マウス肺障害モデルにおける exosome 内の miRNA 発現の変動とその役割について
日本大学大学院医学研究科博士課程 内科系呼吸器内科学専攻
鹿野 壯太郎
2017 年
指導教員 橋本 修
マウス肺障害モデルにおける exosome 内の miRNA 発現の変動とその役割について
日本大学大学院医学研究科博士課程 内科系呼吸器内科学専攻
鹿野 壯太郎
2017 年
指導教員 橋本 修
目次
Ⅰ . 概 要 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・1
Ⅱ . 緒 言 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・3
Ⅲ.材料と方法 ・・・・・・・・・・・・・・・・・・・・・・・・9 1) 動物実験及び exosome の抽出・・・・・・・・・・・・・・・・9 2) 網羅的遺伝子解析・・・・・・・・・・・・・・・・・・・・・・・10 3) リ ア ル タ イ ム P CR・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・11 4) 細胞培養・・・・・・・・・・・・・・・・・・・・・・・・・・12 5) 遺 伝 子 導 入・ ・ ・ ・・ ・ ・ ・ ・・ ・ ・ ・・ ・ ・ ・ ・・ ・ ・ ・・12
6) ELISA・・・・・・・・・・・・・・・・・・・・・・・・・・13
7) Western blot ・・・・・・・・・・・・・・・・・・・・・・・・・13 8) 統計解析 ・・・・・・・・・・・・・・・・・・・・・・・・・・14
Ⅳ.結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・14 1) 経気道 LPS 投与によるマウス BALF 中 exosome の変化・・・・・14
2) miRNAマイクロアレイを用いた網羅的遺伝子解析 ・・・・・・・・15
3) miRNA-466gとmiRNA-466m-5pの発現レベルのPCR法による検証16 4) miRNA-466g と miRNA-466m-5p のIL-1β産生経路への影響・・・16
Ⅴ.考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・17
Ⅵ.まとめ ・・・・・・・・・・・・・・・・・・・・・・・・・・・20
Ⅶ.謝辞 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・21
Ⅷ.図表 ・・・・・・・・・・・・・・・・・・・・・・・・・・22
Ⅸ.図表説明 ・・・・・・・・・・・・・・・・・・・・・・・・・・33
Ⅹ.引用文献 ・・・・・・・・・・・・・・・・・・・・・・・・・・36
Ⅺ.研究業績 ・・・・・・・・・・・・・・・・・・・・・・・・・・43
1
Ⅰ.概要
急性呼吸窮迫症候群 (acute respiratory distress syndrome, ARDS)は、重症 肺炎や敗血症などが誘因となり全身性の炎症が引き起こされ、肺血管の透過性 亢進により急性肺水腫を発症し、急性呼吸不全を呈し予後は不良である。
ARDS の発症には炎症性サイトカインの過剰な産生による肺血管傷害と好中 球の集簇が関係すると考えられており、治療には好中球エラスターゼ阻害薬が 使用されている。しかし効果は部分的であり、好中球以外の炎症を制御する必要 があると考えられている。そこで我々はARDS におけるマクロファージを中心 とした炎症のメカニズムに注目し、Nod (nucleotide-binding oligomerization)- like receptors 3 (NLRP3)インフラマソームとその制御が報告されているエクソ ソーム (exosome)について検討した。
Exosomeとは脂質二重膜で構成され、内部に核酸やタンパクを含む小胞で、
ドナー細胞からのエキソサイトーシスにより細胞外に放出され、レシピエント 細胞にエンドサイトーシスにより取り込まれることで、細胞間の情報伝達に関 与していると考えられている。Exosomeに含まれる情報成分の中でも、特に近 年microRNA (miRNA)についての研究が進められている。miRNAは約20ヌク レオチドで構成される小さな RNA であるが、相補的配列をもつターゲット messenger RNA (mRNA)に結合し、分解または翻訳抑制を行い、タンパク質の
2
産生を抑制することが知られている。 本研究は、マウス肺障害モデルの肺胞洗 浄液 (bronchoalveolar lavage fluid, BALF)を用い、exosome内miRNAの解析
とNLRP3インフラマソームへの制御機構を明らかにすることによりARDS に
対するexosomeの役割を検討した。
Lipopolysaccharide (LPS)をマウスの気道に投与して急性肺障害モデルを作 製し、BALF中のexosome とその中に含まれるRNAについて解析を行った。
LPS 刺激後も exosome 数に変化はみられなかったが、RNA量の著明な増加を
認めた。miRNAの網羅解析においては、miRNA-466ファミリー (miRNA-466g, miRNA-466m-5pなど)がLPS刺激により有意に増加していた。
miRNA-466g と miRNA-466m-5p をマウス骨髄由来マクロファージ (bone marrow derived macrophage, BMDM)に遺伝子導入し、LPSとadenosine 5’- triphosphate disodium salt hydrate (ATP)投与によるinterleukin-1β (IL-1β) の分泌への影響を検討した。miRNA-466gとmiRNA-466m-5pを遺伝子導入し たBMDMはコントロールmiRNA処理した細胞と比較し、細胞内でのpro-IL-
1β増加と、細胞外への IL-1β分泌の増加を認めた。以上から ARDS では、気
道に分泌される exosome 内の miRNA-466g と miRNA-466m-5p が増加し、
NLRP3 インフラマソームを介した IL-1βの過剰な産生が起き、病態が悪化し
ている可能性が示唆された。
3
Ⅱ.緒言
ARDS は、全身性の高度な炎症に伴って肺胞隔壁の物質透過性が亢進するこ とにより生じる非心原性肺水腫である1)。ARDSの概念は、1967年にAshbaugh らが、重症外傷・ウイルス感染症・急性膵炎の患者12名が、呼吸不全の既往が ないにも関わらず、罹患して 1~96 時間後に急性呼吸不全を発症したことを報 告したことに始まる2)。1971年に Pettyらは、上記の病態をARDS と命名し、
肺への直接または間接的な傷害が誘因となりびまん性肺障害が発症する ARDS は、人工呼吸器で持続的陽圧換気 (positive end-expiratory pressure, PEEP)を 使用しながら呼吸管理することが治療に有効であると報告した3)。その後、1988 年にMurrayらは肺損傷スコア (lung injury score, LIS)を作製してARDSの定 義を拡大し、肺損傷が軽度の場合を急性肺障害 (aute lung injury, ALI)とするこ とを提唱した4)。1994年には、米国胸部学会 (American Thoracic Society, ATS) とヨーロッパ呼吸器学会 (European Respiratory Society, ERS)が合同で①発症 時期、②酸素化能、③胸部画像所見、④心不全の否定の 4 項目をもとに ARDS の診断基準を作成し5)、2011年に重症度分類と一部の項目で改定がなされ、2012
年にBerlin定義として診断基準が改善され6)、今日の医療に用いられている。
ARDSの発症率は、2014年に世界50ヵ国459カ所のICUで搬送患者を対象 としたコホート研究で、全患者29144名のうち3022名 (10.4%)がBerlin定義
4
にもとづくARDSの診断基準を満たし、重症度は軽症30%、中等症46.6%、重 症23.4%で、それぞれの死亡率は、軽症34.9%、中等症40.3%、重症46.6%で あったと報告されている7)。
ARDS の原因は直接損傷と間接損傷に分類されるが、原因の違いによる死亡 率の差は認められていない8)。直接損傷の原因は重症肺炎や誤嚥性肺炎などが多 いとされており、間接損傷では敗血症やショックを伴う外傷が多く、なかでも敗 血症はARDSの原因として全体の40%を占めている。敗血症は全身性炎症反応 症候群 (systemic inflammatory response syndrome, SIRS)のなかで感染症を 伴うものと定義される。SIRSの病態においては、炎症性サイトカインが過剰産 生され、ヒトの体内で多臓器機能不全症候群 (multiple organ dysfunction
syndrome, MODS)を引き起こすことが知られており、ARDSもその一部ではな
いかと考えられている9)。
ARDS の発症機序に関しては主に以下の可能性が考えられている。1)活性化 されたマクロファージからIL-1βやinterleukin-8 (IL-8)などの炎症性サイトカ インが過剰産生され、刺激を受けた好中球は肺の微小血管を通過しにくくなり、
また肺血管内皮で接着分子が発現増加するため、結果的に好中球の肺血管内へ の集簇が起こる。2)さらに、好中球はマクロファージから放出された走化性因子 の影響を受け肺間質や肺胞腔内に遊走し、そこで好中球エラスターゼや活性酸
5
素などを産生するため肺胞上皮細胞を傷害する10, 11)。
現在、ARDS の薬物療法は好中球エラスターゼ阻害薬などが主体で、その効 果は部分的であり、新たな治療法に関する研究が進められているが十分な成果 は得られていない。ARDS の新たな治療には好中球以外の炎症制御が必要と考 えられる。そこで我々は、マクロファージを主体とした炎症に関与するNLRP3 インフラマソームに注目した。
NLRP3インフラマソームとは、マクロファージ内で、プロテアーゼの一種で
ある caspase-1 の活性化により IL-1βの分泌を誘導し、炎症を引き起こすタン
パク質複合体である。インフラマソームは病原体関連分子パターン (pathogen- associated molecular pattern, PAMP)やダメージ関連分子パターン (damage- associated molecular pattern, DAMP)をマクロファージのToll like receptor 4 (TLR4)が認識して pro-IL-1βを産生する priming と、組織障害により danger
signalとして細胞外に放出されたATPをマクロファージのP2X7受容体が認識
してインフラマソームである NLRP3 を介して caspase-1 が活性化し、その結 果 と し て caspase-1 に よ り pro-IL-1β が 切 断 さ れ 、IL-1β が 産 生 さ れ る triggeringにより炎症を引き起こす12-16) (図1)。
TamasらはMICUに搬送された敗血症にARDSを合併した患者の血清IL-1 βが上昇し、このARDS は人工呼吸器に誘発されたものとする研究を報告し、
6
ARDS にインフラマソームが関与していることを示唆した16)。しかし、呼吸器 感染症によるARDSとインフラマソームについては不明な点が多く、その制御 機構を明らかにできれば新たな治療標的になると考えられる。呼吸器感染症モ デルを作製するに当たり、一般的に市中肺炎では肺炎球菌・インフルエンザ桿菌 などが起因菌として多いが、本研究では NLRP3 インフラマソームが TLR4 を 介した炎症経路であることから、TLR4 に特異的に結合するE.coli 由来の LPS による急性肺障害をARDSモデルとして実験を行うこととした17)。
近年、細胞間の情報伝達手段としてexosomeが注目されている。興味深いこ
とに exosome がインフラマソームを制御していることが報告されている 18)。
Exosome とは体液中に存在する脂質二重膜で覆われた直径 40~100nm の小胞
で、体内の様々な細胞内で多面小胞体から出芽することにより産生され、エキソ サイトーシスにより細胞外に放出されると考えられている。Exosome内には主 にmRNAや、miRNAなどのnon coding RNA、タンパクが含まれており、外 部は脂質とタンパクで構成された膜で覆われた構造をしている 19,20) (図 2)。
Exosome の存在は古くから知られ、1983 年に Johnstone らはヒツジ網状赤血 球の培養中に細胞外小胞が存在することを発見し、これをexosomeと命名した
21)。その後、1983 年にHarding らとPan らが抗トランスフェリン受容体抗体
を 投 与 し た 網 赤 血 球 を 免 疫 電 子 顕 微 鏡 で 観 察 し 、 細 胞 内 で 産 生 さ れ た
7
multiplevesicular endosomeがexosomeとして細胞外へ放出されることを確認
している22,23)。
Exosomeが生物情報のバイオソースとして近年注目を集めるようになったの
は、2007年に Rotval らがマウスの肥満細胞のexosome 内マウスmRNA をヒ トの肥満細胞に移行させ、マウスのタンパクがヒト細胞で産生されたことを報 告したことによる19)。この実験から、exosomeがmRNAを細胞間で運搬し、レ シピエント細胞に取り込まれることで、メッセンジャーとして作用している可 能性が示唆された。また吉岡らは担癌患者の血液中に含まれる細胞外小胞を解 析し、癌由来の小胞を特定する技術 (liquid biopsy)を 24)、Carla らは好酸球が
IFN-γ刺激によりexosomeの分泌を増加し、さらに気管支喘息患者では健常者
と比較して exosome の分泌が増加することを 25)報告しており、exosome の
biomarkerとしての利用について研究が進められている。
miRNA とは、生物中に存在する 21~25 ヌクレオチドで形成された小さな
RNAである。ターゲットとする特定の mRNAに結合し、分解または翻訳抑制 することでタンパク発現を制御し生体のホメオスターシスに関与している 26,27)。 初めに発見された miRNA は、1993 年に Lee らが報告した C.elegans が持つ 22ヌクレオチドの非コードRNA鎖Lin-4である。Lin-4は、C.elegansの発生 に関与するLin-14の mRNAの3’-untranslated region (UTR)と相補的に結合
8
することが判明し、さらにLin-14の働きを制御することが示された28,29)。
miRNAの生成過程は次のように考えられている。1)細胞核内にコードされ
ている miRNA 遺伝子が転写され、1 回目のプロセシングを受けて miRNA 前
駆体が生成される。2)次に輸送タンパクにより核外に運ばれ、そこで2回目のプ ロセシングを受けて成熟miRNA二量体となる。3)その後、一方のRNAは除去 され、残ったRNAが複合体RNA-induced silencing complex (RISC)を形成し、
相補的な配列をもつ mRNA と結合することで分解または翻訳抑制に働いてい
る30,31)。一部のmiRNAはexosomeに取り込まれて細胞外に放出され、レシピ
エント細胞に運搬された後にエンドサイトーシスにより吸収されることが報告 されている 32)。Exosome に含まれる miRNA についてはその働きが注目され、
traffickingやsorting機能について今日研究が進められている33)。
近年、気管支喘息、慢性閉塞性肺疾患、肺線維症、嚢胞線維症、肺癌などの呼 吸器疾患を持つ患者の肺組織、肺胞洗浄液、血液、喀痰中のmiRNAをbiomarker として応用しようとする研究が進められている34)。LPS誘導性マウス肺障害モ デルの体液や肺組織中の miRNA 発現とその機能解析を行い、さらに発現の亢
進したmiRNAを肺上皮細胞株に遺伝子導入しLPS刺激による遺伝子発現の変
化を評価することにより、ARDS の発症と miRNA との関連について検討がな されている35)。
9
本研究はLPSを経気道投与したマウス肺障害モデルを用いて、そのBALF中
にあるexosome内miRNAの発現を網羅的に解析し、増加するmiRNA をマウ
ス骨髄由来マクロファージに遺伝子導入して NLRP3 インフラマソームへの影 響を評価することで、ARDS の発症における NLRP3 インフラマソームを介し た炎症経路に対する BALF 中 exosome の役割について検討した。その結果、
LPS 刺激により気道に分泌された exosome 内では miRNA-466g と miRNA-
466m-5pの発現が亢進し、このmiRNA がマクロファージのNLRP3 インフラ
マソームに作用することでIL-1β産生を増加させてARDSの発症に関与してい る可能性が示唆された。現在まではARDS治療は好中球の炎症制御を主体とし ていたが、本研究結果からexosomeによるマクロファージのNLRP3インフラ マソームの炎症を制御することが新たな治療標的になると考えられた。
Ⅲ.材料と方法 1) 実験動物及びexosomeの抽出
C57BL/6JマウスはCharles river (Yokohama, Japan)から、E.coli由来のLPS はSIGMA-Aldrich (St. Louis, Missouri)から購入した。8週齢、雄のC57BL/6J マウスを準備し、2群 (各n=5)に分けてそれぞれ50µLのPBSと1µg/µL,50µL
のLPS溶液を2%イソフルラン吸入により麻酔後に経気道投与し、24時間後に
10
PBS1mLで1回肺を洗浄しBALFを回収した。BALFは遠心分離機で4℃, 500
×g, 10分間遠心し、上清を-80℃で保存した。後日、Sera MirTM Exosome RNA Amplification Kit (System Biosciences, Palo Alto, California)のプロトコール に準じて、exosomeとRNAを抽出した。抽出したexosomeは走査電子顕微鏡 で観察し、その数をナノ粒子カウンターqNano (メイワフォーシス, Tokyo, Japan)で カ ウ ン ト し た 。RNA は Agilent RNA6000 pico kit (Agilent Technologies, Santa Clara, California)でRNA量と大きさを測定した。LPS投 与量については、マウス1匹対して60µgのE.coli 由来のLPSを経気道投与す ることにより NLRP3 インフラマソームを介した肺障害モデルを作製したとの 報告がされているが 17)、本研究ではマウス 1 匹に対して 50µg の経気道投与で も BALF 中に著明な好中球の増加を認めたことから、同量でマウス肺障害モデ ルを作製した。
2) 網羅的遺伝子解析
8 週齢、雄の C57BL/6J マウスを準備し、PBS 投与群 (n=4)と LPS 投与群 (n=4)に分け、LPS 投与後に exosome 内で変動する miRNA の同定を試みた。
但し、PBS群はRNA量を確保するため10匹分のサンプルをプールし1検体と して使用した。前述の方法で exosome と RNA を回収し、Affymetrix Flash
11
TaqTM Biotin HSR RNA Labeling Kits for Gene Chip miRNA Array (Affymetrix, Santa Clara ,California) と Gene Chip® miRNA3.0 Array(Affymetrix, Santa Clara ,California)を 用 い て 解 析 し た 。Genechip Fluidics Station 450, Genechip Scanner 3000 (Affymetrix, Santa Clara ,California)を用いて蛍光強度を数値化し、得られた遺伝子データを Partek Genomics Suite (Partek Inc, St. Louis, Missouri)を用いて統計学的解 析を行い、LPS投与により変動したmiRNA遺伝子を同定した。
3) リアルタイムPCR解析
マウスBALF中のexosomeより抽出したtotal RNAをNanoDrop (スクラム,
Tokyo, Japan)で濃度測定し、TaqMan MicroRNA Reverse Transcription Kit (Applied Biosystems, Foster City, Carifornia)を用いて complementary DNA (cDNA)を合成し、Applied Biosystems 7300 standard Real- Time PCR System (Applied Biosystems)を用いてリアルタイムPCR解析を行った。またTaqMan primer は miRNA-466g (Assay ID 241015_mat, Applied Biosystems)と miRNA-466m-5p (Assay ID 465329_mat, Applied Biosystems)、内在性コント ロールにはU6 (Assay ID 0019173, Applied Biosystems)を用いた。Ct値は初 期閾値で決定し、サンプルの RNA 量は比較定量法 (⊿⊿CT 法)を用いて評価
12
した。
4) 細胞培養
骨髄由来マクロファージ (bone marrow-derived macrophage, BMDM)は、
C57BL/6Jマウスの両大腿骨と両脛骨から骨髄を抽出し、L929細胞 (American Type Culture Collection, Manassas, Virginia)の上清で分化させ使用した。L929 細胞の培養液上清は、骨髄抽出後1~2日目までは30%濃度で、3~7日目まで は 25%濃度で使用した。BMDM は、10%FBS (fetal bovine serum)(SAFC Biosciences, Lenexa, KS)、1%penicillin-streptomycin (nacalai tesque, Kyoto, Japan)を含有した dulbeco’s modified eagle’s medium-low glucose (SIGMA- Aldrich, St. Louis, Missouri)に L929 細胞の培養液上清を加えて 100%湿 度,37℃で培養した。
5) 遺伝子導入
培養7日目のBMDMに50nM miRNAをLipofectaminTM RNAiMAX (Life Technologies, Yokohama, Japan)のプロトコールに従って、24ウェルプレート 上でmiRNA mimic Negative Control #2 (Bioneer, Daedeok-gu, Korea)、MMU- MIR-466g mimic (Bioneer, Daedeok-gu, Korea)、MMU-MIR-466m-5p mimic
13
(Bioneer, Daedeok-gu, Korea)を遺伝子導入した。培養液は 10%FBS (SAFC Biosciences)と20%のL929上清を含んだdulbeco’s modified eagle’s medium- low glucose (SIGMA-Aldrich)を使用した。
6) ELISA
BMDMに miRNA を遺伝子導入して2 日後に、10ng/mLの LPSで刺激し、
その6時間後に5mM のATP (SIGMA-Aldrich, St. Louis, Missouri)で刺激し た。1時間後に培養液の上清を回収し、Mouse IL-1beta ELISA Ready-SET-GO!
(Affymetrix, Santa Clara, California)でIL-1βを測定した。また本研究で採用 したBMDMへのLPSとATPの投与量は、Franchiらの報告もあることから妥 当と考えられた36)。
7) Western blot
上 清 回 収 後 に 残っ た細 胞 を RIPA バ ッ ファ ー に よ り タ ンパ ク抽 出 し 、 Criterion XTゲル (4-12%T)Bis-Tris 18well (BIO-RAD, Hercules, California) を用いてSDSページを行い、一次抗体はgoat anti-mouse IL-1β(lone AF-401- NA)(R&D systems, Minneapolis, Minnesota)、二次抗体は donkey anti-goat IgG-HRP (sc2020)(Santa Cruz, Dallas, Texas)を使用して IL-1βの western
14
blot を行った。また、一次抗体はβ-actin antibody (#4967)(Cell signaling technology, Tokyo, Japan)、二次抗体はanti-rabbit IgG HRP-linked antibody (#7074)(Cell signaling technology, Tokyo, Japan)を 使 用 し て β-actin の western blotを行った。バンドはAmershamTM ECLTM Prime Western Blotting Detection Reagent (GE Healthcare, Tokyo, Japan)で発光させ、ルミノ・イメ ージアナライザLAS-4000IR (Fuji Film, Tokyo, Japan)で検出した。
8) 統計解析
デ ー タ は GraphPad Prism 6 (GraphPad Prism Software, La Jolla, California)で解析し、平均値±標準偏差値 (SD)または平均値±標準誤差値 (SE)で記した。群間の有意差は Student’s t 検定または ANOVA 検定と Tukey の多重比較検定を用いて、P<0.05を有意とした。
Ⅳ.結果
1) 経気道的LPS投与によるマウス BALF中exosomeの変化
はじめに気道内に分泌される exosome への LPS 刺激による影響を調べるた め、C57BL/6J マウスを用いて LPS の経気道的投与後のマウス BALF 中
exosome の大きさ、数の変化を測定した。走査電子顕微鏡における観察で、酢
15
酸ウラニル染色によって脂質二重膜で覆われた100~200nm 前後の粒子が回収
されたことを確認した (図3)。ナノ粒子カウンターで測定したexosome数は、
PBS群は平均7.4×10^8 particles、LPS投与群は平均 8.7×10^8 particlesで あり明らかな差を認めなかった (図4A)。
次に、exosomeからtotal RNAを回収し、バイオアナライザーで RNA量を 測定した。RNA 量は PBS 群では平均 32ng、LPS 群は平均 9162ng であり、
LPS 投与により RNA 量は約 280 倍に増加することが分かった (図 4B)。
exosome1個当たりのRNA量に換算しても、LPS群ではRNA量の増加を認め
た (図4C)。
以上からLPS 経気道投与により、マウスBALF 中exosome は数や形態的に 変化が認められないが、その内部の RNA 量は著しく増加することが分かった。
PBS 群と LPS 群のいずれから採取した exosome 内 RNA においても、バイ オアナライザーで測定したRNAサイズと量のヒストグラム50-100bp(図5A,B) の結果から、比較的小さなRNAを中心としていることが観察された。
2) miRNAマイクロアレイを用いた網羅的遺伝子解析
miRNA マイクロアレイを用いた網羅的遺伝子解析から得られた PBS 群と
LPS 群の miRNA 遺伝子発現パターンのヒートマップでは (図6)、平均よりも
16
発現亢進している遺伝子は赤色で、発現低下している遺伝子は緑色で表示して
いる。PBS群をコントロールとし、LPS投与群で発現比 (Fold change)が有意 (p<0.05)に上昇 (1.5倍以上)する遺伝子は23個であり、その中の上位10遺伝子 にはmiRNA-466ファミリーが多く含まれていた (表1)。一方、減少 (1.5倍以 上)する遺伝子は27個であった。
3) miRNA-466gとmiRNA-466m-5p発現レベルのPCR法による検証
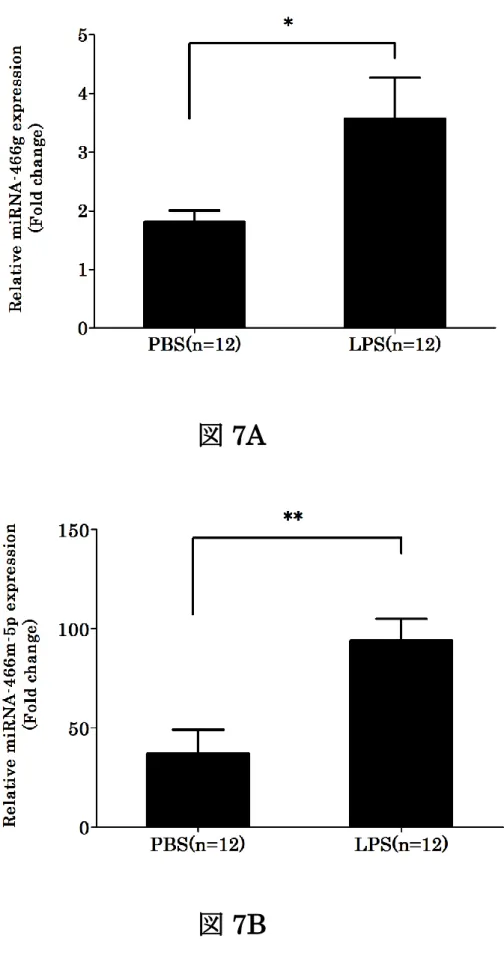
LPS 投与により発現上昇率が高い miRNA ファミリーの中で、リアルタイム PCRによるプライマー配列が確立されているmiRNA-466gと466m-5pに関し てリアルタイムPCRによる解析を行い、発現量がコントロールに比べてそれぞ れ約 2 倍の上昇がみられ miRNA マイクロアレイの結果と同じく遺伝子の発現 量が増加することが確認された (図7)。
4) miRNA-466gとmiRNA-466m-5pのIL-1β産生経路への影響
miRNA-466g または miRNA-466m-5p を遺伝子導入した群では、コントロ ール群と比較して上清中 IL-1β濃度の増加を認めた (図 8A)。次に IL-1βの western blotにより、細胞内のpro-IL-1β量について観察した。LPS刺激によ り 31kDa の pro-IL-1βが上昇し、ATP 添加により pro-IL-1βが減少すること
17
が観察された (図8B)。
Ⅴ.考察
一般的にARDSの発症には活性化した好中球による炎症が主な要因と考えら れる。しかし、ARDS に対して好中球エラスターゼ阻害薬による治療では効果 が不十分であるため、他の炎症経路についても解析を試みる必要がある。そこで、
我々はARDSの発症にマクロファージの NLRP3インフラマソームを介した炎 症の関与について注目し、その制御を担うと考えられているexosomeについて 解析を行った。マウスへのLPS 刺激によりBALF 中exosome内ではmiRNA- 466 ファミリーの発現が亢進し、さらにこの中で発現の確認がとれた miRNA- 466gとmiRNA-466m-5pをBMDMに遺伝子導入するとNLRP3インフラマソ ームを介してIL-1βの産生を亢進することが判明した。以上から、LPS刺激に より気道に分泌されたexosomeはNLRP3インフラマソームに作用することで ARDSの発症に関与している可能性があると考えられた。
まず LPS 投与によるマウス肺障害モデルで、LPS 刺激による BALF 中の
exosome数とRNA量を比較した実験を行った。Exosome数に有意差は認めら
れなかったものの、LPS投与後にexosome内のRNAが著しく増加することが 観察された。この結果から、LPS刺激は RNAをexosome内にsorting するプ
18
ロセスに影響を及ぼすことを示している。バイオアナライザーによる解析結果 では、無刺激のBALF中exosome内RNAは、従来報告されている他の体液同 様に、50-200bpを中心とした小分子 RNAであることが分かった。また、LPS 刺激により劇的に増加するRNAも、200bp以上にもピークがみられるが、主と
して50-200bpに大きなピークを認め、やはり小分子RNAを中心に刺激により
exosome内にsortingされることが分かった。この結果は一般的に、exosome内 の RNAには miRNA などの non-coding RNA が多く含まれていることと一致 しており、LPS による RNA 上昇は miRNA を中心としている可能性が示唆さ れた。LPS刺激によるexosome内におけるmiRNAの劇的な増加は、exosome が内包する情報量を増加させ、病原微生物に対する生体応答を制御するのに必 要である可能性がある。また本研究ではexosome抽出キットを使用してBALF
からexosomeを抽出したが、exosomeに付着したRNAの混入も微量と考えら
れるが否定できない。今後、より高精度にexosomeを抽出できればさらに正確 なRNA情報を得ることが可能と考える。
LPS刺激によりexosome内で増加するRNAがmiRNAを主体とする可能性 があることから、次にmiRNAの網羅的発現解析を行った。これにより、LPS刺
激でmiRNA遺伝子発現が有意に増加する23個の遺伝子を同定した。興味深い
ことに、その中の上位10遺伝子には、miRNA-466 ファミリーが多く含まれて
19
いた。LPSとmiRNAに関するこれまでの報告では、Margaretらがマウス樹状 細胞をLPSで刺激し、細胞外に放出されたexosome内のmiRNAを解析した結 果、炎症を増悪させる miRNA-155 の発現が増加し、炎症を抑制する miRNA- 146aの発現が低下したことを報告している37)。しかし、LPSによる肺障害にお
けるmiRNA-466ファミリーの役割についてはこれまで明らかにされていない。
また miRNA-466 ファミリーについては、マウス腎臓皮質集合管細胞を用いた
実験でserum and glucocorticoid regulated kinase1 (SGK1)を抑制する働きが 報告されているのみで 38)、その機能の詳細については明らかでない。よって次 に我々は、miRNA-466 ファミリーの LPS による肺障害における役割について 検討した。その中でもマクロファージを介して炎症に寄与する NLRP3 インフ ラマソームに注目し、その機序に対するmiRNA-466の影響について検討した。
LPSによりNLRP3インフラマソームのprimingを介して産生されるpro-IL- 1βは、ATP による triggering を介して活性化した caspase-1 により切断され IL-1βとして細胞外に放出されるが、miRNA-466gとmiRNA-466m-5pの細胞 への遺伝子導入は、IL-1βの分泌を亢進させることが分かった。これは sepsis による急性肺障害に exosome の放出を阻害する GW4869 を前投与すると血清
のIL-1β量が減少したという過去の報告39)に矛盾しない。細胞内におけるpro-
IL-1βの観察では、miRNA-466gとmiRNA-466m-5pの遺伝子導入により、LPS
20
刺 激 に よ る pro-IL-1β の 産 生 が 上 昇 し て い る こ と か ら 、miRNA-466g と miRNA-466m-5pは、LPS によるpro-IL-1βの産生を亢進していると考えられ た。しかし今回の検討では、exosome内のmiRNA-466gとmiRNA-466m-5pが インフラマソームの priming のどの因子をターゲットにしているのか特定でき ておらず、さらにexosome内で発現亢進したすべてのmiRNA の働きについて もその働きが明らかでないため、今後検討していく必要がある。
本実験は、LPS 経気道刺激により BALF 中 exosome 内で発現亢進した miRNA-466gとmiRNA-466m-5pが、マクロファージのNLRP3インフラマソ
ームのpriming に作用することにより pro-IL-1βの産生が増加し、その結果と
してIL-1βが過剰に産生されることでARDSの発症に関与している可能性を明
らかにした。以上から、BALF中exosomeやその中に含まれるmiRNA-466gや
miRNA-466m-5pを制御することがARDSの新たな治療法となりえると考えら
れた。
Ⅵ.まとめ
LPS 刺激により気道内に分泌される exosome は miRNA-466g と miRNA-
466m-5pの発現を増加させ、インフラマソームを介するIL-1βの産生を増加さ
せることにより ARDS の発症に関与している可能性が示唆された。また、
21
exosomeの気道への分泌や、それに内包されるmiRNA-466gやmiRNA-466m- 5pは、急性肺障害の新たな治療標的になると考えられた。
Ⅶ.謝辞
本研究に関して、研究を御指導賜りました日本大学医学部呼吸器内科学分 野 橋本修教授をはじめ、権寧博准教授、丸岡秀一郎准教授、水村賢司助教に深 謝いたします。また、本研究に対して多大なる御尽力を頂きました先生方、研究 補助員の皆様方にこの場を借りまして心から厚く御礼申し上げます。
22
Ⅷ.図表
miRNA Fold Change p-value
Mmu-miR-16-1-star 6.17 0.00119
Mmu-miR-466g 6.04 2.55E-05
Mmu-miR-669m-5p 5.27 0.000802
Mmu-miR-466f-5p 5.03 0.00193
Mmu-miR-195-star 4.70 0.000202
Mmu-miR-466f-3p 4.60 8.00E-05
Mmu-miR-669f-5p 4.27 3.63E-05
Mmu-miR-1187 4.17 0.00316
Mmu-miR-466m-5p 4.12 3.62E-06
Mmu-miR-455 3.74 0.00147
表
123
LPS
TLR4 P2X7
受容体
NLRP3
Caspase-1
pro-IL-1β IL-1β
ATP(danger signal)
傷害
組織 病原菌
マクロファージ インフラマソーム
priming triggering
図
1NF-kB
24
図
225
PBS
LPS
図
326
図
4A27
図
4B図
4C28
図
5PBS
LPS
29
図
6P B S L P S
30
図
7B図
7A31
図
8A- - + - - + - - + - + + - + + - + + LPS
ATP
miR-control miR-466g miR-466m-5p
32
図
8B- - + - - + - - + - + + - + + - + + LPS
ATP
miR-control miR-466g miR-466m-5p
33
Ⅸ.図表説明
表1. LPSにより誘導されるmiRNAリスト (上位10遺伝子)
PBS 群 (n=40)または LPS 群 (n=4)より採取した BALF から exosome を抽 出し、さらにexosome内のtotal RNAを抽出し、miRNAアレイを用いて網羅 発現解析を行った。発現の上昇が大きい上位 10 の miRNA のリストを示す。
Fold changeはLPS非投与群との比を表す。
図1. NLRP3インフラマソーム
NLRP3 インフラマソームとはマクロファージ内で炎症を制御するタンパ
ク質複合体であり、PAMPやDAMPをマクロファージのTLR4が認識してpro- IL-1βを産生する priming と、組織障害により danger signal として細胞外に 放出された ATP を P2X7 受容体が認識して NLRP3 を介して活性化された caspase-1 が IL-1β前駆体を切断し、IL-1βを産生する triggering により炎症 を引き起こす。
図2. Exosomeの模式図
Exosomeは脂質二重膜で構成されており、内部には核酸やタンパクが含まれ
ている。
34
図3. Exosomeの走査電子顕微鏡所見
PBSまたは LPSを経気道投与したマウスの BALFより抽出された exosome を走査電子顕微鏡 (×10,000)で観察した。スケールバーはPBS群では0.7cmが 100nm、LPS群では1.5cmが100nmを示している。
図4. LPS投与によるBALF中のexosome数とRNA量に及ぼす影響
A). LPS投与がBALF中のexosome数に及ぼす影響。
ナノ粒子カウンターにより、BALFに含まれるexosome数を測定した。実験 結果は平均値±SDsで示す。n = 5、N.Sはnot significant。
B). LPS投与がBALF中exosome内のRNA量に及ぼす影響 。
BALFのexosomeから、前述のRNA抽出キットを使用してtotal RNAを抽 出した。Total RNA 量はバイオアナライザーにより測定した。実験結果は平均 値±SDsで示す。n = 5 **P < 0.01 対LPS非投与群。
C). LPS投与がBALF中exosome1個当たりのRNA量に及ぼす影響。
図4Bの RNA量を図 4Aのexosome 数で割り、exosome1 個当たりのRNA 量として算出し、比較した。n = 5 *P < 0.05 対LPS非投与群。
35
図5. バイオアナライザーによるRNAの計測
PBS 投与群と LPS 投与群の exosome 内 RNA のサイズをバイオアナライザ
ーで計測した。nt; nucleotide、FU; fluorescence units。
図6. LPS投与によるBALF中のexosome内miRNAの網羅的発現解析
階層的クラスタリング解析による PBS 群と LPS 群の miRNA 遺伝子発現の
違いをheat mapで示す。平均より発現が亢進している遺伝子を赤、発現が減弱
している遺伝子を緑で示した。
図7. miRNA-466gとmiRNA-466m-5pの発現量のPCRによる検証
A, B). LPS投与により発現増加したmiRNA466gと466m-5pについて、リア ルタイムPCRを用いて発現レベルの検証を行った。サンプルの発現量は核内低 分子 RNA の一つ U6 をベースに正規化した。実験結果は平均±SDs で示す。
n=12 *P < 0.05対LPS非投与群, **P < 0.01対LPS非投与群。
図8. miRNA-466gとmiRNA466m-5pのIL-1β産出に及ぼす影響
miRNA-466g と miRNA-466m-5p を BMDM にトランスフェクションし、
LPS 刺激後 ATP を上清中に添加し、1 時間後に細胞上清と細胞溶解液を得た。
36
A). ELISA法で測定した培養上清中のIL-1β濃度。実験結果は平均±SDsで
示す。n=3, ***P < 0.001対コントロールmiRNA群。N.D.:not detected。
B). western blotで検出した細胞内のpro-IL-1βタンパク (31kDa)。
Ⅹ.引用文献
1. ARDS診療ガイドライン2016:日本呼吸器学会, 日本集中治療医学会, 日 本呼吸療法医学会. (2016).
2. Ashbaugh DG, Bigelow DB, Petty TL, et al Acute respiratory distress in adults. Lancet 2, 319-323 (1967).
3. Petty TL, Ashbaugh DG, The adult respiratory distress syndrome.
Clinical features, factors influencing prognosis and principles of management. Chest 60, 233-239 (1971).
4. Murray JF, Matthay MA, Luce JM, et al An expanded definition of adult respiratory distress syndrome. American review respiratory disease 138, 720-723 (1988).
5. Bernard GR, Artigas A, Brigham KL, et al The American-European Consensus Conference on ARDS. Definitions, mechanisms, relevant outcomes, and clinical trial coordination. American journal of
37
respiratory and critical care medicine 149, 818-824 (1994).
6. Ranieri VM, Rubenfeld GD, Thompson BT, et al Acute Respiratory Distress Syndrome: the Berlin Definition, Journal of the American medicine association 307, 2526-2533 (2012).
7. Bellani G, Laffey JG, Pham T, et al Epidemiology, Patterns of Care, and Mortality for Patients with acute respiratory distress syndrome in intensive care units in 50 countries, Journal of the American medicine association 315, 788-800 (2016).
8. Agarwal R, Srinivas R, Nath A, et al Is the mortality higher in the pulmonary vs the extrapulmonary ARDS A meta analysis. Chest 133, 1463-1473 (2008).
9. Bone RC, Balk RA, Cerra FB, et al Definitions for sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. The ACCP/SCCM Consensus Conference Committee. American College of Chest Physicians/Society of Critical Care Medicine. Chest 101, 1644- 1655 (1992).
10. Ware LB, Matthay MA, The acute respiratory distress syndrome, The New England journal of medicine 342, 1334-1349 (2000).
38
11. Grommes J, Soehnlein O, Contribution of neutrophils to acute lung injury, Molecular medicine 17, 293-307 (2011).
12. Grailer JJ, Canning BA, Kalbitz M, et al Critical role for the NLRP3 inflammasome during acute lung injury, The journal of immunology 192, 1-10 (2014).
13. Bauernfeind FG, Horvath G, Stutz A, et al NF-kB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulatoting NLRP3 expression, The journal of immunology 183, 787-791 (2009).
14. L Franchi, T Eigenbrod, R Muñoz-Planillo, et al The Inflammasome: a caspase-1 activation platform regulating immune responses and disease pathogenesis, Nature immunology 10(3):204, 1-15 (2009) 15. TH Lee, HJ Song, CS Park, Role of inflammasome activation in
development and exacerbation of asthma, Asia Pacific allergy 4(4), 187-196 (2014)
16. T Dolinay, YS Kim, J Howrylak, et al Inflammasome-regulated Cytokines Are Critical Mediators of Acute Lung Injury, American journal of respiratory and critical care medicine 185, 1225-1234 (2012)
39
17. Yamada W, Tasaka S, Koh H, et al Role of toll-like receptor 4 in acute neutrophilic lung inflammation induced by intracheal bacterial products in mice, Journal of inflammation research 1, 1-10 (2008) 18. de Rivero Vaccari JP, Brand F 3rd, Adamczak S, et al Exosome-
mediated inflammasome signaling after central nervous system injury, Journal of neurochemistry 136, 39-48 (2016)
19. Valadi H, Ekström K, Bossios A, et al Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nature cell biology 9, 654-659 (2007).
20. Théry C, Zitvogel L, Amigorena S, Exosomes: composition, biogenesis and function. Nature reviews immunology 2, 569-579 (2002).
21. Pan, BT, Johnstone, RM, Fate of the transferrin receptor during maturation of sheep reticulocytes in vivo: selective externalization of the receptor. Cell 33, 967-978 (1983).
22. Harding C, Heuser J, Stahl P, Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes.
Journal cell biology 97, 329-339 (1983).
23. Pan BT, Teng K, Wu C, et al Electron microscopic evidence for
40
externalization of the transferrin receptor in vesicular form in sheep reticulocytes. Journal cell biology 101, 942-948 (1985).
24. Yoshioka Y1, Kosaka N2, Konishi Y, et al Ultra-sensitive liquid biopsy of circulating extracellular vesicles using ExoScreen, Nature
communications 4591, 1-8 (2014)
25. Mazzeo C, Cañas JA, Zafra MP, et al Exosome secretion by eosinophilis: A possible role in asthma pathogenesis, Journal of clinical immunology 135, 1603-16138 (2015)
26. He L, Hannon GJ, MicroRNAs: small RNAs with a big role in gene regulation, Nature 5, 522-531(2004).
27. Wahid F, Shehzad A, Khan T, et al, MicroRNA: synthesis, mechanism, function, and recent clinical trials, Bochemica et biophysica acta 1803, 1231-1243 (2010).
28. Lee RC, Feinbaum RL, Ambros V, The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14, Cell 75, 843-854 (1993).
29. Wightman B, Ha I, Ruvkun G, Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation