平成
19 年度修士論文(工学)
褐藻
Pylaiella littoralis のミトコンドリア group II イントロンと
Chlorella saccharophila と同定された緑藻 KS-1 株の分子系統解析
Molecular phylogenetic analysis of mitochondrial group II introns detected in
a brawn alga Pylaiella littoralis and th phylogenetic position of a green alga '
Chlorella saccharophila ' (KS-1)
平成
20 年 3 月
高知工科大学大学院
工学研究科 基盤工学専攻
物質・環境システム工学コース
1105118 藤村政隆
目次
[1] Chlorella saccharophila と同定された緑藻 KS-1 株の分子系統解析
1. 要旨………1
2. 序論………1
3. 材料と方法………1
4. 結果………4
5. 考察………7
6. 図表………8
7. 参考文献………21
[2]
褐藻
Pylaiella littoralis のミトコンドリア group II イントロンの分子系統解析
1. 要旨………22
2. 序論………22
3. 材料と方法………23
4. 結果………23
5. 考察………25
6. 図表………26
7. 参考文献………21
[1] Chlorella saccharophila と同定された緑藻 KS-1 株の分子系統解析
1. 要旨
鹿児島県の硫黄島の海岸から採取・単離された KS-1 株は、MV(メチルビオロゲン)CdCl2に 対して耐性を持ち、形態的な特徴からChlorella saccharophila と同定された。 本研究では、Chlorella saccharophila と同定された KS-1 株の分子系統上の位置を推定するため に、核ゲノム上の遺伝子として 18S rDNA、ミトコンドリアゲノム上の遺伝子として cox1、葉緑 体ゲノム上の遺伝子として rbcL の DNA 塩基配列を決定した。これを他の緑藻の塩基配列と比較 し、近隣結合法によって分子系統樹を作成することで、KS-1 株の系統位置を解析した。その結 果 KS-1 株は、Chlorella saccharophila あるいは、その近縁種ではないことを示した。従って、形 態的な特徴に基づいた分類が誤りである可能性が高いことが分かった。 また、18S rDNA 遺伝子の塩基配列中に、group I イントロンと思われるイントロンを見出し、 この配列が group I イントロンであることを確かめる為に、group I イントロンの特有な二次構造 を作れるかどうか確かめた。group I イントロンの一般的な特徴を満たす 2 次構造モデルを作るこ とが出来たので、この配列はgroup I イントロンである可能性が高い。2. 序論
本研究は、Chrorella saccharophila と同定された緑藻 KS-1 株の分子系統上の位置を推定するこ とを目的としている。種の同定は現在、緑藻は大きく 5 分類される。陸上植物の起源になったと される車軸藻綱、原始的な緑藻であるプラシノ藻綱、主に海産で多細胞のアオサ藻綱、トレボキ シア藻綱、狭義の緑藻綱に分類される。分子系統学の発達により、近年、緑藻鋼がトレボキシア 藻鋼と狭義の緑藻鋼に 2 分割されて、5 綱となった。トレボキシア藻綱に含まれる代表的な緑藻 がクロレラであり、クラミドモナスやボルボックスは狭義の緑藻に含まれる。KS-1 株は Chlorella saccharophila と同定されている。核ゲノム上の遺伝子として 18S rRNA、ITS1 領域、ミトコンドリアゲノム上の遺伝子としてcox1、葉緑体ゲノム上の遺伝子として rbcL の DNA
塩基配列を決定して、他の緑藻の塩基配列と比較し、近隣結合法によって分子系統樹を作成する ことで、KS-1 株の分子系統的位置を解析した。
(保存されている株)
Chlorella saccharophila として公的に保存され配布されている株としては、UTEX Culture 247 と
CCAP Culture 211-9a がある。
3. 材料と方法
KS-1 株 鹿児島県の薩摩硫黄島から採取され、MV(メチルビオロゲン)でスクリーニングされた株で MV(メチルビオロゲン)に対して 3mM、CdCl2に対して 0.5mM 以上の耐性があることが分かっ ており、KS-1 株と名づけられたこの緑藻は、関西電力株式会社電力技術研究所環境技術センタ ーより譲りうけたものである。 KS-1 株は光学顕微鏡下での目視による観察の結果、鞭毛なし、細胞表面に模様なし、ピレノ イドあり、自生胞子形成様式の特徴から、KS-1 株はクロレラ属の一種、Chlorella saccharophila と同定された。培養条件
KS-1 株は MOM 培地(Modified Okamoto Medium 培地)で培養した。25_、5000lux(24h)の環 境下で静置、あるいは振とう培養を行った。
MOM(Modified Okamoto Medium)培地の組成(1L) NaCl 30g NH4Cl 270mg CaCl2・2H2O 200mg MgSO4・7H2O 250mg FeSO4・7H2O 20mg Thiamine-HCl 100mg VitaminB12 1mg
Trace element stock solution*1 2ml(_500 stock solution) DW 995ml
各成分をDW995ml に溶解し、pH を 8.0 に調整するために 1N NaOH を使用した。
pH8.0 調整後オートクレーブ滅菌し、温度が下がってから無菌的にリン酸 stock*2 を 5ml 加 える。
*1 Trance element stock solution (_500) EDTA・2Na 5g H3Bo3 286mg MnCl2・4H2O 181mg ZnSo4・7 H2O 22mg CuSo4・5 H2O 8mg NaMoO4 2mg CoCl2・6 H2O 1mg DW 200ml *2 リン酸 stock solution 1. 1M KH2PO4、1M KH2PO4を作成 2. 1M K2HPO4 500ml に 1M KH2PO4を徐々に加えpH を 8.0 に調整 3. オートクレーブ滅菌する。 ゲノムDNA の回収 KS-1 株は、細胞壁が非常に硬く、プロテアーゼ、ガス圧による破砕、中性フェノールで処 理をしても、細胞破砕率が低く DNA の回収が出来なかった。そこで、KS-1 株の細胞を破砕する ため石英砂を用いDNA の回収をした。 DNA の回収に用いた KS-1 株は、500ml マイヤーに MOM 培地 300ml を加え通気培養を行い 十分生育した株を用いた。50ml コニカルチューブに移し、遠心(5,000rpm 15min)で藻体を集める。 藻体にDNeasy Plant Mini Kit(QIAGEN)の Buffer AP1 液 500_l で縣濁する。縣濁液をすり鉢に入れ、 石英砂を少量加えて藻体をすり潰し、破砕された細胞の比率が 50-60%であることを顕微鏡によ って確認後、DNeasy Plant Mini Kit(QIAGEN)(日本語 Mini protocol 7 の RNase 処理から 9 まです り鉢上で行った)を用いゲノムDNA を抽出した。
PCR プライマーの設計
PCR プライマーを設計するために、緑藻の 18Sr RNA、ITS1、rbcL、 cox1 のデータを NCBI (National Center for Biotechnology Information)(http://www.ncbi.nlm.nih.gov/)で集め、集めたデ ータから相同性の高い部分をPCR プライマーとして用いた(表 1、2、3、4)。 集めたデータをアラインメントし、相同性の高い領域を見つけるために、ClustalX (ftp://ftp.ebi.ac.uk/pub/software/clustalw2)を用いた。ClustalX はアラインメントと近隣結合法に よる系統樹の作成にも用いた。 PCR による目的領域の増幅 上記で設計したプライマーを用いPCR を行った。PCR 反応液には EX-Taq Polymerase(TaKaRa) を用いた。
PCR 反応液は、EX Taq(TaKaRa)に添付されている_10EX-Taq buffer 3_L、2.5mM dNTP Mixture 3_L、10_M プライマー 各 2_L(最終濃度 0.5_M)、EX-Taq Polymerase 0.3_L(1.5unit)の反応液 にゲノム DNA を 30ng 含み、滅菌水で 30_L に調整した。PCR 反応の温度設定、サイクル数、各 ステップの時間は以下の通りである。18S rDNA 領域と ITS1 領域増幅の条件[94_(5min) _{94_(30sec)_55_(30sec)_72_(30sec)}_30cycles_72_(7min)] 、 rbcL と cox1 領 域 増 幅 の 条 件 [95_(5min)_{95_(30sec)_42_(30sec)_72_(30sec)}_40cycles_72_(7min)] それぞれの PCR 産物 6_L を 0.8%または 1.5%アガロースゲル/TAE で電気泳動し、バンドの有 無を確認した。 分子系統樹の作成 PCR 法で得られた塩基配列を用いて NCBI のデータベースから BLAST 検索を用い、相同性の 高い緑藻データを得た。その緑藻データと KS-1 株の塩基配列から分子系統樹の作成を行った。 多重アラインメントの作成には上記の ClustalX を用い、多重アラインメントの編集には SeaView (http://pbil.univ-lyon1.fr/software/seaview.html)を用いた。系統樹を作成するために ClastalX によ る近隣結合法の系統樹作成を行った。ブートストラップ値のための乱数は 111(初期値)、ブート ストラップ回数は 1000 として、分子系統樹の作成を行った。系統樹の表示と編集には NJplot (http://pbil.univ-lyon1.fr/software/njplot.html )を用いた。進化距離は、Kimura 2-パラメータ法に よる補正距離法を用いた。
4. 結果
PCR の結果(図 5、図 6、図 7、図 8)から、図 5 の 18Sr DNA からレーン_と_、図 6 の ITS1 からレーン_、図 7 の rbcL からレーン_と_、図 8 の cox1 からレーン_と_で示した産物がシ ングルバンドで尚且つオーバーラップしている箇所を基点とし補える領域だと判断し、PCR 産物 をWizardSV Gel and PCR Clean-Up System(Promega)で精製し、pT7blue ベクター(TAKARA) に TA クローニングしたものを大腸菌(E.coli DH5_ Competent Cells(東洋紡))に導入した。形 質転換した大腸菌をLB/amp/X-gal/IPTG 寒天培地上に撒き、カラーセレクションしたものを Wizard
Plus Minipreps DNA Purification System(Promega)で精製し、サンガー法により挿入 DNA 断片の
塩基配列を決定した。
その結果、18S rDNA 領域を 2585bp、18S rDNA 及び 5.8S rDNA を含む ITS1 領域を 364bp、rbcL 領域を1578bp、cox1 領域を 1058bp 得た。 18S rDNA 領域が予想したものより長く、さらに他の緑藻と相同性が見られない領域が挿入さ れており、予測より約700bp 長いバンドが出た。この他の緑藻と相同性が見られない領域は、group I イントロンが特異的に持つ 2 次構造を形成したので group I イントロンだと判断した。(図 13) シークエンシング結果 シークエンシングの結果、オーバーラップしている箇所を基点とし、18S rDNA、rbcL、cox1 の 塩基配列を決定した。 18S rDNA(2585bp) _Primer の箇所は赤で示した。青文字はgroup I イントロン領域を示す。 GTGCCACTGCGAATGGCTCATTAAATCAGTTATAGTTTATTTGATGGTACCTGCTACTCGGATACC CAGTACTAATTGTAGAGCTAATACGTGCGCACATCCCGACTCCTCGGAAGGGACGTATTTATTAG ATAAAAGGCCGACCGGGCTTGCCCGACTCGCGGCGAATCATGATAACTCCACGAATCGCACGG CCTCCGCGCCGGCGATGTTTCATTCAAATTTCTGCCCTATCAACTTTCGATGGTAGGATAGAGGC CTACCATGGTTTTGACGGGTGACGGGGAATTAGGGTTCTATGCCGGAGAGGGAGCCTGAGAAA CGGCTACCACATCCAAGGAAGGCAGCAGGCGCGCAAATTACCCAATCCCGACACGGGGAGGTA GTGACAATAAATAACAATACCGGGCTCTTTCGAGTCTGGTGATTGGAATGAGTACAATCTACATC CCTTAACGAGGATCCAGTGGAGGGCAAGTCTGGTGCCCGCAGCCGCGGTAATTCCAGCTCCAC TAGCGTATATTTAAGTTGTTGCAGTTAAAAAGCTCGTAGTCGGATGTCGGGCGGCGCCCGCCGG TCCGCCGTCAGGCGTGCACCGGCGTGGCGCCGCCTGTGTGCCGGGGACGGGCGCCTGGGCTTG ATTGTCCGGGCCCCGGAGTCGGCGAGGTCACTTTGAGTAAATTAGAGTGTTCAAAGCAGGCAG CCGCTCTGAATACGCCAGCATGGAATGACGCGATAGGACTCTGGCCTATTCCGTCGGTCTGTGG GACCGGAGTAATGATGAACAGGGACGGTCGGGGGCATTCGTATTTCGCTGTCAGAGGTGAAAT TCTTGGATTTGCGAAAGACGAACTTCTGCGAAAGCATTTGCCAAGGATGTTTTCATTGATCAAG AACGAAAGTCGGGGGCTCGAAGACGATTAGATACCGTCCTAGTCTCGACCATAAACGATGCCG ACTAGGGATCGGCGGGCGTTTCTTCGACGACCCCGCCGGCACCTCACGAGAAATCAAAGTGTT CGGGTTCCGGGGGGAGTATGGTCGCAAGGCTGAAACTTAAAGGAATTGACGGAAGGGCACCA CCAGGCGTTTGATGCTCGGCTCTGGCGCCTGAGAGTGGCGGCCGCGAGGCCTGCCGCTAGTGG CCTCCTTCCTTCCCTTGGGGAAGGGAGGGGGTCGCGACACTGTCAAATTGCGGGGACCTCCTA AAGCTTCGGGTGCCAAGCCCGGCCCGGAAACGGGCGGGTGGCCGGGGAGAAACACCCCGGG CGCGGCGACAACCCCGGAGATGGCACAATGGACGACCCGCAGCCAAGTCCCTCCCCGAGTTG CTCGAGGGGGGATGCAGTCCACAGACTAGATGGCAGTGGGCGGCGCCCTCTTTGTTTAAGAGT GCGTCGCTTAAGGTATAGTCGGCCCCCGGCCGAGAGGCCGGCCCGTCGGAGGACCGCCGCTGC TCCCAAACAGGGGGCACAGCGGCGCGAGAGCCGGCGGGGAGCGCAGCGTGCGCTCGGAATCG CGGGAGCCTGCGGCTTAATTTGACTCAACACGGGAAAACTTACCAGGTCCAGACATGGCGAGG ATTGACAGATTGACAGCTCTTTCTTGATTCTATGGGTTGACACGTGCGTCACACACCGCGCACC AACTGCCCGTCACAGTCGGGCGCGAAAGCGTCCGGCTAGTCCGCCTGCTCTCCTACTAGCGGG GAGCAGGCGGGCGAGGCCGTCAAATTGCGGGGACGCCCTAAAGTTCGCACCGCCAAGCCCGG
AGATATAGTCCGGCCCTCGCGAGAGCGAGCCCCCGCGAAGAAGCGCGCCTGGTGCGCGCCGG AGCGCGGGGGGCCGGGCGGGGCGTCCTGGATGGCGCAAACCCGCCGGCGGAGCAAACCGGGT GGTGCATGGCCGTTCTTAGTTGGTGGGTTGCCTTGTCAGGTTGATTCCGGTAACGAACGAGACC TCGGCCTGCTAAATAGCCCCGGGCGGCGCTCGCGCCGGCCGGCTGAGCTTCTTAGAGGGACTC TCGGCGACTAGCCGATGGAAGTGCGAGGCAATAACAGGTCTGTGATGCCCTTAGATGTTCTGGG CCGCACGCGCGCTACACTGACGCAGACAACGGGCGCGGCCTTGGCCGAGAGGCCCGGGTAAT CCGGCAGCCTGCGTCGTGACGGGGCTAGACTCTTGCAATTATCAGTCTTCAACGAGGAATGCCT AGTAGGCGCGAGTCATCAGCTCGCGTCGATTACGTCCCTGCCCTTTGTACACACCGCCCGTCGC TCCTACCGATTGGACGTGCTGGTGAAACGCTCGGACCGGTGGCGTGCCGCGGTTCGCCGCGGC GCACCGCCGGGAAGTCCGTTGAACCCTCCCGTCTAGAGGAAGGAGAAGTCGTAACAAG ITS1(364bp) Primer の箇所は赤で示した。 GGAGAAGTCGTAACAAGGTTTCCGTAGGAGAACCTGCGGAGGGATCATTGCGTCTATCGTTCA CACCGCTCGGCGTCTCTTTATAAAAAAGCCGGGGCCGCGGACCTCGCCAGAGGAATGCGGCTC CCCGCGACCCGCCTCCGGGCCGGCGCTCCCTTCCTCAAGAAGAGGGGGGTGTCGCGCCCGGC GGCCGGCGCGACCCCTTTTTCACACTGACCGCACCGGCCAGCCTGAAGCCGGACGCCGCCCTC CACCAGGAGGCGCCGTCTCAGCAAAACCAGAACAACTCTCAACAACGGATATCTCGGTTCCCG CAACGATGAAGAACGCAGCGAAATGCGATACTTGGTGTGAATTGCAGGAT
rbcL_pratial - nucleotide - (1578bp) Primer の箇所は赤で示す
GCAGGTGTTAAAGATTACCGCCTTACTTACTTTACACCAGATTATCAAGTAAAAGATACAGATAT TCTGGCAGCATTTCGTATGACTCCGCAACAAGGTGTACCTCCTGAAGAAGCAGGTGCAGCAGT TGCCGCAGAATCATCAACAGGAACTTGGACAACAGTATGGACGGACGGTTTAACAAGTTTAGA TCGTTATAAAGGTCGTTGTTATGATATTGAACCAGTTCGTGGAGAAGATAATCAATATATTGCATA TGTAGCATACCCTCTTGACCTTTTTGAAGAAGGATCAGTAACTAACTTATTTACTTCAATTGTAG GTAACGTTTTTGGGTTCAAAGCTCTTCGTGCTCTACGTTTAGAAGATCTTCGTATCCCGCCAGCT TACGTTAAAACATTCCAAGGTGCGCCTCATGGTATCCAAGTTGAACGTGATAAACTAAACAAAT ATGGTCGTTCACTTTTAGGTTGTACTATTAAACCAAAACTTGGACTATCAGCTAAAAACTATGGT CGTGCAGTTTATGAGTGTTTACGTGGTGGACTGGACTTTACAAAAGATGATGAGAACGTAAACT CACAACCATTTATGCGTTGGAGAGATCGATTCTTATTCGTAGCCGAAGCAATTTATAAATCTCAG GCAGAAACTGGGGAAGTTAAAGGACATTACCTAAACGCAACTGCAGGTACTGTTGAAGAAATG CTAAAACGTGCTGCAATGGCAAAAGATTTAGGTGTACCTATTATCATGCATGACTACCTAACTGG TGGTTTCACTGCAAATACTACTTTAGCACAATACTGTCGTGATGAAGGACTTTTATTACACATTC ACCGTGCAATGCATGCTGTAATTGACCGTCAAAGAAACCATGGTATTCATTTCCGTGTACTAGCA AAAGCTTTACGTTTATCAGGTGGTGATCACCTGCATTCAGGTACAGTTGTAGGGAAACTAGAAG GTGAACGTGAAGTAACTTTAGGTTTCGTAGACTTAATGCGTGATGATTACATTGAGAAAGACCG TAGCCGAGGTGTTTATTTCACACCAGACTGGGTTTCTCTTCCTGGTGTTATGCCAGTAGCTTCAG GTGGTATTCACGTATGGCACATGCCTGCACTTGTAGAAATTTTTGGAGATGATGCATGTCTTCAA TTTGGTGGAGGTACTTTAGGACACCCATGGGGTAATGCTCCAGGTGCTGCAGCAAACCGTGTAG CTTTAGAAGCTTGTACACAAGCACGTAATGAAGGTCGTGACCTTGCACGTGAAGGTGGTGATG TTATTCGTGCAGCTTGTAAATGGAGTCCAGAATTAGCAGCAGCTTGTGAAGTTGGAAAGAAATT AAATTTGAATTCGAAACTATCGATAC
cox1 - nucleotide - (1058bp) Primer の箇所は赤で示す
ACTTATCATAAGGATATTGGTACTCTTTATCTGATTTTCGGTGCTTTCTCTGGATTACTGGGAACA TGTTTTTCGATACTTATTCGTATGGAACTTTCTCAACCAGGAAATCAAATTTTAGAGGGGAATCA TCAATTATATAATGTGATTATTACCGCTCATGCTATTTTAATGATTTTTTTTATGGTAATGCCTGCAA TGATAGGTGGTTTTGGTAACTGGTTTGTTCCTATTTTAATTGGTGCTCCAGATATGGCATTCCCAC
TAGAAGTTGGTGCTGGTACTGGTTGGACTATTTATCCACCACTTAGCAGTATTGCTAGTCACTCT GGAGGAGCAGTGGATTTAGCTATTTTTAGCTTACACCTTTCTGGTATTAGTAGTATTTTAGGTGCT ATTAACTTTATTACTACAATTTTTAATATGCGTGCACCAGGATTAAGTATGCACCGTTTACCTTTAT TTGTATGGTCTGTTCTAATTACTGCATTTTTATTATTATTATCTTTACCAGTTTTAGCAGGAGCAAT TACTATGCTTTTAACAGATCGTAACTTTAACACTACTTTCTTTGATCCAGCAGGAGGTGGAGACC CGGTATTATTTCAACACCTTTTTTGGTTTTTTGGTCATCCAGAGGTGTACATTTTAATTTTACCTG GTTTTGGTATAATTAGTCATATCGTATCAACTTTTTCTAAAAAACCTATTTTTGGGTATTTAGGAAT GGTTTATGCAATGTTAAGTATTGGAATTCTAGGATTTATTGTCTGGGCTCATCATATGTATACTGTA GGGTTAGACATAGATACTCGTGCTTATTTTACAGCTGCTACTATGATTATCGCTGTCCCTACTGGA ATTAAAGTGTTCAGTTGGATTGCTACCATGTGGGGGGGAAGTATTGAATTCAAAACACCTATACT CTTTGCTGTGGGTTTTATCTTCTTATTTACTATAGGAGGATTAACAGGTGTCAACTTAACTTTCTT 塩基配列比較の結果 核ゲノム上の遺伝子18S rDNA の塩基配列による比較(図 9) KS-1 株で見出された group I イントロンをコードする領域は、他の緑藻中で相同性が見 出されなかったので、分子系統樹を作成する配列中には含めなかった。
MBI から登録されているデータを赤で示している。非常に高い Bootstrap 値(1000)で、MBI から登録されている Chlorella saccharophila などが強固なクラスターを形成しているが、KS-1 株
はその中に含まれない。また、GenBank に登録されている 18S rDNA 配列の中に KS-1 株と同一 種と思われるほど、酷似したデータは見出せなかった。Chlorella luteoviridis とクラスターを形成
しているが、進化距離が長いため long branch effect により、分子系統樹に使用した他の緑藻と近 縁でないもの同士がクラスターを形成したものだと考えられる。
ITS1 領域の塩基配列比較による比較(図 10)
ITS1 領域でも KS-1 株と近縁になるような緑藻は見出されておらず、Chlorella saccharophila と近縁関係に無いことが示されるデータしか得られなかった。
葉緑体ゲノム上の遺伝子rbcL 領域の塩基配列比較による比較(図 11)
rbcL 配列による分子系統樹では、アミノ酸であれ、塩基配列であれ、18S rDNA の分子系統
樹と矛盾しない系統樹が得られ、Chlorella luteoviridis とクラスターを形成したが、18S rDNA と
同じく進化距離が長く、他の緑藻と近縁に無いもの同士がクラスターを形成していると考えられ る。 ミトコンドリアゲノム上の遺伝子cox1 領域の塩基配列比較による比較(図 12) cox1 の登録されている緑藻の数が限られており、分子系統樹に使用した領域も短く。強固な Bootstrap 値を持つクラスターが、トレボキシア藻鋼の中で形成されておらず、信用できる分子系 統樹を作らなかった。
5. 考察
KS-1 株について KS-1 株は、形態的特徴により Chlorella saccharophila と同定されたが、分子系統解析の結果、 近縁関係になく、形態的な特徴に基づいた分類が誤りである可能性が高いことが分かった。これ は、トレボキシア藻鋼にクロレラと分類されたものが一塊に集まらず、いろんなクラスターに存 在することからも明らかなように、現状の形態的な定義ではクロレラ属が人為分類群となってし まうことに本質的な問題があると考えられる。また、現在KS-1 株の近縁種の塩基配列データは、核 large subunit rRNA、ミトコンドリア cox1、 葉緑体rbcL を含めて Genebank には登録されていないことが分かった。
6. 図表
表1.18S rDNA 領域増幅用プライマー
下記の配列を持つ合成DNA を用いた。
_KS_18Spartial_FP
5’ – GTG CCA CTG CGA ATG GCT CAT – 3’ _KS_18S_FP2
5’ – GCA GCA GGC GCG CAA ATT ACC – 3’ _KS_18S_FP3
5’ – AAC GAT GCC GAC TAG GGA TCG G – 3’ _KS_18S_RP2
5’ – ACT ACG AGC TTT TTA ACT GCA – 3’ _KS_18S_RP3
5’ – TCC AAG AAT TTC ACC TCT GA – 3’ _KS_18S_RP4 5’ – CTT GTT ACG ACT TCT CCT TCC – 3’ 上記のプライマーがアニールする18S rDNA 内のおよその位置を図 1 に示した。 図1. 18S rDNA とプライマーの位置関係 表2. ITS1 領域増幅用プライマー 下記の配列を持つ合成DNA を用いた。 _BMCF
5’ – GGA GAA GTC GTA ACA AGG TTT CCG – 3’ _BMCR
5’ – ATC CTG CAA TTC ACA CCA AGT ATC G – 3’ _KS_ITS_R1(18S rDNA の PCR で使用) 5’ – CGC ATT TCG CTG CGT TCT TCA – 3’ 上記のプライマーがアニールする ITS1 領域内のおよその位置を図 2 に示した。また、18S rDNA 領域と ITS1 領域を含む真核生物におけるリボソーム RNA 遺伝子オペロンの一般的な構造を 同じ図2 で示す。 図2.リボソーム遺伝子オペロン構造の概略と ITS1 領域とプライマーの位置関係 ③KS_ITS_R1 ②BMCR 5’ 3’ ①BMCF 約2kbp 18SrDNA ITS1領域(約250bp) 5.8SrDNA 26SrDNA ITS2領域 ②KS_18S_FP2 ③KS_18S_FP3 ④KS_18S_RP2 ⑤KS_18S_RP3 ⑥KS_18S_RP4 5’ 3’ ①KS_18Spartial_FP
表3. rbcL 遺伝子領域増幅用プライマー
rbcL のほぼ全領域 1.5kb をいくつかの断片として PCR 法で増幅するために、下記配列を持つ
合成DNA を用いた。 _rbcL_KS-1FP1
5’ – GCA GGT GTT AAA GAY TAC CG – 3’ _rbcL_KS-1FP2
5’ – TGG ACT GAY GGT TTA ACT AG – 3’ _rbcL_KS-1FP3
5’ – GGW ATT CAA GTA GAA CGT GA – 3’ _rbcL_KS-1RP1
5’ – TCT CTC CAA CGC ATR CCT ATT – 3’ _rbcL_KS-1RP2
5’ – YTT TGA CGG TCA ATT ACA GC – 3’ _rbcL_KS-1RP3
5’ – ATT ACA TCA CCR CCT TCA CG – 3’ _rbcL_KS-1RP4
5’ – GTA TCG ATW GTT TCG AAT TC – 3’ 但し Y = C/T , W = A/T , R = A/G 図3.rbcL 遺伝子内の上記プライマーの位置 ①rbcL_KS-1FP1 ②rbcL_KS-1FP2 ④rbcL_KS-1RP1 ⑦rbcL_KS-1RP4 ⑤rbcL_KS-1RP2 ⑥rbcL_KS-1RP3 ③rbcL_KS-1FP3 5’ 3’
表4.cox1 遺伝子領域増幅用プライマー
cox1 のほぼ全領域 1.5kb をいくつかの DNA 断片として PCR 法で増幅するために、下記の配
列を持つ合成DNA を用いた。 _cox1_KS-1FP
5’ – ACY YMT CAY AAR GAT ATT GG – 3’ _cox1_KS-1FP2
5’ – TTC TTY ATG NTN ATG CCT GC – 3’ _cox1_KS-1RP1
5’ – TCW GGR TGW CCR AAR AAC CA – 3’ _cox1_KS-1RP2
5’ – GCA CCC ATW GCR CAA ATC AT – 3’ _cox1_KS-1RP3
5’ – AAG AAD GTY AAR TTR ACA CC – 3’ _cox1_KS-1RP4
5’ – AAN ARW GGN ART CTA TGC AT – 3’ _cox1_KS-1RP5
5’ – CCA AMM ATW GGT TTT TTA GA – 3’
但し N = A/C/G/T , Y = C/T , W = A/T , R = A/G , M = A/C , D = A/G/T
図4.cox1 遺伝子内の上記プライマーの位置
①cox1_KS-1FP1
⑥cox1_KS-1RP4 ③cox1_KS-1RP1⑦cox1_KS-1RP5④cox1_KS-1RP2
②cox1_KS-1FP2
5’ 3’
図5.18S rDNA の増幅結果 レーン_(_KS_18Spartial_FP と_KS_18S_RP2 プライマーによる PCR) 予測される産物の長さ:約550bp 増幅された PCR 断片の長さ:約 550bp レーン_(_KS_18Spartial_FP と_KS_18S_RP3 プライマーによる PCR) 予測される産物の長さ:約850bp 増幅された PCR 断片の長さ:約 850bp レーン_(_KS_18Spartial_FP と_KS_18S_RP4 プライマーによる PCR) 予測される産物の長さ:約1700bp 増幅された PCR 断片の長さ:PCR 産物なし レーン_(_KS_18Spartial_FP と KS_ITS_R1 プライマーによる PCR) 予測される産物の長さ:約1950bp 増幅された PCR 断片の長さ:約 800bp (目的でない産物) レーン_(_KS_18Spartial_FP と BMCR プライマーによる PCR) 予測される産物の長さ:約1950bp 増幅された PCR 断片の長さ:PCR 産物なし レーン_(_KS_18S_FP2 と_KS_18S_FP2 プライマーによる PCR)(control) 予測される産物の長さ:?bp 増幅された PCR 断片の長さ:PCR 産物なし レーン_(_KS_18S_FP2 と_KS_18S_RP3 プライマーによる PCR) 予測される産物の長さ:約500bp 増幅された PCR 断片の長さ:約 500bp レーン_(_KS_18S_FP2 と_KS_18S_RP4 プライマーによる PCR) 予測される産物の長さ:約1350bp 増幅された PCR 断片の長さ:約 2200bp レーン_(_KS_18S_FP2 と KS_ITS_R1 プライマーによる PCR) 予測される産物の長さ:約1600bp 増幅された PCR 断片の長さ:約 2300bp レーン_(_KS_18S_FP2 と BMCR プライマーによる PCR) 予測される産物の長さ:約1600bp 増幅された PCR 断片の長さ:約 2400bp レーン_(_KS_18S_FP3 と_KS_18S_FP3 プライマーによる PCR)(control) 予測される産物の長さ:?bp 増幅された PCR 断片の長さ:PCR 産物なし レーン_(_KS_18S_FP3 と_KS_18S_RP4 プライマーによる PCR) 予測される産物の長さ:約700bp 増幅された PCR 断片の長さ:PCR 産物なし レーン_(_KS_18S_FP3 と KS_ITS_R1 プライマーによる PCR) 予測される産物の長さ:約950bp 増幅された PCR 断片の長さ:PCR 産物なし レーン_(_KS_18S_FP3 と BMCR プライマーによる PCR) 予測される産物の長さ:約950bp 増幅された PCR 断片の長さ:PCR 産物なし 500bp 1500bp
図6.ITS1 領域の増幅結果 レーン_(_BMCF と_BMCF プライマーによる PCR)(control) 予測される産物の長さ:?bp 増幅された PCR 断片の長さ:PCR 産物なし レーン_(_BMCF と_BMCR プライマーによる PCR) 予測される産物の長さ:約250bp 増幅された PCR 断片の長さ:約 300bp レーン_(_BMCR と_BMCR プライマーによる PCR)(control) 予測される産物の長さ:?bp 増幅された PCR 断片の長さ:PCR 産物なし
500bp
1500bp
図7.葉緑体ゲノムコード rbcL の増幅結果 レーン_(_rbcL_KS-1FP1 と_rbcL_KS-1RP1 プライマーによる PCR) 予測される産物の長さ:約600bp 増幅された PCR 断片の長さ:約 600bp レーン_(_rbcL_KS-1FP1 と_rbcL_KS-1RP2 プライマーによる PCR) 予測される産物の長さ:約850bp 増幅された PCR 断片の長さ:約 800bp レーン_(_rbcL_KS-1FP1 と_rbcL_KS-1RP3 プライマーによる PCR) 予測される産物の長さ:約1250bp 増幅された PCR 断片の長さ:約 1100bp レーン_(_rbcL_KS-1FP1 と_rbcL_KS-1RP4 プライマーによる PCR) 予測される産物の長さ:約1350bp 増幅された PCR 断片の長さ:約 1200bp レーン_(_rbcL_KS-1FP2 と_rbcL_KS-1RP1 プライマーによる PCR) 予測される産物の長さ:約400bp 増幅された PCR 断片の長さ:約 400bp レーン_(_rbcL_KS-1FP2 と_rbcL_KS-1RP2 プライマーによる PCR) 予測される産物の長さ:約650bp 増幅された PCR 断片の長さ:約 650bp レーン_(_rbcL_KS-1FP2 と_rbcL_KS-1RP3 プライマーによる PCR) 予測される産物の長さ:約1100bp 増幅された PCR 断片の長さ:約 1000bp レーン_(_rbcL_KS-1FP2 と_rbcL_KS-1RP4 プライマーによる PCR) 予測される産物の長さ:約1200bp 増幅された PCR 断片の長さ:約 1100bp レーン_(_rbcL_KS-1FP3 と_rbcL_KS-1RP3 プライマーによる PCR) 予測される産物の長さ:約850bp 増幅された PCR 断片の長さ:約 800bp レーン_(_rbcL_KS-1FP3 と_rbcL_KS-1RP4 プライマーによる PCR) 予測される産物の長さ:約950bp 増幅された PCR 断片の長さ:約 900bp
500bp
1500bp
図8.ミトコンドリアゲノムコード cox1 の増幅結果 レーン_(_cox1_KS-1FP1 と_cox1_KS-1RP1 プライマーによる PCR) 予測される産物の長さ:約700bp 増幅された PCR 断片の長さ:約 650bp レーン_(_cox1_KS-1FP1 と_cox1_KS-1RP2 プライマーによる PCR) 予測される産物の長さ:約800bp 増幅された PCR 断片の長さ:約800bp と複数の産物 レーン_(_cox1_KS-1FP1 と_cox1_KS-1RP3 プライマーによる PCR) 予測される産物の長さ:約1250bp 増幅された PCR 断片の長さ:約 650bp+約 1100bp レーン_(_cox1_KS-1FP1 と_cox1_KS-1RP4 プライマーによる PCR) 予測される産物の長さ:約500bp 増幅された PCR 断片の長さ:PCR 産物なし レーン_(_cox1_KS-1FP1 と_cox1_KS-1RP5 プライマーによる PCR) 予測される産物の長さ:約750bp 増幅された PCR 断片の長さ:約 700bp レーン_(_cox1_KS-1FP2 と_cox1_KS-1RP1 プライマーによる PCR) 予測される産物の長さ:約500bp 増幅された PCR 断片の長さ:約 500bp レーン_(_cox1_KS-1FP2 と_cox1_KS-1RP2 プライマーによる PCR) 予測される産物の長さ:約600bp 増幅された PCR 断片の長さ:約600bp と複数の産物 レーン_(_cox1_KS-1FP2 と_cox1_KS-1RP3 プライマーによる PCR) 予測される産物の長さ:約1050bp 増幅された PCR 断片の長さ:約 800bp (目的でない産物) レーン_(_cox1_KS-1FP2 と_cox1_KS-1RP4 プライマーによる PCR) 予測される産物の長さ:約300bp 増幅された PCR 断片の長さ:PCR 産物なし レーン_(_cox1_KS-1FP2 と_cox1_KS-1RP5 プライマーによる PCR) 予測される産物の長さ:約550bp 増幅された PCR 断片の長さ:約 550bp
500bp
1500bp
図9.核ゲノム上遺伝子 18S rDNA の塩基配列比較による分子系統樹
形 態 的 な 特 徴 か ら KS-1 株 は Chlorella saccharophila と 同 定 さ れ て い る の で 、 Chlorella
saccharophila の 18S データー名を赤で示した。
表5.系統樹に使用した塩基配列のデータ
Species Data source Chlamydomonas reinhardtii M32703 Chlorella angustoellipsoidea AB006047 Chlorella luteoviridis X73997 Chlorella minutissima X56102 Chlorella saccharophila strain:SAG211-9a X63505 Chlorella saccharophila strain:MBIC10037 AB183575 Chlorella saccharophila strain:MBIC10038 AB058300 Chlorella saccharophila strain:MBIC10039 AB058301 Chlorella saccharophila strain:MBIC10040 AB183576 Chlorella saccharophila strain:MBIC10041 AB183577
Chlorella saccharophila strain:MBIC10042 AB183578 Chlorella saccharophila strain:MBIC10067 AB058306 Chlorella saccharophila strain:MBIC10092 AB058310 Chlorella sorokiniana X62441 Chlorella sp. Yanaqocha RA1 Y14950 Chlorella sp. strain:MBIC10043 AB183579 Chlorella sp. strain:MBIC10747 AB183635 Chlorella vulgaris X13688 Closteriopsis acicularis Y17470 Desmodesmus communis X73994 Dictyochloropsis reticulata Z47207 Koliella spiculiformis AF278744 Nannochloris sp. RCC 011 AJ131691 Parachlorella kessleri X56105 Picochlorum eukaryotum X06425 Prototheca wickerhamii X56099 Pseudochlorella sp. CCAP 264-2 AB006049 Viridiella fridericiana AJ439401 Watanabea reniformis X73991
図10.ITS1 領域の塩基配列比較による分子系統樹
表6.系統樹に使用した塩基配列のデータ
Species Data source Chlorela sf minutissima EF030581 Chlorella saccharophila EF030587 Chlorella sorokiniana EF030580 Chlorella sp. AcGK EF030577 Chlorella sp. AcPIB EF030576 Chlorella sp. OCH EF030578 Chlorella sp. PbKM2 EF030584 Chlorella sp. PbPIB EF030582 Chlorella sp. Pbu EF030579 Chlorella sp. PbW EF030583 Chlorella sp. PtPIB EF030573 Chlorella sp. SpPIB EF030575 Chlorella sp. SvPIB EF030572 Chlorella sp. TtPIB EF030574 Chlorella sp. UPIB EF030571 Chlorella vlgaris AB162910 Pseudochlorella pyrenoidosa AM422986 Viridiella fridericiana AJ439402
図11.葉緑体ゲノム上遺伝子 rbcL の塩基配列比較による分子系統樹
表7.系統樹に使用した塩基配列のデータ
Species Data source Actinastrum hantzschii EF113405 Bryopsis maxima X55877 Chlamydomonas reinhardtii NC_005353 Chlorella ellipsoidea D10997 Chlorella luteoviridis EF113428 Chlorella pyrenoidosa AB240145 Chlorella saccharophila AM260446 Chlorella vulgaris NC_001865 Nephroselmis olivacea NC_000927 Ostreococcus tauri CR954199 Pediastrum duplex AB084333 Pseudendoclonum akinetum NC008114 Scenedesmus obliquus NC008101 Tetraselmis suecica DQ173247 Tolypella prolifera AF097175
図12.ミトコンドリアゲノム上遺伝子 cox1 の塩基配列比較による分子系統樹
表8.系統樹に使用した塩基配列のデータ
Species Data source Actinastrum hantzschii D63660 Chara vulgaris AY267353 Chlamydomonas eugametos NC_001872 Chlamydomonas reinhardtii U03843 Chlorella reisiglii AB009363 Chlorella vulgaris AB011523 Coelastrum microporum D63656 Kirchneriella lunaris D63653 Nephroselmis olivacea NC_008239 Oltmannsiellopsis viridis DQ365900 Ostreococcus tauri NC_008290 Pediastrum boryanum D63659 Platymonas subcordiformis Z47795 Prototheca wickerhamii U02970
Pseudendoclonium akinetum NC_005926 Scenedesmus obliquus AF204057 Scenedesmus quadricauda AB011524 Tetraedoron bitridens D63657 Volvox sp. D63661
7. 参考文献
Hayashi-Ishimaru Y, Ohama T, Kawatsu Y, Nakamaru K, Osawa S. UAG is a sense codon in several chlorophycean mitochondria. Curr Genet. 1996; 30: 29-33
Huss VAR, Ciniglia C, Cennamo P, Cozzolino S, Pinto G, Polio A. Phylogenetic relationships and taxonomic position of Chlorella - like isolates from low pH environments (pH < 3.0). BMC Evol Biol. 2002; 2: 13. Published online 2002
Huss VAR, Frank C, Hartmann EC, Hirmer M, Kloboucek A, Seidel BM, Wenzeler P, Kessler E. Biochemical taxonomy and molecular phylogeny of the genus Chlorella sensu lato (Chlorophyta). J. Phycol. 1999; 35: 587-598.
Rot C, Goldfarb I, Ilan M, Huchon D. Putative cross-kingdom horizontal gene transfer in sponge (Porifera) mitochondria. BMC Evol Biol. 2006; 6: 71 Published online 2006
Watanabe KI, Ehara M, Inagaki Y, Ohama T. Distinctive origins of group I introns found in the COXI genes of three green algae. Gene. 1998; 213: 1-7
[2] 褐藻 Pylaiella littoralis のミトコンドリア group II イントロンの分子系統解析
1. 要旨
イントロンには、核型イントロンのほかにgroup I イントロンと group II イントロンと呼ばれる ものが存在し、それらのイントロン RNA 配列は、自分自身を切り出して前後のエキソンを結合 する活性を持つリボザイム分子の一種であることが分かっている。そのうちの一つ group II イン トロンは、オルガネラや細菌、ウィルスのゲノムから見出されている。group II イントロン内に は、スプライシングされたイントロン RNA を DNA の特定配列部に挿入させた後に、これを逆 転写して DNA に変換するのに必要な酵素がコードされている。この酵素の働きは、group II イン トロンが水平伝播する時に使われると考えられている。オルガネラの遺伝子内に侵入したイント ロン DNA は、メカニズムは不明だが、進化的に長い時間のうちに消えてしまう。フランスで採 集された褐藻P.littoralis のミトコンドリア LSU rRNA 遺伝子には 4 つの group IIB イントロンが、 cox1 遺伝子には、3 つの group IIA イントロンの存在が報告されている。6 カ国で採集された P.littoralis について、サンプル毎に、上記 2 種類の遺伝子内に挿入されているイントロンの数は様々であること、また同一サイトに挿入さているイントロンであっても、その内部配列に大きな 違いが見られることが解っている。これらのデータを系統解析することにより、日本で採取され たサンプルで見出された、LSU rRNA 遺伝子内の第 4 イントロンとフランスのサンプルが持つ遺 伝子内の第1 イントロは、ごく最近侵入してきたイントロンである事が明らかになった。
データベースから P.littoralis の group II イントロン内 ORF とホモロジーのある塩基配列、アミ
ノ酸配列得て、P.littoralis の group II イントロンとの相互関係を調べることにより、P.littoralis に
侵入してきたイントロンがどの種由来なのか、または近い種を明らかに出来ると考え分子系統解 析を行った。その結果データベースには P.littoralis の group II 内 ORF と高い相同性を示す配列が
無く、P.littoralis のミトコンドリア内にイントロンを提供したと思われる生物種の特定はできな かった。
2. 序論
group II イントロンは多細胞動物を除く生物のオルガネラや細菌、ウィルスから見出されてい る。このイントロンは group I イントロンと同様に種の壁を越えて、転移できることが分子系統 解析から分かっている。接合可能な近縁種でイントロンを持つ種と持たない種を人為的に交配さ せることにより、イントロンの種間転移を観測できる。しかし、自然界におけるイントロンの種 を越えたメカニズムは未だ不明である。ただ、種外からいったん種内に入り込んだ group II イン トロンは、接合を介して種内に休息に伝播されると考えられている。また、遺伝しないに侵入し たイントロンは、メカニズムは不明だが、進化的に長い時間のうちに跡形も無く消えてしまう。 やがて、回復したターゲット配列内に新たにイントロンが侵入する。遺伝子のある特定サイトに ついて見ると、このようなサイクルが成立していると考えられる。group II イントロンのアーム IV には逆転写酵素活性をもつ領域と、イントロンRNA が正しく折りたたまれるのを助ける maturase 領域、エンドヌクレアーゼ活性を持つDNase 領域を持つ 1 つの ORF(Open Reading Frame)が挿 入されていることがある。このうち逆転写酵素活性領域と DNase 領域は、それまでイントロン が無かったサイトに新たにイントロンが挿入されるために必要であるが、スプライシング反応の 進行には必要ではない。従って、3 つの完全な領域から構成されている ORF が残っているイン トロンは、そのサイトに比較的最近挿入されたイントロンであると考えられる。maturase 領域を 除く、逆転写酵素活性領域や DNase 領域の一部に欠失が生じた不完全な ORF はしばしば見出さ れる。Pylaiella littoralis のミトコンドリアゲノムの全塩基配列は決定されており、全遺伝子中 group II
ことが示されている褐藻 Laminaria digitata でも、ミトコンドリアゲノムの全塩基配列が決定され
ているが、group II イントロンは見出されていない。3 種の褐藻、Dictyota dichotoma、Fucus vesiculosus、
Desmarestia viridis でもミトコンドリアゲノムの全塩基配列が決定されているが、これらの褐藻か
らもgroup II イントロン見出されていない。
Fontain は 1995 年の論文で、他の地域からの同一種のサンプルでは、イントロン構成が異なる と論文で述べているが、データは示されていない。World wide に様々な地域から採集した Pylaiella
littoralis と Pylaiella littoralis 種外のイントロン多型を解析することにより、イントロンの侵入か
らイントロンの喪失までの過程に関する有用な情報を得られることが期待された。
3. 材料と方法
Pylaiella littoralis 固着性の褐藻であり、無性生殖で増殖する。ミトコンドリア全塩基配列が決定しており、今の ところ褐藻では唯一 group II イントロンが見出されている事と、ほぼ完全な ORF を有している ことから、比較的最近種外からgroup II イントロンが挿入したと考えられた。World wide な Pylaiella littoralis の group II イントロンのデータは大阪教育大学 理科教育講座 の生田享介准教授より提供して頂いたものである。(図2、表 1)
group II イントロンデータの採集
Pylaiella littoralis の group II イントロンの ORF のアミノ酸配列を用い、NCBI
(http://www.ncbi.nlm.nih.gov/)から相同性のあるデータを BLAST 検索で集めた。 分子系統樹の作成 多重アラインメントの作成には ClustalX(ftp://ftp.ebi.ac.uk/pub/software/clustalw2)を用いた。 多重アラインメントの編集にはSeaView(http://pbil.univ-lyon1.fr/software/seaview.html)を用いた。 系統樹を作成するために ClastalX による近隣結合法の系統樹作成を行った。ブートストラップ 値のための乱数は 111(初期値)、ブートストラップ回数は 1000 として、分子系統樹の作成を行 った。系統樹を表示と編集には NJplot(http://pbil.univ-lyon1.fr/software/njplot.html )を用いた。 進化距離は、Kimura 2-パラメータ法による補正距離法を用いている。 種内多型解析には、イントロン全体の塩基配列を用いて分子系統樹を作成している。種外多型 解析では、ORF 部分の逆転写酵素活性領域のアミノ酸配列を用いて分子系統樹を作成した。
4. 結果
種内多型解析による結果 LSU rRNA 遺伝子内の第 1 イントロンについて(図 3) France-I 株で報告されているこのイントロンは、地理的にも近接している地域から得られた France-II 株と England 株でのみ検出されている。France-I 株のみ完全な ORF を持つのに対して、 France-II 株と England 株のイントロンから抽出可能な ORF はごく短く、進化距離が France-I は他の2 株との進化距離が大きい。このことは、現在 France-II と England 株の持つイントロンは、こ れらの共通祖先から受け継がれたものであるのに対し、France-I 株のものはそうでなく、最近種 外から完全なORF を持つイントロンが再侵入したと考えれば、他の 2 株に対して長い進化距離 を持つFrance-I 株が完全な ORF を持つことを説明できる。 3 株のイントロンを比較すると、そのリボザイム部分に France-II と England 株にのみ共通する 塩基が見られることから、France-II と England 株は共通祖先から、このイントロンを受け継いで いることの説明ができる。 LSU rRNA 遺伝子内の第 2 イントロンについて(図 4)
France-I 株で報告されているこのイントロンは、France-II 株と England 株で検出されている。 しかしFrance-II、England 株共にイントロン内で抽出できる ORF は短く、16 アミノ酸残基に相 当する部分が検出されるのみだった。3 株の進化距離は完全な ORF を持つか持っていないか関 係なく相互に短かった。このことからFrance-I、France-II、England 株の 3 株の共通祖先にあった イントロンが受け継がれており、France-II と England 株の共通祖先で ORF の大部分に欠失が生 じたと考えられる。
LSU rRNA 遺伝子内の第 3 イントロンについて(図 5)
France-I 株で検出されているこのイントロンは、France-II 株と England 株で検出されている。 どのイントロンも抽出可能なORF が無く、進化距離が短かった。このことから、France-I、France-II、 England 株の共通祖先において、すでに第 3 イントロンの ORF は、ほぼ完全に欠失していると考 えられる。
LSU rRNA 遺伝子内の第 4 イントロンについて(図 6)
France-I 株で検出されているこのイントロンは、France-II、England、Japan、Chile-II 株で検出 されている。Chile-II 株が持つイントロンは完全な ORF を持ち、Japan 株は容易に完全な ORF を 抽出できる配列を持っていた。 Japan 株を除く、France-I、France-II、England、Chile-II 株の進化距離は相互に短く大きな差も なく、これら4 株から得られたイントロンの配列は高い Bootstrap 値で支持されたクレイドを形 成する。それに対して、Japan 株のイントロンだけが、このクレイドに含まれない。また、Japan 株は他の4 株と長い進化距離を持つにもかかわらず、完全に近い ORF を持つので Japan 株のイ ントロンはこのサイトに比較的最近侵入したと考えられる。 従って、現在、France-I、France-II、England、Chile-II 株が持つイントロンが、これら 4 株の共 通祖先から直接受け継がれたものであるのに対して、Japan 株のものだけが、そうではないこと を示している。4 株のうち Chile-II 株だけが完全な ORF を今も持っている。従って、これらの 4 株の共通祖先のイントロンは完全なORF を持つものであり、それが Chile-II 株に、ほぼ完全に近 い状態で受け継がれ維持されているのに対し、France-I、France-II、England 株では、それぞれ独 立または、共通祖先でORF の大部分に欠失したものと考えられる。 cox1 遺伝子内の第 1 イントロンについて(図 7) France-I 株で検出されているこのイントロンは、France-II、Ireland、Germany、Japan、USA、Chile-I 株から検出されている。それぞれ完全なORF か容易に完全な ORF を検出できる配列をもってお り、相互に進化距離が短く、高いBootstrap 値で支持されている、このことから、それぞれの共 通祖先において完全なORF を持つイントロンが存在し、わずかな塩基置換や欠失が生じたのみ で現在まで受け継いで維持していると考えられる。
全なORF を持っており、進化距離も短いことから、共通祖先で持っていた ORF を現在も受け継 いで維持していると考えられる。 cox1 遺伝子内の第 3 イントロンについて(図 9) France-I 株で検出されているこのイントロンは、France-II、England、Ireland、Japan、Chile-I 株 で検出されている。全てのイントロンは完全なORF を持ち、相互に進化距離が短かった。従っ て、これらの共通祖先から完全なORF を持つこのイントロンが現在まで受け継がれ、維持され ていると考えられる。
この種内多型解析から、France-I 株の LSU rDNA の第 1 イントロンと Japan 株の LSU rDNA の 第4 イントロンが比較的最近侵入してきたことが明らかになった。
このイントロンのORF 配列を NCBI の BLAST 検索で相同性のある配列を見出した。共生関係 にあるような水カビなどがgroup II イントロンを持っており Pylaiella littoralis の group II イント ロンとクラスターを作り進化距離も短ければ、同じイントロンが挿入されていることの証明にな り、種を超えた転移機構の解明に役立てるデータが得られると考えられた。
.
種外多型解析による結果
France-I 株の LSU rDNA 内第 1 イントロンについて(図 10)
France-I 株の LSU rDNA 内の第 1 イントロンの ORF と相同性のある領域を NCBI を用いて検 索した。相同性が高くスコアの上位に来るものは、ほとんどバクテリア類だが、系統樹を描いて みると違うクレイドに分けられ、高いBootstrap 値で支持されバクテリア類の group II イントロン とPylaiella littoralis に見られる group II イントロンが別物だと言う事が証明された。スコアが低
くても同じようにLSU rDNA に挿入されている group II イントロンとクレイドを組むことが示さ れたが、第1 イントロンの起源となったイントロンは見つからず、種外から転移してきたと証明 できるデータは示されなかった。
図10 では相同性が高いが系統樹で France-I 株のイントロン ORF とクレイドを組まないバク テリア類は大部分除外している。
Japan 株の LSU rDNA 内第 4 イントロンについて(図 11)
France-I 株のときと同様に、Japan 株の LSU rDNA 内の第 4 イントロンの ORF と相同性のある領 域を NCBI を用いて検索した。France-I 株と同様に相同性が高くスコアの上位に来るものは、バ クテリア類だが、イントロンの近縁関係は見られなかった。ここでは、Chile-II 株の第 4 イント ロンとクレイドを形成した。このことから、Pylaiella littoralis のイントロン起源となっているも のは同一種であり、再び同じイントロンが再侵入してきているものだと考えられた。 図 11 では図 10 と同じく相同性が高いが系統樹で Japan 株のイントロン ORF とクレイドを組ま ないバクテリア類は大部分除外している。
5. 考察
種内多型解析産地で group II イントロンに多型が見られた。これは、Pylaiella littoralis が固着性の褐藻であ ることと、無性生殖で増殖していることが関係し、イントロン多型に更なる拍車をかけているも のだと思われる。
Japan 株の LSU rDNA 内の第 4 イントロンと France-I 株の LSU rDNA 内の第 1 イントロンは 最近侵入してきたイントロンであることが種内多型の分子系統樹で示された。
種外多型解析
Pylaella littoralis の group II イントロンが同一種からの侵入であることが示唆されたが、デー
タベースには起源となった group II イントロンを持った生物種は確認できなかった。一番イント ロンのやり取りをしている可能性が高い、共生生物である水カビが Pylaiella littoralis のイントロ ンとクレイドを形成してくれれば、種外から転移してきた証明になったが、そのようなデータは 見られなかった。データベースには group II イントロンのデータはかなり蓄積されている。種間 転移のメカニズムを解明するには、広い生物種のオルガネラのデータが必要であるが、現状では 十分でなく、その機構は不明である。
6. 図表
図1.France-I France の論文で示されている group II イントロンの配置
表1.世界中から採集された Pylaiella littoralis の採取場所
採取場所 海域 略称
Roscoff, France Atlantic Ocean France-II Mulroy Bay, Ireland Atlantic Ocean Ireland Shetland Is., England North Sea England Helgoland Is., Germany North Sea Germany Hokkaido, Japan Pacific Ocean Japan St. Lawrence Is., USA Bering Strait USA Diego Ramirez Is., Chile Drake Passage Chile-I Puyuhuapi, Chile Pacific Ocean Chile-II
図3.Pylaiella littoralis の LSU rDNA 遺伝子内第 1 イントロンの種内多型解析
図4.Pylaiella littoralis の LSU rDNA 遺伝子内第 2 イントロンの種内多型解析
図5.Pylaiella littoralis の LSU rDNA 遺伝子内第 3 イントロンの種内多型解析
LSU rDNA intron2
図6.Pylaiella littoralis の LSU rDNA 遺伝子内第 4 イントロンの種内多型解析
図7.Pylaiella littoralis の cox1 遺伝子内第 1 イントロンの種内多型解析
図7.Pylaiella littoralis の cox1 遺伝子内第 2 イントロンの種内多型解析
cox1 intron1
図9.Pylaiella littoralis の cox1 遺伝子内第 3 イントロンの種内多型解析
cox1 intron3
図10.France-I 株の LSU rDNA 内の第 1 イントロンの種外における分子系統解析
表2.France-I 株の LSU rDNA 内の第 1 イントロンの種外分子系統樹作成に用いた種 Species name Organism Class Accession Number
Saccharomyces cerevisiae (S.c.) intron-1 Yeast V00694 Saccharomyces cerevisiae (S.c.) intron-2 Yeast V00694
Euglena myxocylindracea (Eu.m.) Euglenoid AY290861
Chlamydomonas sp. CCMP 1619 (Ch.sp.) Chlorophyta AY325305
Scenedesmus obliquus strain UTEX 393 (Sc.o) Chlorophyta DQ396875
Bacteroides thetaiotaomicron VPI-5482 (Ba.t.) Bacterial AE015928
Escherichia coli O157:H7 plasmid pO157 (E.c.) Bacterial AF074613
Azotobacter vinelandii (Az.v.) Bacterial AY057439
図10.France-I 株の LSU rDNA 内の第 1 イントロンの種外における分子系統解析
表3.Japan 株の LSU rDNA 内の第 4 イントロンの種外分子系統樹作成に用いた種
Species name Organism Class Accession Number
Porphyra purpurea (P.p.) intron-1 Red alga AF114794
Porphyra purpurea (P.p.) intron-2 Red alga AF114794
Physcomitrella patens (Ph.pa.) intron-1 Streptophyta AB251495
Physcomitrella patens (Ph.pa.) intron-2 Streptophyta AB251495
Euglena myxocylindracea (Eu.m.) Euglenoid AY290861
Chlamydomonas sp. CCMP 1619 (C.sp.) Chlorophyta AY325305
Trichodesmium erythraeum IMS101 (T.e.) Cyanobacteria CP000393
Gloeobacter violaceus PCC 7421 (G.v.) Cyanobacteria BA000045
7. 参考文献
Zimmerly S, Hausner G, Wu X. Phylogenetic relationships among group II intron ORFs. Nucleic Acids Research. 2001; 29: 5.
Delaroque N, Maier I, Knippers R, Müller DG. Persistent virus integration into the genome of its algal host, Ectocarpus siliculosus (Phaeophyceae). J Gen Virol. 1999; 80: 1367-1370
Ehara M, Watanabe KI, Ohama T. Distribution of cognates of group II introns detected in mitochondrial cox1 genes of a diatom and a haptophyte. Gene. 2000; 256: 157-167
Fontain JM, Rousvoal S, Leblanc C, Kloareq B, Loiseaux-de Goër S. The mitochondrial LSU rDNA of the brown alga Pylaiella littoralis reveals alpha-proteobacterial features and is split by four group IIB introns with an atypical phylogeny. J Mol Biol. 1995; 251: 378-389
Fontain JM, Goux D, Kloareq B, Loiseaux-de Goër S. The reverse-transcriptase-like proteins encoded by group II introns in the mitochondrial genome of the brown alga Pylaiella littoralis belong to two different lineages which apparently coevolved with the group II ribosyme lineages. J Mol Evol. 1997; 44: 33-42
Oudot-Le Secq MP, Fontaine JM, Rousvoal S, Kloareq B, Loiseaux-de Goër S. The complete sequence of a brown algal mitochondrial genome, the ectocarpale Pylaiella littoralis (L.) Kjellm. J Mol Evol. 2001 53; 80-88
謝辞
本研究を始めるにあたり、ご指導ご指摘いただいた高知工科大学 工学部 大濱武教授に深く感 謝いたします。また緑藻 KS-1 株を譲っていただいた関西電力株式会社 電力技術研究所 環境技 術センターの宮坂均史と Pylaiella littoralis のデータを提供して頂いた大阪教育大学 理科教育講 座の生田享介准教授に厚く御礼申し上げます。日本で採集された
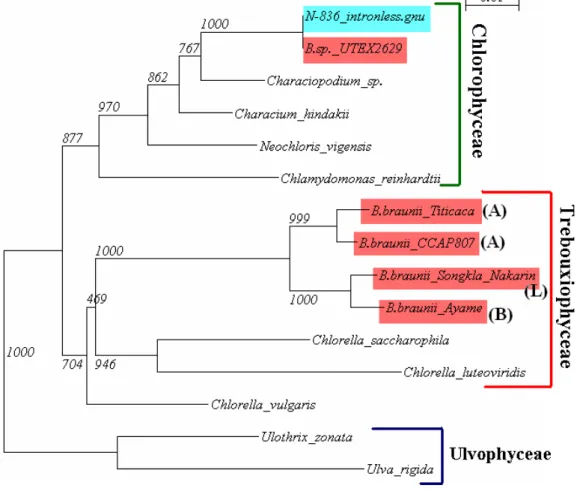
Botryococcus braunii N-836 株の chemical race の分子系統解析
Botryococcus braunii は炭化水素生産能力を持つ単細胞緑藻で、培養中の細胞濃度が高くなると、
相互に付着し直径 30 500_m ほどのコロニーを形成し、生育する。細胞間物質として大量の炭 化水素を保持しており、原油の一部はこの藻が生産した炭化水素が元になっていると考えられて いる。これまでに発見されているBotryococcus braunii は生産する炭化水素の違いによって A・B・
L の 3 系統に分類されている(chemical race)。また、chemical race 毎にその 18S rDNA 配列は、 特徴的な変異を持っている。 日本で発見され、現在国立環境研究所が保管している N-836 株について、chemical race が不明 である。そこで、この株の 18S rDNA を PCR 法によって増幅し、配列を決定し、分子系統解析 を作成することでchemical race を決定した。 ゲノムDNA の回収・分子系統樹作成方法 上記[1]Chlorella saccharophila と同定された緑藻 KS-1 株の分子系統解析の 3 章-材料と方法の項 参照。 表1. 18S rDNA 増幅用プライマー ①B.braunii_18S_FP1
5’ - TGC CAG TAG TCA TAT GCT TGT CTC A - 3’ ②B.braunii_18S_FP2
5’ - ACT ACT GCG AAA GCA TTT GCC A - 3’ ③B.braunii_18S_RP1
5’ - GAC GGT ATC TAA TCG TCT TCG - 3’ ④B.braunii_18S_RP2
5’ - CAA TGA TCC TTC CGC AGG TT -3’
上記のプライマーがアニールする18S rDNA 内のおよその位置を図 1 に示した。 図1. 18S rDNA とプライマーの位置関係 ②B.braunii_18S_FP2 ①B.braunii_18S_FP1 ③B.braunii_18S_RP1 5’ ④B.braunii_18S_RP4 3’
結果
_ と_ 、_ と_ 、_ と_ 全てにおいてシングルバンドが検出された。 その中でほぼ全領域を増 幅できた、_と_番のプライマー産物を WizardSV Gel and PCR Clean-Up System(Promega)で精 製し、pT7blue ベクター(TAKARA)に TA クローニングしたものを大腸菌(E.coli DH5_ Competent Cells(東洋紡))に導入した。形質転換した大腸菌を LB/amp/X-gal/IPTG 寒天培地上に撒き、カ ラーセレクションしたものをWizard Plus Minipreps DNA Purification System(Promega)で精製し、 サンガー法により挿入DNA 断片の塩基配列を決定した。
その結果、18S rDNA 領域を前半部分 787bp、後半部分 817bp 得た。
得られたデータをNCBI で BLAST 検索すると、Botryococcus sp. UTEX 2629 (AJ581914)との相 同性が100%となり、N-836 株と Botryococcus sp. UTEX 2629 の塩基配列を比較するとイントロン を含め全て同じ配列であり、Botryococcus braunii N-836 株は Botryococcus sp. UTEX 2629 と同一種
であることが明らかになった。 図2. 得られた領域およその位置関係 シークエンシング結果 18S rDNA 前半部分(787bp) ※Primer の箇所は赤で示した。青文字はイントロン領域を示す。 TGCCAGTAGTCATATGCTTGTCTCAAAGATTAAGCCATGCATGTCTAAGTATAAACTGCTTATACT GTGAAACTGCGAATGGCTCATTAAATCAGTTATAGTTTATTTGATGGTACTTCTTACTCGGATAAC CGTAAGAAATTTAGAGCTAATACGTGCGTAAAACCCGACTTCTGGAAGGGTCGTATATATTAGAT GAAAGGCCGACCGGACTTTGTCCGACCCGCGGTGAATCATGATATCTTCACGAATCGCATAGCC TTGTGCTGGCGATGTTTCATTCAAATTTCTGCCCTATCAACTTTCGATGGTAGGATAGAGGCCTA CCATGGTGGTAACGGGTGACGGAGGATTAGGGTTCGATTCCGGAGAGGGAGCCTGAGAAACG GCTACCACATCCAAGGAAGGCAGCAGGCGCGCAAATTACCCAATCCTGACACGGGGAGGTAGT GACAATAAATAACATTTCCGGGCACATCGTGTCTGGAAAATGGAATGAGTACAATATAAATCCCT TAACGAGTATCAATTGGAGGGCAAGTCTGGTGAACATTCAACGGAATGTGCTGTTGACGCCAG AGATAGTGGGGCAGCGTACGCTGTTATGCCTGCTAGTCGAGCAACCACAATGCACAAGGAATG GGTGGGTTGCCGGCAAGACGACCTGGTACGGGGAAGGCTAAGTTGCAGACTCTGCAATATGCT AATCCCGTGGTGAGCTTGTGAAGGGTGACTTTCACACAGCCATCGTAACGCACGGTAAGGCGT CGGCTGACTCCTGTGAGAG 18S rDNA 後半部分(817bp) ※Primer の箇所は赤で示した。青文字はイントロン領域を示す。 TTAGAGGGACTATTGGCGTTCTAGCCAATGGAAGTATGAGGCAATAACAGGTCTGTGATGCCCT TAGATGTTCTGGGCCGCACGCGCGCTACACTGATGCGTTCAACGAGCCTATCCTTGACCGAGAG ①B.braunii_18S_FP1 5’ 前半領域787bp 後半領域817bp ④B.braunii_18S_RP4 3’
GTCCGGGTAATCTTTGAAACCGCATCGTGATGGGGCTAGACTATTGCAATTATTAGTCTTTAACG AGGAATGCCTAGTAAGCGCGATTCATCAGATCGCGTTGATTACGTCCCTGCCCTTTGTACACACC GCCCGTCGCTCCTACCGATTGAGTGTGCTGGTGAAGTGTTCGGATCGGCAAAGCTGGGGGCAA CTCTTGCATTGCCGAGAAGTTCATTAAACCCTCCCACTTAGAGGAAGGAGAAGTCGTAACAAG GTTTCCGTATTATCACCATGCGGAAGTCAGCCTTAAAAGGGCTGGCTAGTGGGTTGCCCAGGCA ACCTGCAAGACTGTCAAATTGCCGGAACCTCCTGAATGCTGTTGGTACCGCTGGCTGTGGGAA ACCATGGCCCAGCACCGTAGGGAAACTTGCGGGTATGGTAACAATCCAACAGATAGGGACTATC GGCAGCCAAGTCCTAAGGGCACCTGCCTATGGATGCAGTTCACAGACTAAATGGCAGTCGGCC TGCAATGTGTGCAGGCTTAAGATATAGTCGGGCCCTACCGAGAGGTAGCCTGTCAGAGGAAGC TGAATCAAGCTGAGAGCTGATAGGAAGGGTGGTAATACTTCATGCTGCAAGGCATTGAAGGGC TGTAATGCCTCACCCCTTGGAACAACCGAGGTGAACCTGCGGAAGGATCATTG
図3. Botryococcus braunii の chemical race 分子系統樹
N-836 株のシークエンシングの結果から、イントロン領域とまだ不明な領域が存在するが、そ の部分は分子系統樹を作成する配列中には含めなかった。
表2.Botryococcus braunii の chemical race 分子系統樹作成に用いた塩基配列のデータ Species name chemical race Accession Number
Botryococcus braunii CCAP 807/1 A AJ581913
Botryococcus braunii Titicaca A AJ581912
Botryococcus braunii Songkla Nakarin L AJ581911
Characium hindakii M63000 Chlorella luteoviridis X73997 Chlorella saccharophila X63505 Chlorella vulgaris X13688 Neochloris vigensis M74496 Ulothrix zonata Z47999
Ulva rigida AJ005414
参考文献
Senousy HH, Beakes GW, Hack E. Phylogenetic placement of Botryococcus braunii
(Trebouxiophyceae) and Botryococcus sudeticus isolate UTEX 2629 (Chlorophyceae). J Phycol. 2004; 40: 412-423