
及びPromoter/
Enhancer
領域をinsert
としてG3BP2 promoter-Luc construct
を作成し、さらに
Motif
解析により同定されたARE
候補領域にmutation
を入れたG3BP2 mutation-Luc construct
を作成した(図7
)。まず
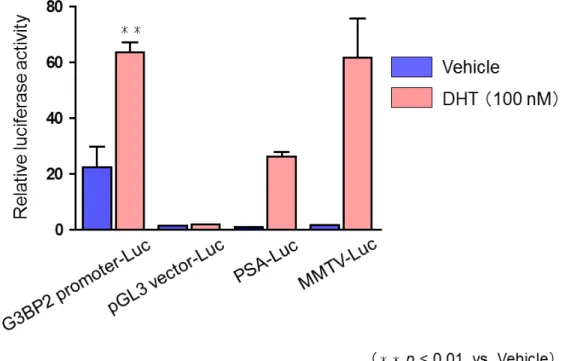
G3BP2 promoter-Luc construct
をLNCaP
細胞にtransfection
しLuciferase assay
にてVehicle
処理及びリガンド処理(DHT 100 nM
)での転写活性化を評価したところ、
Vehicle
処理に比べリガンド処理のもので有 意 な リ ガ ン ド 依 存 的 な 転 写 活 性 化 を 認 め た ( 図8
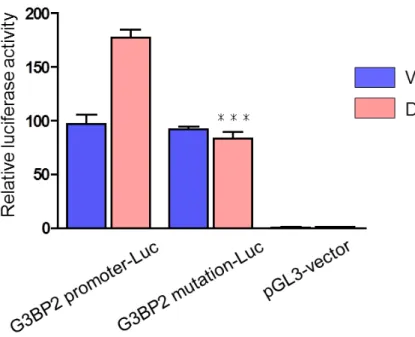
) 。 次 に 、G3BP2 mutation-Luc construct
を 用 い て 同 様 に し て 転 写 活 性 化 を 検 討 し た 。G3BP2 promoter-Luc
ではリガンド依存的な転写活性化が再現性をもって認められ、そのリガンド依存的な転写活性化は
ARE
候補領域にmutation
を入れることにより消失した(図9
)。また、
Electrophoresis Mobility Shift Assay
(EMSA
)にてこのARE
候補24
配列と蛋白の結合を解析したところ、リガンド刺激でより多くの蛋白質と
の結合が認められ、
mutation
を入れた配列ではこの結合の消失を確認した(図
10
左)。さらに、抗AR
抗体とincubation
させることによりAR/ARE
の結合が抗AR
抗体濃度及びincubation
時間に相関して減弱することを確 認しAR
が直接DNA
配列と結合していることが示唆された(図10
右)。以上より
G3BP2 TSS
近傍のARE
候補領域にAR
が結合し転写活性化に重要な役割を担っていることが確認できた。
Ⅲ
G3BP2
のアンドロゲン応答性の検討qRT-PCR
を用いてmRNA
レベルでのアンドロゲン応答性を検討した。Vehicle
処理に比べリガンド処理(DHT 100 nM
)ではリガンド及び時間依存 的 に 有 意 な 増 加 を 認 め ( 図
11
左 ) 、 そ の 増 加 は 抗 ア ン ド ロ ゲ ン 剤(
Bicalutamide
)を用いることにより消失した(図11
右)。次に、同様にしてリガンド処理をした
LNCaP
細胞よりタンパク質を抽出しWestern Blot
法により検討した。するとタンパク質レベルでもリガンド及び時間依存的な増加を認めた(図
12
上)。さらに蛍光免疫染色によりリガ ンド依存的なタンパク質の増加は、細胞内において細胞質有意な局在で高発現していることが認められた(図
12
下)。25
また、別の
AR
陽性前立腺癌細胞株VCaP
細胞を用いて同様にした検討を 行ったところ、LNCaP
細胞と同様にリガンド依存的な転写活性化を認め、mRNA
レベル及びタンパク質レベルでも有意な増加を確認できた(図13
)。以上より
G3BP2
はmRNA
レベル、タンパク質レベルでアンドロゲンにより制御される標的遺伝子であることが示された。
Ⅳ
G3BP2
安定過剰発現細胞株での検討さ ら に
G3BP2
の 前 立 腺 癌 に お け る 機 能 を 解 析 す る た め 前 述 の 通 り に 、Vector
のみをtransfection
し細胞培養して得られた細胞株(Vector #1, #2
)と、
N
末端にFlag
付きのG3BP2
安定過剰発現細胞株(Flag-G3BP2 #1, #2
) を作製した(図14
)。これらの細胞株を用いて
Cell proliferation assay
にてG3BP2
過剰発現の 増殖能へ与える影響について検討したところ、G3BP2
安定過剰発現細胞株 であるFlag-G3BP2 #1, #2
はVector #1, #2
に比べ細胞増殖能が亢進してい た(p < 0.05
)(図15
)。また、同様にCell migration assay
にても有意 差をもって細胞遊走能の亢進を認めた(p < 0.05
)(図16
)。Ⅴ
G3BP2
発現抑制による前立腺癌細胞への影響の検討26
まず初めに前述の
Invitrogen Stealth RNAi #1, #2
の2
種類を用いてG3BP2
を充分に発現抑制できることをLuciferase assay
及びWestern Blot
法により確認した(図
17
)。次にこの
siRNA
を用いてProliferation assay
及びCell migration assay
に て検討したところ、G3BP2
をknock down
することにより細胞増殖能及び 遊走能がともに抑制されることが示された(図18
、19
)。以上より前立腺癌細胞株において
G3BP2
は細胞増殖能及び遊走能の両方 を強く促進的に制御していると考えられた。Ⅵ 前立腺癌臨床検体における
G3BP2
発現解析実際の前立腺癌における
G3BP2
の機能を解析するため、臨床サンプルにおける
G3BP2
蛋白の発現を免疫組織染色により解析した。そのため日本大学医学部附属板橋病院にて前立腺癌の診断で根治的前立腺全摘除術を施行し
た
101
例の臨床検体について抗G3BP2
抗体を用いて免疫組織染色を行い、染色強度を測定し検討した。染色強度の詳細は前述の方法にて行った。す
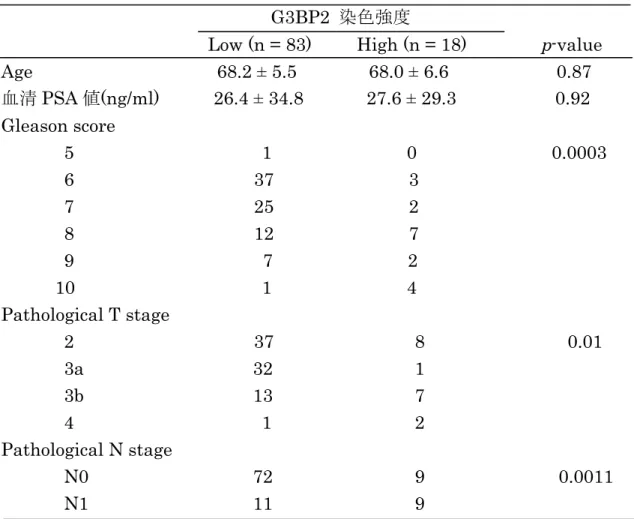
ると
IR score
が7
以上を強発現、6
以下を弱発現で評価したところ、弱発現
83
例、強発現18
例であった。その臨床病理を解析したところpathological
T stage
(p = 0.01
)、リンパ節転移陽性例(p = 0.0011
)、Gleason Score
27
(
p = 0.0003
)と有意に相関した。また、正常前立腺組織ではほとんど染色を得られず、癌で高発現していることが見出された(表
,
図20
)。28
【考察】
Ⅰ
ChIP-chip
法及びChIP-sequence
法による新規アンドロゲン応答遺伝 子の同定本研究では
ChIP-chip
法及びChIP-sequence
法を用いてゲノムワイドにARE
を解析した結果を引用している。東京大学大学院医学系研究科抗加齢医学講座の高山らはこれまでに
ChIP-chip
法によるAR
結合部位の解析結 果を報告している 26-26 。従来のARE
に関する多くの報告は各遺伝子の転 写開始点(TSS
)近傍のプロモーター領域の配列を解析しレポーターアッセ イなどで同定するものであった39 。しかし、これらの方法ではTSS
近傍のAR
結合部位のみの解析であり、ヒトゲノム全領域を解析したものではなかった。今回
ChIP-chip
法及びChIP-sequence
法によりヒトゲノム全領域を 解析しAR
結合部位を同定することに成功している。利点として、標的遺伝 子のプロモーター領域に偏ることなくAR
結合部位を網羅的に同定するこ とが可能であることがあげられる。また、次世代シークエンサーを用いることによって
AR
の結合が強い領域の配列をシークエンスし全ゲノム配列 上のどの位置にAR
結合部位が存在しているか、またターゲット遺伝子のど の位置にARE
が存在するかなどの詳細な情報が得られるようになった。本29
研究により
G3BP2 TSS
近傍(Intron 1
)に1
つのAR
結合部位を同定し、これは新規の
AR
結合部位であった。その部位に対するChIP assay
ではAR
の結合が濃縮して確認され精度の高い検出ができていることが見出された。同定した
AR
結合部位へのAR
の結合により直接的なアンドロゲンに よる転写制御を受け、アンドロゲン応答性にmRNA
及びタンパク質レベル での発現増加が認められ新規アンドロゲン応答遺伝子の発見へと至った。今後ますます
ChIP-chip
法及びChIP-sequence
法が核内受容体ならびに転 写因子の機能解析に応用されていくことが期待される。Ⅱ
G3BP2
の分子機能本研究により
G3BP2
がアンドロゲンによる制御を受けていることを見出 した。G3BP family
はG3BP1, G3BP2a, G3BP2b
の3
つのアイソフォーム が知られているがそれぞれProline rich (PxxP) motif
のsplicing
の程度に より大きく分類されている。G3BP2
はRas GTPase activating protein (SH3 domain) binding protein2
でありRas GAP
にSH3 domain
を介して結合することが知られている。また、
G3BP2
のC
末端にはRNA Recognition Motif
やRGG (arginine-glycine rich) box
などRNA
との結合がよく知られている
domain
やp53
との結合が報告されているdomain
なども認められ30
る 41-43 。このように
G3BP2
は様々なdomain
を有しており癌との関連に関してもいくつかの報告があり近年、癌関連遺伝子としても注目されてい
る37, 44-45 。
G3BP2
はC
末端のdomain
を介して癌抑制遺伝子p53
やMDM2
と直接結合し、
p53
のdegradation
及びubiquitin
化を阻害し、さらにMDM2
自身の
ubiquitin
化も阻害していることが知られている。また、G3BP2
により
p53
の細胞内局在は核内から細胞質へ変化し、その機能は抑制されて いる。さらにG3BP2
の発現を抑制することによりMDM2
は減少し、p53
はタンパク質レベルで増加を認め活性も上昇する 39 。このように多くの報告では
G3BP2
は癌細胞で過剰発現しており、p53
と結合し抑制することで癌細胞の増殖能を亢進させているものと考えられる。また、前立腺癌細胞
におけるドセタキセル感受性は機能的な
p53
により決定されるとの報告も ある46 。さらに臨床検体における前立腺癌細胞のp53
遺伝子変異はゲノム レベルで約40%
との報告もあり前立腺癌の発生や悪性化などに深く関与し ているものと思われる 47 。今回、
p53
と直接的に結合しその活性の制御に関わるG3BP2
が前立腺癌細 胞においてアンドロゲン応答性に発現増加することを初めて報告した。アンドロゲンが
G3BP2
を直接制御し前立腺癌の悪性化に関与している可能 性が示唆される。31
Ⅲ
G3BP2
の臨床病理に関する考察本研究の細胞レベルでの実験結果を元に実際の前立腺癌における機能を解
析するため、日本大学医学部附属板橋病院にて前立腺癌の診断で根治的前
立腺全摘除術を施行した
101
例の臨床サンプルを用いてG3BP2
蛋白の発現 を免疫組織染色により解析した。するとIR score 7
以上を強発現、6
以下を 弱発現として評価したところ、弱発現83
例、強発現18
例であった。さら に臨床病理を解析したところpathological T stage
(p = 0.01
)、リンパ節 転移陽性例(p = 0.0011
)、Gleason Score
(p = 0.0003
)と有意に相関し ていた。これはG3BP2
強発現の臨床サンプルにおいてT stage
が悪い傾向 が認められ、リンパ節転移陽性例が多いことを示している。さらに前立腺癌の悪性度を示す
Gleason score
と強く相関していることから、G3BP2
が 予後因子となる可能性を示唆している。これらの結果は細胞レベルで示された
G3BP2
安定過剰発現細胞株の増殖能の亢進や遊走能の亢進が認められたことと一致する。しかし図
18
、19
で示された結果はリガンド応答性にG3BP2
を介した増殖能・遊走能を直接証明するものではないため、今後の研究課題と言える。
これらより
G3BP2
は何らかのシグナル伝達経路に関与することで前立腺 癌細胞の増殖能や遊走能を亢進していると考えられる。またアンドロゲン32
による直接的な制御を受けていることなどから、前立腺癌に関わるアンド
ロゲン作用の下流シグナルであると考えられる。さらに今回の免疫組織染
色では、正常前立腺組織では
G3BP2
抗体でほとんど染色を得られず、癌で 高発現していることが見出されており、アンドロゲンにより誘導され前立腺癌の発症や進展に働いていることが示唆された。
Ⅳ 今後の研究の展望
(前立腺癌治療標的としての
G3BP2
の応用)これまでの細胞レベルでの機能解析と臨床病理から得られた解析結果から、
G3BP2
はTSS
近傍(Intron 1
)に1
つのARE
をもつ新規アンドロゲン応答遺伝子で、アンドロゲン作用により前立腺癌細胞の増殖能及び遊走能を
亢進させる機能を有する可能性が考えられる。また
G3BP2
を発現抑制する ことにより癌細胞増殖能及び遊走能はそれぞれ抑制され、悪性度の低い癌や正常前立腺組織では発現が低いことなどからも癌細胞において重要な役
割を果たしていることが想定できる。以上より
G3BP2
を治療標的とするこ とで前立腺癌細胞の発症や進展を抑えることができる新しい治療法につながる可能性もある。さらに、
G3BP2
と臨床的予後との関連性やホルモン療 法不応性前立腺癌細胞内でのG3BP2
の役割などを機能解析していくこと33
で新たな臨床マーカーの発見や
CRPC
樹立の機序解明などへつながってい くことも考えられる。今回、
G3BP2
の前立腺癌細胞における役割の一部が初めて報告できた。しかし、実際にどのようなシグナル伝達経路に寄与し、どのような機能を持
っているかは未だに解明されていない点も多く今後の研究課題と言える。
34
【まとめ】
本研究においてヒトゲノムにおける
AR
結合領域データの解析を行い、新規 アンドロゲン応答遺伝子G3BP2
を同定、さらに転写制御に関わるARE
を 同定した。また、in vitro
で前立腺癌細胞の増殖能及び遊走能に深く関与し ていることを明らかにした。前立腺癌臨床検体においては悪性度(Gleason score
)、pathological T stage
分類、リンパ節転移などと相関していた。G3BP2
は様々なシグナル伝達に関与しており、本研究並びにさらなる機能解析により新たな治療標的や診断マーカーの発見などにつながる可能性が
期待される。
35
【謝辞】
本研究の機会を与えてくださり、終始親身な御指導、ご鞭撻を賜りました
日本大学医学部泌尿器科学系泌尿器科学分野主任教授 高橋悟博士に謹ん
で感謝の意を表します。また
1
年半の間、特別研究学生として受け入れ本 研究を行うにあたり、環境を与えて頂き、親身な御指導、貴重な御助言を賜りました東京大学大学院医学系研究科抗加齢医学講座特任教授 井上聡
博士、東京大学大学院医学系研究科抗加齢医学講座特任講師 浦野友彦博
士、東京大学医学部附属病院老年病科 髙山賢一博士、東京大学医学部附
属病院泌尿器科講師 藤村哲也博士、日本大学医学部泌尿器科学系泌尿器
科学分野助教 大日方大亮博士に心から感謝申し上げます。本研究にあた
り細胞培養を初め、各種解析の御指導、ご鞭撻を賜りました東京大学大学
院医学系研究科抗加齢医学講座皆様に深く感謝申し上げます。
本研究をまとめるにあたり、貴重な御助言を賜りました日本大学医学部泌
尿器科学系泌尿器科学分野研究所准教授 山口健哉博士には衷心よりの謝
意を、そして日本大学医学部泌尿器科学系泌尿器科学分野の皆様に深く感
謝申し上げます。
最後に、物心両面にわたり援助をいただいた両親に感謝いたします。
36
【表・図・図説】
表 前立腺癌臨床病理検体を用いた
G3BP2
免疫組織染色(n = 101)
G3BP2
染色強度
Low (n = 83)
High (n = 18)
p-value Age
68.2 ± 5.5 68.0 ± 6.6 0.87
血清PSA
値(ng/ml)
26.4 ± 34.8 27.6 ± 29.3 0.92 Gleason score
5 1 0 0.0003 6 37 3
7 25 2 8 12 7 9 7 2 10 1 4 Pathological T stage
2 37 8 0.01
3a 32 1
3b 13 7
4
1 2 Pathological N stage
N0 72 9 0.0011
N1 11 9
全細胞数のなかで陽性細胞数が何%存在し(
score 0,
陽性細胞なし; score 1, <
1/100; score 2, 1/100 to 1/10; score 3, 1/10 to 1/3; score 4, 1/3 to 2/3; score5, >
2/3
)、染色強度(score 0,
染色されず; score 1,
弱; score 2,
中; score 3,
強)と 合わせてIR score; 0-8
までで評価した。IR score
≧7
を強発現、IR score
≦6
を弱発現とした。37
図
図
11
前前 立立 腺腺 癌癌 診診 療療 アア ルル ゴゴ リリ ズズ ムム「前立腺癌診療ガイドライン
2012
年版」より抜粋・改変38
図
図
22
前前 立立 腺腺 癌癌 細細 胞胞 内内 でで のの アア ンン ドド ロロ ゲゲ ンン 受受 容容 体体 のの 役役 割割39
図
図
33
ゲゲ ノノ ムム ワワ イイ ドド ななA AR R
結結 合合 部部 位位 のの 検検 索索上 図 は
Prostate specific antigen (PSA)
のChromatin
immunoprecipitation (ChIP) -sequence
法の結果である。アンドロゲンを 付加することにより活性型ヒストンの修飾が認められ、エンハンサー・プ ロモーター領域に多くのAR
の結合が認められる。同様にしてゲノムワイド にアンドロゲン応答配列を同定している。40
図
図
44
直直 接接 的的 ななA AR R
のの 制制 御御 をを 受受 けけ るる 遺遺 伝伝 子子 のの 同同 定定41
図
図
55
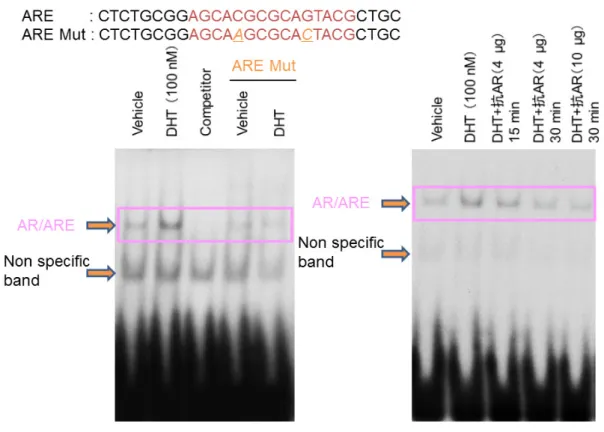
アア ンン ドド ロロ ゲゲ ンン 応応 答答 遺遺 伝伝 子子G G33B BPP22
のの 同同 定定( 上 段)
UCSC genome browser
(hg18
) 上 で のG3BP2
の 位 置 。ChIP-sequence
法によりG3BP2 TSS
近傍にリガンド処理によりAR
の結 合の増加が認められ1
つのARBS
が同定された。(下段)LNCaP
細胞にVehicle
もしくはDHT (10 nM )
処理を行いAR
抗体によりChIP assay
を 行った。濃縮度はqRT-PCR
により定量化しinput
に対する濃縮率を示した。42
図
図
66
G G33B BPP22 TTrraannssccrriippttiioonn SSttaarrtt SSiittee ((TTSSSS))
近近 傍傍A AR RB BSS
内内及
及 びび 周周 囲囲 のの
m moottiiff
検検 索索G3BP2 TSS
下 流645bp-1051bp
にAndrogen Receptor Binding Site
(ARBS)
を認め、内部に1
つのAndrogen Response Element (ARE)
を同 定。また、周囲にOct1
やHNF3
(FOXA1
)などの前立腺癌で知られるAR
関連転写因子群の結合予想配列が多く認められる。43
図
図
77
G G33B BPP22 pprroom mootteerr,, m muuttaattiioonn--LLuucc ccoonnssttrruucctt
のの 作作 製製(上段)
ARBS
を含むG3BP2 enhancer/promoter
領域をinsert
したG3BP2 promoter-Luc
を作製。さらにARBS
内に認められた1
つのARE
候補領域 に2
カ所のmutation
を入れたG3BP2 mutation-Luc
を作製した。(下段)公共データベース(
Transfac
)で公表されているARE consensus
配列。44
図
図
88
G G33B BPP22 A AR RE E
のの リリ ガガ ンン ドド 依依 存存 的的 なな 転転 写写 活活 性性 化化LNCaP
細胞に図7
で作製したconstruct
をtransfection
しそれぞれVehicle
及びDHT (100 nM)
処理し転写活性化を検討した。pGL3 vector-Luc
はnegative control
として、PSA
及びmouse mammary tumor virus (MMTV)
-Luc
はpositive control
として使用した。45
図
図
99
G G33B BPP22 m muuttaattiioonn--LLuucc
のの 転転 写写 活活 性性 化化 能能 のの 消消 失失図
8
と同様にLNCaP
細胞に図7
で作製したG3BP2 promoter-Luc
及びG3BP2 mutation-Luc construct
をそれぞれtransfection
しVehicle
及びDHT (100 nM)
処理し転写活性化を検討した。pGL3 vector-Luc
はnegative
control
として使用した。46
図
図
1100
G G33B BPP22 A AR RE E
のの 同同 定定(左図)
Vehicle
及びDHT (100 nM)
処理48
時間のLNCaP
細胞から核内 タンパク質の分離抽出を行い、それぞれARE, ARE mutation
オリゴ核酸と 室温で15
分間incubation
しGel shift assay
を施行した。(右図)リガンド処理
LNCaP
細胞から抽出した核内タンパク質とオリゴ核酸に加え抗AR
抗体の濃度をそれぞれ
4, 10 (µg)
及びincubation
時間を15, 30
分間として 調整し施行した。47
図
図
1111
G G33B BPP22 m mR RN NA A
レレ ベベ ルル でで のの リリ ガガ ンン ドド 応応 答答 性性 発発 現現 定定 量量 解解 析析(左図)
LNCaP
細胞にVehicle
及びDHT (100 nM)
処理し、処理後各時間 でのmRNA
レベルの発現をqRT-PCR
を用いて定量化した。Vehicle
処理 に対するリガンド処理での発現増加率で示した。(右図)Vehicle
処理及びDHT (100 nM)
処 理 単 独 、 さ ら にDHT
処 理 に 抗 ア ン ド ロ ゲ ン 剤Bicalutamide
付加(それぞれ1 µM, 5 µM
)処理し24
時間後に定量解析を 施行した。48
図
図
1122
G G33B BPP22
タタ ンン パパ クク 質質 レレ ベベ ルル でで のの リリ ガガ ンン ドド 応応 答答 性性 発発 現現 解解 析析(上図)
LNCaP
細胞にVehicle
及びリガンド処理0, 24, 48
時間後にタンパ ク質を回収しwhole cell lysates 30 (µg)
でWestern Blot
法による発現解析 を行った。(下図)LNCaP
細胞を用いて同様にしてリガンド処理し蛍光 免疫染色を施行した。G3BP2
特異抗体により反応させ蛍光に標識された2
次抗体を用いて局在を示した。DAPI
により核染色を行った。49
図
図
1133
別別 ののA AR R
陽陽 性性 前前 立立 腺腺 癌癌 細細 胞胞 株株 ((V VC CaaPP
細細 胞胞 )) でで ののG
G33B BPP22
アア ンン ドド ロロ ゲゲ ンン 応応 答答 性性 のの 検検 討討細胞株を別の
AR
陽性前立腺癌細胞株VCaP
細胞にして図9, 11, 12
と同様 の条件にてアンドロゲン応答性を検討した。50
図
図
1144
G G33B BPP22
安安 定定 過過 剰剰 発発 現現 細細 胞胞 株株 ((SSttaabbllee
細細 胞胞 )) のの 作作 製製G418 (500 µg/ml)
にてselection
を行い、Flag
付きG3BP2
安定過剰発現細 胞株を2
株作製した。(上図)qRT-PCR
によりmRNA
レベルでのG3BP2
の発現を解析した。(下図)Western Blot
法によりタンパク質レベルでのG3BP2
の発現の解析を行った。51
図
図
1155
G G33B BPP22
安安 定定 過過 剰剰 発発 現現 細細 胞胞 株株 のの 細細 胞胞 増増 殖殖 能能 解解 析析Cell proliferation assay
(MTS assay
)を用いてG3BP2
安定過剰発現細胞及び
Vector
コントロール細胞の細胞増殖能を検討した。96 well
にそれぞれの細胞株を
3×10
3 個ずつ散布した(N=4
)。0, 24, 48, 72
時間後にMTS
assay
を行い、吸光度を測定した。52
図
図
1166
G G33B BPP22
安安 定定 過過 剰剰 発発 現現 細細 胞胞 株株 のの 遊遊 走走 能能 解解 析析G3BP2
安定過剰発現細胞及びVector
コントロール細胞をそれぞれ5×10
4 個ずつ24 well
に散布し24
時間培養しCell migration assay
を用いて解析 した。メタノール固定後ギムザ染色しpore
を通過した細胞数を蛍光顕微鏡 にてランダム5
視野で評価しG3BP2
安定過剰発現細胞の遊走能を解析、定量化した。
53
図
図
1177
ssiiG G33B BPP22
にに よよ るる 発発 現現 抑抑 制制 効効 率率 のの 検検 討討LNCaP
細胞に対して各siRNA (10 nM)
になるようにtransfection
施行し、48
時間後にRNA
及びタンパク質を回収、qRT-PCR, Western Blot
法によ り発現を解析した。54
図
図
1188
G G33B BPP22
発発 現現 抑抑 制制 にに よよ るる 細細 胞胞 増増 殖殖 能能 へへ のの 影影 響響図
15, 17
と同様の条件でLNCaP
細胞をsiG3BP2 2
種類で処理しcell
proliferation assay
にて検討した。55
図
図
1199
G G33B BPP22
発発 現現 抑抑 制制 にに よよ るる 細細 胞胞 遊遊 走走 能能 へへ のの 影影 響響図
18
の増殖能assay
と同様に処理して遊走能についてはcell migration
assay
にて定量化し検討した。56
図
図
2200
前前 立立 腺腺 癌癌 臨臨 床床 検検 体体 にに おお けけ るるG G33B BPP22
発発 現現 解解 析析日本大学医学部附属板橋病院にて前立腺癌の診断のもと前立腺全摘除術を 施行した
101
例について、anti-G3BP2 antibody
(1
:100
希釈)を用いた 免疫組織染色を施行した。Gleason score
別及びBenign
の免疫組織染色の 例を示した。Scale bar: 50 (µm)
57
【引用文献】