マウス肥満モデルにおける
Extracellular Signal-Regulated Kinase 2
(ERK2)を介した脂肪細胞分化と血管内皮機能
への影響の検討
大﨑 歩
(循環器病学専攻)
防衛医科大学校
令和2年度
目 次
第1章 緒言 1頁 第2章 脂肪細胞ERK2が脂肪細胞分化とインスリン抵抗性に及ぼす影響の検討
第1節 背景及び目的 5頁
第2節 対象と方法 7頁 第3節 結果 16頁 第4節 考察 25頁 第5節 小括 30頁
第3章 脂肪細胞ERK2が血管内皮機能や血圧に及ぼす影響の検討
第1節 背景及び目的 31頁
第2節 対象と方法 32頁 第3節 結果 35頁 第4節 考察 38頁 第5節 小括 40頁
第4章 全体の考察 41頁
第5章 結論 47頁 謝辞 48頁 略語一覧 49頁 引用文献 51頁 図表 59頁
1
第1章 緒言
厚生労働省によると日本人の死因として第2位に心疾患、第4位に脳血管疾 患となっており(1)、心疾患、脳血管疾患の原因として動脈硬化が危険因子と
なる。動脈硬化の要因としては、高血圧、2型糖尿病、脂質異常症、肥満など
が挙げられ、それらが複合的に関与すると個々の要因よりも心血管疾患の発症
リスクが高くなる。メタボリックシンドローム(Mets)は、肥満によるインス
リン抵抗性を基盤とした耐糖能異常、血圧高値、脂質代謝異常の複合疾患であ
り、虚血性心疾患をはじめとした心血管疾患の重要な危険因子として、1999年
に世界保健機構(WHO)によってその概念と診断基準が提唱された(2, 3)。厚
生労働省(平成28年国民健康・栄養調査の概要)の発表によると、我が国の 肥満者の割合は男性で32.4%、女性で21.6%に至る。Metsでは個々の要因が複 合することにより心血管疾患の発症リスクが増大するため(4-6)、その対策は
心血管疾患の長期予後改善に資する。また、西洋食にさらされる自衛隊衛生に
おいてもMets対策は重要な課題である。しかし、現状のMets対策としては食 事・運動療法と各要因に対する治療のみで、Mets合併症の成因や特異的治療法
は確立されていない。
生活習慣の欧米化、高脂肪食、高糖質食と運動不足は体内のエネルギーバラ
ンスを貯蔵に傾け、糖・脂質調整に関わる遺伝子や生理活性物質の発現を変化
2
させる(7, 8)。特に脂肪細胞はエネルギー貯蔵と供給の中心的役割を担ってお
り、Mets病態において非常に重要な標的臓器である。脂肪組織は大きく皮下脂
肪と内臓脂肪に分けられる。脂肪細胞は内分泌器官として様々な生理活性物質
(アディポサイトカイン)を分泌するが、内臓脂肪の蓄積はアディポサイトカ
イン分泌を変化させ、高血圧、糖尿病、心血管合併症進展に関与する(9)。一
方で近年、皮下脂肪は内臓脂肪とは異なる機能を持つことも注目されてきてお
り(10)、内臓脂肪だけでなく皮下脂肪の蓄積においても、インスリン抵抗性進
展に関与する報告がある(11)。それに加えて脂質が過剰蓄積した場合に、本来
蓄積されるはずの皮下脂肪や内臓脂肪だけではなく、他臓器に脂肪が蓄積す
る、「異所性脂肪沈着」も着目されている(12)。インスリン標的臓器への異所
性脂肪沈着はインスリン感受性の低下やインスリン分泌の増加につながる。例
えばMets病態下では、代謝の中心を担う肝臓で脂肪変性が主体となる非アル コール性脂肪肝の進展を合併し、この肝脂肪変性がインスリン抵抗性と心血管
疾患のリスク増大に関与する(13)。Metsの心血管疾患の進展機序を考えるにあ
たり、内臓脂肪の蓄積のみではなく皮下脂肪機能や異所性脂肪沈着が2型糖尿 病、脂質異常症、高血圧を基盤とした心血管合併症に関連する可能性がある。
Metsに伴うインスリン抵抗性では、臓器特異的なインスリンシグナルの障害 が糖・脂質代謝異常のみならず、Mets合併症の進展に関与する(14)。脂肪細胞
3
における主要インスリンシグナル伝達経路として二つの経路があり、一つは
phosphatidylinositol-3 kinase(PI3K)/AKT経路で糖・脂肪代謝を調節し、
もう一方はRaf/mitogen-activated protein kinase kinase(MEK)
/extracellular signal-regulated kinase(ERK)経路であり脂肪細胞の分 化・増殖を制御する(図1)(15)。過去の報告では培養脂肪細胞において分裂 促進因子活性化蛋白キナーゼ(mitogen-activated protein kinases;MAPKs)
/ERK経路が脂肪細胞の分化や増殖を制御することが報告されている(16-20)。
MAPKsファミリーに属するERK経路の活性化は、分化早期において前駆脂肪細
胞の分化を促進することが報告されたが(17, 21)、MAPKsの活性化は成熟脂肪
細胞分化を抑制する報告もあり(19, 22, 23)、ERK経路の脂肪細胞分化への関
与の詳細は解明されていない。
ERKには主にERK1とERK2の二つのサブタイプがあり、アミノ酸レベルで 80%以上の相同性があり、非常に似通った構造と標的分子を有する(24)。しか し全身でのERK1欠損マウスは生存できるが(25)、全身でのERK2欠損マウスで は胎生致死となることが報告され(26, 27)、その役割や局在・標的に相違があ
ることが想定されている。ERK1欠損マウスから培養した前駆細胞からは、脂肪
細胞分化が抑制されたとの報告がある(28)。脂肪細胞のERK経路は2型糖尿病 の発症とも関連があることから、ERK経路がインスリン抵抗性や2型糖尿病の
4
治療標的になる可能性も示唆されている(29)。当教室では過去にKujiraokaら が、肝細胞特異的ERK2欠損マウスを作出したところ、肥満や脂肪肝炎の悪化 と血管内皮機能障害が認められた(30)。しかしながらMetsにおけるin vivo での脂肪細胞のERK2の役割は未だ解明されていない。
これらの背景を基に、本研究では脂肪細胞特異的ERK2ノックアウト
(AE2KO)マウスを用いて、高脂肪高ショ糖食を与えることでマウス肥満・イ
ンスリン抵抗性モデルを作成し、in vivoにおける脂肪細胞ERK2の細胞分化へ の作用、それが脂質代謝、異所性脂肪沈着及びインスリン抵抗性に与える影響
を検討した。次にこれらの変化が心血管疾患の基盤である高血圧や血管内皮機
能に及ぼす影響を検討した。本研究の最終目的は脂肪細胞ERK2による脂肪組 織と脂質代謝の変化、Mets合併症臓器障害に対する役割について、AE2KOマウ
スを用いて明らかにすることである。
5
第2章 脂肪細胞ERK2が脂肪細胞分化とインスリン抵抗性に及ぼす影響 の検討
第1節 背景及び目的
脂肪細胞は余剰なエネルギーを中性脂肪として蓄積し、全身に再供給する役
割を担う。脂肪細胞の集団である脂肪組織は、皮下や内臓周囲、腎臓や骨格
筋、心筋、血管の周囲などにも存在する。脂肪組織はエネルギーバランスと基
礎代謝の重要な調整組織であり、過剰な脂肪蓄積は2型糖尿病や動脈硬化、心
血管疾患の悪化と関連する。脂肪細胞の分化・増殖においてインスリンシグナ
ル伝達経路は非常に重要な役割を担うが、その役割は完全には解明されていな
い。脂肪細胞ではperoxisome proliferator activated receptor γ(PPARγ) が直接分化に関わる重要な因子として同定されており(31, 32)、PPARγは脂肪
細胞特有の遺伝子発現を誘導し、脂肪蓄積能を獲得させる(33)。糖尿病治療薬
のひとつであるチアゾリン系薬剤は PPARγ の活性を上昇させ、前駆脂肪細胞
から脂肪細胞への分化を促進し、新たに分化した小型の脂肪細胞の増加と共
に、肥大化した脂肪細胞のアポトーシスによりインスリン抵抗性を改善させる
(34)。PPARγの制御因子の一つにERKが同定されており、in vitroの研究では ERKがPPARγの制御に寄与することが多数報告されている(17, 18, 35)。また
6
脂肪組織における脂質調節は、脂肪分解と脂質放出における2つの重要な酵素
であるlipoprotein lipase(LPL)とhormone-sensitive lipase(HSL)によ って制御されている(36, 37)。LPLは脂肪細胞を含む様々な臓器で発現してお
り、中性脂肪及び超低密度リポ蛋白(very low density lipoprotein;VLDL)
の加水分解を触媒することで、リポタンパク質代謝において重要な役割を担っ
ている(38)。LPL 発現はprotein kinase(PKC)/ ERK/ c-Fosシグナル伝達で 制御されており、脂肪細胞の分化・機能獲得へのERKの関与が想定される。ま たMetsで増加するreactive oxygen species(ROS)はLPL発現を低下させる
(39)。従って、Metsの脂肪細胞においてERKはインスリンによる脂肪分化に深
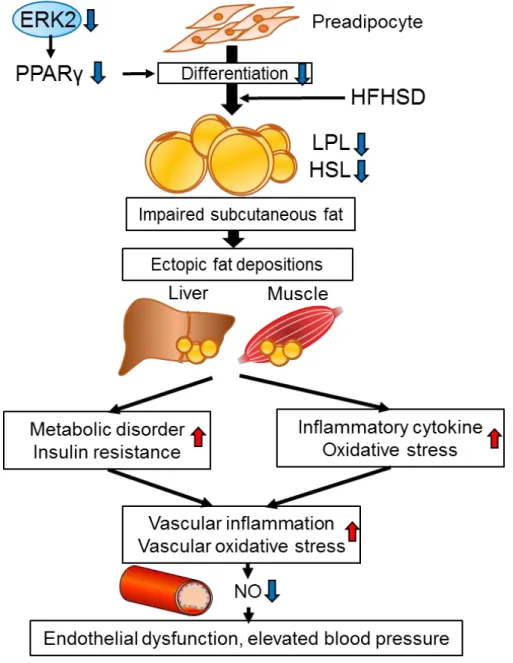
く関わり、Mets合併症に関与しうる(図2)。
そこで本章ではAE2KOマウスを用いて脂肪細胞ERK2の分化・脂質代謝への 作用を評価し、異所性脂肪沈着及びインスリン抵抗性に与える影響を検討し
た。
7
第2節 対象と方法
(1)実験動物
ERK2 floxマウスは防衛医科大学校生化学教室 佐藤泰司教授より供与され
た(40)。Fatty acid binding protein 4(FABP4)はin vitro において、脂肪 幹細胞の分化誘導直後では発現はみられないが、分化誘導2日目以降に発現が
亢進することがわかっており、脂肪細胞の分化マーカーとして用いられる
(41)。そのため脂肪細胞特異的プロモーターであるFabp4の制御下でDNAの組
換え酵素であるCreを発現しているFabp4-CreマウスをThe Jackson
laboratory(Bar Harber ME、USA)より購入した。これらのマウスを交配し、
脂肪細胞特異的ERK2ノックアウトマウスを作成し、遺伝子背景はC57BL/6Jで 揃えた。実験には脂肪細胞特異的ERK2ノックアウトマウス(Fabp4-Cre [+/
-]; ERK2 [lox/lox] マウス [AE2KOマウス])とControlマウス(Fabp4-Cre [-/-]; ERK2 [lox/lox] マウス [Controlマウス])を用いた。また、ERK2 発現の比較検討するためにC57BL/6Jの野生型(wild-type; WT)マウスを日本 クレア株式会社(東京)より購入した。
飼育は防衛医科大学校動物実験施設にて行い、12時間毎の明暗の照明のもと
に行われた。明は7時から19時、暗は19時から7時とした。食餌は通常食
(日本クレア株式会社、東京)及び高脂肪高ショ糖食(炭水化物 28.3%、脂
8
質 54.5%、タンパク質 17.2%)(表1)(オリエンタル酵母株式会社、東京)
を使用し、食餌・飲水は共に自由摂取とした。Controlマウス及びAE2KOマウ スそれぞれに対して、通常食(normal diet;ND)及び高脂肪高ショ糖食
(high fat/high sucrose diet;HFHSD)を6週齢から30週齢までの24週間 負荷し、4群(Control-ND、AE2KO-ND、Control-HFHSD、AE2KO-HFHSD)を作成
し、2週間毎に体重を測定した(図3)。
ERK2 flox遺伝子とCre遺伝子の確認のためのgenotypingは、polymerase chain reaction法(PCR法)を用いて確認した。flox遺伝子の検出のためには 5’-GATCTGATGCTTGCCAAAGCC-3’及び 5’-TGTAAAGTAGCAGCAGATGC-3’をプ
ライマーとして使用し、Cre遺伝子の検出のために、5’-
GCGGTCTGGCAGTAAAAACTATC-3’及び 5’-GTGAAACAGCATTGCTGTCACTT-3’をプ ライマーとして使用した(30)。また、本研究は防衛医科大学校の動物実験倫理
委員会の承認を得て(承認番号:19038)、動物実験規則に基づき実施した。ま
た組換えDNA実験委員会の承認を得て(承認番号:2018-17)、組換えDNA実験 安全管理規則に基づき実施した。
9
(2)組織のタンパク質抽出及びウエスタンブロット法
イソフルランによる麻酔下で開胸腹し、血液を回収した後クレブス・リンゲ
ル重炭酸バッファー(NaCl 118.3 mmol/L、KCl 4.7 mmol/L、CaCl 2.5
mmol/L、MgSO4 1.2 mmol/L、KH2PO4 1.2 mmol/L、NaHCO3 25 mmol/L、D-glucose 5.5 mmol/L)で灌流した。摘出した組織(白色脂肪、褐色脂肪、大動脈、心 臓、肝臓、膵臓、腎臓、筋肉、脳)は1 mmol/Lの
phenylmethylsulfonylfluoride (PMSF)及びprotease inhibitor cocktailを 含んだタンパク質抽出液(Tris-HCl 20 mmol/L、NaCl 150 mmol/L、Na2EDTA 1 mmol/L、EGTA 1 mmol/L、1% NP-40、sodium pyrophosphate 2.5 mmol/L、β- glycerophosphate 1 mmol/L、Na2VO4 1 mmol/L、pH 7.4)を加え、ホモジナイ ザーを用いて溶解した。溶解液は 4℃、15,000 x gで20分遠心し、上清を回 収した。タンパク質濃度の測定はBradford法を用いて行った。白色脂肪は精 巣周囲脂肪、筋肉は腓腹筋を用いた。サンプルの加熱処理を行った後、ポリア
クリルアミドゲル電気泳動を行い、転写の際はpolyvinylidene difluoride
(PVDF)メンブレンを使用した。ブロッキング液(Blocking One、ナカライテ
スク、京都)に1時間浸して 4℃で一晩、1次抗体反応を行った。翌日、
Tween20含有リン酸緩衝液(PBS-T)で3回洗浄後、2次抗体反応を行った。抗
体反応終了後にPBS-Tで3回洗浄し、SuperSignal West Dura Extended
10
Duration Substrate(Thermo Fisher Scientific、 Waltham、 MA、 USA)を 用いて化学発光させ、LAS300-IR(富士フィルム、東京)で検出した。
1次抗体の抗ERK1/2抗体(9102S)はCell Signaling Technology
(Danvers、 MA、 USA)から入手したものを使用した。検出したタンパク質は
Multi Gauge – Image(富士フィルム、東京)で解析した。
(3)脂肪細胞の組織学的評価
イソフルラン麻酔下で開胸腹し、血液を回収した後クレブス・リンゲル重炭
酸バッファーで灌流した。摘出した皮下脂肪及び内臓脂肪を4%パラホルムアル デヒドで24時間固定し、パラフィン切片を作成した。皮下脂肪は腋窩から背 側にある脂肪、内臓脂肪は精巣周囲脂肪を採取した。皮下脂肪及び内臓脂肪の
ERK2の発現を確認するために、抗ERK2抗体(AB7948)(Abcam plc、
Cambridge、 UK)を用いて免疫蛍光染色を行った。さらに脂肪細胞の形態を確 認するために皮下脂肪及び内臓脂肪のパラフィン切片にヘマトキシリン・エオ
ジン染色を行った。画像はKEYENCE社のBZ-X710(大阪)を用いて評価した。
11
(4)RNA抽出及びリアルタイムPCR法
イソフルランによる麻酔下で開胸腹し、血液を回収した後クレブス・リンゲル
重炭酸バッファーで灌流した。摘出した皮下脂肪及び内臓脂肪にQIAzol Lysis Reagent(QIAGEN、PL、NLD)を加え、ホモジナイザーを用いて溶解した。クロロ ホルムを加えて 4℃、12,000 x gで15分遠心し、上清を回収した。さらにRNeasy Lipid Tissue Mini Kit(QIAGEN、PL、NLD)を用いてRNAを抽出した。ReverTra Ace qPCR RT Master Mix(東洋紡、大阪)を用いて逆転写を行い、相補的DNAを 生成した。これを鋳型としてリアルタイム PCR を行った。皮下脂肪及び内臓脂
肪のRNAを用いてERK2、脂肪の分化マーカーであるPPARγ、脂肪組織で分泌・
合成され、脂肪細胞への中性脂肪貯蔵を促進する LPL 及び遊離脂肪酸を放出し 脂肪細胞の中性脂肪分解を促進するHSLを評価した。mRNAの発現はPower SYBR Green PCR Master Mix(Applied Biosystems、 MA、 USA)を用い、ViiaTM7 リア ルタイムPCRシステム(Applied Biosystems、 MA、 USA)でリアルタイムPCR を行い評価した。プライマーの塩基配列は表 2 のものを使用した。内部標準と してのハウスキーピング遺伝子はribosomal protein S18(S18)を使用した。
12
(5)脂質代謝の評価
食餌摂取による影響を排除するため、14時間の絶食後、イソフルラン麻酔下
で血液を回収した後、血液を遠心して血清を回収した。回収した血清の中性脂
肪、総コレステロール、HDLコレステロール、遊離脂肪酸を生化学検査用キッ
ト(富士フィルム和光純薬株式会社、大阪)を用いて測定した。
(6)肝臓及び筋肉への異所性脂肪沈着の評価
異所性脂肪沈着の評価のために肝臓と筋肉の脂肪沈着を評価した。筋肉の評
価には腓腹筋を用いた。摘出した肝臓及び筋肉をoptimal cutting
temperature medium(O.C.T Compound、サクラファインテックジャパン株式会 社、東京)に包埋し、液体窒素で急速凍結し、10 µmの厚さで切片を作成し
た。スライドをオイルレッドO染色液に 37℃で15分染色した後、マイヤーの ヘマトキシリン溶液に5分間浸水した。オイルレッドOはSigma-Aldrich(St Louis、MO、USA)のものを使用した。また第2章(3)と同様の方法で作成し た肝臓のパラフィン切片で、線維化を評価するためにマッソン・トリクローム
染色を行い、組織学的な肝臓の変化を観察した。画像はKEYENCE社のBZ-X710
(大阪)を用いて評価した。
脂肪肝の評価のために回収された血清を酵素測定用キット(富士フィルム和
13
光純薬株式会社、大阪)を用いて、アラニンアミノトランスフェラーゼ
(alanine aminotransferase;ALT)を測定した。
また摘出した肝臓及び筋肉1 g当たり、タンパク質抽出バッファーを5 ml 加えてホモジナイザーを用いて溶解した。溶解液を 4℃、1、5,000 x gで20 分間遠心し、上清を回収した後、中性脂肪及び総コレステロールを生化学検査
用キット(富士フィルム和光純薬株式会社、大阪)を用いて測定した。
(7)インスリン抵抗性及び感受性の評価
空腹時グルコース濃度は14時間の絶食の後、血液をtail cutにて採取し、
血中のグルコース濃度をアキュチェックSTメーター(ロシュDCジャパン株式 会社、東京)を用いて測定した。またガラス毛細管を用いて採取した血液を、
遠心して血清を回収し、回収した血清を用いて血清中のインスリン濃度を
ELISAキット(富士フィルムワコーシバヤギ株式会社、群馬)を用いて測定し
た。インスリン抵抗性の評価に必要なhomeostasis model assessment of insulin resistance(HOMA-IR)は以下の式で計算した。
HOMA-IR=空腹時グルコース濃度(mg/dl)×空腹時インスリン濃度(ng/ml)
より詳細な糖代謝を評価するために、インスリン負荷試験(insulin
tolerance test;ITT)を行った。ITTは4時間の絶食後、速効型インスリン
14
0.5 U/kg体重(Humulin R、Eli Lilly and Company、Indianapolis、IN、
USA)を腹腔内投与し、投与直前(0分)、投与15分、30分、45分、60分、
120分でそれぞれ血液を採取し、グルコース濃度を測定した。
(8)血清アディポサイトカインの測定
食餌摂取による影響を排除するため、14時間の絶食後、イソフルラン麻酔下
で血液を回収した後、血液を遠心して血清を回収した。回収した血清のアディ
ポサイトカインであるアディポネクチン、レプチン濃度をそれぞれELISAキッ ト(富士フィルムワコーシバヤギ株式会社、群馬)を用いて測定し、tumor
necrosis factor-α(TNF-α)をELISAキット(R&D system、Minneapolis、
NM、USA)で測定した。
(9)統計学的解析
すべてのデータを平均±標準誤差(mean±SEM)で表記した。体重、ITTの評
価には繰り返しのあるtwo-way analysis of variance(ANOVA)後、
Bonferroni法でpost hoc解析を行った。2群間の比較にはMann-Whitney U検 定を使用した。すべての統計解析はGraphPad Prism Software Ver.7
(GraphPad Software、La Jolla、CA、USA)を使用し、すべての検定において
15
p<0.05を統計学的有意と判定した。
16
第3節 結果
(1)各臓器におけるERK2の発現
マウスのERK2タンパクの発現を確認するために、WTマウス、FABP4-Creマ ウス、Controlマウス、AE2KOマウスの各臓器を用いて、ウエスタンブロット
法を行った。白色脂肪に加えて、褐色脂肪、大動脈、心臓、肝臓、膵臓、腎
臓、筋肉、脳におけるERK2のタンパク発現を確認したところ、白色脂肪及び 褐色脂肪ではERK2の低下がみられたが、他の臓器ではControlマウスとAE2KO マウスに差がないことを確認した(図4A)。さらに抗ERK2抗体を用いて免疫蛍 光染色を行い、皮下脂肪及び内臓脂肪におけるERK2の発現を確認したとこ ろ、AE2KOマウスではControlマウスと比較してERK2の発現が皮下脂肪及び内 臓脂肪共に低下していた(図4B)。またリアルタイムPCR法において、今回作 成した4群のマウス間(Control-ND、AE2KO-ND、Control-HFHSD、AE2KO- HFHSD)でERK2のmRNAの発現を比較検討したところ、AE2KOマウスのERK2の 発現は皮下脂肪(Control-ND=1.00±0.35[N=7]対AE2KO-ND=0.18±0.06
[N=7]、P=0.011、Control-HFHSD=0.88±0.23[N=7]対AE2KO-HFHSD=
0.11±0.07[N=5]、P=0.005)及び内臓脂肪(Control-ND=1.00±0.06[N=
7]対AE2KO-ND=0.56±0.06[N=8]、P<0.001、Control-HFHSD=1.28±0.33
[N=6]対AE2KO-HFHSD=0.46±0.064[N=7]、P=0.005)共にControlマウ
17
スと比較して低下していた(図4C、D)。
(2)AE2KOマウスの特徴
ControlマウスとAE2KOマウスはND群では体重に変化はみられなかったが、
HFHSD群ではControlマウスと比較してAE2KOマウスで有意に体重増加を認め た(28週齢時の体重:Control-HFHSD=42.95±1.00 g[N=13]対AE2KO- HFHSD=48.75±0.98 g[N=14]、P<0.001)(図5A、B)。30週齢において3日 間食事摂取量を計測したが、各食事間でControlマウスとAE2KOマウスの食事 摂取量に差はみられなかった。また皮下脂肪及び内臓脂肪の重量はND群では ControlマウスとAE2KOマウスに差はみられなかった(図5C、D)。HFHSD群で は内臓脂肪の重量はControlマウスとAE2KOマウスに差はみられなかったが (図5D)、皮下脂肪ではAE2KOマウスで有意な増加がみられた(Control-HFHSD
=0.98±0.095 g[N=10]対AE2KO-HFHSD=1.76±0.12 g[N=8]、P<0.001)
(図5C)。さらに肝臓及び筋肉の重量を測定したところ、HFHSD群において
AE2KOマウスの肝臓及び筋肉の重量増加がみられた(肝臓;Control-HFHSD=
1.41±0.11 g[N=9]対AE2KO-HFHSD=2.42±0.26 g[N=8]、P=0.004、筋 肉;Control-HFHSD=0.44±0.03 g[N=8]対AE2KO-HFHSD=0.58±0.05 g[N
=7]、P=0.029)(図5E、F)。これらの結果からAE2KO-HFHSD群の体重増加の
18
原因として、皮下脂肪重量の増加に加え、肝臓や筋肉における異所性脂肪沈着
が原因として考えられた。
皮下脂肪及び内臓脂肪のヘマトキシリン・エオジン染色を行い(図6A、E)、
単位面積あたりの脂肪細胞数と脂肪細胞のサイズ(直径)を評価した。皮下脂
肪では、単位面積あたり脂肪細胞数がAE2KO-HFHSD群でControl-HFHSD群と比 較して増加しており(Control-HFHSD=485.9±34.0 cells/nm2[N=7]対
AE2KO-HFHSD=721.0±56.6 cells/nm2[N=8]、P=0.001)(図6B)、また脂肪 細胞の直径を測定したヒストグラムではControl群のHFHSD負荷でみられた脂 肪細胞の大型化が、AE2KO-HFHSD群ではみられず、脂肪細胞の大型化が障害さ
れていた(図6D)。内臓脂肪では単位面積あたりの脂肪細胞数は、Control群と AE2KO群に差はみられなかった(Control-HFHSD=376.5±38.8 cells/nm2[N=
7]対AE2KO-HFHSD=323.3±22.9 cells/nm2[N=8]、P=0.39)(図6F)。内臓 脂肪細胞のヒストグラムでは、Control群のHFHSD負荷においても脂肪細胞の 大型化はみられず、AE2KO群においてもControl群と同様の変化であった(図
6G、H)。AE2KO-HFHSD群の皮下脂肪の重量増加と単位面積あたりの脂肪細胞数
が増加していたことから、AE2KO-HFHSD群の皮下脂肪は大型化できない皮下脂
肪細胞の数が増加したものと考えられた。
19
(3)脂肪細胞の分化・機能のリアルタイムPCRによる評価
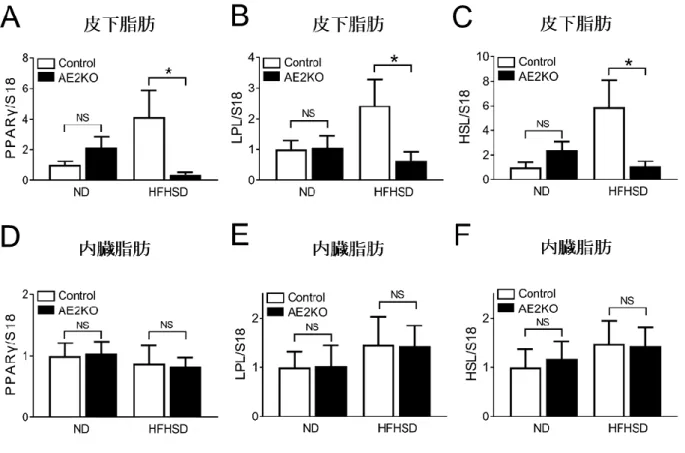
リアルタイムPCR法で内臓脂肪及び皮下脂肪で脂肪の分化に重要な役割を果 たす PPARγ のmRNAの発現を評価したところ、AE2KO-HFHSD群の皮下脂肪で低 下していた(Control-HFHSD=4.14±1.73[N=7]対AE2KO-HFHSD=0.38±0.15
g[N=7]、P=0.001)(図7A)。この変化は内臓脂肪ではみられなかった(図
7D)。次に脂肪組織において中性脂肪貯蔵の促進するLPL、分解を促進するHSL の発現をリアルタイムPCRで評価した。皮下脂肪ではHFHSD群でControlマウ スと比較してAE2KOマウスがLPL及びHSLのmRNAの発現が共に低下していた
(LPL; Control-HFHSD=2.44±0.85[N=5]対AE2KO-HFHSD=0.64±0.29[N=
5]、P=0.05、HSL; Control-HFHSD=5.88±2.22[N=7]対AE2KO-HFHSD=1.07
±0.42[N=8]、P=0.009)(図7B、C)。しかし、内臓脂肪では PPARγ と同様 に、LPL及びHSLにControlマウスとAE2KOマウスで差はみられなかった(図 7E、F)。以上の結果から脂肪のERK2欠損がHFHSD負荷で PPARγ の低下により 皮下脂肪分化障害が生じ、LPL及びHSL低下により脂肪の蓄積・放出能が低下 した可能性が示唆された。
20
(4)脂質代謝の評価
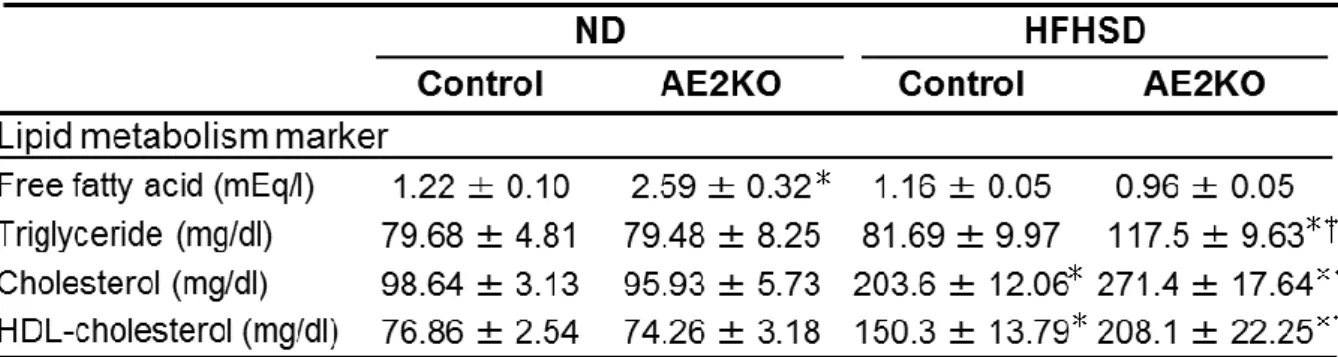
ND群では、中性脂肪、総コレステロール及びHDLコレステロールにおいて、
ControlマウスとAE2KOマウスに差はみられなかったが、遊離脂肪酸はAE2KO マウスが有意に上昇を認めた(Control-ND=1.22±0.10 mEq/l[N=7]対
AE2KO-ND=2.59±0.32 mEq/l[N=8]、P=0.0011)(表3)。Controlマウスは
HFHSD負荷において、総コレステロール、HDLコレステロールの上昇を認め
た。AE2KO-HFHSD群はControl-HFHSD群と比較して中性脂肪、総コレステロー ル及びHDLコレステロールにおいて有意に上昇を認めたが(中性脂肪;
Control-HFHSD=74.81±8.55 mg/dl[N=6]対AE2KO-HFHSD=117.5±9.63 mg/dl[N=6]、P=0.022、総コレステロール;Control-HFHSD=203.6±12.06 mg/dl[N=7]対AE2KO-HFHSD=271.4±17.64 mg/dl[N=8]、P=0.029、HDL- コレステロール;Control-HFHSD=150.3±13.79 mg/dl[N=7]対AE2KO-HFHSD
=222.2±19.88 mg/dl[N=7]、P=0.017)、遊離脂肪酸では差はみられなかっ
た(表3)。これらの結果からAE2KO-HFHSD群はControl-HFHSD群と比較して血 中の中性脂肪及び総コレステロールの上昇が判明した。
21
(5)肝臓への異所性脂肪沈着の評価
肝臓組織への脂肪沈着の程度を評価するためオイルレッドO染色、肝臓組織 中の中性脂肪及び総コレステロール含量の評価を行い、脂肪肝炎の評価として
線維化を評価するためマッソン・トリクローム染色、また血清の肝酵素である
ALTの評価を行った。AE2KO-HFHSD群ではControl-HFHSD群と比較してオイル レッドO染色で脂肪滴の増大を認め(図8A)、肝臓内の中性脂肪及び総コレス テロール含量も有意に増加していた(肝臓1gあたりの肝臓内中性脂肪含有 量;Control-HFHSD=51.07±4.01 mg/g[N=9]対AE2KO-HFHSD=65.4±3.06
mg/g[N=7]、P=0.016、肝臓1gあたりの肝臓内総コレステロール含有量;
Control-HFHSD=5.72±0.55 mg/g[N=9]対AE2KO-HFHSD=9.27±0.50 mg/g
[N=9]、P<0.001)(図8B、C)。またAE2KO-HFHSD群の肝臓組織においてマッ ソン・トリクローム染色で線維化を認め(図8D)、血清ALTは有意に増加して いた(Control-HFHSD=57.06±10.21 U/L[N=7]対AE2KO-HFHSD=
204.50±17.22 U/L[N=8]、P<0.001)(図8E)。以上の結果からAE2KOマウス
ではHFHSD負荷によって肝臓の脂肪沈着が増大し、線維化を伴う脂肪肝炎が進
行したことが判明した。
22
(6)筋肉への異所性脂肪沈着の評価
筋肉への脂肪沈着を評価するために、腓腹筋のオイルレッドO染色を行い、
また腓腹筋組織中の中性脂肪及び総コレステロール含量の評価を行った。
AE2KO-HFHSD群ではControl-HFHSD群と比較してオイルレッドO染色で筋線維 間の脂肪滴の増大を認め(図9A)、また筋肉内の中性脂肪及び総コレステロー
ル含量もAE2KO-HFHSD群で有意に増加していた(筋肉1gあたりの筋肉内中性
脂肪含有量;Control-HFHSD=41.28±8.01 mg/g[N=10]対AE2KO-HFHSD=
75.04±12.47 mg/g[N=9]、P=0.04、筋肉1gあたりの筋肉内総コレステロー ル含有量;Control-HFHSD=1.45±0.47 mg/g[N=11]対AE2KO-HFHSD=
6.96±1.55 mg/g[N=8]、P=0.003)(図9B、C)。これらのことからAE2KOマ
ウスではHFHSD負荷によって筋肉への異所性脂肪沈着の増加が判明した。
(7)インスリン抵抗性及び感受性の評価
ND群ではControlマウスとAE2KOマウスのインスリン濃度に差は認めなかっ た。HFHSD群ではAE2KOマウスはControlマウスと比較してインスリン濃度の 上昇を認め(Control-HFHSD=1.18±0.33 ng/ml[N=11]対AE2KO-HFHSD=
3.06±0.73 ng/ml[N=9]、P=0.013)(図10A)、インスリン抵抗性の指標であ るHOMA-IRの上昇を認めた(Control-HFHSD=8.34±2.17[N=11]対AE2KO-
23
HFHSD=29.72±7.13[N=9]、P=0.0097)(図10B)。AE2KO-HFHSD群でインス リン抵抗性の悪化がみられたことから、ITTを行いインスリン感受性に関して
詳細に検討した。ITTではND群ではControlとAE2KOマウスに差はみられなか ったものの、HFHSD群ではAE2KOマウスでインスリン投与後の45分、60分で 有意に血中グルコース濃度の低下率が低値であり、インスリンの反応が悪化し
ていた(45分;Control-HFHSD=77.05±2.49%[N=7]対AE2KO-HFHSD=
95.33±3.19%[N=7]、P=0.011、60分;Control-HFHSD=79.65±3.66%[N
=7]対AE2KO-HFHSD=107.00±3.50%[N=7]、P<0.001)(図10C)。以上の 結果から、AE2KO-HFHSD群はControl-HFHSD群と比較してインスリン抵抗性と インスリン感受性の悪化が明らかとなった。
(8)アディポサイトカインの評価
アディポネクチン濃度はControl-ND群と比較してAE2KO-ND群で増加を認 め、HFHSD群で低下が認められた。しかしながらControl-HFHSD群とAE2KO- HFHSD群では差はみられなかった(Control-HFHSD=1045.0±112.0 ng/ml[N=
21]対AE2KO-HFHSD=903.8±64.1 ng/ml[N=21]、有意差なし)(表4)。ND群
ではControlマウスとAE2KOマウスにレプチン濃度の差はみられなかったが、
HFHSD負荷によりControl-HFHSD群のレプチン濃度はControl-ND群と比較して
24
増加しており、またAE2KO-HFHSD群ではControl-HFHSD群と比較してさらに増 大していた(Control-HFHSD=33.48±4.52 pg/ml[N=12]対AE2KO-HFHSD=
53.07±3.43 pg/ml[N=12]、P=0.006)(表4)。TNF-α 濃度はND群では ControlマウスとAE2KOマウスで差はみられなかったが、HFHSD群ではAE2KO マウスのTNF-α濃度がControlマウスと比較して増加していた(Control- HFHSD=4.16±0.99 pg/ml[N=9]対AE2KO-HFHSD=8.37±1.03 pg/ml[N=
9]、P=0.017)(表4)。 このことからAE2KOマウスではHFHSD負荷によって炎 症性アディポサイトカインであるレプチンやTNF-α の上昇が明らかとなっ た。
25
第4節 考察
白色脂肪細胞は皮下脂肪と腹腔内にある内臓脂肪とに分類される。内臓脂肪
は皮下脂肪より多くの遊離脂肪酸や炎症性アディポサイトカインを放出すると
いわれているが、内臓脂肪は腹腔という制限された空間にあるため、その増大
には限界がある。一方、皮下脂肪は内臓脂肪のような解剖学的な空間の制限を
受けずに外側に拡張することが可能であり、長期的に多量のエネルギー貯蔵が
可能である。皮下脂肪の機能障害が全身性の炎症に関連があり、皮下脂肪を除
去した場合に、内臓脂肪蓄積とインスリン抵抗性、炎症性アディポサイトカイ
ンであるTNF-α が増加したとの報告がある(42)。また皮下脂肪を腹腔内に移
植すると、インスリン感受性や耐糖能等の代謝因子が改善されたが、内臓脂肪
の移植では改善されなかったことも報告された(43)。これらの報告からは、皮
下脂肪が内臓脂肪とは異なる機能を持ち、Metsにおいても重要な役割を担って
いることが示唆される。
脂肪細胞の分化誘導において PPARγ は中心的な役割を担っている。PPARγ 遺
伝子の強制発現により線維芽細胞から脂肪細胞が分化することや、PPARγ アゴ
ニストにより脂肪細胞への分化が増えることが確認されている(44)。さらに
PPARγ 欠損の細胞は脂肪細胞に分化しないことから(45)、PPARγ は脂肪細胞の
分化に決定的な役割を果たす。肥満のようなエネルギーの過剰な状態では、脂肪
26
前駆細胞の活性化が生じており、脂肪幹細胞から脂肪細胞への分化が亢進する
ため、HFHSD負荷でのControlマウスで PPARγ の発現が上昇したと考えられ、
これに伴い皮下脂肪細胞の大型化が認められた。一方でAE2KOマウスはHFHSD負 荷によって皮下脂肪細胞の大型化の障害が観察され、また皮下脂肪における
PPARγ の発現低下がみられたことからも皮下脂肪の分化障害が示唆された。さ
らには脂肪組織において脂肪分解と脂質生成における2つの重要な酵素である
LPLとHSLの発現が、AE2KO-HFHSD群の皮下脂肪でControl-HFHSD群と比較して 著減しており、このことからAE2KO-HFHSD群の皮下脂肪の脂肪蓄積・放出能が低 下していることが示唆された。興味深いことに、このような変化はAE2KO-HFHSD 群の内臓脂肪ではみられなかった。以上からHFHSD負荷の脂肪細胞ERK2欠損に おいては PPARγ の低下により皮下脂肪の脂肪分化障害が生じ、LPL及びHSL 発 現の低下により脂肪の蓄積・放出能が低下したものと考えられた。AE2KOマウス
は HFHSD 負荷によって Control 群と比較して血中中性脂肪及び総コレステロー
ル濃度の増加がみられており、脂肪細胞の分化障害による脂質蓄積機能低下が
血中の脂質増加に繋がった可能性が考えられた。
脂質は脂肪組織の脂肪細胞に主に蓄えられるが、肥満や糖尿病等の病的環境
において脂肪組織以外の組織に脂肪が沈着することがある(46)。こうした脂肪
を異所性脂肪といい、肝臓や筋肉が主要な組織である。肥満患者における脂肪
27
由来の脂肪酸や生理活性物質は門脈を介して肝臓へ注がれるため、脂肪組織と
肝臓のネットワークは代謝異常における臓器連関の中心的な役割を果たす。本
章において肝臓の評価を行ったところ、AE2KOマウスはHFHSD負荷によって
Controlマウスと比較して脂肪沈着の増大と線維化を伴う脂肪肝炎の進展を認
めた。次に筋肉への異所性脂肪を評価したところ、AE2KOマウスはHFHSD負荷
によってControlマウスと比較して筋線維間への著明な脂肪沈着を認めた。こ
れらは皮下脂肪の分化障害によって脂肪組織の脂肪蓄積能が低下し、本来脂肪
に蓄えられるべき脂質が血中に多く存在し、肝臓や筋肉での脂肪沈着を惹起し
たことが想定された。肝臓による糖放出抑制及び筋肉による糖取り込み増加は
インスリンによる血糖制御の中核であり、肥満やMetsにおいては脂肪肝炎が 進展し、インスリン抵抗性の原因となることは多数報告されている(47-50)。
また筋肉への脂肪沈着には筋線維内脂肪滴と筋線維間脂肪細胞に分けられ、臨
床的には肥満(51, 52)や2型糖尿病(53, 54)で認められるが、特に筋線維間脂 肪細胞はインスリン抵抗性と関連することが報告されている(52, 54)。本章に
おいてもAE2KO-HFHSD群において著明なインスリン抵抗性とインスリン感受性
の低下がみられており、HFHSD負荷における脂肪細胞ERK2欠損による皮下脂肪 の分化・機能障害が引き起こした肝臓の脂肪肝炎の進展や筋肉での脂肪沈着
が、AE2KO-HFHSD群のインスリン抵抗性を助長したと考えられた。
28
Metsにおいてインスリン抵抗性と慢性炎症には深い関連がある。アディポネ
クチンは抗糖尿病・抗動脈硬化作用を有し、脂肪細胞の肥大化とインスリン抵
抗性の悪化に伴い、血中アディポネクチンは低下する(55-58)。本研究におい
てもHFHSD負荷によってControlマウスとAE2KOマウスの血清アディポネクチ ン濃度は低下した。しかしながらAE2KO-HFHSD群とControl-HFHSD群に差はみ られなかった。このことはアディポネクチンが主に内臓脂肪から分泌されおり
(10)、HFHSD負荷のAE2KOマウスでは主に皮下脂肪での分化・機能障害が主体
で、内臓脂肪蓄積にControl-HFHSD群と明らかな差がなかったことが原因であ ると考えられた。
肥大化した脂肪細胞からはTNF-α などの炎症性サイトカインが産生・分泌 され、インスリン抵抗性を惹起する(59)。レプチンは皮下脂肪で発現が多く、
主に視床下部の受容体を介して摂食量やエネルギー消費を調節する(60)。また
レプチンは視床下部のみならず、腎やマクロファージなどの末梢組織にも発現
し、炎症性サイトカインとしての役割をもっている(61)。TNF-αは肥満やイン
スリン抵抗性と相関するとされる炎症性サイトカインであり、培養脂肪細胞の
実験ではTNF-αは主にマクロファージに由来すると報告されている(62)。血清
TNF-α 濃度は、AE2KO-HFHSD群でControl-HFHSD群と比較し上昇していた。こ のことから脂肪細胞ERK2欠損が引き起こした皮下脂肪主体の脂肪分化・機能
29
異常により、マクロファージによる炎症性変化が誘導され、炎症性サイトカイ
ンが増加した可能性がある。さらにAE2KO-HFHSD群異所性脂肪沈着を起こした 肝臓、筋肉等の炎症も、血清TNF-α の上昇に関与したと考えられた。このモ デルでは皮下脂肪分化異常と異所性脂肪沈着、インスリン抵抗性、そして慢性
炎症との関連が認められ、Mets合併症の進展機序を考慮するにあたり興味深い
モデルであると考えられた。
30
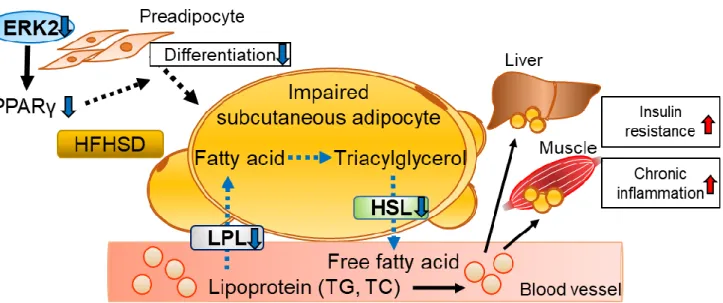
第5節 小括
AE2KOマウスのHFHSD負荷では、皮下脂肪において PPARγ、LPL, HSLの発現 を低下伴う細胞分化・機能障害を呈し、この障害は脂質の血中濃度上昇を介し、
肝臓及び筋肉に異所性脂肪沈着を引き起こし、インスリン抵抗性の増悪と慢性
炎症に関与すると考えられた(図11)。一方で内臓脂肪細胞においてはAE2KOマ
ウスの HFHSD 負荷で皮下脂肪と同様の変化は認められず、血清アディポネクチ
ンの低下はControlと同程度であった。このことから脂肪細胞ERK2はin vivo において、皮下脂肪を主体とした脂肪細胞分化や脂肪蓄積能に関与すると考え
られた(図11)。
31
第3章 脂肪細胞ERK2が血管内皮機能や血圧に及ぼす影響の検討
第1節 背景及び目的
前述したとおり、脂肪細胞ERK2欠損ではHFHSD負荷の条件において、皮下 脂肪分化、脂肪蓄積機能を障害し、脂質代謝異常を介した異所性脂肪沈着が起
こり、その結果インスリン抵抗性増悪と炎症性サイトカインによる慢性炎症が
認められた。肥満患者における脂肪酸や炎症性アディポサイトカインは、肝臓
や筋肉でインスリン伝達を障害し、インスリン抵抗性を引き起こすとされ(34,
59)、これは心血管疾患の基盤である高血圧や血管内皮機能障害に密接な関わ りがある。高血圧の機序として、血管トーヌスを調整する一酸化窒素(NO)の
生理活性が低下し、血管内皮機能が障害されることで収縮期血圧が上昇するこ
とがいわれている(63)。また多くの疾患で血管壁におけるスーパーオキシド産
生がNOの生理活性低下と血管内皮機能障害の主要因であると報告されてい る。血管壁のスーパーオキシド産生は、高血糖、高脂血症、炎症で増加するこ
とが報告されており、前章でのAE2KO-HFHSD群ではこれら多くの要因が存在す る。そこで本章ではHFHSD負荷における脂肪細胞ERK2が血管内皮機能、血圧 へどのような影響を及ぼすか、またその悪化の原因となりうるスーパーオキシ
ド産生及びNOの関与について検討した。
32
第2節 対象と方法
(1)実験動物
前章と同様にControlマウス及びAE2KOマウスそれぞれに対して、ND及び HFHSDを6週齢から30週齢までの24週間負荷し、4群(Control-ND、AE2KO- ND、Control-HFHSD、AE2KO-HFHSD)を作成して比較検討を行った。
(2)血管におけるスーパーオキサイド産生の評価
過去に報告された方法を参照して、dihydroethidium(DHE)染色(Thermo
Fisher Scientific)を使用し、摘出した大動脈におけるスーパーオキシド産 生の評価を行った(64)。マウスから摘出した胸部大動脈から血管周囲組織を剥
離し、3 mm長に分割して大動脈リングを作成し、O.C.T Compoundに包埋した
後に、液体窒素で急速凍結した。10 µmの厚さで切片を作成し、DHE(2x10-6
mol/L)を室温で遮光し30分間反応させた。またDHE染色がスーパーオキシド
産生によるものかを確認するために摘出した大動脈リングをスーパーオキシド
除去剤であるtempol(10-4 mol/L)に1時間反応させた後に、同様の方法で DHE染色を行った。画像はKEYENCE社のBZ-X710(大阪)を用いて、540 nmの 励起光を使用し、605 nmの蛍光波長で撮影した。DHEの蛍光強度はKEYENCE社 の解析ソフトであるBZ-X Analyzerを用いて解析した。
33
(3)血管拡張反応試験
等張性張力測定による血管反応試験を過去に報告された方法を参照して行っ
た(30, 65)。装置として3Dイージー・マグヌス(いわしや岸本医科産業株式 会社、京都)を使用した。マウスから摘出した胸部大動脈から血管周囲脂肪を
剥離し、3 mm長に分割した後、クレブス・リンゲル重炭酸バッファーで満たさ
れた恒温槽の張力測定用フックに大動脈リングを吊るした。バッファーは95%
O2と5% CO2の混合ガスを溶存させて 37℃に保った。初期張力を1.0 gとし、張 力測定はトランスデューサ―で増幅してポリグラフ上に記録した。拡張反応の
前にKCLバッファー(60 mmol/L)にて15分間の過収縮を起こし、その後クレ ブス・リンゲル重炭酸バッファーにて3回洗浄した。L-phenylephrine(10-5.5 mol/L)で15分間前収縮させた後、acetylcholine(ACh)を恒温槽内に漸増投 与し(10-9 mol/L-10-5 mol/L)、AChによる血管内皮依存性拡張反応の張力を測 定した。さらに血管内皮機能障害がスーパーオキシド産生に関与するものかを
検討するため、tempol(10-4 mol/L)を恒温槽に加えて1時間後に、同様にACh 投与による内皮依存性血管拡張反応を行い、血管内皮依存性拡張反応へのスー
パーオキシドの影響を検討した。
34
(4)収縮期血圧、脈拍、血中の酸化ストレス指標及びNO代謝産物の評価 収縮期血圧と脈拍をtail cuff法によって無麻酔下で測定した(MK-2000、
室町機械株式会社、東京)。血清中の過酸化物代謝物(reactive oxygen
metabolites; ROM)の測定を行った。回収した血清をFree carpe diem
(Wismerll、東京)を使用して、derivatives of reactive oxygen
metabolites(d-ROM)テストを行い、血清ROM値を測定した(66)。ROMの測定 単位は、1 CARR Uが0.8 mg/L H2O2に相当する。また回収した血清を用いて、
NOの代謝産物である硝酸塩及び亜硝酸塩(NO2-+NO3-)を専用の測定キット(株 式会社 同仁科学研究所、熊本)を使用して測定した。
(5)統計学的解析
すべてのデータを平均±標準誤差(mean±SEM)で表記した。血管拡張反応
試験の評価には繰り返しのあるtwo-way analysis of variance(ANOVA)後、
Bonferroni法でpost hoc解析を行った。2群比較にはMann-Whitney U検定を 使用し解析を行った。すべての統計解析はGraphPad Prism Software Ver.7
(GraphPad Software、La Jolla、CA、USA)を使用し、すべての検定において
p<0.05を統計学的有意と判定した。