博士論文
炎症性腸疾患の病態に基づいた
5-アミノサリチル酸の輸送制御機構に関する研究
(Study on transport mechanisms of
5-aminosalicylic acid based on the pathology of
inflammatory bowel disease)
2020 年 9 月
立命館大学大学院薬学研究科
薬学専攻博士課程
立命館大学審査博士論文
炎症性腸疾患の病態に基づいた
5-アミノサリチル酸の輸送制御機構に関する研究
(Study on transport mechanisms of
5-aminosalicylic acid based on the pathology of
inflammatory bowel disease)
2020 年 9 月
September 2020
立命館大学大学院薬学研究科
薬学専攻博士課程
Doctoral Program in Pharmacy
Graduate School of Pharmacy
Ritsumeikan University
由利 龍嗣
YURI Tatsushi
研究指導教員:藤田 卓也 教授
Supervisor:Professor FUJITA Takuya
目次
略語一覧 -1- 総論の部 緒言 -3- 第1 章 モノカルボン酸トランスポーターを介した 5-ASA の大腸における輸送特性に 関する検討 -5- 第1 節 モノカルボン酸トランスポーターを介した 5-ASA 輸送評価 -7- 1-1 マウス消化管におけるモノカルボン酸トランスポーターの発現評価 1-2 Nicotinate の輸送特性とモノカルボン酸による阻害効果の検討 1-3 Nicotinate 輸送におよぼす 5-ASA の影響 1-4 5-ASA のマウス大腸組織での輸送特性評価 1-5 MCTs 阻害剤の 5-ASA の輸送におよぼす影響 1-6 5-ASA の輸送に対するモノカルボン酸の阻害効果 1-7 5-ASA の濃度依存的な輸送特性評価 1-8 Na+ 濃度依存的な 5-ASA 輸送特性評価 1-9 5-ASA 輸送におよぼす ibuprofen の阻害効果 第2 節 考察 -14- 第2 章 PEPT1 を標的とした 5-ASA 誘導体の代謝・輸送特性に関する検証 -18- 第1 節 5-ASA 誘導体の合成 -20- 第2 節 薬物代謝酵素による 5-ASA 誘導体の分解特性に関する検討 -21- 2-2-1 Caco-2 細胞における薬物代謝酵素の活性評価 2-2-2 Caco-2 細胞における 5-ASA 誘導体の代謝特性 第3 節 5-ASA 誘導体の PEPT1 を介した輸送特性評価 -23- 2-3-1 PEPT1 の基質輸送におよぼす 5-ASA 誘導体の影響 2-3-2 5-ASA 誘導体の取り込みに対する pH の影響 2-3-3 5-ASA 誘導体の輸送に対する Gly-Sar の影響 2-3-4 5-ASA 誘導体の濃度依存的な輸送特性評価2-3-5 Gly-Sar の取り込みに対する 5-ASA 誘導体の阻害形式評価
2-3-6 hPEPT1 発現 two-electrode voltage clamp 法を用いた X. oocytes を介した 5-ASA 誘導体の輸送機構解析 第4 節 TNBS 誘発性大腸炎モデルラットでの 5-ASA 誘導体の薬物動態変動評価 -31- 2-4-1 TNBS 誘発性 IBD モデルラットの病態解析 2-4-2 TNBS 誘発による Val-ASA の血中移行変動 2-4-3 5-ASA および Val-ASA の大腸炎症組織蓄積性の評価 2-4-4 大腸炎症部位からの Val-ASA の全身循環移行性におよぼす Gly-Sar の影響 第5 節 考察 -35- 結論 -42- 謝辞 -44- 論文目録 -45- 実験の部 -46- 第1 章 実験の部 -46- 第2 章 実験の部 -51- 引用文献 -58-
- 1 -
略語一覧
IBD : inflammatory bowel disease
SMCT1 : sodium-coupled monocarboxylate transporter 1 MCT1 : proton-coupled monocarboxylate transporter 1 PEPT1 : oligo-peptide transporter 1
5-ASA : 5-aminosalicylic acid SASP : salazosulfapyridine SP : sulfapyridine
Kdeg : apparent first-order degradation rate constants
t1/2 : degradation half-life
AMC : 7-amino-4-methylcoumarin MCA : 4-methylcoumaryl-7-amide Gly-Sar : glycylsarcosine
IC50 : half maximal (50%) inhibitory concentration
Km : michaelis-menten constant
Vmax : maximum velocity
Ki : inhibition constant
𝐾!.#$% : the concentration of Na+ necessary for half-maximal activation BCS : biopharmaceutics classification system
fMLP : N-formylmethionine-leucine-phenylalanine MDP : muramyl dipeptide
Tri-DAP : L-Ala-γ-D-Glu-meso-diaminopimelic acid BCA : bicinchoninic acid
α-CHCA : α-cyano-4-hydroxycinnamic acid HRP : horseradish peroxidase
HTAB : hexadecyltrimethylammonium bromide MPO : myeloperoxidase
TNBS : 2, 4, 6-trinitrobenzensuflonic acid DSS : dextran sodium sulfate
HEPEs : 2-[4-(2-Hydroxyethl)-a-piperazinyl] ethanesulfonic acid MES : 2-(N-morpholino) ethanesulfonate
- 2 -
Tris : tris(hydroxymethyl) aminoethane AcCN : acetonitrile
PBS : phosphate buffered saline TCA : trichloroacetic acid SDS : sodium dodecyl sulfate
- 3 -
総論の部
緒言
潰瘍性大腸炎やクローン病に大別される炎症性腸疾患 (inflammatory bowel disease; IBD) は、小
腸や大腸に炎症が生じることでびらんや潰瘍ができる疾患である。IBD は、下痢や血便、腹痛や
発熱などの症状を慢性的に繰り返すため、IBD 患者の quality of life (QOL) が極めて悪い。IBD は 厚生労働省から難病指定されている疾患の中で患者数が最多の慢性疾患であり、本邦における潰 瘍性大腸炎の患者数は16 万人以上、クローン病では 3.9 万人以上であることが報告されており、 この患者数は増加の一途を辿っている (1-3)。また、IBD の好発年齢は 10 代後半から 20 代前半 と若くして発症することが統計学的に明らかにされている。IBD の発症メカニズムとして、細菌 や化学物質などの外来因子、ストレスや自己免疫異常などが関与すると考えられているが、未だ 明確な原因は解明されておらず、根治療法も確立されていない。そのため、寛解期を維持するた めに生涯に渡る医薬品の服用が必要とされている。また、IBD 患者は大腸がんの発生リスクが健 康なヒトと比べて有意に高いことからも (4-7)、効率的かつ安全な治療戦略の考案が望まれる。 現在の IBD 治療法として、tumor necrosis factor-α (TNF-α) 抗体であるインフリキシマブやアダ リムマブ、シクロスポリンなどの免疫抑制剤、副腎皮質ステロイド製剤、5-aminosalicylic acid (5-ASA) などを用いた対症療法が行われている。抗体医薬品や免疫抑制剤、副腎皮質ステロイドは 強い抗炎症効果を示す一方で、甲状腺や前立腺などのがんや骨粗鬆症、糖尿病様症状などの発症 に影響することが報告されている (8-13)。そのため、これらの医薬品の使用は、1 日の排便回数が 6 回以上、大部分が血液の血便、37.5℃以上の発熱、ヘモグロビン (Hb) が 10 g/dL 以下の貧血や 赤血球沈降速度が 30 mm/h 以上などの重症な症状の患者に制限されている。一方で、中程度まで の IBD 治療の第一選択薬として、メサラジン (5-ASA) 製剤が寛解導入・維持療法に用いられて いる。5-ASA は、大腸上皮細胞の炎症局所において cyclooxygenase や lipoxygenase の阻害、およ びラジカル捕捉などにより薬効を発揮する (14,15)。5-ASA は小腸において速やかに吸収される ため、pH 7.0 以上で溶解する Eudragit® S でコーティングしたアサコールや時間依存的に崩壊す るエチルセルコースでコーティングしたペンタサ、さらにマルチマトリックスによるコーティン グを施したリアルダなど大腸へ効率的に送達するための製剤が開発・上市されている (16-20)。し かし、5-ASA は大腸内においても高い吸収性を示すことが報告されているため、有効な治療効果 を得るためには、高用量 (2.0 g/day 以上) の投与が求められているが、5-ASA の過量投与は下痢 などの消化器症状に加えて、全身循環への 5-ASA 吸収により発熱やネフローゼ症候群、心機能障
- 4 - 害など多くの副作用を引き起こすことが知られている (21-23)。 5-ASA は生理的条件下では電荷を帯びた状態で存在し (24)、比較的高い水溶性 (12.4 mg/mL) を示す。そのため、消化管で認められる 5-ASA の速やかな吸収は単純拡散のみでは説明すること ができず、トランスポーターの関与が示唆されている (25,26)。よって、消化管でのトランスポー ターを介した 5-ASA の輸送特性を明らかにすることができれば、5-ASA の消化管における高い 膜透過性の要因を解明できると考えられる。さらに、大腸炎症組織に高発現するトランスポータ ーを介して、5-ASA を大腸内の炎症部位特異的に蓄積させる製剤的な工夫を施すことにより、投 与量を減少することができ、副作用発現リスクを低下させることが可能になる。また、5-ASA の 服用量の減少は、IBD 患者のコンプライアンスの向上にもつながることが期待される。コンプラ イアンスが悪い患者では 39% まで寛解維持率が低下するが、寛解期にコンプライアンスを遵守 した場合、2 年後まで高い寛解維持率 (89%) を維持することが可能であり、QOL の向上につなが ることが期待される。 そこで本研究では、5-ASA の消化管内での高い膜透過性のメカニズムを解明するため、大腸に 発現する H+ 共役型あるいは Na+ 共役型モノカルボン酸トランスポーターに着目し、5-ASA 輸 送に対するこれらトランスポーターの関与について検討した。さらに、IBD 発症時に大腸炎症部 位特異的に高発現するペプチドトランスポーターを標的とした薬物送達により 5-ASA の治療効 率を改善できると考え、アミノ酸修飾を施した 5-ASA 誘導体を開発し、その動態特性について検 討を行った。 以下、これら結果について2 章にわたり論述する。
- 5 -
第
1
章 モノカルボン酸トランスポーターを介した
5-ASA
の
大腸における輸送特性に関する検討
潰瘍性大腸炎やクローン病に大別される炎症性腸疾患 (inflammatory bowel disease: IBD) は、細 菌や化学物質などの外来因子、ストレスや自己免疫異常などが発症に関与するとされているが、 未だ明確な発症メカニズムは解明されておらず、根治療法も確立されていない難治性疾患である。 現在、中程度までの IBD 症状の第一選択薬としてメサラジン (5-aminosalicylic acid; 5-ASA) が、 経口投与あるいは注腸投与により使われている。5-ASA は消化管の上皮細胞内で cyclooxygenase の阻害や lipoxygenase の阻害、またラジカル捕捉などにより抗炎症作用を示す (14,15)。5-ASA に ついて、その物理的性質に着目すると、5-ASA は NH2 基と COOH 基を持ち、生理的条件下では
COOH 基が解離し、電荷を帯びた状態で存在する (24)。そのため、5-ASA は比較的高い水溶性を 有する化合物であり、biopharmaceutics classification system (BCS) 分類において膜透過の低い化合 物に分類され、脂質二重膜である細胞膜などへの分配は低いことが知られている (24,27)。それに も関わらず、5-ASA は小腸で速やかに吸収されるため (17)、小腸からの吸収を抑制し、大腸へ選 択的に 5-ASA を送達するための放出制御型の製剤的工夫が施された 5-ASA 製剤が開発された (18-20)。しかし、大腸においても 5-ASA は良好な吸収性を示すことから、大腸炎症局所での十分 な薬物濃度を維持するためには投与量を増加する必要があり、それに伴う発熱や心機能障害、ネ フローゼ症候群などの全身性の副作用の発現が問題となっている (21-23)。この高い 5-ASA の吸 収は、単純拡散による膜透過だけではなく、トランスポーターなどの能動輸送の関与が考えられ る。この輸送メカニズムを明らかにすることで、5-ASA を有効かつ安全に使うための治療戦略を 立案することが可能になりうる。 消化管は、組織により役割が大きく異なり、小腸はグルコースやアミノ酸などの栄養吸収に関 与し、大腸は水分の吸収を担う。さらに、消化管は体内へ異物の進入を防ぐ排泄機構としての役 割も担っている。これらの機能を果たすために、消化管には様々なトランスポーターの発現が確 認され、栄養素の吸収や生体異物の排泄のみならず、医薬品の吸収にも大きく影響している。 5-ASA の膜透過に影響する可能性があるトランスポーターとして、モノカルボン酸トランスポータ ーが考えられる。モノカルボン酸トランスポーターは solute carrier (SLC) family である H+ 共役
型モノカルボン酸トランスポーター (SLC16A; MCTs) および Na+ 共役型モノカルボン酸トラン
スポーター (SLC5A; SMCTs) の 2 種類が同定されている。MCTs は主に 14 種類のサブタイプに 分類され、中でも MCT1−MCT4 は機能的に明らかにされている (28-31)。MCTs は low-affinity ト ランスポーターであること (Km values : millimolar range) が知られており、また、それぞれのサブ
- 6 -
タイプにより発現組織が異なる。MCT1 は全身に発現する一方で、MCT2−4 の発現は限局化され ている。MCT2 の発現場所は精巣や肝臓、腎臓、骨格筋、心臓であり、MCT3 は網膜上皮細胞や
胎盤や腎臓に局在する。さらに、MCT4 は白色筋や白血球、脳、腎臓、胎盤、小腸に発現が報告
されている (27,32,33)。また、MCT1 は apical および basolateral membrane に発現するが、MCT2−4 の発現は basolateral membrane に限局している。一方、 Na+ 共役型モノカルボン酸トランスポー
ターは SMCT1 と SMCT2 の 2 種類のサブタイプに大別される。これら SMCTs は細胞膜に発現 するトランスポーターとしてニコチン酸やピルビン酸、またプロピオン酸や酪酸などの短鎖脂肪 酸 (short-chain fatty acids; SCFAs) 輸送に関与する (34-36)。SMCT1 は、脳や甲状腺、すい臓、前 立腺、肺や大腸などを含む様々な組織に発現し、プロピオン酸や酪酸などの基質と高い親和性を 示す (Km = ~130 μM) (37,38)。一方で、SMCT2 の発現は、腎臓や小腸に限局しており、SMCT1 と 比較して基質との親和性は低く、SCFAs 輸送の Km 値は 10 mM 以上を示すことが報告されてい る (39,40)。これらトランスポーターの中で MCT1 と SMCT1 が大腸上皮細胞の管腔側に発現し ており、主に SCFAs の吸収を担う。吸収された SCFAs は免疫寛容やがん抑制因子、エネルギー 源として機能している (41,42)。MCT1 や SMCT1 はモノカルボン酸骨格をもつ生理活性物質や SCFAs を輸送するだけでなく (34,35)、5-ASA の構造類似体であるサリチル酸もこれらにより輸 送されることが報告されている (43-45)。さらに、hSMCT1 を発現した Xenopus laevis oocytes (X. oocytes) を用いた評価システムにより 5-ASA が SMCT1 の基質として認識されることも明らか にされており (43)、これらトランスポーターが消化管内での高い 5-ASA 輸送に関与しているこ とが考えられる。しかし、SMCT1 はがん細胞においてアポトーシスを誘発するため (46,47)、細 胞株を用いた SMCT1 の輸送特性について評価した研究は行われておらず、さらに、大腸での機 能性についてもほとんど検討されていない。 そこで本章では、5-ASA の高い膜透過性に影響する要因を明らかにするため、マウス消化管に 発現する MCT1 および SMCT1 に着目して、マウス摘出大腸における 5-ASA の輸送特性を検討 した。
- 7 -
第
1
節 モノカルボン酸トランスポーターを介した
5-ASA
輸送評価
1-1 マウス消化管におけるモノカルボン酸トランスポーターの発現評価 消化管におけるトランスポーターや代謝酵素の発現量は部位により大きく異なることが知られ ている (48-53)。まず、5-ASA 輸送に関与する可能性があるモノカルボン酸トランスポーターとし て SMCT1 と MCT1 の遺伝子発現の解析を行った。SMCT1 は小腸上部の十二指腸では発現が認 められなかったが、空腸、回腸および大腸において発現が認められた (Fig. 1)。一方で、MCT1 は 小腸および大腸の消化管全体での発現が確認された (Fig. 1)。Figure 1. RT-PCR analysis of mSMCT1/SLC5A8, mMCT1/SLC16A1 and GAPDH in mouse intestinal tissues. The total RNA (1.0 μg) was isolated from the mucous membrane of mouse intestinal tissues and
reverse-transcripted into cDNA and subjected to PCR using their transporters specific primers. mRNA expression SMCT1, MCT1 and GAPDH in Duodenum (Duo), jejunum (Jej), ileum (Ile), cecum (Cec), proximal colon (PC) and distal colon (DC) were prepared from mouse gastrointestinal tract. The RT-PCR products were analyzed by 1% agarose gel electrophoresis.
1-2 Nicotinate の輸送特性とモノカルボン酸による阻害効果の検討
マウス大腸に発現が認められた SMCT1 および MCT1 の輸送活性について検討するため、 SMCT1 および MCT1 双方の基質である nicotinate のマウス摘出大腸における輸送特性を評価 した。まず、SMCT1 による輸送特性に関して評価するため、MCTs の特異的な阻害剤である α-cyano-4-hydroxycinnamic acid (α-CHCA) (5 mM) の存在下での [3H]nicotinate 輸送について検討し
た (Fig. 2) (29,44)。SMCT1 は Na+ と基質を共輸送するトランスポーターであるため、マウス摘
出大腸の nicotinate 取り込みに対する Na+ の影響を評価した結果、[3H]nicotinate 輸送は Na+ 存
在下において取り込み量が有意に増加し、Na+ 依存性が認められた (Fig. 2A)。また、マウス摘出
大腸への Na+ 依存的な nicotinate 取り込みは、5 分まで直線性が認められたため、以降の SMCT1
MCT1 GAPDH
- 8 -
[3H]nicotinate を用いた輸送特性評価は、取り込み時間を 5 分に設定して行った。次に、 SMCT1
を介した輸送に nicotinate や L-lactate などの基質および 5-ASA が影響を及ぼすか評価した (Fig. 2B and C)。マウス摘出大腸への Na+ 依存的な [3H]nicotinate の取り込みは、過剰量の
nicotinate や L-lactate、5-ASA の存在下において有意に減少した (Fig. 2B)。さらに、Na+ 依存的な
[3H]nicotinate の輸送は 5-ASA の濃度依存的に阻害され、その IC
50 値は 2.8 ± 1.0 mM であるこ
とが算出された (Fig. 2C)。これらの結果から、マウス摘出大腸において SMCT1 は機能性を維持 し、SMCT1 を介した基質輸送に 5-ASA が影響を及ぼすことが示唆された。
Figure 2. Transport property of [3H]nicotinate into murine colonic mucosa. (A) Time course of
[3H]nicotinate uptake in mouse colonic mucosa. Uptake of [3H]nicotinate (0.040 μM radiolabeled plus 40
μM unlabeled nicotinate) was measured in the absence (▲) or presence (●) of Na+ at pH 7.4. In Na+ free
buffer, the NaCl in the uptake buffer was replaced with LiCl. (B) Inhibitory effect of nicotinate, L-lactate and
5-ASA on Na+-dependent [3H]nicotinate uptake. Na+-dependent uptake of [3H]nicotinate was measured in
the presence of 10 mM nicotinate, L-lactate and 5-ASA. (C) Inhibition of Na+-dependent nicotinate uptake
by 5-ASA in mouse colonic mucosa. Uptake of [3H]nicotinate (0.040 μM radiolabeled plus 40 μM unlabeled
nicotinate) was measured with a 5-min incubation in the NaCl- or LiCl-containing buffer at pH 7.4 over the 5-ASA concentration range of 0-30 mM. All uptake studies were carried out in the presence of 5 mM α-CHCA, which is a MCT inhibitor. Na+-dependent uptake of nicotinate was obtained by subtracting the uptake
in the LiCl-containing buffer from the uptake in the NaCl-containing buffer. Results are mean ± SD (n = 5-6). *p<0.05, **p<0.01 and ***p<0.001, compared with the presence of Na+ or Control group.
1-3 Nicotinate 輸送におよぼす 5-ASA の影響
前項より、マウス摘出大腸における [3H]nicotinate 輸送に Na+ 依存性が認められること、さら
に、SMCT1 を介した [3H]nicotinate の輸送は 5-ASA により阻害されることが示された。そこで、
5-ASA による [3H]nicotinate 輸送の阻害形式を明らかにするため、濃度依存的な [3H]nicotinate の
輸送に及ぼす 5-ASA の影響を評価した。マウス大腸上皮細胞への Na+ 依存的な [3H]nicotinate Time (min) 0 2 4 6 8 10 [ 3H]Nicotinate upta ke (pm ol/mg-wet tiss ue) 0 5 10 15 20 Na +-d ep en d en t n ic ot in at e u p tak e (p m ol /m g -w et t is s u e/ 5 m in ) 0 2 4 6 8 10 Co
ntrol + Nicotin
ate + L-L ac tate + 5-A SA [5-ASA] (mM) Na +-d ep en d en t n ic ot in at e u p tak e (p m ol /m g t is s u es /5 m in ) 0 5 10 15 0 0.01 0.1 1 10 100 * ** ** ** ** *** A B C
- 9 -
の取り込みには飽和性が認められ、その Km 値は、0.99 ± 0.064 mM であり Vmax 値は 107 ± 2.1
pmol/mg tissue/5 min であった。一方で、5 mM の 5-ASA を添加することにより [3H]nicotinate 輸
送の Km 値が 3.4 ± 0.22 mM に低下したが、Vmax 値には変化が認められなかった (103 ± 3.4
pmol/mg tissue/5 min)。これら結果から、5-ASA は SMCT1 を介した Na+ 依存的な [3H]nicotinate
輸送を競合的に阻害する可能性が示された。さらに、Eadie-Hofstee plot は 1 本の直線性 (r2 = 0.889)
を認めたことから、マウス摘出大腸への [3H]nicotinate は 1 種類の輸送機構を介し、その輸送は
SMCT1 であることが示唆された。
Figure 3. Transport characteristics of [3H]nicotinate uptake in mice colon. (A) Kinetics of
5-ASA-induced inhibition of Na+-dependent nicotinate uptake in mouse colonic mucosa. The concentration of
nicotinate ranged between 0.04 and 5000 μM in the presence (○) or absence (●) of 5 mM of 5-ASA (B). Eadie-Hofstee plot analysis of data (A). The study were carried out in the presence of 5 mM α-CHCA, which is a MCT inhibitor. Na+-dependent uptake of nicotinate was obtained by subtracting the uptake in the
LiCl-containing buffer from the uptake in the NaCl-LiCl-containing buffer. Results are mean ± SD (n = 5-6).
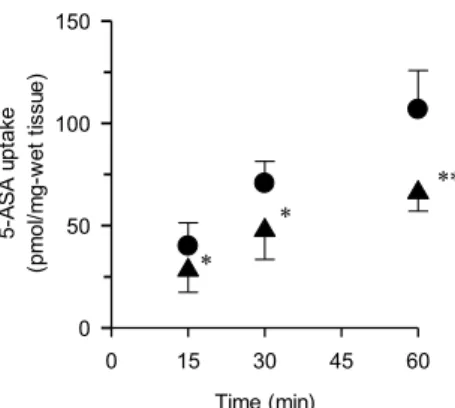
1-4 5-ASA のマウス大腸組織での輸送特性の評価
Fig. 2 および 3 の結果から、大腸に発現する SMCT1 を介した基質輸送を 5-ASA が競合的に
阻害することが示唆されたため、次に 5-ASA がマウス大腸に発現する SMCT1 の基質として認 識されるか検討を行った (Fig. 4)。5-ASA の輸送は、Na+ 存在下において有意に増加し、Na+ 依存
性が認められたことから、マウス大腸に発現する SMCT1 が 5-ASA 輸送に関与する可能性が示 された。また、Na+ 依存的な 5-ASA 輸送は 60 分まで直線的な増加が認められた。そこで、取り 込みの直線性、およびNa+ 依存性が認められた時間である 30 分を以降の検討における 5-ASA 輸 送のインキュベート時間とした。 [Nicotinate] (mM) 0 1 2 3 4 5 Na + -d epe nde nt nicot inat e u pta ke (p mo l/m g-wet tissue /5 min ) 0 30 60 90 120 Uptake/[nicotinate] 0 30 60 90 120 Uptake 0 30 60 90 120 B A
- 10 -
Figure 4, Time course of Na+-dependent 0.5 mM 5-ASA uptake. 5-ASA uptake was carried out at pH 7.4
for the various incubation period in the absence (♦) or presence (●) of Na+. After the incubation for various
incubation period at 37℃, mucous membrane was isolated and the amounts of 5-ASA extracted from the colon tissues were measured by HPLC. Each point represents the mean ± SD (n = 5-6). *P<0.05, **P<0.01, compared with presence of Na+.
1-5 MCTs 阻害剤の 5-ASA の輸送におよぼす影響 大腸上皮細胞には、モノカルボン酸トランスポーターとして SMCT1 だけでなく MCT1 が発 現していることが報告されている (28,32)。実際に、MCT1 が大腸に発現していることを mRNA レベルで確認したため (Fig. 1)、マウス摘出大腸における 5-ASA 輸送に及ぼす MCTs の影響を評 価した。MCTs は H+ 共役型のトランスポーターであるため、5-ASA 輸送に与える pH の影響お よび MCTs 阻害剤による阻害効果を検討した。その結果、5-ASA の輸送は pH 7.4 と比較して pH 6.0 の条件下で多く、5-ASA の輸送に pH 依存性が認められた。しかし、pH 6.0 および pH 7.4 の 条件下において、MCTs の特異的阻害剤である α-CHCA による 5-ASA の阻害は認められなかっ た (Fig. 5)。このことから、MCT1 は 5-ASA の輸送に影響しないことが示された。
Figure 5. Effect of α-CHCA on 5-ASA uptake by mouse colon tissues. Mouse colon tissues were incubated
with 0.5 mM 5-ASA in the absence (■) or presence (■) of α-CHCA for 30 min at 37℃, pH 6.0 or 7.4. After the incubation, the amounts of 5-ASA extracted from the colon tissues were measured by HPLC. Each point represents the mean ± SD (n = 5-6).
5 -A S A u pt a ke (p m o l/m g tis su e /3 0 m in ) 0 50 100 150 200 pH6.0 pH7.4 n.s. n.s. Time (min) 0 15 30 45 60 5-A S A u pt ake (p m ol /m g-w et t iss ue ) 0 50 100 150 * ** *
- 11 -
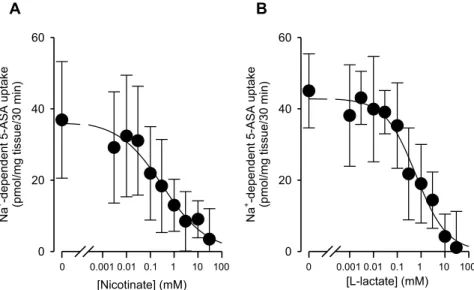
1-6 5-ASA の輸送に対するモノカルボン酸の阻害効果
次に、SMCT1 の基質として知られている nicotinate および L-lactate がマウス摘出大腸におけ る5-ASA 輸送に及ぼす影響について検討した。Na+ 依存的な 5-ASA の大腸上皮細胞への輸送は
nicotinate および L-lactate により濃度依存的に阻害され、その IC50 値はそれぞれ 0.33 ± 0.16 mM
と 0.66 ± 0.11 mM であった (Fig. 6)。
Figure 6. Effect of nicotinate and L-lactate on 0.5 mM 5-ASA uptake by mouse colon tissues. Mouse
colon tissues were incubated for 30 min at pH 7.4, in the various concentration of nicotinate (A) and L-lactate (B). After the incubation for various incubation period at 37℃, mucous membrane was isolated and the amounts of 5-ASA extracted from the colon tissues were measured by HPLC. Each point represents the mean ± SD (n = 5-6).
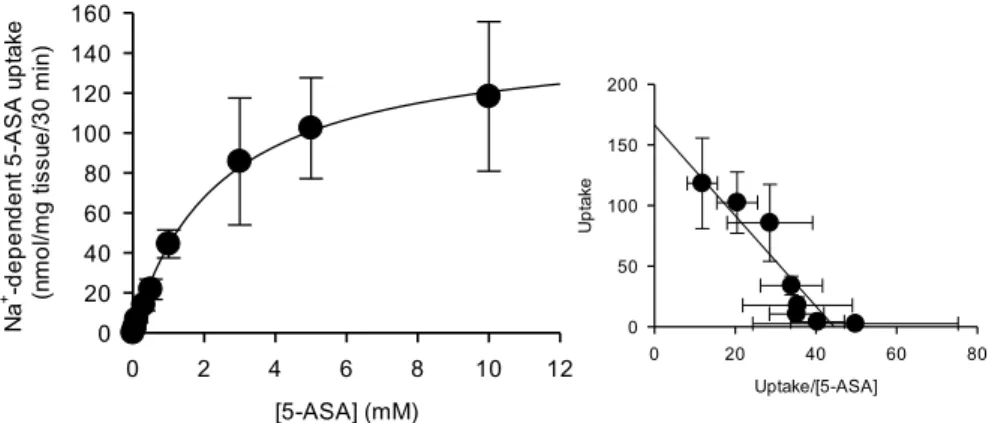
1-7 5-ASA の濃度依存的な輸送特性評価
次に、Na+ 依存的な 5-ASA 輸送の濃度依存性に関して検討を行った。Na+ 依存的な 5-ASA の
輸送には飽和性が認められ、その Km 値は 2.4 ± 0.4 mM、Vmax 値は 149 ± 14.3 nmol/mg tissue/30
min であった (Fig. 7)。さらに、Eadie-Hofstee Plot が直線性を示し、マウス摘出大腸への 5-ASA 取り込みは1 つの輸送機構を介した輸送であることが確認され、その輸送機構は SMCT1 輸送で あることが示唆された。 [Nicotinate] (mM) Na + -dependent 5-AS A uptake (pmol/ mg t issue/30 mi n) 0 20 40 60 0 0.0010.01 0.1 1 10 100 [L-lactate] (mM) Na +-depen dent 5-ASA upt ake (pm ol/ mg ti ssue /30 min) 0 20 40 60 0 0.0010.01 0.1 1 10 100 A B
- 12 -
Figure 7. Concentration dependence of 5-ASA uptake by mouse colon tissues. Nonspecific 5-ASA uptake
was evaluated by measuring in the absence of Na+. The transport of 5-ASA was measured over the
concentration range of 0.05-10 mM. The colon tissues were incubated at 37℃ for 30 min with various concentration of 5-ASA, pH 7.4. Inset, Eadie-Hofsterr plots of the data (A). After the incubation, the amounts of 5-ASA extracted from the colon tissues were measured by HPLC. Each point represents the mean ± SD (n = 5-6).
1-8 Na+ 濃度依存的な 5-ASA 輸送特性評価
SMCT1 は Na+ と基質を共輸送するトランスポーターであり、これまでの検討により 5-ASA
輸送に Na+ 依存性が認められることを明らかにしている。そこで、5-ASA 輸送に及ぼす Na+ 濃
度の影響を評価した (Fig. 8)。5-ASA の輸送は Na+ の濃度依存的に増加が認められ、その Hill 係
数は 2.2 ± 0.75 であった。また、最大取り込み速度 (Vmax) に対する 50% 活性化時の Na+ 濃度
(𝐾!.#$%) は 36 ± 7.1 mM であった (Vmax = 32 ± 3.8 pmol/mg tissue/30 min を用いて標準化した)。これ
らの結果から、5-ASA と Na+ は、 Na+ : 5-ASA = 2 : 1 の stoichiometry で輸送されることが確認
された。 [5-ASA] (mM) 0 2 4 6 8 10 12 Na +-d e p e nd e nt 5 -A S A u p ta ke (n m o l/m g ti ss ue /3 0 m in ) 0 20 40 60 80 100 120 140 160 Uptake/[5-ASA] 0 20 40 60 80 Up ta ke 0 50 100 150 200
- 13 -
Figure 8. Na+ activation kinetics of SMCT1-mediated 5-ASA (0.5 mM) transport. Mouse colon tissues
were incubated with 0.5 mM 5-ASA in the presence of increasing concentrations of Na+ (0-140 mM) for 30
min at 37℃, pH 7.4. Osmolality of the uptake buffer was maintained by iso-osmotic substitution of NaCl with LiCl. The uptake amount in the presence of 140 mM Na+ in mouse colon tissue was taken as 1 to correct
for variations. The inset shows the Hill plot. After the incubation, the amounts of 5-ASA extracted from the colon tissues were measured by HPLC. Each point represents the mean ± SD (n = 5-6).
1-9 5-ASA 輸送におよぼす ibuprofen の阻害効果 最後に、5-ASA が SMCT1 の基質として認識されることを明確にするため、SMCT1 の選択的 な阻害剤として知られる ibuprofen による 5-ASA 輸送に対する阻害効果を検討した (36)。その結 果、5-ASA 輸送は ibuprofen により濃度依存的に阻害され、その IC50 値は 0.19 ± 0.015 mM であ った (Fig. 9)。これらの結果から、マウス大腸に発現が認められる SMCT1 が 5-ASA 輸送に関与 することが明らかにされた。
Figure 9. Effect of ibuprofen on 5-ASA uptake by mouse colon tissues. The colon tissues were incubated
for 30 min with 0.5 mM 5-ASA at pH 7.4 in the presence of various concentration of ibuprofen. After the incubation, mucous membrane was isolated and the uptake amounts of 5-ASA extracted from the colon tissues were measured by HPLC. Each point represents the mean ± SD (n = 5-6).
[Na+] (mM) 0 20 40 60 80 100 120 140 160 V/ Vma x 0.0 0.2 0.4 0.6 0.8 1.0 1.2 Log [Na+] (mM) Log[ v/ (V ma x -V )] -3.0 -2.5 -2.0 -1.5 -1.0 -0.5 0.0 0.5 1.0 1 2 [Ibuprofen] (mM) Na +-dep en de nt 5-ASA up take (pmol /mg t issue /30 mi n) 0 20 40 60 0 0.001 0.01 0.1 1 10
- 14 -
第
2
節 考察
5-ASA が臨床において使用され始めた当初は、スルファピリジン (SP) をアゾ結合により修飾 したプロドラッグであるサラゾスルファピリジン (SASP) として用いられていた。SASP は、腸 内細菌によりアゾ結合が切断され、活性代謝物である 5-ASA を放出し、薬効を発揮する (54)。 SASP は消化管膜透過性が低いことに加えて、breast cancer resistant protein (BCRP: ABCG2) や multidrug resistance-associated protein 2 (MRP2: ABCC2) などの排出トランスポーターの基質として 認識されるため、消化管吸収率は非常に低い (55,56)。そのため、小腸からの吸収を受けず大腸へ 移行し、SASP のアゾ結合が大腸の腸内細菌により切断されるため、大腸に 5-ASA を送達するこ とが可能であった。しかし、代謝副生成物の SP が嘔気やめまい、消化器症状、皮膚障害や男性 不妊症などの副作用をきたすため、現在ではほとんど使用されていない。次いで、5-ASA をアゾ 結合で 2 分子結合した olsalazine (Dipentum) も開発されたが、下痢などの副作用の発現が報告さ れ市場より撤退した。そのような背景もあり、化学的な修飾を施さない 5-ASA そのものを用いた 製剤が着目された。しかし、5-ASA は、単純拡散による膜透過性は低い化合物であるが消化管内 で高い膜透過性を示すため、pH 7.0 以上で溶解するポリマーのメタクリル酸コポリマーS や放出 制御型コーティングであるエチルセルロースなどの大腸デリバリー技術と組み合わせた製剤が開 発され (18-20)、これら 5-ASA 製剤が IBD 治療の第一選択薬として現在用いられている。この ような製剤化により大腸選択的に 5-ASA を送達することは可能であるが、大腸においても 5-ASA は高い膜透過性を示し、全身性の副作用発現が生じている。この高い膜透過性に影響する要因と して、モノカルボン酸トランスポーターの関与が示唆されたため、大腸上皮細胞の管腔側に発現 が認められている SMCT1 と MCT1 に着目し、5-ASA 輸送との関係について検討を行った (32,43)。 SMCT1 は大腸のみならず前立腺や膵臓、肺など多くの正常組織に発現が確認され、ピルビン酸 や乳酸などの生理活性物質やニコチン酸などの水溶性ビタミンの輸送を介して生体の恒常性維持 に関わっている。しかし、がん細胞内においては、ピルビン酸や乳酸、SCFAs などのモノカルボ ン酸化合物は、histone deacetylase やカスパーゼの活性化を阻害し、アポトーシスを引き起こす (41,57)。そのため、がん細胞では、DNA メチルトランスフェラーゼの高発現により SMCT1 プロ モーター領域が高度にメチル化され、 SMCT1 遺伝子の転写が抑制されている (41,42,57,58)。そ こで、SMCT1 を介した基質輸送特性は X. oocytes やがん細胞株に遺伝子導入した強制発現系に おいて評価されてきたが、正常組織での評価はこれまでほとんどされていない。そのため、実際 の組織レベルでの 5-ASA の輸送を評価することは重要である (34,35,43,57,59)。
- 15 -
まず、マウス摘出大腸における nicotinate の輸送特性に関して検討を行った結果、nicotinate の 取り込みに Na+ 依存性が認められ (Fig. 2A)、この nicotinate の輸送は、nicotinate や L-lactate な
どの SMCT1 の基質および 5-ASA により阻害された (Fig. 2B and C)。これらの結果から、SMCT1 を介した nicotinate の輸送を 5-ASA が阻害する可能性が示された。さらに、Na+ 依存的な
nicotinate の取り込みには飽和性が認められ、その Km 値は 0.99 mM が算出された (Fig. 3)。
SMCT1 は、high-affinity および low-capacity トランスポーターであり、X. oocytes やヒト網膜上 皮細胞株である HRPE 細胞に hSMCT1 を発現させた評価系における nicotinate 輸送の Km 値 はそれぞれ 0.23 mM と 0.7 mM であることが報告されているが、本検討でもとめたマウス摘出 大腸における nicotinate の SMCT1 との親和性はこれらの報告より低いものであった (43,59)。一 般に、がん細胞株や X. oocytes に異所性に発現させたタンパク質は、遺伝子を導入した 12~15 時 間後に評価に用いられることが多い。しかし、この遺伝子導入後の処理時間では翻訳後修飾や新 規トランスポーターの発現が生じるには不十分であることが推察され、これがマウス摘出大腸組 織と SMCT1 の発現細胞株とで nicotinate の輸送特性に差が生じた原因であると考えられる (60)。また、nicotinate の濃度依存的な取り込みは 5-ASA を添加することにより Km 値が 0.99 mM から 3.4 mM へシフトすることが認められる一方で、Vmax 値には変化が認められなかった (Fig. 3)。これらの結果から、5-ASA は競合的に nicotinate の輸送を阻害していることが示された。 大腸に発現する SMCT1 を介した基質輸送に 5-ASA が影響を及ぼす可能性が示されたため、 次に、大腸における5-ASA の輸送特性について検討を行った。その結果、マウス摘出大腸におけ る Na+ 依存的な 5-ASA 輸送には飽和性が認められ、その K m 値は 2.4 mM であった (Fig. 7)。
さらに、Fig. 6 および 7 の結果から、Na+ 依存的な nicotinate および 5-ASA 輸送の K
m 値 (0.99
mM、2.4 mM) と互いの輸送に対する Ki 値 (2.1 mM、2.3 mM) の一致が示され、nicotinate と
5-ASA の輸送は同じ輸送機構を介することが示された。さらに、SMCT1 は Na+ と基質を共輸送す
るトランスポーターであり、この輸送の化学量論比は Na+: 基質が 2: 1 であることが算出された。
SMCT1 を介した SCFAs や nicotinate などの輸送の化学量論比も 2: 1 であり、5-ASA の輸送特 性との一致が認められた (43,61)。さらに、この輸送の hill 係数は 2.2 であり、𝐾!.#$% は 36 mM で あることが示された (Fig. 8) これらの結果は X. oocytes を用いた評価系で報告されている 5-ASA の輸送の Km 値 6.5 mM、𝐾!.#$% 54 ± 6 mM および hill 係数 2.0 ± 0.1 と同程度であった (43)。 一方、5-ASA の取り込みは MCTs の特異的阻害剤として知られる α-CHCA による 5-ASA 輸送 の阻害効果は認められず、5-ASA は MCTs の基質として認識されないことが示された (Fig. 5) (44)。ヒト結腸癌由来細胞株 Caco-2 細胞にも MCTs の発現が認められているが、Caco-2 細胞に
- 16 -
5-ASA 輸送に MCTs が関与しないことが強く示唆された (27,45,62)。一方、マウス大腸組織への 5-ASA 取り込みは、pH 7.4 より pH 6.0 の条件下で取り込み量が多かった (Fig. 5)。これはトラン スポーターを介した輸送ではなく、酸性薬物である 5-ASA の pH 分配仮説に従う輸送特性であ ることが推察される (24)。さらに、5-ASA の濃度依存的な輸送の Eadie-Hofstee plot が 1 本の直 線を示したことから、大腸上皮細胞への 5-ASA 取り込みは、1 種類の輸送機構を介していること が示された (Fig. 7)。これらの結果から、消化管に発現するモノカルボン酸トランスポーターの中 で SMCT1 が 5-ASA 輸送に関与する可能性が示された。
そ こ で 次 に 、SMCT1 の 阻 害 剤 で あ る 非 ス テ ロ イ ド 性 酸 性 抗 炎 症 薬 (Non-Steroidal Anti-Inflammatory Drugs: NSAIDs) による、5-ASA 輸送の阻害効果を検討した (36,59,63)。NSAIDs は サリチル酸系やアントラニル酸系、プロピオン酸系に分類され、中でもプロピオン酸系抗炎症薬 である ibuprofen や ketoprofen、fenoprofen、naproxen は SMCT1 の阻害剤として働く。特に ibuprofen は SMCT1 を介した基質輸送を強力に阻害することが報告されている (36)。そこで、 Na+ 依存的な 5-ASA 輸送に及ぼす ibuprofen の影響を評価した。その結果、Na+ 依存的 5-ASA
輸送は、ibuprofen の濃度依存的に阻害され、その IC50 値は約 0.2 mM が算出された。さらに、
過剰量の ibuprofen の存在条件下では、Na+ 依存的な 5-ASA 輸送をほぼ完全に阻害した (Fig. 9)。
これらの結果から、大腸に発現する SMCT1 が ASA 輸送に寄与し、この SMCT1 を介した 5-ASA の取り込みが大腸全体における高い 5-5-ASA の吸収性に関与すると考えられる。 SMCT1 は、nicotinate、L-lactate や SCFAs など MCTs と共通する基質が多く、サリチル酸系抗 炎症薬の中でサリチル酸もこれらの基質として認識される (43,46,47)。一方で、プロピオン酸系に 分類される ketoprofen や naproxen は MCTs の基質であるが、SMCT の阻害剤として機能する (36,64)。SMCT1 と MCTs はアミノ酸配列において相同性は高くないものの、MCTs の基質とし て認識される化合物は SMCT1 の阻害剤として機能する場合もある。そのため、SMCT1 の基質 であるが MCTs の基質として認識されないサリチル酸骨格をもつ 5-ASA は、MCTs を阻害する 可能性が推察される。 また、消化管には SMCT1 以外に Na+ 依存的にモノカルボン酸を輸送する low-affinity、high-capacity トランスポーターである SMCT2 も発現する (39,40)。消化管での SMCT2 の発現は小 腸上部に高発現する一方で、小腸下部に進むにつれて発現量が低下し、盲腸から大腸ではほぼ発 現が認められない (65)。そのため、小腸での速やかな 5-ASA 吸収には SMCT2 が関与しており、 大腸での 5-ASA 輸送は SMCT1 の寄与が大きいことが推察される (Fig. 1) (39)。よって、現行の 5-ASA 製剤は、製剤化工夫により小腸での吸収を抑制することにより投与量は減少しているが、 SMCT1 を介した炎症部位以外からの 5-ASA 吸収を阻害することで、5-ASA による治療効率の更
- 17 - なる向上が可能となりうる。 以上、本章では、大腸に発現する SMCT1 が 5-ASA 輸送に関与することを明らかにした。こ の SMCT1 による輸送が 5-ASA の高い膜透過に影響する一因となり、副作用発現に関与してい ると推察された。そこで、大腸全体に発現する SMCT1 を介した吸収を回避し、IBD 大腸炎症部 位選択的に吸収するトランスポーターを介して 5-ASA を送達することにより、5-ASA の治療効 率を向上できると考え、次章では IBD 大腸炎症部位選択的な 5-ASA 送達を目指した誘導体の開 発に関する検討を行った。
- 18 -
第
2
章
PEPT1
を標的とした
5-ASA
誘導体の代謝・輸送特性
に関する検討
第1 章では、IBD 治療薬である 5-ASA の大腸における輸送特性について検討し、小腸下部か ら大腸にかけて発現する SMCT1 による輸送が 5-ASA の高い膜透過性に関与している可能性を 明らかにした。このような大腸に広く分布するトランスポーターを介した取り込みにより、大腸 内での 5-ASA の分布に炎症局所への選択性がなくなる。その結果、薬物濃度の維持に高い投与量 が必要となり、副作用が発現することが臨床において問題となっている (21-23)。また、5-ASA は、 消化管で高い吸収性を示すため、大腸デリバリーを目的にしたプロドラッグが開発されている。 5-ASA は、NH2 基と COOH 基の 2 つの反応性の官能基を持つため、5-ASA に dextran やcyclodextran、hydroxypropyl cellulose (HPMC)、chondroitin sulfate などの polysaccharide を修飾した 5-ASA プロドラッグがこれまでに考案されている (66)。しかし、IBD の炎症症状は、潰瘍性大腸 炎では大腸全体に炎症が発症する場合もあるが、下行結腸型や直腸型など局所的に炎症を誘発す ることも多く、クローン病では飛び石状病変を示すため、大腸に到達した 5-ASA をさらに大腸炎 症部位選択的に送達する方法の確立が望まれている。 現在、組織選択的に薬物を送達するためのターゲットとして、細胞表面受容体やトランスポー ターが注目され、特に疾患特異的に発現変動が認められるトランスポーターは、薬物の選択的な 送達に有効な標的になることが期待されている。例えば、アミノ酸を輸送する L-type amino acid transporter 1 (LAT1; SLC7A5) や amino acid transporter system B0,+ (ATB0,+; SLC6A14) などは、がん
細胞で発現量が増加することが報告されている。これら発現量が増加するトランスポーターをタ
ーゲットとして、LAT1 の基質であるメチオニンの構造類似体や、グルタミンなどのアミノ酸を
修飾した nanoparticle、あるいは ATB0,+ の基質であるカルニチン修飾 nanoparticle が開発され、
がん細胞選択的に薬物を送達する技術が構築されている (67-69)。IBD 発症時においてもトラン スポーターの発現変動が報告されており、oligo-peptide transporter 1 (PEPT1) や ATB0,+ の発現量
の増加、および P-glycoprotein (P-gp) の発現量の低下が報告されている (70-72)。そのため、IBD 発症時に発現量が増加するこれらのトランスポーターをターゲットにすることで、効率的に薬物 を送達できると考えられる。
PEPT1 は、ヒトの小腸に高発現し、ジペプチド・トリペプチドの吸収に関与している。PEPT1 は、low-affinity および high-capacity トランスポーターであり、ジ/トリペプチド様化合物である β-ラクタム系抗菌薬や angiotensin-converting enzyme (Ace) 阻害剤であるカプトプリルなどを基質 として認識し、これらの薬物を小腸より効率的に吸収する (50,73-75)。この PEPT1 の輸送特性を
- 19 -
利用して、消化管吸収バイオアベイラビリティ (Fa) を改善したプロドラッグである valacyclovir や valganciclovir が開発・上市されている (76,77)。これらプロドラッグは、Fa の低い acyclovir や ganciclovir に valine を修飾することで PEPT1 の基質として認識されるようにしたものであ
り、Fa が改善されたことにより投与量や服用回数を減少できる医薬品として汎用されている。通
常、PEPT1 の発現は大腸にはほとんど認められないが (50,78)、IBD 発症時における大腸炎症組 織特異的に PEPT1 の発現量が増加することが報告されている (70)。そこで、PEPT1 を標的とす ることで、5-ASA を大腸炎症組織選択的に送達できると考えられる。これにより、現行の 5-ASA 製剤の投与量を減少させることができ、副作用発現の回避につながると期待される。
そこで、PEPT1 を標的とした IBD 大腸炎症組織特異的な 5-ASA の送達を目指し、5-ASA の NH2 基あるいは COOH 基に、種々の L-アミノ酸 (glycine、lysine、glutamic acid、valine と tyrosine)
を導入したジペプチド型 5-ASA 誘導体を合成した。これら 5-ASA 誘導体の代謝特性および輸送 特性についてヒト結腸癌由来細胞株Caco-2 細胞および hPEPT1 を発現させた X. oocytes を用い て検討を行った。さらに、5-ASA 誘導体の PEPT1 を介した大腸炎症組織への蓄積性および全身 循環移行特性に関して、IBD モデルラットを用いて検討した。
- 20 -
第
1
節
5-ASA
誘導体の合成
本章において、5-ASA の NH2 基にアミノ酸を導入した 5-ASA 誘導体 (amino acid-ASA
derivatives; Gly-ASA、Lys-ASA、Glu-ASA、Val-ASA および Tyr-ASA)、COOH 基にアミノ酸を導 入した 5-ASA 誘導体 (ASA-amino acid derivatives; ASA-Gly、ASA-Lys、ASA-Glu、ASA-Val およ び ASA-Tyr) を合成した (Summary 1)。
Summary 1. Structure of 5-ASA derivatives and its title method.
5-ASA 誘導体の合成経路について Scheme 1, 2 に示した。
Scheme 1. Synthesis of COOH-terminal conjugates of 5-ASA with various amino acids.
·HCl
(i) (ii) (iii)/(iv)
N H2 CO2Me R1 R1= H = CH(CH3)2 = CH2C6H4OH = CH2CH2CO2Me = CH2CH2CH2CH2NHBoc
Reagents: (i) Boc2O, ET3N, DMF; (ii) WSCD·HCl, HOBt, DMF; (iii) NaOH/MeOH; (iv) 4M HCl/dioxane
N H2 OH CO2H BocHN OH CO2H BocHN OH O NH CO2Me R1 N H2 OH O NH CO2H R R = H (Gly) = CH(CH3)2 (Val) = CH2C6H4OH (Tyr) = CH2CH2CO2Me (Glu) = CH2CH2CH2CH2NH2 (Lys) NH2 OH O OH
5-aminosalicylic acid Amino acid-ASA derivatives ASA-amino acid derivatives NH OH O OH A.A NH2 A.A O OH Glycine Lysine A.A = Glutamic acid
Valine Tyrosine
- 21 -
Scheme 2. Synthesis of NH2-terminal conjugates of 5-ASA with various amino acids.
第
2
節 薬物代謝酵素による
5-ASA
誘導体の分解特性に関する検討
本節では、ヒト結腸癌由来細胞株である Caco-2 細胞に発現する薬物代謝酵素の活性評価およ び 5-ASA 誘導体の酵素的分解特性に関する検討を行った。 2-2-1 Caco-2 細胞における薬物代謝酵素の活性評価 ペプチド代謝に関わる酵素として、エンドペプチダーゼやエキソペプチダーゼがある。本検討 で合成した 5-ASA 誘導体の代謝に関わる酵素として、N 末端のアミノ酸を認識するアミノペプ チダーゼや C 末端のアミノ酸を認識するカルボキシペプチダーゼなどのエキソペプチダーゼが 挙げられるが、Caco-2 細胞にはカルボキシペプチダーゼがほとんど発現していない (79,80)。そこ で、Caco-2 細胞 homogenate におけるアミノペプチダーゼ活性を評価した。各種アミノ酸に 4-methylcoumaryl-7-amide (MCA) を修飾した Arg-MCA、Met-MCA、Ala-MCA、Leu-MCA と Gly-Pro-MCA を用いて aminopeptidase N (APN)、aminopeptidase M (APM)、aminopeptidase B (APB) と dipeptidylpeptidase (DPP IV) の活性を測定した (Table 1)。いずれのアミノ酸修飾体も分解が認めら れ、Caco-2 細胞のペプチダーゼ活性は、APM > APB, DPP IV > APN であることが示された。これ らの結果から、Caco-2 細胞には活性の異なる各種アミノペプチダーゼの発現が確認された。·HCl
(i) Base (ii)
Reagents: (i) Boc2O, ET3N, DMF; (ii) 4M HCl/dioxane; (iii) WSCD·HCl, HOBt, DMF; (iv) H2Pd/C; (v) 4M HCl/dioxane
R2= H = CH(CH3)2 = CH2C6H4OH = CH2CH2CO2Me = CH2CH2CH2CH2NHCbz N H2 OH CO2H BocHN OH CO2H NO2 Br BocHN O O O NO2 NO2 N H2 O O O NO2 NO2 NH CO2H R2 Boc NH O O O NO2 NO2 R2 NH Boc O NH OH O OH R N H2 O ·HCl R = H (Gly) = CH(CH3)2 (Val) = CH2C6H4OH (Tyr) = CH2CH2CO2Me (Glu) = CH2CH2CH2CH2NH2 (Lys)
- 22 -
Table 1. Specific aminopeptidase activities in Caco-2 homogenates.
Peptide-MCA Aminopeptidase aminopeptidase activity (nmol/mg-protein/min) Arg-MCA aminopeptidase B 2.46 ± 0.189 Met-MCA aminopeptidase N 1.89 ± 0.278 Ala-MCA aminopeptidase N 0.91 ± 0.110 Leu-MCA aminopeptidase M 4.45 ± 0.374 Gly-Pro-MCA dipeptidylpeptidase IV 2.49 ± 0.315
10 μM Arg-MCA (APB), Met-MCA (APN), Ala-MCA (APN), Leu-MCA (APM) or Gly-Pro-MCA (DPP IV) was co-incubated with an equal volume of the Caco-2 homogenate (1 mg protein/mL) at 37°C for an appropriate time. These aminopeptidase activity was calculated at liner range. Results are expressed as mean ± SD (n = 4).
2-2-2 Caco-2 細胞における 5-ASA 誘導体の代謝特性
次に、5-ASA 誘導体の代謝特性について Caco-2 細胞 homogenate を用いて検討し、得られた分 解速度定数 (kdeg) と半減期 (t1/2) を Table 2 に示した。いずれの 5-ASA 誘導体も、37℃ の
PBS(−) 中での加水分解は認められなかった。また、ASA-amino acid derivatives は Caco-2 細胞の homogenate と co-incubation することによる代謝も認められなかった。一方で、amino acid-ASA derivatives は Caco-2 細胞 homogenate と co-incubation することにより 1 次反応に従う分解が認 められた。さらに、修飾するアミノ酸により t1/2 が異なり、Lys-ASA の t1/2 が最も短く、約3 分
であった。また、その他の t1/2 は Val-ASA が 30 分、Tyr-ASA が 54 分、Glu-ASA が 136 分で
あり、Gly-ASA が 199 分と最も長い半減期が算出された。
Table 2. Degradation rate constants and half-lifves of 5-ASA derivatives in Caco-2 cell homogenate.
kdeg (min-1) kdeg (min-1) t1/2 (min)
ASA-Gly no degradation Gly-ASA 3.56 ± 0.67 199 ± 38.1 ASA-Lys no degradation Lys-ASA 211 ± 18.5 3.27 ± 0.02 ASA-Glu no degradation Glu-ASA 5.23 ± 1.05 136 ± 25.9 ASA-Val no degradation Val-ASA 22.9 ± 1.27 30.4 ± 1.51 ASA-Tyr no degradation Tyr-ASA 12.9 ± 0.79 53.8 ± 3.38
The Caco-2 homogenate (1 mg protein/mL) was mixed with an equal volume of 10 μM 5-ASA derivatives, and incubated at 37 °C for an appropriate time. The apparent first-order degradation rate constants (kdeg) of
5-ASA derivatives were determined by plotting the logarithm of 5-ASA derivatives remaining as a function time. The degradation half-life was calculated by the following equation: t1/2 = ln2/k. Results are expressed
- 23 -
そこで次に、amino acid-ASA derivatives の代謝生成物に関して解析するため、Val-ASA を Caco-2 細胞 homogenate と co-incubation する前後において、HPLC により分析し、その chromatogram を以下の Fig. 10 に示した。インキュベート前の 5-ASA および Val-ASA のピークは、保持時間 がそれぞれ 2.5 min および 7.4 min に確認された (Fig. 10A and B)。一方、Val-ASA を Caco-2 細 胞 homogenate と 15 分間 co-incubation した際、2 本のピークが認められ、Fig. 10C の矢印が示 す代謝物ピークは、5-ASA と同じ保持時間を示した。そのため、5-ASA 誘導体と Caco-2 細胞を co-incubation することにより、代謝物として 5-ASA が生成されることが示された。
(A) 5-ASA (B) Val-ASA (C) Val-ASA (15 min)
Figure 10. HPLC analysis of degradation of 5-ASA derivatives in Caco-2 cell homogenate.
Chromatogram of 5-ASA (A), Val-ASA (0 min) (B) and incubation with Caco-2 homogenate for 15 min (C). Arrow indicates the peak derived from byproduct of 5-ASA.
第
3
節
5-ASA
誘導体の
PEPT1
を介した輸送特性評価
本節では、5-ASA にアミノ酸を修飾した 5-ASA 誘導体が PEPT1 の基質として認識されるか否 か、PEPT1 の発現が報告されている Caco-2 細胞を用いて検討した。さらに、hPEPT1 遺伝子を 発現させた Xenopus laevis oocytes (X. oocytes) により、5-ASA 誘導体が PEPT1 の基質として認識 されるか直接的な検討を行った。前節で 5-ASA 誘導体が aminopeptidase により代謝を受けるこ とが示されたため、本節の輸送特性評価は、酵素的分解の阻害剤 (protease inhibitor cocktail) の存 在下で行った。
2-3-1 PEPT1 の基質輸送におよぼす 5-ASA 誘導体の影響
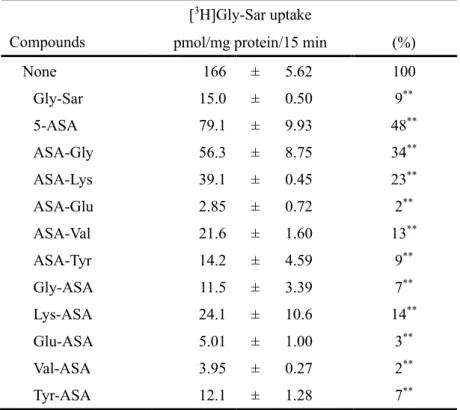
まず、非代謝性のペプチドであり、PEPT1 の基質として知られる glycylsarcosine (Gly-Sar) の Caco-2 細胞における輸送に及ぼす 5-ASA 誘導体の影響を評価した (Table 3)。その結果、[3
- 24 -
示された。また、5-ASA 誘導体は 5-ASA より強力に Gly-Sar の輸送を阻害することが確認され た。特に、ASA-Glu、Gly-ASA、Glu-ASA、Val-ASA および Tyr-ASA を添加することにより、[3
H]Gly-Sar の取り込みは、ほぼ完全に阻害された。これらのことから、5-ASA および 5-ASA 誘導体は PEPT1 の基質輸送に影響を及ぼすことが示された。
Table 3. Inhibitory effects of 5-ASA derivatives on Gly-Sar uptake in Caco-2 cells.
Compounds
[3H]Gly-Sar uptake
pmol/mg protein/15 min (%)
None 166 ± 5.62 100 Gly-Sar 15.0 ± 0.50 9** 5-ASA 79.1 ± 9.93 48** ASA-Gly 56.3 ± 8.75 34** ASA-Lys 39.1 ± 0.45 23** ASA-Glu 2.85 ± 0.72 2** ASA-Val 21.6 ± 1.60 13** ASA-Tyr 14.2 ± 4.59 9** Gly-ASA 11.5 ± 3.39 7** Lys-ASA 24.1 ± 10.6 14** Glu-ASA 5.01 ± 1.00 3** Val-ASA 3.95 ± 0.27 2** Tyr-ASA 12.1 ± 1.28 7**
[3H]Gly-Sar (1 μM) uptake was measured in Caco-2 cells at an extracellular pH of 6.0. Unlabeled compounds
were used at a concentration of 10 mM. Results are expressed as mean ± SD (n = 3). **P<0.01, compared with absence of unlabeled compound.
2-3-2 5-ASA 誘導体の取り込みにおける pH の影響
PEPT1 は内向き H+ 勾配を駆動力として、基質: H+ = 1: 1 で共輸送するトランスポーターであ
る。そのため、異なる細胞外 pH 条件下における 5-ASA 誘導体の輸送特性について評価した。 その結果、すべての 5-ASA 誘導体は pH 7.5 よりも pH 6.0 において取り込み量が多いことが示 され、5-ASA 誘導体の輸送に pH 依存性が認められた (Fig. 11)。また、ASA-amino acid derivatives の輸送は、pH の増加に伴い減少することが認められた。一方で、amino acid-ASA derivatives の取 り込みは、細胞外 pH が 5.0 から 6.5 へ増大することに伴い取り込み量が増加し、さらに高塩基 性条件下では減少する bell 型を示した。これらの結果から、5-ASA 誘導体の輸送に H+ 勾配が
- 25 -
Figure 11. Influence of pH on 5-ASA amino acid derivatives uptake in Caco-2 cells. Uptake of 5-ASA (1
mM) was measured in Caco-2 cells. The uptake was carried out for 15 min in the uptake buffers of varying pH. Values are mean ± SD (n = 4).
pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 50 100 150 200 pH 5 6 7 8 9 up tak e (n m ol /m g pr ot ei n/ 15 m in ) 0 20 40 60 80 pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 10 20 30 40 pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 50 100 150 200 pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 20 40 60 80 100
ASA-Gly ASA-Lys ASA-Glu
ASA-Val ASA-Tyr pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 50 100 150 200 pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 20 40 60 80 100
ASA-Gly ASA-Lys ASA-Glu
ASA-Val ASA-Tyr pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 50 100 150 pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 10 20 30 40 pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 50 100 150 200 pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 100 200 300 400 pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 20 40 60 80 100
Gly-ASA Lys-ASA Glu-ASA
Val-ASA Tyr-ASA pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 100 200 300 400 pH 5 6 7 8 9 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 20 40 60 80 100
Gly-ASA Lys-ASA Glu-ASA
- 26 -
2-3-3 5-ASA 誘導体の輸送に対する Gly-Sar の影響
5-ASA 誘導体は PEPT1 を介した基質輸送に影響を与えること、また、細胞外 pH が 5-ASA 誘 導体の輸送に影響することが明らかとなったため、5-ASA 誘導体が PEPT1 の基質として認識さ れる可能性が示された。そこで次に、5-ASA 誘導体が PEPT1 の基質になることをより詳細に検 証するため、5-ASA 誘導体の取り込みに対する Gly-Sar の阻害効果を検討した。検討の結果、い ずれの ASA-amino acid derivatives の輸送も Gly-Sar による阻害が認められなかった。一方で、 amino acid-ASA derivatives の中で Gly-ASA、Glu-ASA と Val-ASA の取り込みは Gly-Sar の濃度 依存的に阻害され、その IC50 値はそれぞれ 3.4 ± 0.6 mM、4.6 ± 1.5 mM と 10.2 ± 2.1 mM であっ
た。Lys-ASA と Tyr-ASA の輸送に関しては過剰量 (10 mM) の Gly-Sar 存在下においても阻害が 認められなかった。これらの結果から、3 種類の 5-ASA 誘導体 (Gly-ASA、Glu-ASA、Val-ASA) は PEPT1 を介して輸送されることが示唆された (Fig. 12)。
- 27 -
Figure 12. Inhibitory effect of Gly-Sar on the uptake of 5-ASA and 5-ASA amino acid derivatives in Caco-2 cells. Uptake of 1 mM 5-ASA derivatives was measured with a 15-min incubation in the uptake
buffer at pH 6.0 over Gly-Sar concentration range of 0-10 mM. Each point represents mean ± SD (n = 4).
0 0.1 1 10 0 0.1 1 10 0 0.1 1 10 [Gly-Sar] (mM) Up ta ke (n m ol / m g pro te in /1 5 m in ) 0 20 40 60 80 100 0 0.1 1 10 [Gly-Sar] (mM) Up ta ke (n m ol /m g pro te in /1 5 m in ) 0 20 40 60 80 0 0.1 1 10
ASA-Gly ASA-Lys ASA-Glu
ASA-Val ASA-Tyr [Gly-Sar] (mM) up tak e (n m o l/m g p ro te in /1 5 m in ) 0 50 100 150 200 250 [Gly-Sar] (mM) Up ta ke (n m o l/m g p ro te in /1 5 m in ) 0 30 60 90 120 150 0 0.1 1 10 [Gly-Sar] (mM) Up ta ke (n m o l/m g p ro te in /1 5 m in ) 0 10 20 30 40 0 0.1 1 10 0 0.1 1 10
Gly-ASA Lys-ASA Glu-ASA
0 0.1 1 10 0 0.1 1 10 Val-ASA Tyr-ASA [Gly-Sar] (mM) up tak e (n m o l/m g p ro te in /1 5 m in ) 0 30 60 90 120 150 0 0.1 1 10 0 0.1 1 10 0 0.1 1 10
Gly-ASA Lys-ASA Glu-ASA
[Gly-Sar] (mM) Up ta ke (n m o l/m g p ro te in /1 5 m in ) 0 100 200 300 400 500 0 0.1 1 10 0 0.1 1 10 Val-ASA Tyr-ASA [Gly-Sar] (mM) up tak e (n m o l/m g p ro te in /1 5 m in ) 0 30 60 90 120 150 0 0.1 1 10 [Gly-Sar] (mM) Up ta ke (n m ol /m g pro te in /1 5 m in ) 0 20 40 60 80 0 0.1 1 10 0 0.1 1 10 [Gly-Sar] (mM) Up ta ke (n m ol /m g pro te in /1 5 m in ) 0 10 20 30 40 50 0 0.1 1 10 [Gly-Sar] (mM) Up ta ke (n m ol /m g pro te in /1 5 m in ) 0 20 40 60 80 0 0.1 1 10
ASA-Gly ASA-Lys ASA-Glu
ASA-Val ASA-Tyr [Gly-Sar] (mM) up tak e (n m o l/m g p ro te in /1 5 m in ) 0 30 60 90 120 150
- 28 -
2-3-4 5-ASA 誘導体の濃度依存的な輸送特性評価
続いて、amino ASA derivatives の濃度依存的な輸送特性に関して検討を行った。Amino acid-ASA derivatives の中で、Gly、Glu と Val を修飾した 5-acid-ASA 誘導体の輸送には飽和性が認められ、 その Km 値はそれぞれ 1.3 ± 0.1 mM、1.0 ± 0.1 mM と 0.59 ± 0.08 mM であった。一方で、Lys-ASA
と Tyr-ASA の輸送は、濃度の増加に伴い直線的に取り込み量が増加した (Fig. 13)。これらの結果 から、Gly-ASA、Glu-ASA と ASA はトランスポーターを介して輸送され、その親和性は Val-ASA > Gly-Val-ASA > Glu-Val-ASA であることが示された。
Figure 13. Saturation kinetics of various amino acid-ASA derivatives transport in Caco-2 cells. Uptake
of various amino acid-ASA derivatives was measured in Caco-2 cells with a 15-min incubation in the uptake buffer (pH 6.0) over their concentration range of 0.01-10 mM. Each point represents mean ± SD (n = 4).
また、0.5 mM における 5-ASA および amino acid-ASA derivatives の Caco-2 細胞への取り込み 量を比較した結果を Table 4 に示した。その結果、Gly-ASA、Glu-ASA および Val-ASA の取り込 み量は 5-ASA の取り込み量 (110 nmol/mg protein/15 min) と比較して多く、中でも Val-ASA の取 り込みが最も高いことが明らかになった。一方で、Lys-ASA と Tyr-ASA の取り込みは、5-ASA よ り少ないことが示された。 [Gly-ASA] (mM) 0 3 6 9 12 Up ta ke ( nm o l/m g p ro te in /1 5 m in ) 0 200 400 600 [Lys-ASA] (mM) 0 3 6 9 12 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 100 200 300 [Glu-ASA] (mM) 0 3 6 9 12 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 100 200 300 400 500 [Val-ASA] (mM) 0 3 6 9 12 U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 200 400 600 800 [Tyr-ASA] (mM) 0 3 6 9 12 Up ta ke ( nm o l/m g p ro te in /1 5 m in ) 0 100 200 300 400 500
Gly-ASA Lys-ASA Glu-ASA
- 29 -
Table 4. Uptake amounts of 5-ASA and 5-ASA derivatives in Caco-2 cells.
Uptake (nmol/mg protein/15 min) 5-ASA 110 ± 3.76 Gly-ASA 126 ± 2.24 Lys-ASA 6.69 ± 0.36 Glu-ASA 122 ± 9.01 Val-ASA 271 ± 32.2 Tyr-ASA 34.2 ± 5.28
Uptake of 5-ASA (0.5 mM) or 5-ASA derivatives (0.5 mM) was measured in Caco-2 cells with a 15-min incubation in the uptake buffer at pH 6.0. Results are expressed as mean ± SD (n = 4).
2-3-5 Gly-Sar の取り込みに対する 5-ASA 誘導体の阻害形式評価
次に、これまでの検討により、PEPT1 の基質として最も高い親和性を持つことが示唆された Val-ASA について、Val-Val-ASA が Gly-Sar の取り込みに対して及ぼす影響を評価した (Fig. 14)。Caco-2 細胞における Gly-Sar の取り込みは飽和性が認められ、その Km 値は 2.1 ± 0.1 mM、Vmax 値は
315 ± 2.7 nmol/mg protein/15 min であった。一方、0.5 mM の Val-ASA の添加した条件下では Km
値は 10.2 ± 0.6 mM へのシフトが認められたが、Vmax 値は変化しなかった (404 ± 13.7 nmol/mg
protein/15 min)。これらの結果から、Val-ASA は Gly-Sar の取り込みを競合的に阻害することが示 された。また Gly-Sar の取り込みは Eadie-Hofstee plot において一相性の直線関係が示された (Fig. 14A)。また Gly-Sar の取り込みは Val-ASA の濃度依存的に阻害され、Val-ASA の Ki 値は
0.14 mM であった (Fig. 14B)。この値は、Val-ASA の取り込みの Km 値 (0.59 mM) と一致するこ
とから、Val-ASA は Gly-Sar と同じ輸送機構を介した輸送であることが示され、その輸送は PEPT1 である可能性が示された。
- 30 -
Figure 14. Inhibition of Gly-Sar transport by Val-ASA, and kinetics of Val-ASA-induced inhibition of
Gly-Sar transport in Caco-2 cells. Uptake of [3H]Sar (0.06 μM radiolabeled plus 1 μM unlabeled
Gly-Sar) was measured with a 15-min incubation in the uptake buffer (pH 6.0) over Val-ASA concentration range of 0-10 mM (A). The uptake of Gly-Sar was measured with a 15-min incubation in the uptake buffer (pH 6.0) (B). The concentration of Gly-Sar ranged between 0.06 -10 mM in the presence (▵) or absence (●) of 0.5 mM Val-ASA. Inset: Eadie-Hofstee plot analysis of data. Lines were fitted by using linear-regression analysis. Results are mean ± SD (n = 4).
2-3-6 hPEPT1 発現 two-electrode voltage clamp 法を用いた X. oocyte を介した 5-ASA 誘導の
輸送機構解析
最後に、5-ASA 誘導体が PEPT1 の基質となることを明確にするため、hPEPT1 を発現した X. oocytes を用いて、amino acid-ASA derivatives の PEPT1 を介した輸送を直接的に評価した。その 結果、5-ASA 誘導体の中で、Gly-ASA、Glu-ASA と Val-ASA の輸送に内向き電流が認められ、膜 電位を −50 mV に固定し、0.25 mM Gly-Sar 誘発性内向き電流をコントロール (100% = 151 ± 31 nA) としたとき、それぞれの内向き電流は 44 ± 6.2%、56 ± 11%、59 ± 8.6% であった (Fig. 15A)。 また、Lys-ASA および Tyr-ASA の輸送は 10 ± 4.5% と 5.2 ± 3.4% のわずかな起電流が示された。 さらに、Gly-Sar の輸送は、X. oocytes の膜電位の増加に伴い減少が認められたが、amino acid-ASA derivatives の輸送には膜電位の影響がほとんど認められないことが示された (Fig. 15B)。これらの 結果から、5-ASA 誘導体の中で amino acid-ASA derivatives は PEPT1 の基質として認識されるこ とが明らかにされた。また、修飾するアミノ酸により PEPT1 との親和性が異なり、Gly-ASA、Glu-ASA と Val-との親和性が異なり、Gly-ASA、Glu-ASA は PEPT1 に対して高い親和性を示すが、Lys-との親和性が異なり、Gly-ASA、Glu-ASA と Tyr-との親和性が異なり、Gly-ASA、Glu-ASA は、PEPT1 と の親和性は低いことが示された。 [Gly-Sar] (mM) 0 2 4 6 8 10 G ly -S ar u pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0 50 100 150 200 250 300 V/S 0 50 100 150 200 V 0 100 200 300 400 500 A [Val-ASA] (µmol/L) G ly -S ar U pt ak e (n m ol /m g pr ot ei n/ 15 m in ) 0.00 0.05 0.10 0.15 0.20 0.25 0.30 0 1 10 100 1000 B B A

![Figure 2. Transport property of [ 3 H]nicotinate into murine colonic mucosa. (A) Time course of [ 3 H]nicotinate uptake in mouse colonic mucosa](https://thumb-ap.123doks.com/thumbv2/123deta/6578782.1133754/12.892.64.819.383.621/figure-transport-property-nicotinate-murine-colonic-nicotinate-colonic.webp)
![Figure 3. Transport characteristics of [ 3 H]nicotinate uptake in mice colon](https://thumb-ap.123doks.com/thumbv2/123deta/6578782.1133754/13.892.154.749.352.596/figure-transport-characteristics-h-nicotinate-uptake-mice-colon.webp)