Title
食物繊維ペクチンによる炎症制御機構に関する研究( 本文
(Fulltext) )
Author(s)
石其, 慧太
Report No.(Doctoral
Degree)
博士(農学) 甲第732号
Issue Date
2020-03-13
Type
博士論文
Version
ETD
URL
http://hdl.handle.net/20.500.12099/79374
※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。食物繊維ペクチンによる炎症制御機構に関する研究
2019年
岐阜大学大学院連合農学研究科
生物資源科学
(岐阜大学)
石 其 慧 太
食物繊維ペクチンによる炎症制御機構に関する研究
i 目次 第1章 緒論...1 1-1 食物繊維とその生理機能...1 1-2 ペクチン...2 1-3 腸管免疫...5 1-4 炎症関連疾患について...10 1-5 研究の目的...11 参考文献... ... ... 12 第2章 ペクチンによるエンドトキシンショック緩和機構の解明 ...19 第1節 緒言...19 第2節 実験材料および実験方法...20 2-1 実験材料...20 2 - 2 エ ン ド ト キ シ ン シ ョ ッ ク の 病 態 に ペ ク チ ン が 与 え る 影 響 の 解 析 . . . 2 0 2 - 3 ペ ク チ ン 摂 取 が マ ウ ス パ イ エ ル 板 細 胞 の I L - 6 産 生 に 与 え る 影 響 の 解 析... ... ..29 2-4 磁気ビーズ分離法によるパイエル板細胞の精製 ... ... ....30 2-5 細胞の培養...33 2-6 ペクチンの調製...35 2-7 ペクチンの細胞毒性試験...35 2-8 ペクチンが MΦの炎症性サイトカイン mRNA 発現に及ぼす影響...36 2-9 ペクチンが MΦの IL-6 産生に及ぼす影響...37
ii 2-10 MΦ細胞表面 TLR4 発現量の測定...39 2-11 RAW264.7 細胞に対する蛍光標識 LPS の結合試験...41 2-12 ペクチンの酵素分解...42 2-13 酵素分解ペクチンの細胞添加試験...43 2-14 ポリガラクツロン酸の細胞添加試験...44 2-15 統計処理...45 第3節 結果...46 3-1 ペクチン経口投与は LPS 腹腔投与によって誘導される低体温を軽減す る... ...46 3-2 ペクチン経口投与は小腸 IL-6 産生を抑制するが体温低下の改善には寄与しな い . . . 4 9 3-3 ペクチン経口投与は MLN、パイエル板における炎症性サイトカイン発現を抑制 す る. .. .. .. .. .. .. .. .. .. .. .. . .. .. .. .. .. .. .. .. .. .. .. .. . . . .. .. .. .. .. .. .. .. .. .. .. .. .. . . 51 3 - 4 ペ ク チ ン 経 口 投 与 は パ イ エ ル 板 C D 1 1 c+ 細 胞 の I L - 6 産 生 を 抑 制 す る... ...53 3-5 in vitro においてペクチンは MΦの IL-6 産生を抑制する...55 3-6 ペクチンは TLR-MyD88 経路を抑制する...59 3-7 ペクチン側鎖構造は抗炎症作用に重要である ...63 第4節 考察...66 参考 文献. ... .. .. ... .. .. .. .. .. ... .. .. .. ... .. .. . .. .. .. .. .. ... .. .. .. .. . .. .. .. .... ... .. ... 69 第3章 ペク チン側鎖構 造による大腸腸炎緩和 機構の解明 .... ... .... ... ... .... .. .74
iii 第1節 緒言...74 第2節 実験材料および実験方法... ...75 2-1 実験材料...75 2-2 DSS 大腸炎モデルにペクチン摂取が与える影響の解析 ... ...75 2-3 T NBS 大腸炎モデルにペク チン給餌が与え る影響の解析 ... ....82 2-4 粘膜固有層細胞の分離・精製...84 2-5 T 細胞サブセットのフローサイトメトリー解析...85 2-6 糞中 DNA の抽出・精製...86 2-7 糞中短鎖脂肪酸の測定...88 2-8 抗生剤処理マウスの作成...89 2-9 細胞培養...90 2-10 ペクチンの調製...90 2-11 ペクチンの細胞毒性...90 2-12 ペクチンが M Φの炎症性サイトカイン産生に与え る影響 ...90 2-13 統計処理...90 第3節 結果...91 3-1 オレンジペクチンは DSS 大腸炎を緩和する...91 3-2 オレンジペクチンは TNBS 大腸炎を緩和する...94 3-3 オ レ ンジペク チン は CD 4+ T 細胞サブ セ ッ トの集積に影響する .... ..9 7 3-4 抗生剤投与は腸内発酵を抑制する ... ...99 3-5 抗生剤処理条件下において、オレンジペクチンは TNBS 大腸炎を緩和す る . . . 1 0 1 3-6 抗生剤処理条件下におけるオレンジペクチンおよびシトラスペクチンは CD4+ T 細胞サ ブセ ッ トの集積 に影響する . ... ... .. ... . . .... .. ... ... .. ... ... ... .. .. .... 103
iv 3-7 オレンジペクチンは MΦ様細胞 RAW264.7 の IL-6 産生を抑制する...105 第4節 考察...107 参 考文 献. . .. . .. . .. . .. . ... . . .. . .. . .. . .. . .. . .. . .. . .. .. . . .. . .. . .. . .. . .. . .. . .. . .. .. . . .. . 11 0 第4章 総合考察...115 参考文献... ...120 謝辞...123
v
略語一覧
Ara
arabinose
BSA
bovine serum albumin
CD
Crohn's disease
COX
cyclooxygenase
DAI
disease activity index
DC
dendritic cell
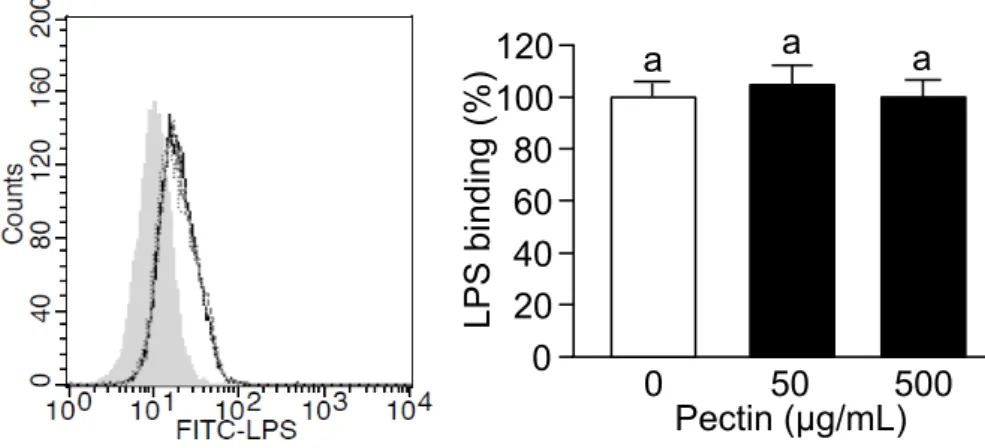
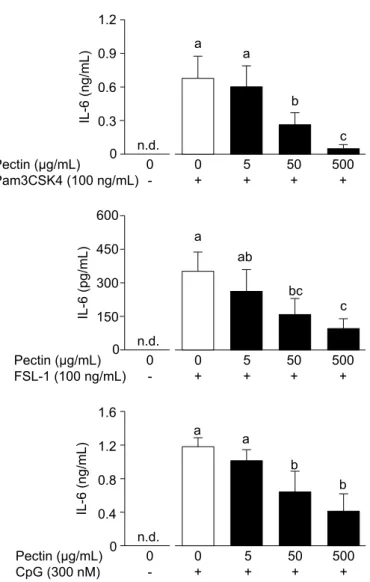
Dha
3-deoxy-lyxo-2-heptulosaric acid
DM
degree of methyl esterification
DMEM Dulbecco’s modification Eagle’s medium
DSS
sodium dextran sulfate
ELISA enzyme-linked immunosorbent assay
FAE
follicle-associated epithelium
FBS
fetal bovine serum
FITC
fluorescein isothiocyanate
Foxp3 forkhead box P3
Gal
galactose
GalA
galacturonic acid
GAPDH glyceraldehyde-3-phosphate dehydrogenase
GPR
G protein-coupled receptor
HDAC histone deacetylases
HG
homogalacturonan
HM
high methoxy
HRP
horseradish peroxidase
vi
IFN
interferon
Ig
immunoglobulin
IL
interleukin
Kdo
3-deoxy-manno-2-octulonic acid
LM
low methoxy
LPS
lipopolysaccharide
MACS magnetic-activated cell sorting
MLN
mesenteric lymph node
MyD88 myeloid differentiation primary response 88
MΦ
macrophage
PAMP pathogen-associated molecular pattern
PE
phycoerythrin
PRR
pathogen recognition receptor
qPCR quantitative polymerase chain reaction
RG
rhamnogalacturonan
Rha
rhamnose
ROR
retinoic-acid-receptor-related orphan receptor
RPMI
Roswell park memorial institute
SCFA short chain fatty acid
SOCS suppressor of cytokine signaling
SHP
Src homology region 2 domain-containing phosphatase
T-bet
T-box family transcription factor
TGF
transforming growth factor
TLR
Toll-like receptor
TNBS 2,4,6-trinitrobenzenesulfonic acid
TNF
tumor necrosis factor
Treg
regulatory T cell
UC
ulcerative colitis
1 第 1 章 緒論 1-1 食物繊維とその生理機能 食物繊維は、ヒトの消化酵素により分解されない食品中の難消化性成分と定義され、主に多 糖類によって構成されている。食物繊維はその消化吸収への耐性から、エネルギー源になら ず不要な食品成分だと考えられてきた。しかし、古くから繊維質の摂取は便量の増加と関連す ることが指摘されていた。Burkitt らはアフリカ原住民と欧米諸国における食習慣と、欧米諸国 で社会問題となっていた心血管疾患、腸疾患、糖尿病、大腸がんの増加について疫学調査 を行った1–3。そこで、欧米におけるこれらの疾患の増加と、食物繊維の摂取不足との関連を指 摘した。以後、急速に食物繊維と健康維持・疾病予防に関連する研究が盛んに行われることと なった。 食物繊維は不溶性食物繊維と水溶性食物繊維に大別される。不溶性食物繊維にはセルロ ース、リグニン、キチンが分類され、水溶性食物繊維にはペクチン、へミセルロースなどが分類 される。一般的にどちらの食物繊維も、水分を含むことで嵩が増加し、便通改善作用をもたら す4。水溶性食物繊維については、消化管上部でのゲル形成に伴い粘度が上昇し、消化管内 容物の移動速度を低下させることで糖吸収や脂質吸収を抑制することが判明している。また、 胆汁酸を吸着・排出することで、再吸収を抑制する。さらに近年では、水溶性食物繊維がヒト に対して有用な腸内細菌の増殖を支持することで健康維持・増進するプレバイオティクス効果 が注目を集めている。 水溶性食物繊維はヒト消化酵素による消化吸収は受けずに、小腸を通過し、大腸に定着し ている腸内細菌によって資化される。このとき、水溶性食物繊維を資化することができる腸内 細菌はBifidobacterium 属や Lactobacillus 属といった一部の種に限定されており、これらの腸 内細菌が優勢となることで、ウェルシュ菌(Clostridium perfringens)や病原性大腸菌といった 毒素産生菌の増殖を抑制し、下痢症、便臭の改善や重篤な腸感染症を予防する 5。さらに、
2
オン酸、酪酸といった短鎖脂肪酸(Short chain fatty acid、SCFA)やコハク酸、乳酸といった有
機酸を生成する 6。これらの腸内細菌代謝産物は大腸上皮細胞に吸収され、エネルギー源と
して利用されることで、大腸上皮の恒常性維持に寄与している 7,8。また、大腸で吸収された代
謝産物の一部は肝臓まで輸送され、エネルギー源や脂肪酸合成に利用される。とりわけ、 SCFA はその受容体として G protein-coupled receptor (GPR) 41、GPR43/FFAR2、GPR109a が 同定されており、これらの受容体を介した抗炎症効果、インスリン抵抗性改善効果が報告され ている。さらに酪酸は細胞内の Histone deacetylases(HDAC)の活性化阻害を示すことで、免 疫応答を制御することが報告されている9–11。 このように、食物繊維の生理機能について、物理・化学的特性による効果と、プレバイオティ クス効果が確立されており、医学や微生物学領域において注目されている。とりわけ、プレバ イオティクス効果についての研究は、DNA シーケンサーの進歩と普及により、2008 年以降か ら急速に増加し、新しい効果が報告されている。 1-2 ペクチン ペクチンは野菜や果実に含まれる主な水溶性食物繊維であり、陸上植物の細胞壁に普遍 的に含まれている。細胞壁中葉において細胞同士の接着に関与し、植物の機械的強度、浸 透圧の維持、外部環境からの防御壁といった役割を担っている 12。また、その分子構造は非 常に複雑であることも知られており、主に Homogalacturonan(HG)、Rhamnogalacturonan I (RG-I)、RG-II と呼ばれる 3 つの構造単位から構成されている(Fig.1-1)。HG は、 α-D-galacturonic acid(GalA)が α-1,4-グリコシド結合した直鎖状の構造であり、ぺクチン分子の 約65%を構成する。HG はさらにメチルエステル化、アセチル化、xylose(Xyl)残基による修飾 を受けている。メチルエステル化はメチルエステル基(-OCH3)修飾されている GalA の割合によ
3
ってメチルエステル化度(degree of methyl esterification、DM)として表わされ、DM が 50%以 上のペクチンを High methoxy(HM)ペクチンと分類し、DM が 50%未満のペクチンを Low methoxy(LM)ペクチンと分類している。HM ペクチンと LM ペクチンでは、溶解性、ゲル形成 能、酵素の耐性に大きな違いがあることが知られている。RG-I は GalA 残基と rhamnose(Rha) 残基の二糖の繰り返し構造([-4-α-D-GalA-1,2-α-L-Rha-1-]n)から成り、ペクチン分子の、 20-35%を構成する。Rha 残基からは主に arabinose(Ara)残基と galactose(Gal)残基で構成さ れる側鎖が分岐している。Ara 残基および Gal 残基はそれぞれ[-5-α-Ara-1,5-α-Ara-1-]n、 [-4-β-Gal-1,4-β-Gal-1-]n といった構造をとる Arabinan、Galactan、Arabinogalactan といった様 式で存在している。Arabinan、Galactan、Arabinogalactan の鎖長や分岐構造は植物種によっ て大きく異なっている。RG-II では HG 構造から Gal、Ara、Rha、apiose(Api)、fucose(Fuc)、 3-deoxy-lyxo-2-heptulosaric acid(Dha)、3-deoxy-manno-2-octulonic acid(Kdo)などから構成
される比較的短い側鎖が分岐していると考えられている。RG-II は植物種によらず、高度に保
存されていることが知られており、植物の成長・発達に必須であることが報告されている 13。HG、 RG-I および RG-II はそれぞれ共有結合しており、RG-I や RG-II が多い領域は毛状領域 (Hairy region)と分類され、HG が多い領域は平滑領域(Smooth region)と分類されている。
Fig.1-1 ペクチン構造の模式図 D
-GalA
L-Rha
D-Gal
RG-I D-Dha
D-Kdo
D-Api
L-Ara
L-Fuc
HG RG-IIO-Methyl
O-Acetyl
4 ペクチンも他の水溶性食物繊維と同様に、血糖上昇抑制作用、血中コレステロール低減作 用14,15が報告されている。興味深いことに、ペクチンは腸管上皮細胞に直接はたらきかけてい る可能性が指摘されている。Tasman-Jones らは、ペクチンを含まない食餌を 12 週間与えたラッ トと10%ペクチンを添加した食餌を同期間与えたラットの空腸および回腸の絨毛の形態を比較 し、ペクチン無給餌群ではそれらの絨毛が規則的な薄い葉状であるのに対して、ペクチン給 餌群の空腸絨毛は不規則で隆起を形成し、回腸絨毛は木の葉形で規則的であることを報告 している 16。また、McCullough らは、通常ラットならびに無菌ラットに対し、成分栄養剤に食物 繊維混合物を 30%添加して与えた場合、腸内細菌の有無にかかわらず小腸での杯細胞の増 加と小腸及び近位結腸において陰窩の分化増殖が認められ、さらに無菌ラットの小腸におい て腸内分泌細胞数が低下することを報告している 17。哺乳類に限らず、ニワトリにおいても、ペ クチン摂取による小腸上皮の形態変化が報告されている15。これまでに当研究室において、こ のペクチンの腸絨毛伸長作用の機序について解析を行ったところ、腸管上皮細胞は細胞外 マトリクスを構成するFibronectin を介してペクチンを認識することを明らかにした18。さらに、ペ クチンを認識した腸管上皮細胞は、細胞外分泌タンパク質である Wingless-type MMTV integration site family, member 3(Wnt3a) を分泌することで増殖上皮細胞を活性化することを 報告している19。 ペクチンの抗炎症効果については、1988 年に Rolandelli らが酢酸誘導性腸炎モデルを用 いてペクチンの経口投与の影響を評価して以来、複数の炎症モデルについてその影響が調 べられてきた 20。これら研究の多くが、ペクチンはプレバイオティクス効果を介して抗炎症効果 を示すことを報告している 21–23。ペクチンはそのほとんどが大腸内の細菌において資化され、 短鎖脂肪酸(SCFA)が生成される。SCFA は上皮細胞に吸収されるだけでなく、粘膜固有層に 存在する、マクロファージ(MΦ)、樹状細胞(DC)、リンパ球に作用することで、炎症応答の起 点となる炎症性サイトカイン、ケミカルメディエーター、ケモカインの産生を抑制する 24–26。さら に血中に移行したSCFA は骨髄における DC の成熟や全身免疫系における抗体産生に影響
5 を及ぼすことで気道アレルギー疾患といった、腸管から遠位部での炎症応答を改善する可能 性が指摘されている22。 興味深いことに、いくつかの先行研究ではペクチンが直接炎症応答を抑制し得る可能性が 指摘されている。in vitro 試験において、Chen らは市販されているシトラス由来ペクチンを MΦ 様 培 養 細 胞 株 RAW264.7 に添加した後に、グラム陰性菌の細胞壁構成成分である Lipopolysaccharide(LPS)で刺激したところ、ケミカルメディエーターである Cyclooxygenase (COX)-2 の産生が抑制されることを報告している 27。Salman らはヒト末梢血単核球にシトラス ペクチンを添加しLPS で刺激したところ、炎症性サイトカインである Interleukin(IL)-1β の産生 が抑制され、抗炎症性サイトカインであるIL-10 の産生が増加することを報告している28。ともに、 これらの効果にはペクチンのメチルエステ化度(DM)が関係していた。さらに Fransen らは水溶 性食物繊維の一種であるフラクタンを無菌マウスに 5 日間投与したところ、小腸における抗体 産生に関連する遺伝子群の発現が亢進していた 29。これらの先行研究より、フラクタンと同様 の水溶性食物繊維であるペクチンも、プレバイオティクス効果と並行して、少なくとも腸管免疫 細胞に作用することで、抗炎症効果を発揮する可能性が考えられた。 1-3 腸管免疫 腸管は栄養成分の吸収を担う吸収上皮細胞の他、粘液を分泌する杯細胞や抗菌物質を分 泌するパネート細胞といった多様な細胞から構成されている。また、腸管には全身の約70%の 免疫細胞が集積しており、全身免疫系とは異なる腸管免疫系を構築している。腸管免疫系で は有害な病原体の侵入に対しては免疫応答を起動する必要があるが、共生細菌や食品成分 といった無害な抗原に対しては不応答(免疫寛容)であることが求められる。このようなニーズ を可能にするために、多様な免疫細胞が複雑なネットワークを構築している。腸管上皮細胞の
6
直下にはCX3CR1+ MΦ が配置されており、樹状突起を伸長し、管腔内の食品抗原や共生細 菌を収集し、抗原として上皮下に転送する30。この抗原をCD103+ DC が受け取り、近傍のリン パ 節 ( 腸 間 膜 リ ン パ 節 ;Mesenteric lymph node ( MLN ) や 孤 立 リ ン パ 節 に 遊 走 す る 。 CD103+DC の多くは Transforming growth factor(TGF)-β と Retinoic acid を高産生し、抗原提 示したCD4+ ナイーブT細胞をCD4+CD25+Fxop3+ regulatory T cell(Treg)へと分化誘導する
31,32。Trge は抗炎症性サイトカインである IL-10、TGF-β や CD25 による IL-2 の枯渇作用を介し
て免疫応答や炎症反応を終息することができる。この Treg のはたらきにより共生細菌や食品
抗原に対する免疫寛容が成立していると考えられている。さらに、一部の CX3R1+ MΦ は
IL-10 を恒常的に産生することで、免疫寛容に寄与している可能性が指摘されている 33,34。一
方で、病原体や、大量の細菌が上皮下に侵入した場合、CX3R1+ MΦ や CD103+ DC は病原 体 に 共 通 す る 分 子 構 造 (Pathogen-associated molecular pattern 、 PAMP ) を Pathogen recognition receptor(PRR)により認識し、IL-1β、IL-6、IL-12、IL-23、Tumor necrosis factor (TNF)-α といったサイトカインを産生することで周辺から免疫細胞を動員する35。さらに、CD4+ ナイーブT 細胞から Th1 と Th17 といったヘルパーT 細胞サブセットを誘導する。Th1 および Th17 はそれぞれ IFN-γ もしくは IL-17A/F を高産生することで、ウィルス、細菌、真菌感染に対 して効果的な免疫応答を惹起する36。 腸管には特徴的な免疫器官として、パイエル板と呼ばれるドーム状の組織が点在している 37。 パイエル板はリンパ球が密に集積したリンパ濾胞と、その上部を覆う特殊な上皮組織である Follicle-associated epithelium (FAE)といった構造からなる。FAE には特殊な上皮細胞 M 細胞 が配置されており、この細胞は細菌、ウィルスやラテックスビーズといった高分子を取り込むこと
ができ、上皮下に輸送するトランスサイトーシスという機能が発達している。M 細胞によって取り
込まれた抗原は、M 細胞の直下に配置されている DC に受け渡され、濾胞領域に存在する T
細胞やB 細胞に提示することで Immunoglobulin(Ig)A の産生を選択的に誘導する38。産生さ
7
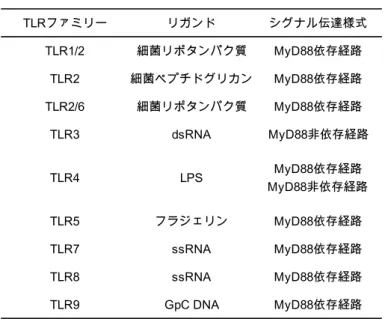
上記にも示した通り、腸管免疫の起動にはMΦ や DC における PRR による抗原の認識が鍵 となる。これまでに、PRR は複数の種類が同定されており、なかでも Toll-like receptor(TLR)フ ァミリーは代表的なPRR であり、ヒトでは TLR1-10、マウスでは TLR1-9、11-13 が同定されてい る。既知のTLR が認識する PAMP については Table 1-1 に示した39。
8
Table 1-1 TLR ファミリーとそのリガンド
TLR は I 型の膜貫通タンパク質であり、細胞外にリガンド認識部位をもち、細胞内には細胞 内にシグナルを伝達する構造をもつ。シグナル伝達の様式としては Myeloid Differentiation Primary Response 88 (MyD88)依存経路と MyD88 非依存経路もしくはその両方によりシグナ ルを細胞内に伝達する(Fig. 1-2)。MyD88 依存経路では PAMPs と結合した TLR が二量体を 形成すると、TLR の細胞質中の Toll/IL-1 receptor (TIR)ドメインが近接し、そこへ MyD88 およ びMyD88-adapter-like (MAL) というアダプター分子が会合する。これらはセリン/スレオニンキ ナーゼIL-1 receptor associated kinase (IRAK) を引き寄せ、このリン酸化およびポリユビキチ ン化を誘導する。ポリユビキチン化したIRAK は MyD88 を離れ TNF receptor associated factor 6 (TRAF6) に結合し、これを活性化する。TRAF6 は TAK1-binding protein (TAB)1、TAB2、 TGF-β activated kinase 1 (TAK1)を介して Inhibitor of NF-κB kinase (IKK) を活性化し、これ が転写因子Nuclear factor-kappa B (NF-κB) の核内移行を抑制している Inhibitor of NF-κB (IκB) の分解を誘導する。IκB の抑制がなくなった NF-κB は核内へと移行し、IL-1β、IL-6、 IL-12p40 および TNF-α の転写を誘導する。一方で TLR3 と TLR4 では MyD88 非依存経路 でシグナルを伝達する。ウィルスに特有の PAMPs である dsRNA をエンドソームで認識した TLRファミリー リガンド シグナル伝達様式 TLR1/2 細菌リポタンパク質 MyD88依存経路 TLR2 細菌ペプチドグリカン MyD88依存経路 TLR2/6 細菌リポタンパク質 MyD88依存経路 TLR3 dsRNA MyD88非依存経路 TLR4 LPS MyD88依存経路 MyD88非依存経路 TLR5 フラジェリン MyD88依存経路 TLR7 ssRNA MyD88依存経路 TLR8 ssRNA MyD88依存経路 TLR9 GpC DNA MyD88依存経路
9
TLR3 では、アダプター分子 TIR-domain-containing adapter-inducing interferon-β (TRIF) を TLR3 の TIR ドメインに動員する。TRIF は TNF receptor associated factor 3 (TRAF3)、 TANK-binding kinase 1 (TBK1) および IKKε を介して転写因子 interferon regulatory transcription factor 3 (IRF-3) をリン酸化し、リン酸化 IRF-3 の核内移行を誘導する。IRF-3 は 抗ウィルス作用を備えるサイトカインであるIFN-β の転写を強力に誘導する。 TLR は自然免疫・獲得免疫の起動に重要である一方で、近年では自己免疫疾患や慢性炎 症疾患の引き金となることが指摘されている。 Fig.1-2 TLR シグナル伝達様式 IRAK IRAK IRAK TRAF6 TAB3 TAB2 TAK1 IKKβ IKKα NF-κB NF-κB JNK p38 AP-1 CREBP AP-1 CREBP IRAK IRAK IRAK TRAF6 TRAF3 IKKα IFR7 IFR7 TRAF3 IKKα TAB2 IFR3 IFR3 TLR5 TLR1/2 TLR2/6 TLR4 TLR4 TLR3 TLR9 TLR7 Endosome Endocytosis IL-1β,IL-6,
IL-12p40, TNF-α IFN-α, IFN-β
TRIF MyD88 MAL TIR domain Nuclear Cytoplasm
10 1-4 炎症関連疾患について 生体防御の観点から、免疫システムは極めて重要なシステムのひとつである。しかし、免疫 細胞の制御異常による過剰な炎症応答は宿主を傷害することで、自己免疫疾患や慢性炎症 疾患といった難治性疾患の原因となり得る。これらの疾患は、生命の危機や Quality of life (QOL)の低下をもたらす。さらに、生活習慣病にまとめられる 2 型糖尿病、肥満、心血管疾患 や、がんといった疾患の基盤業態としても慢性炎症が関与していることが近年指摘されている 40。免疫細胞のなかでも MΦ は、TLR を介して炎症惹起を担うことからも、全身性もしくは局所 における炎症関連疾患の病態形成に対して密接に関与していることが指摘されている。 全身性炎症応答として、敗血症などの感染症が進行することで急激な血圧・体温の低下と いったショック症状が観察されるエンドトキシンショックが挙げられる。上皮バリアの破綻などに より、体内に病原体が大量に出現すると MΦ を中心とした免疫細胞がこれを認識し、大量の IL-1β、IL-6、IL-12、TNF-α を産生する 41。このとき産生された炎症性サイトカインによって、周 囲の細胞の活性化が促され、サイトカインストームと呼ばれる炎症性サイトカインやケミカルメ ディエーターの正のフィードバックループが形成される。産生された過剰量の炎症性サイトカイ ンやメディエーターにより、血管内皮の透過性亢進や臓器障害が引き起こされる。敗血症およ びエンドトキシンショックは急性期の疾患であり、死亡率は 30~80%と非常に高い。また、罹患 者数も年間約3000 万人と多く、これは先進国と途上国で罹患率に大きな差はない42,43。エンド トキシンショックの動物モデルである盲腸結紮・穿刺法 (Cecal ligation puncture、CLP) による 腹膜炎モデルおよびLPS 腹腔投与による LPS 血症モデルにおいて、TLR4 の欠損により症状 が改善することが報告されている44,45。
腸管における炎症疾患としては、潰瘍性大腸炎(Ulcerative Colitis、UC)と、クローン病 (Crohn's Disease、CD)が主に挙げられ、これらは炎症性腸疾患(Inflammatory bowel disease、 IBD)と総称されている。UC では、炎症は直腸の粘膜表層から始まり、連続的に結腸に向かっ
11 能性があり、また炎症が粘膜下層よりも深部にまで広がることが特徴的である。どちらの疾患も 下痢、血便、体重減少、発熱といった全身性の症状が現れ、再燃と寛解を繰り返すことで、炎 症が慢性化する。本邦においては、IBD は厚生労働省により特定難病に指定されており、約 20 万人が罹患している。また、近年の疫学調査によると、欧米においては 320 万人が罹患して おり、くわえて、アフリカ、アジア、南米において工業化が進展している国々においても罹患者 が増加しており、世界的な社会問題となっている46–50。IBD の正確な病因は未だ明らかにされ ておらず、遺伝的要因、環境、共生菌の影響、免疫学的要因といった多数の要素が組み合わ せることで病態が形成されている可能性が指摘されている。とりわけ免疫細胞の異常な活性化 は病態形成に重要であることが認識されており、エフェクター T 細胞の異常な活性化が IBD の病態に深く関与している 51,52。また、マウス腸炎モデルの解析より、エフェクターT 細胞の活 性化について腸管MΦの TLR4 の活性化と、その後の IL-1β および IL-6 産生が病態形成を 担っていることが指摘されている34,53。 1-5 研究の目的 これまでの先行研究より、ペクチンは上皮細胞に直接作用することで、その機能を変化させ ることから、また in vitro において免疫細胞の活性化を制御することから、ペクチンはプレバイ オティクス効果とは独立して、免疫細胞に直接作用することで、炎症疾患を緩和していることが 予測された。本研究ではこの可能性を検証し、ペクチンがどのような細胞に作用することで、抗 炎症効果を示すのかを明らかにすることを目指した。このようなペクチンの新規な機能を見出 すことは、慢性炎症の予防・治療に対して積極的にペクチンのような水溶性食物繊維を利用 する論拠を与えると考えた。そこで、第2章では全身性炎症応答としてマウスエンドトキシンショ ックモデルにペクチン給餌が与える影響を評価した。また,第3 章ではペクチンが腸炎モデル のような局所炎症に与える影響を評価した。
12 参考文献
1. Burkitt DP. Related disease--related cause? Lancet (London, England). 1969;2(7632):1229-1231.
2. Burkitt DP. Epidemiology of cancer of the colon and rectum. Cancer. 1971;28(1):3-13. 3. Burkitt DP, Walker AR and Painter NS. Dietary fiber and disease. J Am Med Assoc.
1974;229(8):1068-1074.
4. Ebihara K. Studies on Nutritional and Physiological Effects of Dietary Fiber. Nippon
Eiyo Shokuryo Gakkaishi. 2008;61(1):3-9.
5. Slavin J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients. 2013;5(4):1417-1435.
6. Watabe J. Carbohydrate Fermentation in the Colon. J Intest Microbiol. 2005;19(3):169-177.
7. Hara H. Physiological Effects of Short-Chain Fatty Acid Produced from Prebiotics in the Colon. J Intest Microbiol. 2002;16(1):35-42.
8. den Besten G, van Eunen K, Groen AK, Venema K, Reijngoud D-J and Bakker BM. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J Lipid Res. 2013;54(9):2325-2340.
9. Maslowski KM and Mackay CR. Diet, gut microbiota and immune responses. Nat
Immunol. 2011;12(1):5-9.
10. Furusawa Y, Obata Y, Fukuda S, Endo T a, Nakato G, Takahashi D, Nakanishi Y, Uetake C, Kato K, Kato T, Takahashi M, Fukuda NN, Murakami S, Miyauchi E, Hino S, Atarashi K, Onawa S, Fujimura Y, Lockett T, Clarke JM, Topping DL, Tomita M, Hori S, Ohara O, Morita T, Koseki H, Kikuchi J, Honda K, Hase K and Ohno H. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature.
13 2013;504(7480):446-450.
11. Park J, Kim M, Kang SG, Jannasch AH, Cooper B, Patterson J and Kim CH. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone
deacetylases and regulation of the mTOR-S6K pathway. Mucosal Immunol. 2015;8(1):80-93.
12. Caffall KH and Mohnen D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr Res. 2009;344(14):1879-1900.
13. Funakawa H and Miwa K. Synthesis of borate cross-linked rhamnogalacturonan II.
Front Plant Sci. 2015;6(APR):223.
14. Jenkins DJ, Newton C, Leeds AR and Cummings JH. Effect of pectin, guar gum, and wheat fibre on serum-cholesterol. Lancet (London, England). 1975;1(7916):1116-1117. 15. Langhout DJ, Schutte JB, Van Leeuwen P, Wiebenga J and Tamminga S. Effect of
dietary high- and low-methylated citrus pectin on the activity of the ileal microflora and morphology of the small intestinal wall of broiler chicks. Br Poult Sci.
1999;40(3):340-347.
16. Tasman-Jones C, Owen RL and Jones AL. Semipurified dietary fiber and small-bowel morphology in rats. Dig Dis Sci. 1982;27(6):519-524.
17. McCullough JS, Ratcliffe B, Mandir N, Carr KE and Goodlad RA. Dietary fibre and intestinal microflora: Effects on intestinal morphometry and crypt branching. Gut. 1998;42(6):799-806.
18. Nishida M, Murata K, Kanamaru Y and Yabe T. Pectin of Prunus domestica L. alters sulfated structure of cell-surface heparan sulfate in differentiated Caco-2 cells through stimulation of heparan sulfate 6-O-endosulfatase-2. Biosci Biotechnol Biochem. 2014;78(4):635-643.
14
19. Nishida M, Murata K, Oshima K, Itoh C, Kitaguchi K, Kanamaru Y and Yabe T. Pectin from Prunus domestica L. induces proliferation of IEC-6 cells through the alteration of cell-surface heparan sulfate on differentiated Caco-2 cells in co-culture. Glycoconj J. 2015;32(3-4):153-159.
20. Rolandelli RH, Saul SH, Settle RG, Jacobs DO, Trerotola SO and Rombeau JL.
Comparison of parenteral nutrition and enteral feeding with pectin in experimental colitis in the rat. Am J Clin Nutr. 1988;47(4):715-721.
21. Andoh A, Bamba T and Sasaki M. Physiological and anti-inflammatory roles of dietary fiber and butyrate in intestinal functions. JPEN J Parenter Enteral Nutr. 1999;23(5 Suppl):S70-3.
22. Trompette A, Gollwitzer ES, Yadava K, Sichelstiel AK, Sprenger N, Ngom-Bru C, Blanchard C, Junt T, Nicod LP, Harris NL and Marsland BJ. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat Med.
2014;20(2):159-166.
23. Sherry CL, Kim SS, Dilger RN, Bauer LL, Moon ML, Tapping RI, Fahey GC, Tappenden KA and Freund GG. Sickness behavior induced by endotoxin can be mitigated by the dietary soluble fiber, pectin, through up-regulation of IL-4 and Th2 polarization. Brain Behav Immun. 2010;24(4):631-640.
24. Maslowski KM, Vieira AT, Ng A, Kranich J, Sierro F, Yu D, Schilter HC, Rolph MS, Mackay F, Artis D, Xavier RJ, Teixeira MM and Mackay CR. Regulation of
inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature. 2009;461(7268):1282-1286.
25. Sina C, Gavrilova O, Förster M, Till A, Derer S, Hildebrand F, Raabe B, Chalaris A, Scheller J, Rehmann A, Franke A, Ott S, Häsler R, Nikolaus S, Fölsch UR, Rose-John S,
15
Jiang H-P, Li J, Schreiber S and Rosenstiel P. G protein-coupled receptor 43 is essential for neutrophil recruitment during intestinal inflammation. J Immunol.
2009;183(11):7514-7522.
26. Cox MA, Jackson J, Stanton M, Rojas-Triana A, Bober L, Laverty M, Yang X, Zhu F, Liu J, Wang S, Monsma F, Vassileva G, Maguire M, Gustafson E, Bayne M, Chou C-C, Lundell D and Jenh C-H. Short-chain fatty acids act as antiinflammatory mediators by regulating prostaglandin E(2) and cytokines. World J Gastroenterol.
2009;15(44):5549-5557.
27. Chen C-H, Sheu M-T, Chen T-F, Wang Y-C, Hou W-C, Liu D-Z, Chung T-C and Liang Y-C. Suppression of endotoxin-induced proinflammatory responses by citrus pectin through blocking LPS signaling pathways. Biochem Pharmacol. 2006;72(8):1001-1009. 28. Salman H, Bergman M, Djaldetti M, Orlin J and Bessler H. Citrus pectin affects
cytokine production by human peripheral blood mononuclear cells. Biomed
Pharmacother. 2008;62(9):579-582.
29. Fransen F, Sahasrabudhe NM, Elderman M, Bosveld M, El Aidy S, Hugenholtz F, Borghuis T, Kousemaker B, Winkel S, van der Gaast-de Jongh C, de Jonge MI, Boekschoten MV, Smidt H, Schols HA and de Vos P. β2→1-Fructans Modulate the Immune System In Vivo in a Microbiota-Dependent and -Independent Fashion. Front
Immunol.
30. Mazzini E, Massimiliano L, Penna G and Rescigno M. Oral tolerance can be established via gap junction transfer of fed antigens from CX3CR1+ macrophages to CD103+
dendritic cells. Immunity. 2014;40(2):248-261.
31. Coombes JL, Siddiqui KRR, Arancibia-Cárcamo C V, Hall J, Sun C, Belkaid Y and Powrie F. A functionally specialized population of mucosal CD103+ DCs induces
16
Foxp3+ regulatory T cells via a TGF-beta and retinoic acid-dependent mechanism. J Exp
Med. 2007;204(8):1757-1764.
32. Schulz O, Jaensson E, Persson EK, Liu X, Worbs T, Agace WW and Pabst O. Intestinal CD103+, but not CX3CR1+, antigen sampling cells migrate in lymph and serve classical dendritic cell functions. J Exp Med. 2009;206(13):3101-3114.
33. Takada Y, Hisamatsu T, Kamada N, Kitazume MT, Honda H, Oshima Y, Saito R, Takayama T, Kobayashi T, Chinen H, Mikami Y, Kanai T, Okamoto S and Hibi T. Monocyte chemoattractant protein-1 contributes to gut homeostasis and intestinal inflammation by composition of IL-10–producing regulatory macrophage subset. J
Immunol. 2010;184(5):2671-2676.
34. Medina-Contreras O, Geem D, Laur O, Williams IR, Lira SA, Nusrat A, Parkos CA and Denning TL. CX3CR1 regulates intestinal macrophage homeostasis, bacterial
translocation, and colitogenic Th17 responses in mice. J Clin Invest. 2011;121(12):4787-4795.
35. Saleh M and Trinchieri G. Innate immune mechanisms of colitis and colitis-associated colorectal cancer. Nat Rev Immunol. 2011;11(1):9-20.
36. Maynard CL and Weaver CT. Intestinal effector T cells in health and disease. Immunity. 2009;31(3):389-400.
37. Heel K a, McCauley RD, Papadimitriou JM and Hall JC. Review: Peyer’s patches. J
Gastroenterol Hepatol. 1997;12(October 1996):122-136.
38. Sato A, Hashiguchi M, Toda E, Iwasaki A, Hachimura S and Kaminogawa S. CD11b+ Peyer’s patch dendritic cells secrete IL-6 and induce IgA secretion from naive B cells. J
Immunol. 2003;171(7):3684-3690.
17
innate immunity. Nat Rev Immunol. 2013;13(6):453-460.
40. Franceschi C and Campisi J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J Gerontol A Biol Sci Med Sci. 2014;69 Suppl 1:S4-9.
41. Schulte W, Bernhagen J and Bucala R. Cytokines in sepsis: potent immunoregulators and potential therapeutic targets--an updated view. Mediators Inflamm.
2013;2013:165974.
42. Adhikari NKJ, Fowler RA, Bhagwanjee S and Rubenfeld GD. Critical care and the global burden of critical illness in adults. Lancet. 2010;376(9749):1339-1346.
43. Vincent J-L, Sakr Y, Sprung CL, Ranieri VM, Reinhart K, Gerlach H, Moreno R, Carlet J, Le Gall J-R and Payen D. Sepsis in European intensive care units: results of the SOAP study. Crit Care Med. 2006;34(2):344-353.
44. Takeuchi O, Hoshino K, Kawai T, Sanjo H, Takada H, Ogawa T, Takeda K and Akira S. Differential roles of TLR2 and TLR4 in recognition of gram-negative and gram-positive bacterial cell wall components. Immunity. 1999;11(4):443-451.
45. Kawai T, Adachi O, Ogawa T, Takeda K and Akira S. Unresponsiveness of MyD88-deficient mice to endotoxin. Immunity. 1999;11(1):115-122.
46. Burisch J, Jess T, Martinato M and Lakatos PL.The burden of inflammatory bowel disease in Europe. J Crohns Colitis. 2013;7(4):322-337.
47. Kappelman MD, Rifas-Shiman SL, Porter CQ, Ollendorf DA, Sandler RS, Galanko JA and Finkelstein JA. Direct Health Care Costs of Crohn’s Disease and Ulcerative Colitis in US Children and Adults. Gastroenterology. 2008;135(6):1907-1913.
48. Kappelman MD, Moore KR, Allen JK and Cook SF. Recent trends in the prevalence of Crohn’s disease and ulcerative colitis in a commercially insured US population. Dig Dis
18
Sci. 2013;58(2):519-525.
49. Rocchi A, Benchimol EI, Bernstein CN, Bitton A, Feagan B, Panaccione R, Glasgow KW, Fernandes A and Ghosh S. Inflammatory bowel disease: a Canadian burden of illness review. Can J Gastroenterol. 2012;26(11):811-817.
50. Ng SC, Shi HY, Hamidi N, Underwood FE, Tang W, Benchimol EI, Panaccione R, Ghosh S, Wu JCY, Chan FKL, Sung JJY and Kaplan GG. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: a systematic review of population-based studies. Lancet. 2017;390(10114):2769-2778.
51. Powrie F, Leach MW, Mauze S, Menon S, Caddle LB and Coffman RL. Inhibition of Th1 responses prevents inflammatory bowel disease in scid mice reconstituted with CD45RBhi CD4+ T cells. Immunity. 1994;1(7):553-562.
52. Fina D, Sarra M, Fantini MC, Rizzo A, Caruso R, Caprioli F, Stolfi C, Cardolini I, Dottori M, Boirivant M, Pallone F, Macdonald TT and Monteleone G. Regulation of gut inflammation and th17 cell response by interleukin-21. Gastroenterology.
2008;134(4):1038-1048.
53. Kayama H, Kohyama M, Okuzaki D, Motooka D, Barman S, Okumura R, Muneta M, Hoshino K, Sasaki I, Ise W, Matsuno H, Nishimura J, Kurosaki T, Nakamura S, Arase H, Kaisho T and Takeda K. Heme ameliorates dextran sodium sulfate-induced colitis
through providing intestinal macrophages with noninflammatory profiles. Proc Natl
19 第 2 章 ペクチンによるエンドトキシンショック緩和機構の解明 第 1 節 緒言 ペクチンは、プレバイオティクス効果による抗炎症効果が報告されている一方で、免疫細胞 に直接作用することで、炎症応答を抑制する可能性が指摘されている。in vitro 試験では、ヒト 単 球 や マ ク ロ フ ァ ー ジ (MΦ ) 様 培 養 細 胞 に お い て 、 LPS 刺 激 に よ っ て 誘 導 さ れ る Cyclooxygenase-2、IL-1β の産生がペクチン添加によって抑制されることが報告されている1,2。 Popov らは、ペクチン単回投与が全身性炎症応答であるエンドトキシンショックからマウスを保 護することを報告している3。これらの研究ではペクチンのDM がその生理活性と相関している ことから、ペクチン化学構造には薬理学的な効果が備わっていることが示唆された。一方で、 ペクチンがエンドトキシンショックを緩和する詳細な機構は未だ明らかにされていない。 本章では、ペクチンによるエンドトキシンショック緩和機構を明らかにするために、ペクチン 給餌がマウスエンドトキシンショックモデルに与える影響を評価した。この時、パイエル板、 MLN、脾臓の遺伝子発現量を測定することで、どのような組織にペクチンが作用しているのか を検討した。さらに、組織から細胞をソーティングすることで、どのような細胞にペクチンが作用 しているのかを調査した。最後に、ペクチンが炎症応答を抑制する機構と、どのような構造が、 抗炎症効果に重要であるのかをin vitro 試験により検討した。
20 第 2 節 実験材料および実験方法 2-1 実験材料 ・実験動物 実験には日本SLC 株式会社(静岡)から購入した 7~12 週齢の雄性 C57BL/6 マウスを使用 した。マウスは12 時間周期で明暗が制御されている飼育室において、気温 23±2℃で E-2(日 本クレア株式会社、東京)を給餌することで飼育した。動物実験に関しては、岐阜大学の動物 実験委員会へ申請書を提出し、承認を得た後、法規に従い適切に実験を実施した。また、動 物実験に関しては、動物愛護上の理念から、苦痛に対して充分に配慮して実験を行った。 ・マウスMΦ 様細胞株 RAW264.7
RAW264.7 は ATCC(Manassas、VA)から購入した。RAW264.7 は、Mus musculus (mouse) の腹水から樹立された株化細胞で、CD11b および F4/80 を発現し、単球および MΦ 様の形態 を示す。 ・シトラスペクチン 本章ではキシダ化学(大阪)から購入したシトラスペクチンを使用した。 試薬は、特に記述のない限り和光純薬工業株式会社(大阪)より購入したものを使用した。 2-2 エンドトキシンショックの病態にペクチンが与える影響の解析 2-2-1 試薬の調製 ・PBS
KH2PO4 1 g、 NaCl 40 g、 Na2HPO4・12H2O 14.5 g、KCl 1 g を Milli Q 水に溶解し、500
mL に定容して 10×PBS を調製した。これを 10 倍希釈することで、1×PBS を調製した。121˚C、 20 分間オートクレーブしてから使用した。
21
LPS from E. coli O111 B4(Sigma-Aldrich、St. Louis、MO)は 1 mg を Milli Q 水 1 mL に溶 解し、50 µLずつ小分けに分注した後、-20°Cで保存した。使用時は1 mg/mL LPS溶液をPBS で希釈し、100 µg/mL LPS 溶液となるように調製して使用した。 ・10 mg/mL ペクチン水溶液 シトラスペクチン(キシダ化学、大阪)を200~300 mg 量りとり、10 mg/mL となるように RO 水 を加え、完全に溶解するまで加熱した。 2-2-2 マウスへのペクチン給餌 マウスを1 週間予備飼育した後、ペクチン水溶液を 10 日間自由摂取させた。コントロール群 には、溶媒であるRO 水を用意した。ペクチン水溶液は用事調製し、毎日交換した。 2-2-3 エンドトキシンモデルの構築 ペクチン水溶液の給餌 10 日目に、26G の注射針(テルモ、東京)を用いて、LPS 溶液を腹 腔投与した(5 mg/kg)。 2-2-4 採血、直腸温測定および解剖 LPS 投与後の直腸温を 0、2、4 時間ごとに測定した。採血は LPS 投与 2 時間後および 4 時 間後に尾採血および心採血により 200~500 µL 程度採血した。頸椎脱臼により安楽死させた 後、LPS 投与 2 時間後および 4 時間後にそれぞれ開腹し、脾臓、MLN およびパイエル板を 採取した。採取した臓器は液体窒素を用いて凍結し、タンパク質とTotal RNA を抽出するため に-80˚C で保存した。 2-2-5 血清炎症性サイトカインの濃度測定
22
assay (ELISA)を行った。ELISA には DuoSet ELISA development system(R&D systems、 Minneapolis、MN)を用いた。 ・PBS 第2 章 2-2-1 と同様に調製した。 ・Wash buffer PBS に Tween 20 (ナカライテスク、京都)を加え、0.05 % Tween 20 溶液とした。 ・Reagent diluent
PBS に Bovine serum albumin (BSA) (純度 98 %≤、ナカライテスク)を加え、1 % BSA 溶 液とした。
・anti-mouse IL-1β、IL-6、TNF-α capture antibody
rat anti-mouse IL-1β、rat anti-mouse IL-6、goat anti-mouse TNF-α capture antibody(R&D systems)は Kit の指示に従い、500 μL の PBS に再溶解した。使用時は PBS で 100~130 倍 希釈して使用した。
・mouse IL-1β、IL-6、TNF-α standard
付属のmouse IL-1β、IL-6、IL-17A、TNF-α standard は Kit の指示に従い、200~500 μL の Reagent diluent に再溶解した。使用時は Kit の指示に従い、Reagent diluent で一度 100~200
倍希釈した後に2 倍公比で段階希釈を行い、8 点で検量線を作成した。
・anti-mouse IL-1β、IL-6、TNF-α detection antibody
biotinylated goat anti-mouse IL-1β、biotinylated goat anti-Mouse IL-6、biotinylated goat anti-mouse TNF-α detection antibody(R&D systems)は Kit の指示に従い、1000 μL の Reagent diluent に再溶解した。使用時は Reagent diluent で 60 倍希釈して使用した。
・Streptavidin-Horseradish peroxidase (HRP)
DuoSet ELISA development system に付属の 2.0 mL Streptavidin conjugated HRP を Reagent diluent で 40 倍希釈して使用した。
23 ・基質溶液
ELISA POD 基質 TMB キット(ナカライテスク)を使用した。Tetramethylbenzidine(TMB)を主 成分とする発色液と、H2O2溶液をあらかじめ室温に戻し、体積比1:1 で混合して使用した。 ・反応停止液 濃硫酸(98 % ≤ 、ナカライテスク)を Milli Q 水で希釈し、1 M 硫酸溶液を調製して使用し た。 ・サンプル溶液 調製した血清はReagent buffer によりそれぞれ 3~100 倍希釈した。 ・血清の分離 全血は4˚C で一晩静置した。その後、3000 × g で 15 分間遠心分離を行った。下層の血餅 に注意して上層の血清を回収した。回収量が不十分な場合は 5000~10000 × g で再遠心し た。 ・固相化プレートの調製
希釈したmouse IL-1β capture antibody、mouse IL-6 capture antibody、mouse TNF-α capture antibody を Costar assay plate (Corning、New York、NY)に 25 µL ずつ加え、ラップで密閉し、 24 時間室温で静置した。Capture antibody をアスピレーターで除去し、Wash buffer を 100 µL 加えた後に吸引除去する操作を3 回繰り返した。3 回目の後、Wash buffer を完全に除くため に、プレートをペーパータオルに押し当て、水切りを行った。プレートへの非特異的な抗原の 結合を防ぐために、Reagent buffer を 100 µL 加え、ラップで密閉し、1 時間室温で静置した。そ の後、Wash buffer で 3 回洗浄した。
・ELISA
事前に調製したサンプル溶液とmouse IL-1β standard、mouse IL-6 standard、mouse TNF-α standard を 25 µL ずつ固相化プレートに加え、ラップで密封し、2 時間室温で静置した。2 時間 後、プレート上の溶液を除去し、Wash buffer で 3 回洗浄した。希釈した mouse IL-1β detection
24
antibody、mouse IL-6 detection antibody、mouse TNF-α detection antibody を 25 µL ずつ固相
化プレートに加え、ラップで密封し、2 時間室温で静置した。2 時間後、プレート上の溶液を除
去し、Wash buffer で 3 回洗浄した。希釈した Streptavidin-HRP を 25 µL ずつ固相化プレート に加え、ラップで密封し、20 分間室温で静置した。20 分後、プレート上の溶液を除去し、Wash buffer で 3 回洗浄した。基質溶液を 50 µL ずつ固相化プレートに加え、ラップで密封し、20 分 間室温で静置した。反応停止液を50 µL ずつ固相化プレートに加え、450 nm における吸光度 を測定した。 2-2-6 小腸組織のホモジナイズ ・Lysis buffer 1×PBS に対してプロテアーゼ阻害剤カクテル(ナカライ)を 1/100 倍量加え、4˚C で保存し た。 第 2 章 2-2-4 で採取した小腸重量を測定した(約 600~800 mg)。試験管に小腸を移し、 Lysis buffer を 200 mg/mL となるように加えた。アイリス剪刃を用いて組織を 2~4 mm 程度に 細断した。ポリトロンホモジナイザ―を用いて氷冷下でホモジネートを調製した。ホモジネート を15 mL 遠心管に移し 10 分間、4˚C、2000 × g で遠心した。上清を 1.0 mL ずつ 1.5 mL チ ューブに移し、10 分間、4˚C、10000 × g で遠心した。上清を 0.9 mL とり、新しい 1.5 mL チュ ーブに移し、-20℃で保存した。 2-2-7 小腸組織中炎症性サイトカインの測定 第2 章 2-2-5 と同様に、組織上清中の IL-1β、IL-6、TNF-αはサンドイッチ ELISA により測 定した。測定にはDuoSet ELISA development system を使用した。
25 2-2-8 BCA 法によるタンパク質濃度の測定
小腸ホモジナイズ中の炎症性サイトカイン量は総タンパク質量で補正した。総タンパク質量
はBCA Protein Assay Kit(Thermo Fisher Scientific Inc.、Waltham、MA)を購入し使用した。 Kit に同封された BSA スタンダードまたはサンプル 25 μL を 96 well マイクロプレートに加えた。 その後、Protein Assay Reagent A 液と B 液を 50:1 で混合した Working Reagent 200 μL を 加えて、37℃ で 30 分間インキュベートした。その後、540 nm の吸光度を測定した。 2-2-9 各臓器における炎症性サイトカイン mRNA 発現量の解析 ・DEPC 水 ドラフト内で、Milli Q 水 400 mL に DEPC(ナカライテスク)400 µL を加え、混合させた。こ れを一晩静置させた後、オートクレーブを2 回行った。 ・70%エタノール 99 %エタノールに DEPC 水を加えて、70%に希釈した。 ・3 M 酢酸ナトリウム 酢酸ナトリウムとDEPC(0.1%)を Milli Q 水に溶解させ、2 回オートクレーブした。 ・1×TAE
Tris 121 g、酢酸 28.55 mL、EDTA·2Na 3.715 g を Milli Q 水に溶解して、500 mL に定容し た。これをオートクレーブして、50×TAE を調製した。これを Milli Q 水で 50 倍希釈して 1× TAE を調製した。 ・Total RNA の抽出 Total RNA 抽出の操作は、クリーンベンチ内で行なった。クリーンベンチ内は、あらかじめ UV ランプを 30 分程度照射し、RNase Quiet(ナカライテスク)とエタノールでよく拭いた。 -80˚C で凍結保存した動物組織サンプル約 20~50 µg を速やかに 1.5 mL チューブに移し、 Total RNA 抽出試薬 RNAiso Plus(タカラバイオ、滋賀)を 500 µL 加え、ディスポーザブルホモ
26 ジナイザ―(ニッピ、東京)を用いて組織をホモジナイズした。この溶液に RNAiso Plus を 500 µL 加え、全体を 1000 µL とした。5 分間静置した後、12000 × g、5 分間、4℃で遠心した。沈殿 を採取しないように注意して上清を回収し、新しい1.5 mLチューブに移した。この溶液にクロロ ホルム(ナカライテスク)を200 µL(開始容量の 0.2 倍量)加え、乳白状になるまでよく振りまぜ、 室温で5分間静置した。次に、12000 × g、4˚C、15分間で遠心し、分離した上層(RNA を含む) を、中間層(ゲノムDNA を含む)に触れないよう十分に注意して採取し、新しい 1.5 mL チュー ブに移した。2-プロパノール(ナカライテスク)を 1000 µL(開始容量と等量)加えてよく混合し、 室温で10 分間静置した。12000 × g、4˚C、10 分間で遠心し、Total RNA を沈殿させた。上清を 捨て、70% 冷エタノールを 1000 µL(開始容量と等量)加え、よく撹拌し沈殿物を洗浄した。 7500 × g、 5 分間、4˚C で遠心し、上清を捨てた。室温で Total RNA の沈殿を数分間乾燥させ、 DEPC 水 42.5 µL を加えて沈殿を溶解した。 ・Total RNA の精製
Total RNA 溶液にゲノム DNA が混入する可能性を除くために、Recombinant DNase I(タカラ バイオ)処理を行った。Total RNA 溶液 42.5 µL に対して、Table 2-1 のように調製して、37˚C で 30 分間インキュベートした。
Table 2-1 DNaseI 反応液の組成
0.5 M EDTA を 2.5 µL 加え、80˚C、2 分間インキュベートして DNase I を失活させた。DEPC 水を加え溶液を100 µL とし、3 M 酢酸ナトリウム 10 µL と冷エタノール 250 µL を加え、-80˚C で20 分間静置し、12000 × g、4˚C、10 分間で遠心し、Total RNA を沈殿させた。次に、上清を 捨て、70%エタノール 1000 µL を加え、沈殿を洗浄し、12000 × g、4˚C、5 分間遠心した。上清 Total RNA 溶液 42.5 µL 10 × DNase 1 Buffer 5.0 µL Recombinant DNase I 2.0 µL RNase Inhibitor(40 U/mL,東洋紡,大阪) 0.5 µL
27
を捨て、Total RNA の沈殿を数分間乾燥した。DEPC 水 10~40 µL を加え、沈殿を溶解させ、 Total RNA 溶液の濃度および純度を測定した。
・Total RNA の純度測定
Total RNA 溶液 1 µL を用いて、260 nm および 280 nm における吸光度を測定した。その吸 光度比(A260/A280)から、RNA の純度を測定した。
・Total RNA の濃度測定
Total RNA 溶液 1 µL を用いて、260 nm における吸光度を測定した。RNA は A260=40
µg/mL という特徴を持つことより、得られた吸光度から RNA 濃度を測定した。 ・cDNA の合成
1.5 mL チューブに Table 2-2 に示した通りの反応溶液を調製した。cDNA 合成に使用する Total RNA 量は 1 µg としたため、DEPC 水と Total RNA の合計量が 4.5 µL となるようにした。
Table 2-2 逆転写反応液の組成
調製した反応液を 42˚C、30 分間インキュベートし、次に 99˚C で 5 分間インキュベートし、 ReverTra Ace を失活させた。この後、DEPC 水 20 µL を加え、容量を 40 µL とし、-20 で保存し た。
・Real-time quantitative PCR
Thermal Cycler Dice Real Time System (タカラバイオ)を使用し、Quantitative polymerase chain reaction(qPCR)を行った。測定には THUNDERBIRD SYBR qPCR Mix (東洋紡)を使
1 µg Total RNA 溶液 4.5 µL 5 × RT Buffer 4.0 µL ReverTra Ace (100 U/µL,東洋紡) 1.0 µL RNase Inhibitor(40 U/µL,東洋紡) 0.5 µL dNTPs (各 2.5mM,タカラバイオ) 8.0 µL Oligo (dt) 15 (10 pmol/µL,タカラバイオ) 1.0 µL
28
用した。反応液はTable 2-3 のように調製した。THUNDERBIRD SYBR qPCR Mix は SYBR Green 1 検出系を利用しており、PCR 反応により合成された DNA 存在量は蛍光強度により検 出される。蛍光データ回収ポイントはextend の際に行い、サイクル数は 45 とした。データ解析 ソフト Thermal Cycler Dice Real Time Systemsingle Software を用いて、2nd Derivative Maximum 法(SDM 法)により CT 値を求めた後、比較 CT 法(⊿⊿CT 法)により遺伝子発現量 を定量解析した。
Table 2-3 Real-time quantitative PCR の反応液組成
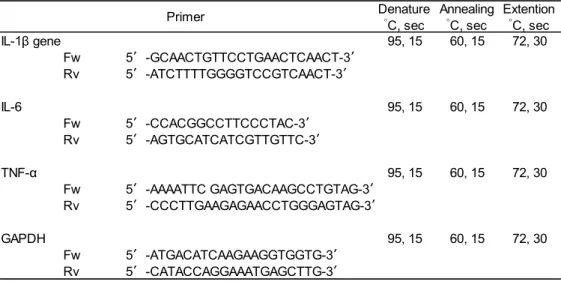
本実験で使用したプライマーおよびPCR 反応条件は Table 2-4 に示した。
Table 2-4 プライマーおよび PCR 反応条件 THUNDERBIRD SYBR qPCR Mix 7.5 µL
MilliQ 6.4 µL 5' primer 0.3 µL 3' primer 0.3 µL Template 0.5 µL IL-1β gene 95, 15 60, 15 72, 30 Fw 5′-GCAACTGTTCCTGAACTCAACT-3′ Rv 5′-ATCTTTTGGGGTCCGTCAACT-3′ IL-6 95, 15 60, 15 72, 30 Fw 5′-CCACGGCCTTCCCTAC-3′ Rv 5′-AGTGCATCATCGTTGTTC-3′ TNF-α 95, 15 60, 15 72, 30 Fw 5′-AAAATTC GAGTGACAAGCCTGTAG-3′ Rv 5′-CCCTTGAAGAGAACCTGGGAGTAG-3′ GAPDH 95, 15 60, 15 72, 30 Fw 5′-ATGACATCAAGAAGGTGGTG-3′ Rv 5′-CATACCAGGAAATGAGCTTG-3′
29 2-3 ペクチン摂取がマウスパイエル板細胞の IL-6 産生に与える影響の解析 2-3-1 試薬の調製 ・8 %炭酸水素ナトリウム溶液 炭酸水素ナトリウム(和光純薬工業、大阪)8 g を Milli Q 水に溶解し、100 mL にメスアップし た。その後培地瓶に入れ、121˚C、20 分間オートクレーブした。 ・3 % L-グルタミン溶液
L-グルタミン(Thermo Fisher Scientific)3 g を MilliQ に溶解し、100 mL にメスアップした。そ の後フィルター濾過滅菌(pore size 0.22 μM、Millipore)した。
・FBS
-20˚C で保存していた Fetal bovine serum (FBS)を 37˚C で解凍し、56 ˚C、30 分間インキュ ベート(非動化)した。
・ペニシリン-ストレプトマイシン混合溶液
ペニシリン-ストレプトマイシン混合溶液は 10 mL ずつ分注し、-20˚C で保存した。 ・10% FBS を含む RPMI 培地
Roswell park memorial institute(RPMI)1640 培地(日水製薬、東京)粉末 10.2 g を Milli Q 水 1000 mL に加え、培地瓶中で溶解した。その後 121 ˚C、20 分間オートクレーブした。8%炭 酸水素ナトリウム溶液20 mL、FBS 100 mL、3% L-グルタミン溶液およびペニシリン-ストレプト マイシン混合溶液を10 mL ずつ加え撹拌し、10 % FBS 含有培地として、4˚C で保存した。 ・1000 ng/mL LPS、10 % FBS を含む RPMI 培地 1 mg/mL LPS を PBS で希釈し、1000 ng/mL LPS、10 % FBS を含む RPMI 培地とした。 2-3-2 マウスへのペクチン給餌 第2 章 2-2-2 と同様に実施した。
30 2-3-3 パイエル板細胞の分離 パイエル板の採取はクリーンベンチ内で無菌的に行った。10 日間ペクチン水溶液を給餌し たマウスに頸椎脱臼を処置した後、開腹し小腸を露出させ、冷 PBS 上で慎重にパイエル板を 採取した。10 mL の冷 10% FBS を含む RPMI を 10 cm dish に加え、その上に 70 µm ナイロン メッシュ(CORNING)を置いた。ナイロンメッシュに採取したパイエル板を移した。マウス 1 匹あ たり5 個前後のパイエル板が回収できた。これを 1 mL シリンジの注射筒で丁寧にホモジナイ ズし、単細胞懸濁液を調製した。懸濁液を15 mL チューブに移し、300 × g、25˚C、5 分間遠 心分離を行い、細胞が沈殿していることを確認した。10% FBS RPMI 培地を 1 mL 加え、沈殿 した細胞を懸濁し、Neubauer 型血球計算板(エルマ販売)を用いて細胞数を計算した。再び 300 × g、25˚C、5 分間遠心分離を行い、上清を吸引除去し、細胞数が 2.5×105 cells/100 µL と なるように1000 ng/mL LPS、10% FBS 含有 RPMI 培地を加えた。96 well プレートに細胞懸濁 液を 100 µL ずつ加え、37˚C、5% CO2条件下で培養した。24 時間後、培養上清を回収し、 -20˚C で保存した。 2-3-4 パイエル板細胞が産生する IL-6 の定量 第2 章 2-2-5 と同様に実施した。 2-4 磁気ビーズ分離法によるパイエル板細胞の精製 2-4-1 試薬の調製 ・10 % FBS を含む RPMI 培地 第2 章 2-3-1 と同様に調製した。 ・1000 ng/mL LPS、10 % FBS を含む RPMI 培地 第2 章 2-3-1 と同様に調製した。 ・Digestion medium
31 60 mg ディスパーゼ(合同酒精、東京)および 0.30 mg DNase I を 10 mL RPMI 培地に溶解 し、0.22 µm フィルターで滅菌した。これに FBS を 10 %となるように加えた。 2-4-2 マウスへのペクチン給餌 第2 章 2-3-2 と同様に実施した。 2-4-3 パイエル板 B220+ 細胞の分離・精製 第2 章 2-3-3 と同様にパイエル板細胞を採取した。分離した細胞を 0.5 % FBS を含む PBS 100 µL に再懸濁した。Anti-mouse CD16/32 Fc receptor (2.4G2、TONBO bioscience、兵庫) を 1 × 106 cells あたり 1 µL 加え、15 分間、氷上でインキュベートした。biotin-labeled anti-CD45R/B220 (clone RA3-6B2、Biolegend、San Diego、CA)を 1 × 106 cells あたり 1 µL
加え、15 分間、氷上でインキュベートした。5 分間、4˚C、800 × g で遠心した後に、0.5% FBS を含む PBS 100 µL に再懸濁した。次いで、anti-biotin magnetic beads(Miltenyi Biotech、 Bergisch Gladbach、Germany)を 5 µL 加えて 15 分間、氷上でインキュベートした。5 分間、4˚C、 800 × g で遠心した後に、0.5 % FBS を含む PBS 500 µL に再懸濁した。細胞懸濁液を Magnetic-activated cell sorting (MACS) セパレーターにセットした MACS MS カラムにアプラ イし、0.5 % FBS を含む PBS 500 µL で 3 回洗浄した。溶出液を 2.0 mL チューブに回収した (B220‐細胞が含まれる)。MACS MS カラムを MACS セパレーターから外し、0.5 % FBS を含 むPBS を 1 mL に加えて、プランジャーにより溶出した(B220+細胞が含まれる)。溶出液を1.5 mL チューブに回収し、2.0 mL チューブに回収した溶出液と共に 5 分間、4˚C、800 × g で遠 心した。0.5 % FBS PBS 1 mL に再懸濁した後に、細胞数を計算した。再び 5 分間、4˚C、800 × g 遠心分離を行い、上清を吸引除去し、B220‐細胞とB220+細胞は2.5×105 cells/100 µL と なるように1000 ng/mL LPS、 10 % FBS を含む RPMI 培地を加えた。細胞懸濁液はそれぞれ 96 well プレートに 100 µL ずつ播種し、37˚C、5 % CO2条件下で培養した。24 時間後、培養上
32 清を回収し、-20˚C で保存した。 2-4-4 パイエル板 CD11c+ 細胞の分離・精製 第2 章 2-3-3 と同様にパイエル板を採取した。採取したパイエル板を Digestion medium に移 し、90 分間 37˚C で穏やかに回転させながらインキュベートした。インキュベートした組織を 70 µm ナイロンメッシュ上でホモジナイズし、細胞懸濁液を調製した。懸濁液を 15 mL チューブに 移し、300 × g、25˚C、5 分間遠心分離を行い、0.5 % FBS を含む PBS 1000 µL に再懸濁した。 細胞数を計算し、5 分間、4˚C、800 × g で遠心した後に、0.5 % FBS を含む PBS 100 µL に 再懸濁し、Anti-mouse CD16/32 Fc receptor (2.4G2、TONBO bioscience)を 1 × 106 cells あ たり1 µL 加え、15 分間、氷上でインキュベートした。biotin-labeled anti-CD11c(Clone N418、 Miltenyi biotech)を 1 × 106 cells あたり 5 µL 加え、15 分間、氷上でインキュベートした。5 分
間、4˚C、800 × g で遠心した後に、0.5 % FBS PBS 100 µL に再懸濁した。次いで、 anti-biotin magnetic beads(Miltenyi Biotech)を 5 µL 加えて 15 分間、氷上でインキュベートし た。5 分間、4˚C、800 × g で遠心した後に、0.5 % FBS を含む PBS 500 µL に再懸濁した。 細胞懸濁液を MACS セパレーターにセットした MACS MS カラムにアプライし、0.5 % FBS PBS 500 µL で 3 回洗浄した。溶出液を 2.0 mL チューブに回収した(CD11c-細胞が含まれる)。 MACS MS カラムを MACS セパレーターから外し、0.5 % FBS PBS を 1 mL 加えて、プランジャ ーにより溶液した(CD11c+細胞が含まれる)。溶出液を 1.5 mL チューブに回収し、2.0 mL チュ ーブに回収した溶出液と共に5 分間、4˚C、800 × g で遠心した。0.5 % FBS を含む PBS 1 mL に再懸濁した後に、細胞数を計算した。再び 5 分間、4˚C、800 × g 遠心分離を行い、上 清を吸引除去し、CD11c- 細胞と CD11c+ 細胞を 1.0×104 cells/100 µL もしくは 2.5×104 cells/100 µL となるように 1000 ng/mL LPS、10 % FBS を含む RPMI 培地に加えた。細胞懸濁 液はそれぞれ96 well プレートに 100 µL ずつ播種し、37˚C、5% CO2条件下で培養した。24 時間後、培養上清を回収し、-20˚C で保存した。
33 2-4-5 パイエル板細胞が産生する IL-6 の定量 第2 章 2-2-5 と同様に実施した。 2-5 細胞の培養 2-5-1 試薬の調製 ・マウスMΦ 様細胞株 RAW264.7 第2 章 2-1 に示した。 ・8 %炭酸水素ナトリウム溶液 第2 章 2-3-1 と同様に調製した。 ・3 % L-グルタミン溶液 第2 章 2-3-1 と同様に調製した。 ・FBS 第2 章 2-3-1 と同様に調製した。 ・10 % FBS を含む DMEM
DMEM(Dulbecco’s Modification Eagle’s Medium、日水製薬)粉末 1.9 g を Milli Q 水 200 mL 加え、培地瓶中で溶解した。その後 121 ˚C、20 分間オートクレーブした。8%炭酸水素ナト リウム溶液 4 mL、FBS 22 mL、3 % L-グルタミン溶液 を 2.2 mL 加え撹拌し 10 % FBS 含有培 地とし、4˚C で保存した。 ・PBS 第2 章 2-2-1 と同様に実施した。 2-5-2 細胞の継代 10 cm dish を用いて、10 % FBS を含む DMEM、37˚C、5 % CO2条件下で培養した。継代は
34 3 日ごとに行った。80 %コンフルエントの状態で、細胞上清を吸引除去し、PBS を 10 mL 加え た。ピペッティングで接着している細胞を剥し、細胞懸濁液を 15ml 遠心チューブに移した。こ れを300 × g、25˚C、5 分間遠心分離した。上清を吸引除去した後、新しく PBS を 1 mL 加えた。 沈殿した細胞をピペッティングで穏やかに洗浄し、この細胞懸濁液にPBS を 9 mL 加え、300 × g、25˚C、5 分間遠心分離した。上清を吸引除去した後、新しく PBS を 1 mL 加え、細胞懸濁液 1 mL を調製した。調製した細胞懸濁液より 50µL を 1.5 mL チューブに移し、PBS を加えて 10 倍希釈した。この懸濁液50 µL をトリパンブルー溶液 50 µL と懸濁した。Neubauer 型血球計算 板を用いて4 箇所の合計生細胞数を数え、その数から細胞懸濁液中の細胞濃度を計算した。
10 % FBS を含む DMEM を 10 mL 加えた 10 cm dish に、細胞濃度が 1×106 cell/dish(3 日継
代)となるように細胞懸濁液を加えた。顕微鏡を用いてシャーレに細胞が存在することを確認し 37˚C、5 % CO2条件下で培養した。 2-5-3 細胞の保存 コンフルエントに達した細胞を 10 cm dish から剥がし、遠心分離により上清除去後、 Neubauer 型血球計算板を用いて細胞数を計算した。再び 300 × g、25˚C、5 分間遠心し、上清 を吸引除去した。その後、細胞濃度が1×106cells/mL となるように Bambanker(日本ジェネティ クス、東京)を1 mL 加えた。細胞懸濁液を、-30˚C で冷却したセラムチューブ(住友ベークライ ト、東京)に1mL ずつ分注し、その後-80˚C で凍結保存した。 2-5-4 凍結細胞の融解 凍結保存しておいたセラムチューブを37˚C の湯浴中で温め、氷が少々残る程度まで解凍し た。全量を15 mL チューブ中の PBS 9 mL に移し、300 × g、25˚C、5 分間遠心分離した。上清 を除去し、新たにPBS を 10 mL 加えて懸濁し、300 × g、25˚C、5 分間遠心分離した。上清を除 去し、10 % FBS を含む DMEM を 10 mL 加えて懸濁し、10 cm dish に移し、37˚C、5 % CO2