九州大学学術情報リポジトリ
Kyushu University Institutional Repository
ショウジョウバエ概日時計突然変異体の分子遺伝学 的解析
松本, 顕
https://doi.org/10.11501/3159113
出版情報:Kyushu University, 1999, 博士(理学), 論文博士 バージョン:
権利関係:
0
Molecular genetic analysis of circadian clock mutants in Drosophila melanogaster
Akira Matusmoto
Research Center for Higher Education, Kyushu University,
Ropponmatsu,
Fukuoka 810-8560, Japan
Contents
Acknowledgments . . . ... . . 1
General Introduction . . . ... . . ... . . ..... . . ... ... . . .. . .. .. .. . . . ....... . . 2
Chapter 1 ... . . ... . . ... . . ..... . ... . . . ... ..... . . ... ... . ... . . .... . . . ... . . ..... . .. .. . . .. .. . . ... . . . 5
"Chronobiological analysis of a new clock mutant, Toki, in Drosophila melanogaster." Abstract . . ... ... . . ... . ... ..... . . ..... . . ... . .. .... . . ... . . ... . .. ... . . ..... . . ... . . ..... . ... . . ... ..... . . 6
Introduction . . . ... . . .... .. . ... . . .. .... . . . ...... . . .... . . . ... . .... .. . . ....... . . .. . .. . . . ... . . 6
Materials and Methods . . . ... .. .. . . ... .... . . ... . . .. . . .. . ... . . ... ... ..... . . . ... . . ... . . ... . . . .. .... ... .. .. . . 7
Results . . . .. .. ....... ..... . . ......... . . ..... .... .... ..... . . . ... . . ..... . . . ... . . ..... . . ... . . .... . . ... . . 9
Discussion . . . ... . ... . . .. . .. . . . .. .. . . ... . . .. .. . . ... . . ... .. .. . . ... . . ... . 11
Tables ... 15
Figures . ... . . ..... . ... ... . . ... . ... . . . .. . .... ... . . ... . ... . . ..... . . ... . . ... . . ..... . . ..... . .. 19
Chapter 2 ... 3 0 " tirrt1 lengthens circadian period in a temperature dependent manner through suppression of PERIOD protein cycling and nuclear localization." Abstract . . ... . . ... . . ... ... . . ... . ... . . ... . . . ... . . ... .... . . .. .. . . ... ... . . ... . ... ... . ..... . .. 3 1 Introduction .. . . .. . . .. . . .. . ... .. .. . . .. .. ....... . . . ... . . ... . . .. . .. . . .. .... . . ... . . .. .. .. . .. . . 31
Materials and Methods . . . ... . . ... . . 32
Results . . . ... . . 35
Discussion . . . ... . . .... ....... . . ... . . ... . . .. .. . . ... . . . ... . . .. .... . .. .. ... . . ... . . ... . . . 42
Tables . . . ......... . . ..... . ..... . . ........ . . ... . . ... . . ..... ... . . .... .. . ... . ... ......... . .... 48
Figures . . . . ..... . . ... .. .. . . ... . . .... . . ..... . . ... . . .. .. . . ... . . .. .. . . ..... . ... ... . . .. . . . .. . . . 51
References . . . ... . . ... . . ..... . . ... . . .... . . ..... ..... . . ... . . ... .. . .. . . ... . . ... ... ....... ..... . . ... . . 69
ACKNOWLEDGMENTS
I thank Teiichi Tanimura for his instruction and encouragement in the course of this study.
He has made this thesis possible. I thank Paul Hardin for use of his laboratory to perform RNase protection assays and comments on the manuscript. I thanks Michael Rosbash for theper-lacZ strain, Joan Rutila for the rh-per strain, Jeffrey C. Hall for period mutants, and Michael W. Young for timeless mutants. Ralf Stanewsky and Justin Blau kindly provided me with anti-PER and anti-TIM antibody, respectively. Flies for mating procedures and two strains carrying a tim duplication were provided by Bloomington stock center, Bowling Green stock center and Umea stock center. I also thank Dave Allen, Lisa Lyons and staffs of my lab for their technical supports and advice, HaiPing Hao and Jan Qiu, J. C. Hall and M. W. Young for comments on the manuscript of chapter II. I thank Yoshihiko Chiba, Kenji Tomioka and Yoshitaka Kobayakawa for useful discussion.
- p. 1 -
GENERAL INTRODUCTION
Circadian rhythms are universal biological phenomena found in eucaryotes and some procaryotes (for review, see Dunlap,
1999),
and are thought to be an adaptation to environmental cycles. A circadian clock governs the rhythms through physiological and endocrinological processes. The circadian clock has several features. First, the circadian clock keeps its period of about 24 hrs even when there are no environmental time cues. Second, the circadian rhythm can synchronize to various environmental cycles if the period is around 24 hrs. For instance, circadian rhythm can be entrained by the daily change of light intensity, temperature or humidity. A few days of environmental cycle is required to reset their circadian rhythms to a new Zeitgeber (time cue) regimen, and this lag is called "transients". Third, a single perturbation of environment such as light, temperature and some chemicals induces a persistent phase change of a freerunning rhythm.The direction and magnification of phase shifts are depend on both the phase of rhythm when the perturbation is applied and strength of the given pertubation. The correlation between a phase where the pertubation is given and the phase shift induced is illustrated as a "phase response curve (PRC)". Fourth, a freerunning period lengthens or shortens as a function of light intensity, and when the light intensity becomes much higher, organisms become arrhythmic. Fifth circadian period remains relatively constant with a change in temperature of
1
OOC, and the temperature quotient,Q10,
is approximately1.
The biochemical mechanisms underlying circadian rhythms are clearly distinct from biochemical reactions observed in other physiological and developmental events, because theQ10
of those reactions is nearly 2�3.Since these circadian mechanisms have a genetic basis, genetic analysis has provided a powerful tool to elucidate molecular mechanisms underlying circadian rhythm. Organisms contributing toward the progress of understanding the clock mechanism are Drosophila, Neurospora, Arabidopsis, and mice (Dunlap,
1999).
Among them, Drosophila has been a key organism to elucidate the molecular basis of circadian rhythm. Five genes in Drosophila, period (per), timeless (tim), dClock (dClk), cycle (eye), and double-time (dbt) have been identified as clock genes that contribute to a central oscillator mechanism (Konopka and Benzer,1971,
Sehgal et al.,1995;
Allada et al,1998;
Price et al,1998;
Rutila et al.,1998).
The abundance of per and tim - p. 2-mRNA and their protein levels fluctuate in a circadian manner (for review, see Sehgal, 1996), and dCLK and CYC are thought to form a heterodimer to act as transcriptional activators of
per
andtim
(Darlington et al., 1998). PER interacts with TIM (Gekakis et al., 1995), moves from the cytoplasm into the nucleus (Curtin et al., 1995) and feeds back to repress the level of the
per
andtim
transcripts (Hardin et al., 1990; Hunter-Ensor et al., 1996). DBT is a kinase phosphorylating PER, and this phosphorylation triggers a relatively quick PER degradation.
There have been identified mammalian clock genes corresponding to the clock genes in
Drosophila.
Interactions between clock proteins have also been investigated in mammals and the mode of interactions is principally same toDrosophila
(for review, see Dunlap, 1999). However, there are some differences in the circadian system between inDrosophila
and mammals. In vertebrates, there are three homolgues ofper
gene in a genome and their protein levels were reported to fluctuate in the antiphase against PER inDrosophila
(see review, for Whitmore et al.,1998).
Clk
gene was identified through studies of arrhythmic mice mutant induced by a chemical mutagen and its protein level does not cycle whiledCLK
shows a clear diurnal rhythm (Vitatema et al., 1994; King et al., 1997).In
Neurospora,
three genes,fre
que
nc
y(jrq), white collar-] (we-i)
and2 (wc-2),
were identified as clock genes (McClung et al., 1989; Crosthwaite et al., 1997).WC-1
andWC-2
interact to act as a transcriptional activator ofjrq. FRQ, in tum, negatively regulateswe-i
andwc-2
(Crosthwaite et al., 1997). Clock genes in
Neurospora
regulate a negative feedback loop.Additionally,
WC-1
andWC-2
have a PAS domain to interact with its partners (see review, for Ponting and Aravind, 1997). PAS domain is a dimerization domain found in some kind of nuclear proteins including PER, CLK and CYC. Thus, the negative feedback loop at which the molecules with PAS domain contribute is postulated as a pacemaking model. The molecular mechanism of circadian rhythm has been investigated even in procaryote, cyanobacteria (for review, see Dunlap,1999). There is three clock genes reported but none is homologue to known clock genes in eucaryote (Ishiura et al., 1998). However, the molecular model of generating circadian rhythm proposed in eucaryotes can be extended to procaryote because two of three clock genes are transcriptional activators to the other and it acts as a negative regulator to others (Iwasaki et al.,
- p. 3-
1999).
Though we have reached some state to understand how the main gear of circadian rhythm endogenously keeps on moving, little is known neither how circadian rhythm responds to
environment nor how circadian system controls the physical homeostasis. For the pacemaker to generate the temporal information, there would likely be a number of unidentified genes whose products would interact with each other. The present study was made from this viewpoint to determine whether new circadian clock mutations in Drosophila could be induced. As the result, nine clock mutants were isolated. There are two novel clock mutants, one tim allele and six per alleles. I chose two mutants to further analyze, because they showed the most remarkable circadian phenotype among mutants isolated in the present study. In chapter 1, I described fundamental circadian properties of a new clock mutant, Toki, examined under different environmental
conditions and its genetic interactions with the known circadian rhythm mutations. In chapter 2, I described the abnormal temperature compensation oftimnt, which derived from a natural population (Murata et al., 1995) and the genetic interaction with per at both the behavioral and molecular levels. I discussed the molecular mechanism of temperature compensation through the results that ti�t shows abnormal temperature compensation of period and reduces PER and TIM levels.
- p. 4-
CHAPTER 1
Chronobiological analysis of a new clock mutant, Toki, in Drosophila melanogaster.
-p. 5--
ABSTRACT
I have isolated a new semidominant clock mutant Toki on the second chromosome in Drosophila melanogaster. This mutant differs from the wild-type Canton-S in several properties as follows. Larger values are obtained in the phase angle difference
('¢,
the time from lights-off in a24
hr light-dark cycle to an activity offset), the ratio of activity time to rest time
(alp
ratio) and the activity level. The free-running period(,-;) is25.3
hr, one hour longer than in the wild-type. In the phase response curve (PRC), the ratio between the delay and the advance portion is larger and the cross-over point occurs later, although there is no difference in type of PRC (Type I). The rhythm is more sensitive to the light intensity, becoming obscure in darker condition. Toki interact with the other X-linked clock mutation,per, pe,.t and And in such a way that,-; is lengthened and'¢ becomes smaller.INTRODUCTION
The circadian rhythm is under genetic control. Remarkable progress has been recently made in the molecular genetics of the rhythm. In Drosophila melanogaster, five genes have been so far identified as affecting the circadian rhythm. A gene, which identified as the first clock gene, is period (per) on the X-chromosome (Konopka and Benzer,
1971
). Then timeless (tim) gene wascloned as a partner of PER (Sehgal et al.,
1995,
Gekakis et al.,1995).
Remaining three, dCloek, eye and double-time (Allada et al,1998;
Rutila et al.,1998;
Price et al,1998)
were located on the third chromosome. Among them, the per mutations show the most notable phenotypes. Extensive molecular studies have been done on the per gene (for review, see Hall,1990).
On the Xchromosome, another clock mutation Andante (And) is mapped (Konopka et al.,
1991)
but its molecular properties are unknown.For the pacemaker to generate the temporal information, there would likely be a number of unidentified genes whose products would interact with each other in a complicated way. The present study was made from this viewpoint to find a new circadian rhythm mutation on the second chromosome in Drosophila. As the result, a new mutation 'Toki' was isolated. Toki alters almost
- p. 6-
fundamental circadian properties observed under different environmental conditions. The genetic interactions with the known circadian rhythm mutations linked to the X chromosome indicates that Toki is one of the component genes functioning in the per feedback loop.
MATERIALS AND METHODS
The fruit fly, Drosophila melanogaster, was reared on glucose-cornmeal-yeast medium at 24°C under a cycle of 12 hr light and 12 hr darkness (LD 12: 12), hereafter called 'the standard
environmental condition'. The Canton-S strain was used for the wild-type.
Mutagenesis and screenings Point mutations were induced using an alkylating agent, ethyl methanesulfonate (EMS) according to the procedure described by Lewis and Bacher (1968). The EMS-treated second chromosomes were made homozygous using the balancer chromosome SMJ.
A total of 10 24 homozygous strains were established.
Candidates of circadian rhythm mutants were screened through two steps. In the first step, strains that showed an abnormal eclosion rhythm were selected. Pupae were transferred from LD 12:12 to constant darkness. Observation on the number of flies emerging was continued for
3
days beginning at33
hr after the last lights-off(3
hr before the projected lights-on when the first freerunning emergence peak was expected to occur soon). The number of flies emerging during the first 12 hr span of each day was summed
(NT)
and its ratio to the sum for the remaining 12 hr span of each day<No)
was obtained. Strains showing N/ND ratio of one or less were regarded as candidates for mutant. In Canton-S, NL /ND ratio was around four to five. The first step was adopted forefficient screening, because all the previous mutations affecting the circadian locomotor rhythm also affect the emergence rhythm, although a possibility was suggested in Drosophila
pseudoobscura that the two rhythms are controlled by separate mechanisms (Engelmann and Mack, 1978). The candidate strains were then examined in the second step for the locomotor rhythmicity.
Recording of locomotor activity rhythm Male flies of five days old were individually housed in transparent acrylic rectangle tubes
(3
x3
x 70mm). The tube was plugged at one end withagar/glucose medium as food and was sealed with paraffin, and at the other end with cotton. A - p. 7-
moving fly blocked an infra-red beam and the number of bloclcings during each six minutes was recorded using a computerized system (Chiba et al., 1 990). For the first three days, flies were held in the standard environmental condition, and then placed in constant darkness (DD) until otherwise stated.
Estimation of circadian parameters Free-running periods (t) of the locomotor activity rhythm were estimated from the slope of a straight line eye-fitted to activity offsets (Fig.1) which are the most reliable phase. The phase-angle difference ('¢) between the light cycle and the
entrained rhythm was defined as the time from lights-off (18 :00) to the activity offset (Fig.1 ). When the offset was timed later than the lights-off, '¢took a positive value. Another straight line was eye-fitted to activity onsets. The intervals between the activity onset and offset lines were measured on the first and the seventh day of DD. The average of the two intervals was regarded as activity time
(
a)
per one circadian cycle and its ratio to the rest time, p, was obtained. I defined an activity level as the ratio of a total activity per one circadian cycle in DD to that per one LD cycle.Analysis of phase response curve The activity offset, designated as circadian time (CT) 12 (beginning of subjective night), was chosen as the phase reference point of the rhythm. A 15 min light pulse (ca. 2000 lux) was given at various circadian times to flies free-running in DD. Phases were determined by fitting straight lines visually through activity offsets before and after the light pulse, respectively. Difference between the two phases was taken as the magnitude of phase shift and was plotted against the circadian time at which the pulse was given.
Genetic mapping and producing double mutants Told gene was preliminarily mapped using recessive morphological markers, which did not affect the locomotor rhythm. Females
homozygous for Told were crossed with males carrying recessive mutations,
aristaless (a/, 2-0.01), dumpy (dp, 2-13.0), black (b, 2-48.5), purple (pr, 2-54.5), curved (c, 2-75.5)
andplexus (px,
2-100.5)
(Lindsley and Zimm, 1992). Homozygous recombinants thus obtained were assayed forthe locomotor rhythmicity. I produced double mutants carrying Told on the second chromosome and another clock mutation on the X chromosome in standard genetic crosses using the balancer chromosome SMJ.
- p. 8-
RESULTS
On the basis of�/ND in eclosion rhythm, 77 strains were selected as candidates for circadian rhythm mutant from 1024 strains homozygous for an EMS-treated second chromosome.
Three strains were selected on the basis of abnormal circadian rhythms in locomotor activity. One of them,
Toki
("Time" in Japanese), showed the most distinguishable and stable phenotype.Circadian parameters of Toki
I
nToki (Toki!Toki)
the activity rhythm was entrained to theLD
cycle, showing a biphasic pattern peaking around lights-on and -off, and free-ran inDD
(Fig.1 C). The activity pattern, entrained or free-running, was similar to that of the wild-type (Fig.1A). However,
Toki
had larger values in some circadian parameters thanCanton-S
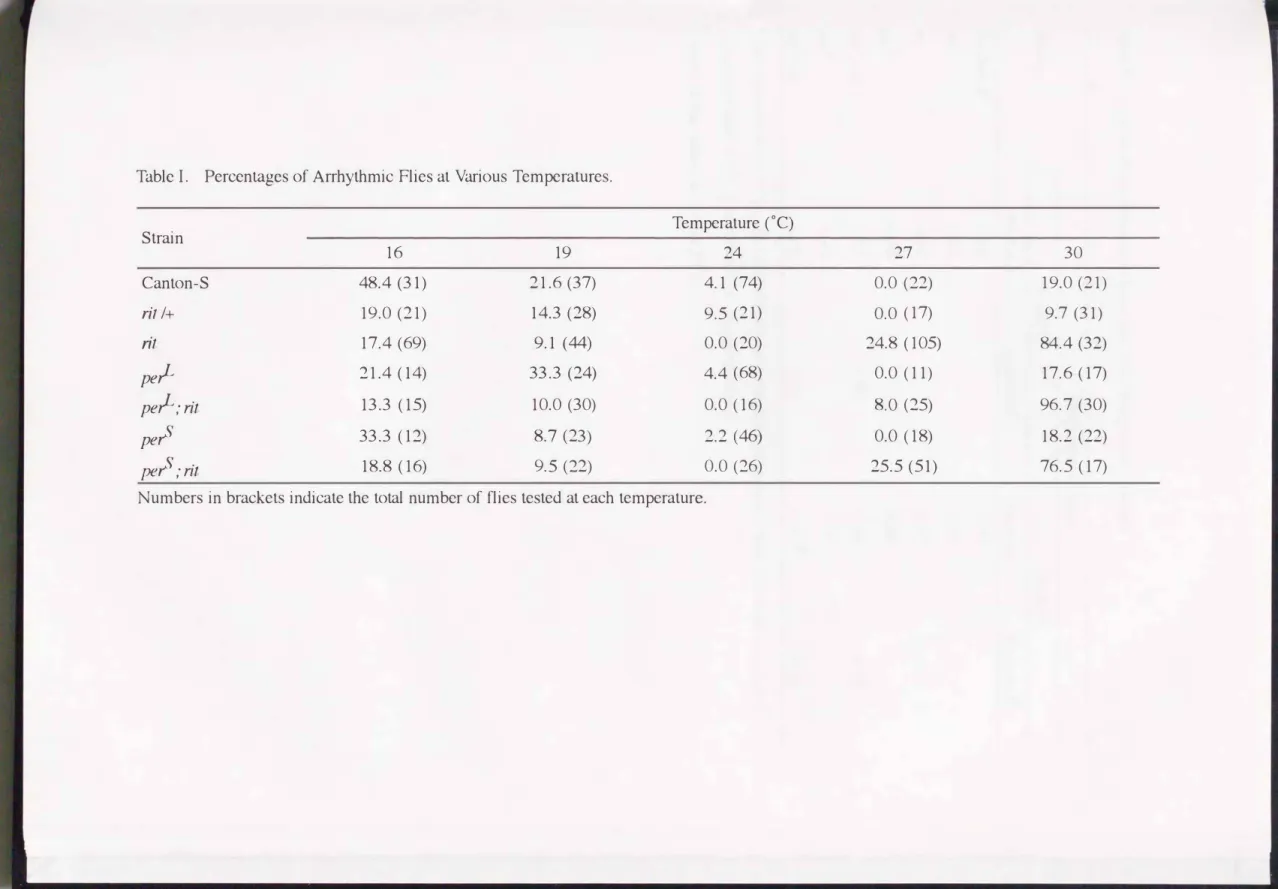
did (TableI);
the average free-running period (t=25.3 hr) was one hour longer (t-test, p<0.01) and activity time(a)
relative to rest time(p)
in one free-running cycle was also much longer (t-test, p<O.Ol).Toki
showed an obscured biphasic free-running pattern asCanton-S
did, but the activity level was three times higher (t-test, p<0.01).When
Toki
was heterozygous withCanton-S (Tokil+,
Fig.1B), -r anda/p
were intermediate between those of the two original strains (t-test, p<0.05; TableI),
but 1tJ and an activity level were almost same as those ofCanton-S
(t-test, p>0.05; TableI).
Phase response curve
Canton-S
showed a typical phase response curve (PRC), which can be classified as Type 1, since the magnitude of phase shifts was not large in both advance and delay directions (upper panel, Fig.2). Phase shifts were minimal during the subjective day from CT 3 to10. Phase delays occurred during the early subjective night from CT 11 to 18, while phase advances were induced during the late subjective night from CT 19 to 2.
The PRC of
Toki
showed the similar pattern toCanton-S.
However, statistically significant differences were found in the magnitude of phase shifts at four circadian phases (Fig.2; t-test, p<0.05), so that theToki
PRC as a whole was drifted toward a delay direction. Thus, the ratio between the delay and the advance portion(D/
A) was 36, much larger than four inCanton-S.
The- p. 9-
cross-over point occurred about two hours later.
Temperature compensation
The free-running periods underDD
at three temperatures, l9°C, 24°C and 28°C, were compared in Canton-S and Toki. The period did not changedsignificantly by temperature (t-test, p>0.05). The temperature quotient
Q10
obtained from 't1/
't28 was1.004 in Canton-S and 0.996 in Toki (Table II).
Light intensity dependency
Canton-S was clearly rhythmic throughout the experiments in DD and dim LL at 0.1 lux, showing the average ts of 24.2 and 26.6 hr, respectively (Fig.3A and Table II). However, under LL at 1 .0 lux, most flies were rhythmic only for the first several days, showing 't of 28.1 hr (Fig.3B). As the light intensity was increased from zero to 0.1 lux, a/p ratio was also increased from 1 . 7 to 2.0, but the activity level was significantly lowered from 1 .0 to 0.8 (t-test, p<0.05). At 1 .0 lux, the a/p ratio could not be obtained because activity onsets were obscure, and the activity returned to the original level atDD
(t-test, p>0.05).Toki was more sensitive to light intensity so that the damped rhythm, which was observed at 1.0 lux in Canton-S, occurred even at 0.1 lux; most flies were rhythmic only for the first several days (Fig.3A). Also at 1.0 lux, all flies were circadian rhythmic for the first several days (Fig.3B).
In this case, the manifested rhythm showed a complicated pattern, so the exact period could not be detected visually or by chi-square periodogram. Thus, the free-running period could be obtained only in the two darker conditions, increasing from 25.5 hr at zero lux
(=DD)
to 26.0 hr at 0.1 lux (Table II); the rate of increase was lower than in the wild-type (t-test, p<0.01 ).Mapping of the
Tokigene
Table III shows the result of recombination mapping of Toki using recessive markers. The locomotor rhythm was normal in recombinants showing b prphenotype. Reciprocal recombinants in this region showed Toki phenotype. These results suggest that Toki is between band pr.
Comparison with the known
xchromosome-linked mutants
Toki, the X-linked mutants and - p. 1 0 -Canton-S were compared for four circadian parameters: 't, '¢, a/p and activity level (Table I, Fig.4).
-r becomes longer in the order of per, Canton-S, Toki, And and per (Table I, Fig.S). In proportion
to this change,'¢ increased from -0.9 hr in per to 10.4 hr in per (Fig.S); in other words, daily activity became to terminate later. Pearson's correlation coefficient (r) between the two parameters was 0.843 (N=S) which was close to the significant level (0.878 at P=O.OS) and Spearman's rank correlation coefficient was 1.00. Another high correlation was observed between a/p and an activity level (r=0.965, P<0.01).
Genetic interactions In double mutants of Toki with each of the three X chromosome
linked mutants, all the parameters examined were affected in a similar way that 't was lengthened (t-test, p<0.01), '¢was reduced (t-test, p<0.01), and a/p and activity level were increased (t-test, p<O.OS)(Table I and Fig.4). Correlation coefficients between two of the four circadian parameters were calculated among four strains carrying Toki mutation. The a/p ratio was highly correlated with an activity level (r=0.960, P<O.OS). Correlation coefficient between 't and'¢ was 0.948 close to the significant level (0.950 at P=O.OS).
DISCUSSION
In the present study, chemical mutagenesis was utilized to isolate mutants. An alternative method is to use the inserted mutagenesis using P-element. This method has several advantages; the loci of interest can be immediately mapped by in situ hybridization using a P-element DNA as a probe and it is possible to clone the tagged gene (Cooley et al., 1988). However, mutants in some gene can not be recovered by insertional mutagenesis because of the preference of the P-element insertion site (Kidwell, 1986). I have screened about ten thousands strains with a single or multiple P-element inserted on the autosomes. However, no clock mutant has been obtained so far. Chemical mutagenesis induces mutations in any genes and, therefore would be more effective way to obtain mutants.
I have obtained several clock mutant candidates besides Toki. Some showed a slightly - p. 11 -
long-period, others showed a scalloped free-running pattern or damping rhythm. However the phenotype of these mutants were unstable, that is, the abnormal phenotype can not be detected in all individuals examined. Strains with these unstable phenotype are not amenable to genetic analyses.
Told showed significantly longer free-running period and was analyzed in detail. This mutation showed larger values in four circadian parameters; -r,
a/p,
'¢and activity level (Table I). The-randa/p
phenotype showed semidominancy, while the others showed recessive character.By analysing to the PRC for locomotor rhythm of per mutants, Orr (1982) reported that a relatively light-insensitive region (the "subjective day") is lengthened in per and shortened in per, while the "subjective night" remains unchanged, showing a normal 12 hr duration. In Told, the difference in free-running period from the wild-type is too small to decide this inconsistency as in
per1k
(Dushay et al, 1990) and And (Konopka et al., 1991). Some clock mutants of Drosophila show a greater phase shift than the wild-type does, as is typically shown in the eclosion rhythm inper
(Konopka et al., 1991; Orr, 1982). In Told, however, phase delay is larger but advance is smaller. In addition, the relation between D/ A ratio and -r is against the rule found by Daan and Pittendrigh (1976) in nocturnal rodents; -r is longer but D/ A ratio is larger as compared with the wild-type. Whether this is an inconsistency between the nocturnal and diurnal animals deserves further study.There are two kinds of mode of temperature-compensation in clock mutants; one is well compensated
(Q10
is very close to 1.0) and the other is relatively incomplete(Q10
is relatively apart from 1.0). The wild-type, And (Konopka and Benzer, 1971) andperctk
(Dushey et al., 1990) are classified into the former, while two alleles at per locus in Drosophila (Konopka et al., 1989) and a number of alleles at.frq gene in Neurospora (for review, see Feldman, 1982) are into the latter. My mutant belongs to the former group, showingQ10
approximating 1.0.Aschoff (1960) described an empirical rule, called Aschofrs circadian rule (Hoffman, 1965), where frequency,
a/p
ratio and a total activity increase with increasing light intensity in diurnal organisms and decrease in nocturnal organisms. It has been reported that, in Canton-S, frequency is affected by light intensity in the opposite way to this rule (Konopka et al., 1989). This wasconfirmed here and was found to be so also in Told. But, as the light intensity increased, -r became - p. 12 -
longer in Toki to less extent than in the wild-type. Furthermore, Toki became arrhythmic in brighter condition ( 1 lux) than the wild-type. These observations suggest that, as in pe� and per (Konopka et al., 1989), Toki requires less light than in the wild-type for both maximum lengthening of period and production of arrhythmia .
On the basis of the study on conidiation rhythm of Neurospora, Lakin-Thomas and Brody (1985) described that there are two modes of effects; additive and multiplicative when different loci affecting the same quantitative character such as --r have no interactions and affect the phenotype independently. On the other hand, when two loci interact, they affect the phenotype in two ways;
epistatic (or hypostatic) and interactive. In Drosophila, only an additive effect was reported in clock mutations (Orr, 1982), suggesting that they act independently on a pacemaking system. Toki is neither epistatic nor hypostatic to per and And with respect to--r, because the double mutants
showed a longer period than the corresponding single mutant (Table I). The difference in --r between the double mutant and the corresponding X-linked single mutant becomes larger as the latter shows Ionger--r (Table
1).
Since --r is only one hour longer in Toki than in Canton-S, the predicted values of�in the additive and multiplicative effects are very close to each other in all of double mutants. This makes it difficult to judge whether the observed value is the result of additive or multiplicative effect.
In general, when a circadian oscillation is entrained to an environmental cycle, there is a positive correlation between� and'¢ (Pittendrigh 1981 ); when --r becomes longer, '¢ becomes larger.
My observation (Fig.S) suggests that this holds true among inter-strain variations of--r and'¢.
However, as far as the effect of Toki on each of the other mutants is concerned, there seems to be the opposite relation that the lengthened --r is accompanied by the reduced'¢ (arrows in Fig.S). The
�-'¢ relationship may depend not only on --r, but on the other circadian properties such as wave
form. The discrepancy should be investigated carefully in future from this viewpoint.
Our data is still insufficient to decide whether the Toki gene product is directly involved in the pacemaker function, although the fact that the Toki gene interacts with other clock genes suggests that this gene cooperates with other genes to express the rhythmicity under LD as well as under DD. It would be possible to argue that the product of Toki controls the locomotor activity
- p. 13 -
level, and that the change in the activity level by the Toki mutation resulted in the change in another circadian parameters. To extend further the study of Toki gene, it is necessary to map the Toki gene cytologically. If the Toki gene is directly involved in the pacemaker system, it is expected that the allelic mutants with more severe phenotype would be isolated as in the case of per and And gene.
The result of recombination mapping of the Toki gene suggests that the gene is located somewhere between band pr in which clock mutations have not been isolated. I am in the process of doing mapping experiments using several chromosomal aberrations. Another important issue is to
examine whether circadian rhythm in eclosion or ultradian rhythm in the courtship song is affected or not, as has been shown in per (Hall, 1990) and percik (Dushay et al., 1990). The molecular studies on the Toki gene should help to understand the circadian mechanism at the molecular level.
- p. 14-
TABLES
- p. 15 -
Table
I
Circadian parameters (mean+SD) of clock mutants and the wild-typeCanton-S.
Strain
N 1:1lJ a/p Activity
(hr) (hr)
Canton-S 31 24.2+0.2 0.4+0.2 1.7+0.3 1.0+0.4 Toki/+ 28 24.6+0.2 0.4+0.2 2.4+0.9 1.0+0.6 Toki 25 25.3+0.3 0.6+0.3 4.2+1.8 2.9+1.1 per5 10 20.3+0.9 -0.9+0.7 1.6+0.8 1.2+0.4 per5;Toki 12 21.0+0.6 -1.3+1.3 2.1+0.6 1.3+0.5 peyL 10 28.5+0.7 10.4+2.5 2.1+0.8 1.6+0.6 peyL;Toki 10 30.6+1.6 7.7+0.8 3.0+1.8 2.0+0.7
And 9 25.4+0.3 2.4+0.8 2.4+0.5 1.4+0.2 And;Toki 8 26.3+0.7 1.4+0.6 3.2+1.0 2.4+1.6
N: Number of animals
1:: free-running period
llJ:
phase angle difference between the lights-off and the entrainedrhythm
a/p:
the ratio between the active phase and the rest phase Activity: the ratio of a total activity (times) per one circadian cycleunder DD to that per one LD cycle
- p. t6-
Table II
Strain
Canton-S
Toki
Free-running periods (mean+SD hr) of Canton-S and Toki in different environments.
Light Temperature
intensity
(lux) 19°c 24°C 28°c QlO
O(=DD) 24.2±0.3(11) 24.2±0.2(31) 24.1±0.3(14)
1.004
0.1 26.6±0.5(20)
1.0 28.1±1.8(10)
O(=DD) 25.1±0.3(11) 25.3±0.3(25) 25.2±0.3(14) 0.996
0.1 26.0±0.4(11)
1.0 arrhythmic( 14)
Numbers in parenthesis indicate the number of animals.
- p. 17-
Table III Recombination mapping of Toki.
Phenotype of Recombinant
Wild-type
b pr 40
+ + 5
b + 2
+ pr 14
- p. 18-
locomotor rhythm Mutant
4 44
1 4
FIGURES
- p. 19 -
FIG. 1 Locomotor activity records at 24°C for single individuals from the following strains:
(A)
wild-typeCanton-S, (B)
heterozygousToki
and (C) homozygousToki.
Flies were held in LD12:12 for the first three days and then transferred into DD. Light regimes are indicated above actograms.
White,
light;black,
dark. Three circadian parameters ('t,'¢
anda/p)
were visuallyestimated on these double-plotted actograms, as shown in
A.
Note that homozygousToki
(C) shows larger values in all the circadian parameters than the wild-type(A)
and heterozygousToki (B).
Heterozygote shows intermediate values between the wild-type and homozygous
Toki
in-randa/p.
- p. 20-
B Toki/+
C Toki/Toki
Fj������---�-�, .. .r,tr-•w-ww--rt�j--Tjrt--nl .. ._� ... --r---�---������--.rw-���---:�--�\�:�.nj�j--�����---w.�·=,l
• • I atrTTWM I I II •111 I M a II I - M II I M I t I *I
r-WTI - W II IM I II tttMM Wt I ·---- WIIMWM ...
�--- ... ··- ..-.. ··- --· -
. -�----�._ -·. .. -
._,.,. ..-...-..r-•r-ra -
__._ • .-...,___._,_,_ __,ill ·--··---
---·Ill ·--··-- I**-- -
24
I W * M IM WI MM •• _....111aa -·
** Ml ·- •• _..WTiiiMM - --11_11 ______ 1 I MIMWMa
•-11-nw .. .-.r-.-wt .. _ 1 .. ..-nr-...-.-wt 1 --·-
- ,--•nw•• w •• 1 -- • .. •• ''' 111!11!'_,..
MTni ···- -·-·-�Tlll·-
_,_._�.,., Ma ... aa II -- I
WMM-1 WMII -- I
12 24
Time of day
12 24
LD DD
LD DD
LD
DD
FIG. 2 Phase response curves for the wild-type and Toki induced by 15 min light pulses (ca.
2000 lux). The mean± S.E.M. calculated by collapsing the data in two hr bins was plotted as a
function of the time of day at while the light pulse was given. Values of Toki marked with asterisks (*)were significantly different from those of Canton-S
-l). 22-
2 Canton-S
!
_,
_,
0 ·-·-·-+
-2 '!
...-...
en Jo-
..c
-4
_..
.t::
..c
(f) Q) en
ctS
2 Toki
..c
a..
0
*'t
-2
'',r-'\
,
.-4 '+,
0 6 12 18 24
Circadian Time (hrs)
FIG. 3 Double plotted actograms of Canton-S (left panels) and Toki (right panels) under LD, dim LD and constant dim-light conditions. For the first several days light intensity was at least 50 lux, then lowered to 0.1 (A,
C)
or 1.0 lux (B,D). All flies could be entrained to dim LD cycles.Flies except Toki at 1.0 lux show longer free-running periods than those in DD. The light intensity becomes higher, the period of Canton-S becomes longer. For mean values, see Table II.
- p. 24-
Can.ton-S
;��{:{�.t>Z'..;.>r���� ::�����
�::: X«-���£.���K������XY.J'..<510l
A 0.1 lux
I
.-
I 8 Mi id I I I I lilt liliM IIl • •• .-.mJJ Tl TJTTIIII- II I I II Iii I ft ill J-
8 1: lux
I M II I M lA I I I I J MIM •- II M Ill -
����iili:�����tt1����!:iiitw;��l:�;�
Toki -
-
�
D
I II I-· I •11111 ililih 1111 •. T •tllliili-•1••11-HIT I-TTU��
AIAI8ii-Mia•l _.,.If I I 1111.1111.11118
..
, • .,... -J-.J-IJa••rWll" -·••n IIIIIIIWIIIMIJIII u• •••• MIMMIII..
iill WWIWIIIIIIIIIMIII8iiillriilri1Mili IIIIIMillihi.llilillill IU lh IMIHIUIIIIIIIM lh 11118111 I IIIIMIIIMI*Iiillill I ih JW I Ml itillfiili 118 II A I ill IIIII •1111 I II II IIIMD I IJ ltifi IIIIIIMJIIIIIIIIIH ill II • I iii I ltN I M fill I I I II h lW N I if lith I I II II Mil II I Ill I M I II I 11•11 ii h I II I I Ifill II I II I ill I II I Ill I I iii IIIII II II II Ill H II 111811tllllliilltttllliillliill iiliilidiliilliiliillllilll IIIIIIIMillii II 11111181 1111111 1111811 I U Jill I Jl ill IIIIMIIAI II II hliMI lrtiiii 1111811 I h till I214 I
24 I I
24 24 I
24 I .
24
Time of day Tlme·ot day
LD dimLD dimLL
LD dimLD
dimLL
FIG. 4 Double plotted actograms at 24°C of six strains representing three clock mutants
linked to the X chromosome (upper panels in each figures) and their double mutants with
Toki
(lower panels) .
(A)per andper;Toki, (B) per andper;Toki
and (C)AndandAnd;Toki.
As compared to single mutants, free-running periods of corresponding double mutants are lengthened by the effect ofToki.
For circadian parameters, see Table I.- p. 26-
A
B
c
I 1 I .r--.---=-..,,--.1 �� ·ri 1 ��-=-�rrr--.1,_----
-r-TWI ill I I I I ..--r-ra•ltar---1•-IIJI1111 •
-"TOaMiifi I* IIWIIII Mil lltmmrnr.-rw--.·--lliUIIIIII Milln rmwmm-nt'rlr-.•--n•n• 11 It 1 w-nmmtrtn,.--•-•mtnmtttrTrltn•nmr-nr••
'"''"',.---•mrmmnrrt 1111*""" n1•11 ttTIIttmrrannnnllrtltntlnl•
• 1111rn IITfllfllmltmTTtlntllt�llt tn!lltla,trtmTimllnm ltiiT--I••r•
Mnlt...,ll11lltYliiWYrm1rlfWTMI-I•nm nrrrr.r-rmrr11 -.---IWIIllntm-rmnwm
• IIIII I 181-II!WrW -· MMAfiWIRiflm11TlT111TnhTnmw MIJilmtJriiiMlnm•ITTlWJIIII..n tntnlnltw Mlll1nmrnrtmlm•!T'T1WI111•• tmntllttrnniiYit•lllllltalr•-•1
111111 mnnniiTII*IIflii .. IT'I-••• ram•-mllll1rwrTnmTM•r�tllllfnnm
Jll1111M 1 lt*llflat """ •••11 lilllnmnnnrmnnMT111mtn••nT'MII•IlJinmtwTmrm-mmnrrtrt th iriWIIWNI ill MWIIi&l--.mtflll'WmTlmllTITil111 PI hi lilt 11f111TJT-n M I I Ill lilt ltlilllllTWrrmrJO rlllliM hi 11111 II 8 II I IIIII lllllillnMJrliiiWll
-1 II I __ Ml ... M
... II -·MlWWM••al M II I I II--
IIIWI I II I I I II - •n I II I II I -nil •••
Mt I M I 11 I -i1111·--l • • I • .-..wr-.- --· • • I • ..-.ar-.- ---"1,...,----.11 I M .- w--.,..,-.11 1 •----••-• , ... ,-trW---..
-.n -- •••• n • 1.--.-. e-. wrar••- .--. •rwr-r
---..-.---·-- ·--···1 . • -. 1... *-'�...-
• • .. ,.. •• MIITI'It-..--arM I I * -I I ···-· - -..wr•r--r • •• 1 • •-• __ ,,,.,�_." 1 1 -•-••n-11•-•
IMMIWY �-.n 1 - w-w•n-111--1--t 8---rr--l-•trnn•r-n•••
-.-*'"• 11 1 ••r•m•,_•..,•••-•--... n--rnn-••••rnra•
�·-·--•.wn,...-namM•tmnra•
'·.
I I I M I I n I I I *I M **
II I M I t II I I i Mt I M •• WI I -· f II 111 H I I M I II I *I I -1 I II Ill M I I M I II - • r • Mlh iii I I I I II -Ill 1_,- M ••• Mlh ttl I It iII -iii 1-iliMi Ril I 1111111 i I Iii Ill th i --- liM II tllililii iMill M I --1 II lllilliili illlhMIII-11.-.--..
_, 11 I 811 I I I ill I I I llli Ill-Ill I -· I I M I I II inlll1illllfh1ITlllmwnMW-M
•• I I M II II iltllnwMI1fllWm.-rtMM Ml I MMII I Ill I I I 8-lllhlnll1l Mil *Mil i Ill I II i-llrtTTlnt111111··-Mam-·-il I I I I I tr W1111Wi·�---�.---. I II I illfllhllnl•t,-.1111-111MI•.-t�._.._Wif8111 rrtnnrn•n•i11�111Wl••••--••••JniTTlrlmnnwJtnWlirnww.--••�
I iII n II I I •rtlf811111ill18W.--·-,_--ftlfllll81ilnl1�trlrMII-rtrlW.Tlf"lli ... IIIWW8lli llttrTT18'WrMfll_,....,.. r-�---,_IT'IIIJIIIIl8-.riiTTMIIWI.i11il ---�IT'W IJIIWI8--IITTWII*IM118l Mll m•• -ntl- .Watwwtlillllf!W 1n·1a 1·a.t1
• ""'_. ....-mw • ., .. _"""*"• .rrta ..-.... ....-w -n•mn rmna·11a·wnlf .. , •• ,,,., trM
.rr,:,::.-::.":"�r:���=·.;.·.:�=�:�!��r:�·�����.n,:.� -1ntlttmran 1 1 11 1 •n•--..,•-.rt..-. ••rrllTTltlt••••• •J'IfWirlllriiiYitfW8'i11•m.I.-:J ,.,.,....,... ... 111- ••• lllltli88'i11_1TT111...,
Jrlllnrmnn I ill I I I I lh til ta II W.lrllr-trTl itiiiNt I lhJIJiiii IMIII II lliliMrr-llrTIIIIII Iii IIMiiilliiilll lllrtT'IUI-11 IIIII r11 It Ill n•illlllll iii I Ill Jmll-11 ill II ar.-JTTTTlllti I I I I II lni'WI--illm-mr n1 II'TfTtl*lt I I I I II lnrw..-..-mrrr-mnllrllliWWl•ll I Mil Wrnww-•t..--n-TrTI in-lilt MtfWII 1 •• 1 • mw.--•m•-.rmn•rnrn n•tmmrrnl..-..nn••li-IIMmt•-•n
�rtn1Wmnlilhila *"''**ll•twMTnr.-•rnrtrm-.ttrnrwaumnatmwnt•l•'**'-'**fW*
"'lrTTlrlirmrwnannwtttTWnratllll1I•,_,.I"T'W**lfM'Tn•liWTIITfTWiw-lll-mti"lm ---•
MhWniWliWIIIIIIMIIIMNIII!WIIf -- II II II lilliiWIW-Il'RliMf8Mi-- li II it lilt 1181M-Iiill .. f8MI-MIMM- llWJTTTWIItnWPI11MMM11li"iMl111a
·--- IIWtil IMMhiiPinii •• IIIMIIM MnWM 11111-MM*iiii-IMMI•
-••• •n••-••••••n-••• •-••-•,_. •n•••.u ••••mnr ..
WIW-W 111181811 IDIIWJJ--.nl...,._ •rnmr.-rmw
IWIIIIWHM- IMIIIIIIIIIIIII...-YfiiiWT-... 11. _....
-.-wn·-· ... -····-.n..-••1- --
···--·--···- ____ ...
-- ···-- --··
---n II
24 I
•
24 I
Time of day
•
24 I
pers
DO LD perS;Toki
lLD perL DO
LD perl;Toki DO
LD And
DO
LD And;Toki
DO
FIG. 5 'ljJ is plotted against,; in each of eight strains. Filled circles: three X-linked mutants
and the wild-type Canton-S. Open circles: four strains carrying Toki on the second chromosome.
There is a positive correlation between,; and 'ljJ (r=0.869, p<0.05). Note that Toki lengthens,; but reduces 'ljJ of the flies carrying X-linked clock mutations (arrows).
- p. 28-
0
pel
•
pei-; T�O
25 30
T (hours)
CHAPTER2
timrit lengthens circadian period in a temperature dependent manner through suppression of PERIOD protein cycling and
nuclear localization.
- p. 30-
ABSTRUCT
A fundamental feature of circadian clocks is temperature compensation of period. The freerunning period of
rits
u(ti�t),
a novel allele oftimeless,
is drastically lengthened in atemperature dependent manner. PER and TIM protein levels become lower in
timnr
as temperature becomes higher. This mutation reducesper
mRNA but nottim
mRNA abundance. PERconstitutively driven by the
rhodopsin]
promoter is lowered inrit,
indicating thatti�t
mainly affects theper
feedback loop at a post-transcriptional level. An excess ofper+
gene dosage can ameliorate allrit
phenotypes including the weak nuclear localization of PER, suggesting thattimnt
affects circadian rhythms by reducing PER abundance and it's subsequent transportation into nuclei as temperature increases.
INTRODUCTION
The circadian clock keeps its period even when there are no environmental time cues. The clock's freerunning period remains relatively constant with a change in temperature of
1 0°C,
and the temperature quotient,Q10,
is approximately1.
The biochemical mechanisms underlying circadian rhythms are clearly distinct from biochemical reactions observed in other physiological and developmental events, because theQ10
of those reactions is nearly2�3.
Although the molecular mechanism to generate circadian fluctuation has been extensively studied, there are only a few molecular studies in
Drosophila
on the temperature compensation mechanism. At a behavior level,per
mutants affect not only period length but also temperature compensation;perr andper
mutants slightly shorten and lengthen their periods, respectively, as temperature increases (Konopka et al.,1989;
Ewer et al.,1990;
Konopka et al.,1994).
Several molecular studies suggested that the temperature compensation is closely associated with PER.Huang et al.
(1995)
reported that PER can undergo a temperature independent intramolecular dimerization, while Gekakis et al.( 1995)
showed that PERL exhibits a temperature dependent defect in binding to TIM although the molecular interaction between TIM and PER is temperature compensated. Moreover, an allele of thetim
gene,timsL,
can compensate a temperature dependent- p.
31
-period lengthening of
per
(Rutila et al., 1996). The length of the Thr-Gly repeat in PER is also reported to affect the temperature compensation (Sawyer et al., 1997).I isolated
ritsu (rit),
a clock mutant on the second chromosome from a natural population (Murata et al., 1995). I have now investigated features ofrit
and its interaction withper
andtim
at both the behavioral and molecular levels.rit
shows abnormal temperature compensation of period and reduces PER and TIM levels. Molecular genetic analyses show thatrit
has a point mutation in thetim
gene that leads to a single amino acid change, indicatingrit
is an allele oftim.
Since an excess of theper
gene dosage ameliorates the weak and delayed nuclear localization of PER as well as all other phenotypes ofrit
at both the molecular and behavioral levels, PER abundance in nuclei seems to be a key factor in the temperature compensation mechanism.MATERIALS AND METHODS
Stocks, locomotor rhythms recording and mating procedures. Flies were kept under LD 12: 12 at 24 °C. Can
t
on-S
was used as wild-type. Double mutants betweenrit
andper
mutants weresynthesized by standard crosses. Flies were grown at 24 °C. Locomotor activity was recorded in the same way described elsewhere (Matsumoto et al., 1994). The period of a locomotor rhythm was calculated by the chi-square periodogram analysis (Sokolove and Bushell, 1978). The mating procedure for the recombination test between
tim
andrit
were done as follows (see Fig. SA). To produce flies carrying both therit
mutation and theper-
lacZ fusion gene on the secondchromosome, I crossed
rit; ry
females toper-SG:3; ry
males which carry theper-
lacZ fusion gene on the second chromosome. These strains carriedry
mutation on the third chromosome and this eye color should be rescued if a fly has theper
-lacZ fusion gene. After two generations I selectedrit
per-lacZhomozygous flies based on the
ry+
eye color and the long periodrit
phenotype. The four lines were selected as arit per-
lacZ strain.rit rh-per
strain was produced according to the standardmating procedures usingSMJ/Sco;
TM3/Pr
(Lindsley and Zimm, 1992)).+/w+Y; rit,
C(J)DX,y w f lw+Y; rit, Dp(2;Y)odd4·31; rit, Dp(2;Y)odd2.31; rit
andw; rit
strains were produced the according to the mating procedures described elsewhere (Matsumoto et al., 1994) with the minor changes. The- p. 32 -
recombination test was done by two different mating procedures. The rit/tim females were mated to tim01 males in one cross and mated to SMJ!Pm males in the other crosses. SMJ/Pm flies show a normal rhythmicity, and were designated by+/+ in Fig. 3A (right). In both cases, progenies from these crosses were then monitored for locomotor rhythms at 30°C. If the recombination between tim and rit occurs, there should be rif tim+ /tim01 progenies whose rhythm is normal in the former cross.
In the latter cross, the double mutant ofrit tim!SMJ or rit tim/Pm whose rhythm is abnormal could be obtained if recombination occurs.
RNase protection assay. Flies were entrained in LD12:12 for five days before they were collected. Total RNA were extracted from 50 fly heads in 500 Jll extraction buffer (15 mM NaOAc,
5 mM EDT A, 1% SDS, 0.01% diethyl pyrocarbonate, 50 mM Tris, pH 9.0). RNase free DNase (Boehringer) were used to remove contaminated DNA. RNase protection assays were done as described elsewhere (Hardin et al., 1990) with minor modifications. I used the per5 and TIMAX1 probe and rp49 as a control. The per5 probe is a genomic fragment of the per gene containing about 210bp (per5849-6060) of the per exon5. TIMAXI is a eDNA fragment of the tim gene from 4963 to 5192nt (Sehgal et al., 1995). I found that the abundance of rp49 in samples obtained at 30°C was half as much as that at 24 OC, although the reason was unknown. So each measurement was
nonnalized by the value of the peak level in wild-type at each temperature.
Immunoblot analyses. Protein extracts were made from 50 fly heads for each time point as
in Edery et al. (1994) with minor modifications. E ach sample was homogenized in 15 Jll ice-cold extraction buffer, and the tip of homogenizing pestle was rinsed twice with another 15 Jll extraction buffer. Amount of proteins in total of 45 Jll extraction buffer were measured by Bradford protein assay system (Bio-Rad). After the measurement, extraction buffer was added to make protein concentration to 5 Jlg/Jll in each sample. Five microliters of 3 X SDS sample buffers were add to 10
�-tl of samples and boiled for 5 min. Supernatants were loaded on 5% SDS-PAGE gel. Western blotting was done as described (Edrey et al., 1994) with minor modifications. I used anti-TIM antibody gifted by J. Blau diluted in 1:5000. HRP-conjugated anti-Rat lgG antibody (Cappel) was used as a secondary antibody with diluting 1:3000. For quantitating of chemiluminesence
(SuperSignal CL-HRP Substrate system, Pierce), an exposure on X-lay film (FUn FILM) was
- p. 33 -
digital-imaged by Densito Graph (ATTO) and quantified by Nlli-image (Nlli) software. After the exposure, the membrane used was incubated for 3 hrs in the substrate buffer in order to eliminate theHRP activity. Then the membrane was used to detect PER abundance as follows. I used anti
PER antibody gifted by R. Stanewsky diluted in 1:10000. HRP-conjugated anti-Rabbit IgG antibody (Cappel) was used as a secondary antibody with diluting 1:3000. The exposure and quantification were done as described above.
Histology. Expression of the per gene in rit flies were assayed histologically using the per-lacZ fusion gene as a reporter. Flies at ZT18 and ZT6 were frozen in a liquid nitrogen and mounted into O.C.T. compound (Tissue-Tek). Sections of 10 �m were cut and stained by X-gal. The staining procedure was done according to Liu et al. (1988). Head sections of wild-type and rit flies were embedded side-by-side on the same slide to compare the staining profiles of the two strains.
Photographs were taken with a Zeiss AxioPhot microscope. For fluorescent immunostaining with anti-PER antibody, the white eyed strain was used as wild-type to eliminate a background
fluorescence of eye pigment. For the same purpose the double mutant ofw; rit was used. The sections (14 Jlm) of two strains were embedded side-by-side on the same slide to ease to compare the strength of signals. Anti-PER antibody was applied at dilution of 1:15,000. Anti-rabbit IgG conjugated to peroxidase labelled-dextran polymer (En Vision+ , Dako) was applied as a secondary antibody. Signals were enhanced with FITC-Tyramide (NEN). TSA reaction was usually done for 7.5 min. In order to obtain a maximum amplification, it was extended to 15 min. Counter staining of nuclei was done by propidium iodide (1 �g/�1, Sigma) after RNase treatment (10 �g/�1,
Boehringer) for 30 min. The double staining images were visualized with a Zeiss LSM41 0 conforcal laser scan microscope equipped with a Krypton/ Argon laser.
RT -PCR and eDNA sequence. Total RNA from 50 heads obtained from rit flies at ZT18 at 24 OC were reverse-transcripted by Ready-To-Go T -primed First-Strand Kit (Pharmacia). Using four
tim-specific primer sets (tim237-258 and tim1245-1226, tim914-933 and tim1835-1816, timl813- 1834 and tim3404-3383, and tim 3122-3142 and tim4474-4453; these numbers are based on the nucleotide position of the tim eDNA according to Myers et al., 1994), fragments were amplified and cloned into the pCRll vector (Invitorogen). When amplified fragments were too large to be
- p. 34 -