第 70 回日本電気泳動学会シンポジウム:電気泳動によって解き明かされる生命現象
論文種目:総合論文
細胞質 DNA によって活性化される自然免疫シグナル
茂 谷 康 *
徳島大学藤井節郎記念医科学センター・細胞情報学分野 (受付 2020 年 12 月 28 日,受理 2021 年 2 月 2 日)SUMMARY
Cytosolic DNA derived from pathogen or host cells activates cyclic GMP-AMP synthase (cGAS) to produce cyclic GMP-AMP dinucleotide (cGAMP). The binding of cGAMP to a downstream adaptor protein, stimulator of interferon genes (STING), induces innate immune response. Here, we reported two useful methods to analyze the cGAS-cGAMP-STING signaling pathway. One is a sensitive in vitro assay to detect cGAMP using clear-native PAGE, based
on its ability to induce STING dimerization. Using this method, we successfully detected cGAMP in the fetal liver and spleen of DNase II deficient mice, which cause a strong inflammatory response that leads to lethal anemia or polyarthritis. Another is an improved proximity-dependent biotin identification (BioID) method, which is a powerful technology to identify protein-protein interactions in living cells. By using the improved BioID method, we identified previously known and unknown interactors of STING, such as several palmitoyl transferases, interferon regulatory factor 5 (IRF5) and interferon-induced transmembrane protein 3 (IFITM3). The STING interactome obtained in this method will provide new insights into the cGAS-cGAMP-STING signaling network.
Key words: cytosolic DNA, innate immune response, cyclic GMP-AMP (cGAMP), stimulator of interferon genes (STING), proximity-dependent biotin identification (BioID)
はじめに
細胞質内 DNA センサー cyclic GMP-AMP synthase(cGAS) は,DNA ウイルスなどの病原体に対する自然免疫応答に 必須の分子である.病原体 DNA を細胞質内で感知した cGASは,GMP と AMP がホスホジエステル結合で連結し た環状ジヌクレオチド cyclic GMP-AMP(cGAMP)を生成 する.そしてこれがセカンドメッセンジャーとして下流ア ダプター分子 stimulator of interferon genes(STING)にシ グナルを伝達し,最終的に I 型インターフェロン(IFN) や炎症性サイトカインの遺伝子発現を誘導する.一方, cGAS-cGAMP-STING経路の異常な活性化は慢性炎症を引 き起こし,様々な病態に関与することが明らかとなりつ つある.例えば,STING をコードする TMEM173 遺伝子 の機能獲得型変異によって,I 型 IFN の恒常的産生を伴う 全身性の血管炎を発症する.このような背景から,cGAS-cGAMP-STING経路の阻害剤開発などの臨床応用研究が加 速し,その基盤となるシグナル伝達経路の制御機構を明ら かにすることが重要な課題となっている.本稿では,細胞 質 DNA によって活性化される cGAS-cGAMP-STING 経路 について,筆者らが開発した電気泳動や質量分析を利用し た解析方法と,それらを用いて明らかとなった病態生理的 役割やシグナル制御因子を紹介する. 高感度 cGAMP 検出法の開発と cGAS-cGAMP-STING 経路の病態生理的役割 cGASは,細胞質内に局在することで,核やミトコン ドリアに格納されている自己の DNA には通常応答しない 仕組みになっている.しかし,外的ストレスや遺伝的要 因で自己 DNA が細胞質内に漏出した場合,cGAS が恒常 的に活性化される可能性が考えられた.このような自己 DNA誘発性の炎症モデルとして DNase II 欠損マウスが知
Innate immune signaling by cytosolic DNA Kou Motani
Division of Cell Signaling, Fujii Memorial Institute of Medical Sciences, Tokushima University
* Corresponding author: Kou Motani; Division of Cell Signaling, Fujii Memorial Institute of Medical Sciences, Tokushima University, 3-18-15, Kuramoto-cho, Tokushima 770-8503, Japan
E-mail: [email protected]
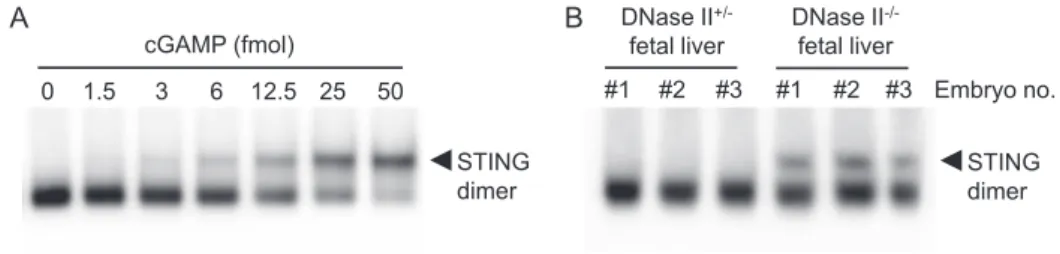
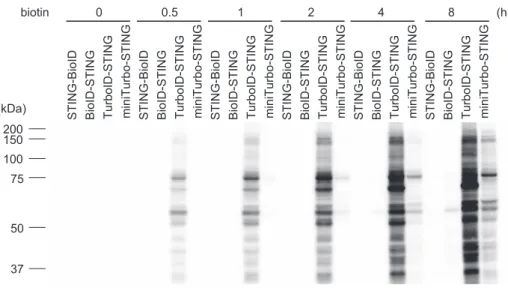
発現させ,約 10 nm 範囲の近接する(相互作用する)タ ンパク質をビオチン標識して同定する技術である3).しか し従来の方法は,①十分なビオチン標識に 16 時間以上を 要するため時間分解能が低い,②ストレプトアビジンに対 する非特異的結合が原因でバックグラウンドノイズが非常 に高い,などの欠点があった.筆者らは,これらの問題を 克服した改良型 BioID 法を構築したので紹介する. ごく最近 Ting らのグループは,従来の BioID 酵素に変 異を導入し TurboID や miniTurbo と呼ばれる高比活性酵素 を 開 発 し た4). そ こ で, こ れ ら の ビ オ チ ン 化 酵 素 と STINGの融合タンパク質を安定的に発現する細胞株を樹 立し,STING 相互作用タンパク質のビオチン化効率をウ エスタンブロットにて評価した.この時,融合タンパク質 が本来の機能を保持していることを確認するため,内在性 STINGを欠失させた細胞を用いた.また,融合位置(N 末端か C 末端)やリンカーの有無,発現量などがビオチ ン標識能に影響するので最適条件を検討しておく必要があ る.Fig. 2 に示すように,BioID-STING 細胞ではビオチン 添加後 8 時間でビオチン化タンパク質がわずかに検出され た一方,TurboID-STING 細胞では 30 分で十分なビオチン 化 が 誘 導 さ れ た5).cGAMP の 代 わ り に 5,6-Dimethyl-9-oxo-9H-xanthene-4-acetic acid(DMXAA)という膜透過性 の合成リガンドを使用し,リガンド刺激依存的に標識され ち,目的の試料と STING タンパク質を試験管内で反応さ せ,CN-PAGE を用いて STING のバンドシフトの程度を 調べることで試料中の cGAMP 量を算出できる(Fig. 1A). また,検出限界がわずか数フェムトモルと非常に高感度で, LC-MS/MS解析に比べて 100 倍以上の感度を示した.な お,ここではマウスの STING を用いており,理由は不明 だがヒトの STING では CN-PAGE における cGAMP 依存 的なバンドシフトが検出されにくいため注意が必要であ る.この検出法を用いて,DNase II 欠損マウスの組織中の cGAMP量を調べたところ,コントロールの野生型または ヘテロ欠損マウスでは全く検出されなかったのに対し,ホ モ欠損マウスの胎仔肝臓では約 20 フェムトモル,脾臓で は約 100 フェムトモルの cGAMP が検出された(Fig. 1B). さらに,cGAS と DNase II の二重欠損マウスを作製し解析 した結果,I 型 IFN 産生量の低下および貧血症状の改善が 認められた.これらの結果は病原体 DNA のみならず自己 DNAが細胞質内に漏出すると cGAS が活性化されること を示し,無菌性炎症における cGAS-cGAMP-STING 経路の 病態生理的意義が明らかとなった. 高時間分解能と低ノイズ化を実現する 新たな BioID 法の構築 小胞体膜タンパク質である STING は,cGAMP と結合
Fig. 1 Quantitative STING dimerization assay for cGAMP detection
The indicated amounts of cGAMP (A) or the nucleotide fraction of fetal liver from DNase II+/− and DNase II− / − mice (B) was
incubated with the purified murine STING-Flag protein on ice for 30 min and analyzed by CN-PAGE, followed by Western blotting with anti-Flag mAb.
非ビオチン化ペプチド由来のノイズを大幅に低減できるこ とが示された.別の研究グループは,タマビジンの代わり に抗ビオチン抗体によってビオチン化ペプチドを濃縮する 方法を報告しているが7),筆者らが試した限りでは同定数 や特異性そして何よりも簡易性の点でタマビジンの方が優 れていたことを強調したい. STING 相互作用因子の大規模同定 上述の TurboID とタマビジンを組み合わせた改良型 BioID法を行い,リガンド刺激前後の STING 近接・相互 作用タンパク質を比較したところ,期待通り既知の下流 因子である TBK1 のビオチン化がリガンド刺激によって 顕著に増加していた.そして,リガンド刺激前の状態で はビオチン化タンパク質のほとんどが小胞体膜タンパク 質であった一方,刺激後には多数のゴルジ体膜タンパク 質やエンドソーム / リソソーム膜タンパク質,そして一部 の細胞質タンパク質のビオチン化が検出された.その中 に は zinc finger DHHC domain containing(ZDHHC) パ ル ミトイル化酵素群や,interferon regulatory factor 5(IRF5), interferon-induced transmembrane protein 3(IFITM3)など が含まれ,新規の相互作用因子であると考えられた5). Taguchiらのグループは,ゴルジ体に局在する ZDHHC-3, -7または -15 を過剰発現すると STING のパルミトイル化 が誘導されることを示し,パルミトイル化した STING が 下流の TBK1 や IRF3 と会合するモデルを提唱している8). 実際に,汎 ZDHHC 酵素阻害剤を細胞に投与すると TBK1 や IRF3 のリン酸化が抑制された.今回のスクリーニング では ZDHHC-3,-7 に加えて ZDHHC-13,-17,-20 が新た るオルガネラを免疫蛍光細胞染色により調べたところ,1 時間後に TurboID-STING がゴルジ体へ移行すると同時に ゴルジ体上でのビオチン標識が観察された.したがって, TurboIDは STING の時空間的相互作用の解析に有用であ ると考えられた. 現在の BioID 法では,ストレプトアビジンを用いてビオ チン化タンパク質をオンビーズ消化し,得られたペプチド を LC-MS/MS 解析によって同定する手法が広く用いられ ている.しかし,ビオチンとストレプトアビジン間の不可 逆的な相互作用のため,ビオチン化部位がストレプトアビ ジンビーズに結合したままとなり,その結果ほとんどが非 ビオチン化ペプチドとして検出されてしまう.この非ビオ チン化ペプチドは,相互作用タンパク質由来なのか非特異 的結合タンパク質由来なのか区別がつかないため,ビオチ ン化を直接的に検出することが信頼性の高いデータを得る ために不可欠である.タマビジン 2-REV は,タモギタケ から単離された新規アビジン様タンパク質タマビジン 2 の 遺伝子改変体で,可逆的にビオチンに結合するため過剰 量のビオチンで競合溶出が可能である6).そこで筆者らは, ビオチン標識タンパク質を先にトリプシン消化してビオチ ン化ペプチドと非ビオチン化ペプチドに断片化し,その 後タマビジン 2-REV を用いた補足と競合溶出によってビ オチン化ペプチドのみを濃縮・精製する方法を確立した5). 従来のストレプトアビジン法ではビオチン化ペプチドの同 定数が 10 個(総ペプチド同定数の 0.1%)であったのに 対し,新規タマビジン法では 4000 個(総ペプチ同定数の 80%)と飛躍的な増加が認められた.またこの結果は,単 純に同定数が上がっただけでなく,特異性の向上を意味し,
Fig. 2 Biotin labeling of STING-interacting proteins
STING-null RAW264.7 cells expressing STING-BioID, BioID-STING, TurboID-STING or miniTurbo-STING were cultured in the presence of 500 μM biotin for the indicated times. The cell lysates were analyzed by Western blotting with HRP-conjugated streptavidin.
た.そしてこれに伴って STING の分解が誘導されるとと もに,TBK1 のリン酸化が 1 時間をピークに徐々に減衰し ていくことが示された.したがって IFITM3 は,後期に起 こる STING の分解やシグナルの不活性化に寄与している のではないかと予想している. おわりに 細胞質 DNA に対する自然免疫応答は,cGAS,cGAMP, STINGといった主要なシグナル分子の同定によって急速 に理解が深まり,現在ではウイルス感染の研究分野に留ま らず,自己免疫疾患やがん,老化,肥満など,多くの研 究分野で関心を集めている.本稿で紹介した CN-PAGE を 用いた cGAMP 検出法や,TurboID とタマビジンを組み合 わせた改良型 BioID 法は,cGAS-cGAMP-STING シグナル の有用な解析ツールであり,様々な疾患モデルにおける cGAMPの検出や,シグナル制御機構の解明に貢献できる と考えている.実際に改良型 BioID 法を用いたスクリーニ ングでは,IRF5 や IFITM3 といった興味深い新規相互作 用因子の同定に成功した.しかし,今回は既知の下流因子 である IRF3 を同定することができなかった点についても 述べておく.これは IRF3 の発現量が非常に低かったこと が原因と思われる.このような低発現タンパク質を検出す るため,筆者らの研究室では BioID 法の更なる改善に取り 組んでおり,近い将来に実現することが期待される. 本論文に関して開示すべき利益相反状態は無い. 文 献
1) Kawane K, Motani K, Nagata S. DNA degradation and its de-fects. Cold Spring Harb Perspect Biol. 2014;6:a016394. 2) Motani K, Ito S, Nagata S. DNA-mediated cyclic GMP-AMP
synthase-dependent and -independent regulation of innate immune responses. J Immunol. 2015;194:4914–4923. 3) Roux KJ, Kim DI, Raida M, et al. A promiscuous biotin ligase
に同定されたことから,これら複数の酵素が協調的ある いは補完的に STING のパルミトイル化に寄与している可 能性が考えられた.IRF5 は IRF3 と同じ IRF 転写因子群 に属するが,STING 経路における役割は不明である.そ こで,STING を介して TBK1 依存的に IRF5 がリン酸化 される可能性について,Phos-tag ウエスタンブロットを 用いて解析した.内在性 STING を発現する RAW264.7 細 胞を STING リガンドで刺激したところ,約 1~2 時間後 に IRF5 のごく一部がバンドシフトする様子が観察された (Fig. 3A).このリン酸化のキネティクスは,STING のゴ ルジ体への移行と TBK1 のリン酸化と一致することがわ かった.さらにこのバンドシフトはホスファターゼ処理 または TBK1 阻害剤によって完全に消失した(Fig. 3B and 3C).免疫沈降法により IRF5 と STING の直接的な結合を 解析したが,検出することができなかった.これは言い換 えると,BioID 法を用いることで初めてこのような微弱ま たは一過性の相互作用が明らかになったといえる.以上の 結果から,STING の下流でリン酸化した IRF5 が何らかの 遺伝子発現を誘導すると考えられるが,リン酸化部位の特 定とともに今後の詳細な解析が待たれる.最後に STING と IFITM3 の相互作用について紹介する.IFITM3 はエン ドソームあるいはリソソーム膜に局在する膜タンパク質で, その N 末端領域が細胞質側を向く場合と内腔側に向く場 合の 2 つのトポロジーモデルが存在した.同定したビオチ ン化部位を調べると,N 末端領域の 24 番目と 88 番目のリ ジンがビオチン修飾されていた.TurboID 酵素は STING の細胞質領域と融合させているため,IFITM3 の N 末端は 細胞質側を向いていることがわかる.このように,BioID 法においてビオチン化を検出するもう一つのメリットは, トポロジーやおおまかな相互作用領域の情報が得られるこ とである.IFITM3 と STING は内在性レベルで強く結合 することが判明し,両者の結合はリガンド刺激 1 時間後 から 4 時間後にかけて徐々に増加することが明らかとなっ
fusion protein identifies proximal and interacting proteins in mammalian cells. J Cell Biol. 2012;196:801–810.
4) Branon TC, Bosch JA, Sanchez AD, et al. Efficient proxim-ity labeling in living cells and organisms with TurboID. Nat Biotech. 2018;36:880–887.
5) Motani K, Kosako H. BioID screening of biotinylation sites using the avidin-like protein Tamavidin 2-REV identifies glob-al interactors of stimulator of interferon genes (STING). J Biol Chem. 2020;295:11174–11183.
6) Takakura Y, Sofuku K, Tsunashima M. Tamavidin 2-REV: an engineered tamavidin with reversible biotin-binding capabil-ity. J Biotechnol. 2013;164:19–25.
7) Udeshi ND, Pedram K, Svinkina T, et al. Antibodies to biotin enable large-scale detection of biotinylation sites on proteins. Nat Methods. 2017;14:1167–1170.
8) Mukai K, Konno H, Akiba T, et al. Activation of STING requires palmitoylation at the Golgi. Nat Commun. 2016;7:11932.