アミノ酸における結晶形状制御
長谷川和宏
1*・佐野千明
2Crystal Shape Control for Amino Acids Crystals
Kazuhiro Hasegawa
1*and Chiaki Sano
2Amino acid crystal shape is affected by impurities those are included in the fermented broth. To control crystal shape is important for final product properties and purification efficiency. To control crystal shape, it is necessary to know how affect to the crystal shape by impurities. Crystal shape is depending on the crystal growth of each growth surface. To know the effect of impurity effects for crystal growth, relation between crystal structure is important. In this report, we estimated the inhibition mechanisms of crystal shape and crystal growth using L-sodium glutamate and Leucine DMBS complex crystals by studying between crystal structure and crystal growth.
1. はじめに
生体中に存在するタンパク質は 20 種類のアミノ酸か ら構成されている.それらのアミノ酸の基本骨格は同一 であるが,側鎖の違いにより溶解度,等電点(pI)など基 本物性に違いがあり,Table 1 に示すように分類されてい る 1).現在,これらのアミノ酸は主として発酵法により 製造されており,飼料,食品,医薬品等の分野で広く利 用されている.アミノ酸の用途として最もよく知られて いるのは,L-グルタミン酸ナトリウム(商品名「味の 素」)の調味料としての利用であり,そのルーツは昆布の うまみの主成分として,晶析操作により1908 年に池田菊 苗博士(東京大)によって単離精製された. 一方,スポーツを愛好する多くの方々にもアミノ酸の 働きが理解されるようになり,幅広く使われるようにな っている.アミノ酸は体を動かす筋肉の主成分であるタ ンパク質の構成成分であり,運動によりその必要量が高 まることが知られている.アミノ酸は摂取してから吸収 までの時間が早く,必要な時にすぐに利用できるという すぐれた特徴から,アミノ酸含有補助食品(サプリメン ト)として,エネルギー源,疲労回復・疲労予防源とし て有効である. また,アミノ酸は医薬品等の医療関連の用途でも用い られている.手術の前後等で充分な栄養をとることがで きない患者に対して,必要なアミノ酸をアミノ酸輸液と して患者の静脈内に投与して栄養を補給するものである. 1956 年に発売されて以来各国で改良が進められ,アミノ 酸,糖質,電解質などを含んだ高カロリー輸液が定着し, 患者の栄養管理だけでなく,手術の成功率を高くするこ解説
1 味の素株式会社 イノベーション研究所 〒210-8681 神奈川県川崎市川崎区鈴木町 1-1 2 公益財団法人 日本農芸化学会 〒113-0032 東京都文京区弥生 2-4-161 Institute for innovation, AjinomotoCo., Inc. 1-1 Suzuki-cho, Kawasaki-ku, Kawasaki, 210-8681 Japan
2 Japan Society for Bioscience, Biotechnology, and Agrochemistry, 2-4-16, Yayoi, Bunkyo-ku, Tokyo 113-0032, Japan * Corresponding author (e-mail: [email protected])
MW water solubility (g/100g 25℃) Isoelectric pH 1 Alanine 89.1 16.5 6.01 2 Arginine 174.21 15 10.76 3 Asparagine 132.12 3.1 5.41 4 Aspartic acid 133.11 0.5 2.77 5 Cysteine 121.16 v. sol 5.02 6 Glutamic acid 147.14 0.84 3.24 7 Glutamine 146.15 3.6 5.65 8 Glycine 75.07 25 5.97 9 Histidine 155.16 7.59 7.59 10 Isoleucine 131.18 4.12 6.02 11 Leucine 131.18 2.3 5.98 12 Lysine 146.19 v. sol 9.82 13 Methionine 149.22 3.5 5.74 14 Phenylalanine 165.2 2.97 5.48 15 Proline 115.14 162.3 6.3 16 Serine 105.1 5 5.68 17 Threonine 119.12 20.5 6.16 18 Tryptophane 204.23 1.14 5.89 19 Tyrosine 181.2 0.05 5.66 20 Valine 117.15 8.85 5.96 Table 1 Some physicochemical properties of the primary
とも認められてきている. こうした,種々の機能を持ったアミノ酸のほとんどの 製品形態は結晶である.従ってアミノ酸を製品として取 得する為の晶析操作は,製品の物性,純度等を決定する 重要操作であり,アミノ酸の種々の性質を考慮した様々 な工夫がなされている. アミノ酸の製造工程を簡単に述べると,ほとんどのア ミノ酸は糖を原料とした発酵法により生産され,アミノ 酸を蓄積した発酵液から除菌,粗製,精製工程を経て製 品化される.発酵工程では発酵菌や発酵培地組成成分を はじめ,副生アミノ酸,有機酸,蛋白質など多くの成分 が夾雑している.これらの夾雑物と,目的のアミノ酸を 分離するためには膜分離,樹脂操作などの方法も用いら れるが,精製工程では晶析技術に負うところが大きい. また製品として結晶を析出させる場合,結晶形状や大き さ,外観,易溶性の観点も重要になってくる.それら粉 体としての特性は晶析工程で決められる部分も大きい. この様に商品価値を決定する製品晶析に際しては,晶析 条件・設備への配慮だけでは不十分であり,晶析系に持 ち込まれる夾雑物量を常に安定に保つこと,即ち精製系 全体の構成も視野に入れることが必須になる.
2. L-グルタミン酸ナトリウム結晶の外観
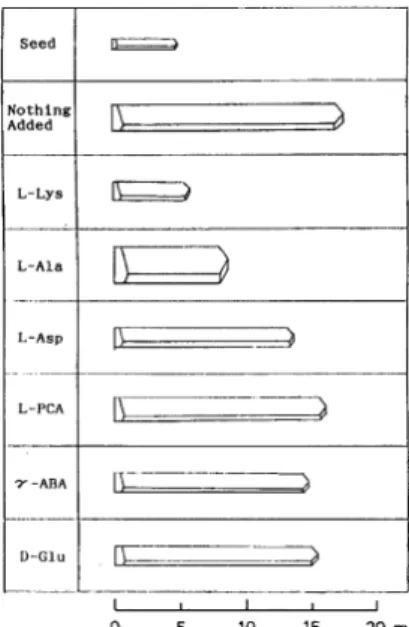
アミノ酸結晶析出時の結晶形状や大きさ,光沢等の外 観の制御について,L‐グルタミン酸ナトリウム(Mono Sodium Glutamate, MSG)を例に述べる. Fig. 1 に概略を示したように,L‐グルタミン酸ナトリ ウムの代表的な製造方法は,大きく二つの工程から成る. 先ず天然原料である澱粉を酸糖化又は酵素糖化して得ら れるブドウ糖,或いは砂糖を製造する際の副産物である 糖蜜を主原料に,微生物を利用して L‐グルタミン酸の 分子骨格を形成させる発酵工程である.次にその発酵液 から種々の精製手段を駆使し,最終的に L‐グルタミン 酸ナトリウムを結晶として取り上げ商品化する単離精製 工程がある. この L‐グルタミン酸ナトリウムの晶析工程では, 種々晶析条件(例えば温度・晶析速度)を操作し,結晶 の大きさ・光沢等を制御している.また,大きな結晶を 晶析する場合は,結晶が破砕しない様に設備仕様も考慮 する必要がある. 一方,結晶の外観,特に結晶の大きさと形を制御する という意味では晶析系に微量ながら存在するアミノ酸量 をコントロールする場合がある.発酵法による L‐グル タミン酸ナトリウムの製造に於いては,微生物の代謝副 生物であるアミノ酸が製品晶析系に微量ながら混入して くる場合が多くなる.これらのアミノ酸は晶析の持つ不 純物淘汰能により製品晶析過程で除去されるので,製品 純度の観点で問題となることはないが,ごく微量のアミ ノ酸が結晶の形を変化させる効果により商品価値に影響 を及ぼす場合がある. L‐グルタミン酸ナトリウム結晶の外形に及ぼす夾雑 アミノ酸の影響を基礎的に解明した結果を Fig. 2 に示す 2).各種アミノ酸を共存させた L‐グルタミン酸ナトリウ ム溶液の中で L‐グルタミン酸ナトリウムの結晶を成長 させた時に,得られた結晶の形がどの様に変化するか観 察した結果である.最も良く成長した無添加系の結晶に 対し,例えば L‐アラニンが共存する場合は長軸方向が 相対的に短くなり,ずんぐりとした見かけの結晶になる. これはL‐アラニンが L‐グルタミン酸ナトリウム結晶の 長軸端に選択的に吸着することで長軸方向の結晶成長を 阻害することによる.一方,L‐アスパラギン酸が共存す る場合は,その形は L‐アラニンの場合とは大きく異な り殆ど純系の結晶の相似形であるが,添加物の存在しな い系に比べて小さな結晶となる.この現象は L‐アスパ ラギン酸がL‐グルタミン酸に対してメチレン鎖が一つFig. 1 Sodium glutamate production scheme (From Ajinomoto Co., Inc. HP).
Fig. 2 L-sodium glutamate crystal shape changing by additives.
短いだけの非常に類似した結晶構造を持つので,結晶の 各面にほぼ均一に吸着し全体的に結晶成長を阻害するた めと考えられる.また D‐グルタミン酸や有機酸の様な 物質はより細長い形状に変化した.これは D‐グルタミ ン酸などは単軸方向に吸着し,成長を阻害した結果と考 えられる.
3. L-グルタミン酸ナトリウム結晶の形状と
結晶構造との関係
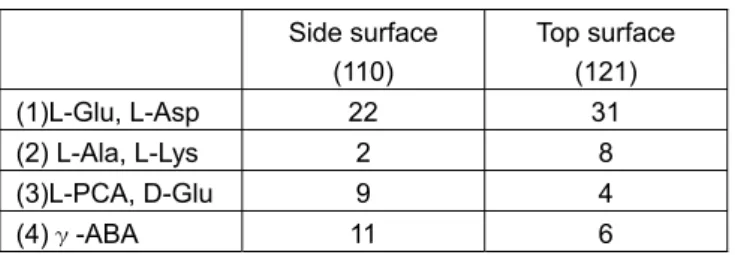
これらの現象からそれぞれの夾雑物は結晶の特定の面 に吸着し,その面の成長を阻害しているためと考えられ る.続いて各夾雑物による各成長面への吸着機構につい て解析する. 結晶成長は構成する分子が成長面へ吸着して構造中に 取り込まれてゆく.したがってまず,成長面の構造につ いて解析する必要がある.そこでまず L‐グルタミン酸 ナトリウムの結晶構造を Fig. 3 に示す 3).また結晶外形 と結晶構造から推定した分子のパッキング様式を Fig. 4 に示す2). L‐グルタミン酸ナトリウムのグルタミン酸分子は 2 種 類の配置をしており,それらを GluA,GluB として表記 している.これらGluA と GluB は互い違いにある角度を 持って2分子づつジグザグにつながりながら c 軸方向に 重ねあっており,この角度が{121}面の角度に反映されて いる.側面の{110}面は Na+イオンが c 軸方向に柱状に 連なっている.これらのパッキング様式に従って分子が結 晶に取り込まれ,成長しているが,分子が取り込まれる 場所としてキンクと呼ばれる部分で分子が結合を形成し, 次の分子の吸着により結晶に取り込まれ,安定化される と考えられる.従って,これらの結晶構造のデータから 各結晶面のキンクにおける分子結合の様式から成長特性 が推定でき,夾雑物による阻害機構も推定できると考え る. そこで続いて各キンクサイトにおける夾雑物分子の結 合強度について評価を行った.各キンクにおいて夾雑物 が結合するためには本来結合するべきグルタミン酸分子 と同様に水素結合と Na+イオンへの配位による静電的な 結合が関与していると考えられる.そのうち水素結合は H-O の距離,角度等が結合形成のためには制限されるた め,分子認識により関与する.従って,夾雑物のキンク サイトへの吸着には水素結合の形成の可否,水素結合形 成の数を評価し,より多くの水素結合を形成するキンク サイトの数が多いほど夾雑物の影響を受けると考えた. また各結晶面におけるキンクサイトは結晶成長面をXY 面 とすると,各成長面でのキンクサイトは Fig. 5 に示すよ うな4種類が考えられる.L‐グルタミン酸ナトリウムの 結晶(空間群 P212121)の単位格子中には等価点が4個あ り,等価点には2分子(GluA,GluB)が存在する.従って, 成長面のキンクへの取り込みには4種類のキンクサイト への8分子の取り込み,すなわち32通りについて評価 を行った. グルタミン酸のα‐アミノ基,γ‐カルボキシル基と 同様に水素結合を4つ形成するパターンを各夾雑分子と 成長面について評価を行った結果をTable 2 に示す.Fig. 3 Crystal structure of L-Sodium glutamate.
Fig. 4 L-sodium glutamane crystal shape and schematic drawing of molecular packing.
Fig. 5 Kink site shapes on the L-sodium glutamane crystal grawing surface.
Side surface (110) Top surface (121) (1)L-Glu, L-Asp 22 31 (2) L-Ala, L-Lys 2 8 (3)L-PCA, D-Glu 9 4 (4)γ-ABA 11 6 これらの結果から,本来の L-Glu 分子は先端面に吸着し やすい.この結果は先端面の方で成長速度が高く,その 結果形成される結晶形状と一致している.L-Asp も L- Glu と同様に吸着されるため,結晶形状には影響を与え ない.一方L-Ala や L-Lys は先端面の方がより吸着しや すく,また L-PCA,D-Glu,γ-ABA は側面の方が吸着 しやすい.これらの結果は夾雑物による結晶形状の影響 と一致しており,結晶構造解析から夾雑物による結晶形 状の与える影響について予測可能であることが示された と考えられる.
4. ロイシン複合体結晶の成長阻害機構
筋肉の組織は,アクチンとミオシンというタンパク質 でできている.このアクチンとミオシンの主成分はバリ ン,ロイシン,イソロイシンという 3 種類の必須アミノ 酸 で , こ れ ら は 分 岐 鎖 ア ミ ノ 酸 (BCAA = Branched Chain Amino Acid)と呼ばれている.ロイシンはこの BCAA の一つであり,食餌により摂取する必要がある必 須アミノ酸の一種である.そのため.先に述べた機能性 食品や輸液等に用いられている.ロイシンはタンパク質 の加水分解や発酵液から単離・生成される.タンパク質 の加水分解物中には同じ BCAA であるイソロイシンやバ リンも混在しており,これらを効率的に分離することが 工業化プロセスのポイントになる.しかしこの3種のア ミノ酸の結晶構造はよく似ており 4-6),ロイシン結晶中に はイソロイシンやバリンが取り込まれ精製効率が高くな く,高度な精製をするためには結晶化を繰り返さなくて はならない.そのような結晶化を繰り返すことなく純度 の良いロイシンを得るために,ロイシンと選択的に結合 する芳香族スルホン酸の開発はなされ,その中で 3,4‐ジ メチルベンゼンスルホン酸はロイシンとの親和性が高く, 溶解度が低く,イソロイシンやバリンを高い効率で排除 することができる特殊沈殿剤として用いられてきた7). ロイシンと 3,4‐ジメチルベンゼンスルホン酸との複合 体は pH2.3 以下になるとロイシンが正に帯電し,3,4‐ジ メチルベンゼンスルホン酸が有する負電荷と引き合い複 合体を形成するようになる.(Fig. 6)またロイシン(Leu) と 3,4‐ジメチルベンゼンスルホン酸(DMBS)との複合体 の結晶構造8)の c 軸投影図を Fig. 7 に示した. この結晶では ac 面に沿った方向に水素結合のネットワ ークが形成されており,b 軸方向ではそのネットワーク同 士が van der Waals 力で積層している構造を持っている. このような構造が結晶析出時の分子認識にどのような影 響を与え,精製度,結晶成長にどのように寄与するかを 検討した. まず Leu.DMBS 結晶の他のアミノ酸の排除性(淘汰 性)について有効分配係数という因子で精製度を評価し た結果を Fig. 8 に示す.有効分配係数は次式であらわさ れ,keffが1より小さければ結晶化を行うことで精製効果 を得ることができる9). keff = Ric / RiL ここで,Ricは結晶中の不純物アミノ酸のモル濃度,RiL は溶液中の不純物アミノ酸のモル濃度である.Leu を単 独で結晶化を行うとよく似た構造であるイソロイシンの Table 2 Impurity adhering site analysis of L-sodiumglutamate crystal.
Fig. 6 Molecular structures of Leucine and 3,4-dimethylbenzene sulfonate.
Fig. 7 Leu.DMBS crystal packing (projection figure from C-axis)2 unit cells are shown for a-axis Glay; Carbon, Blue: Nitrogen, Red: Oxigen, Yellow: Sulfur, Dotted line shows hydrogen bonds.
淘汰性は keffであらわすと 0.300 であるが,Leu.DMBS
結晶を用いることで0.0084 になり,効率的にイソロイシ ンを排除し,精製できることを示している.
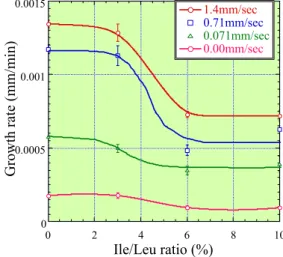
このように淘汰性が向上する要因として,結晶中の分 子パッキングの状態が挙げられる.Leu.DMBS 結晶では van der Waals 力で積層している方向で Leu 側鎖が DMBS のベンゼン環及びメチル基と重なり合わないよう にパッキングしており,その構造の密さから分子認識で できているものと考えられる.そのような分子認識の高 さにおいても若干のIle の取り込みが認められることから, この取り込みがLeu.DMBS の結晶成長速度にどのように 影響するかを検討した. アミノ酸を結晶化する過程においては晶析槽など流れ のある中で結晶が成長することが多い.そのため本検討 においては,流れのない静止系での結晶成長速度に加え て,結晶に対して溶液の流れのある場合の結晶成長速度 と,その時の影響について検討を行った.Leu.DMBS の 結晶はFig. 9 の形をしており,そのうち今回は(011)面の 成長速度の測定を行った. 実験系の模式図を Fig. 10 に示す.図のように(001)面 に対して平行になるように溶液の流れを作り,成長速度 の測定を行った. 各流速での成長速度の測定結果を溶液中のイソロイシ ン濃度に対してプロットした図をFig. 11 に示す. このグラフから,流れをかけるに従い,結晶成長速度 が増加していることがわかる.また流れのない場合や流 速 1.4mm/sec までの流れのある場合いずれも,Ile/Leu 濃度 3%程度までは成長速度の低下はわずかであるが, 3%を越えると急激に成長速度の低下が見られる.そして, その後一定値になっていることが分かった. 次に成長速度と不純物濃度の関係から不純物の成長速 度抑制機構について考察する.不純物が吸着して成長速 度を抑制するサイトはキンク,ステップ,テラスの3種 類がある.キンクやテラスに不純物が吸着し,それが原 因で成長速度が抑制される場合には,不純物濃度が増す につれて成長速度が低下するような現象が現れるはずで あ 10).しかし,今回の実験ではIle/Leu が 3%までは成 長速度の抑制がほとんどなく,それ以上で急激に抑制さ れている.この挙動は特徴的であり,不純物がステップ の掃引を抑制する場合に相当する.この場合,不純物が 完全に成長を止めてしまわない場合には,ステップは不 0 0.0005 0.001 0.0015 0 2 4 6 8 10 1.4mm/sec 0.71mm/sec 0.071mm/sec 0.00mm/sec
Growth rate (mm/min)
Ile/Leu ratio (%)
Fig. 8 Isoleucine rejection by Leu.DMBS crystallization. Ile mole ratio in crystal vs. Ile mole ration in mother liquor.
Fig. 9 Leu DMBS crystal shape and growth faces.
Fig. 10 Leu.DMBS crystal and flow direction at the experiment.
Fig. 11 Leu.DMBS crystal growth difference by adding Isoleucine.
純物の間をすり抜けるように前進してゆく.このメカニ ズムでは,不純物濃度より小さい場合には,吸着した不 純物間隔が大きく,ステップが不純物をすり抜ける時の 曲率が大きくても抜けられるので成長速度の抑制はほと んど起こらない.しかし,一定の濃度を超えると不純物 によりステップが曲げられるので,成長速度が急激に低 下する 11).今回の実験では不純物である Ile が成長面に 取り込まれたとき,その次の層のステップが不純物上を 掃引するときにFig. 12 の様な不純物による‘ピニング’ 効果を生じ,成長速度が抑制されていると考えられる. 続いて結晶構造を考えて,Leu.DMBS 結晶中に Ile が どのような場合に入り込み,また,入り込んだIle がどの ような機構で成長を阻害するかを検討した. 結晶構造から(011)面上で Leu.DMBS がどのように入 り込むかを調べたところ,Fig. 13 に見られるように2種
類の入り方が考えられた.Fig. 13 の CaseA の様に Leu の側鎖を結晶側に向けている場合と,CaseB の様に外側 を向いている場合とがあげられる.
もし,Ile が Leu のサイトに置換して取り込まれる場合, アミノ基やカルボキシル基の部分は Leu,Ile とも共通で あるので,側鎖の違いが取り込みに反映すると考えられ る.そのため,Fig. 8 の CaseA の様な場合に Leu サイト を Ile に置換すると,Ile 側鎖のβ位のメチル基が結晶側 を向いてしまうので,この部分が結晶側の分子と立体障 害を生じると考えられる.従って,CaseA の様な場合に は Leu は Ile に置換することはできない.そのため,さ らに積層し,成長する場合での障害となることはない. しかしFig. 8 の CaseB の様な場合,β位にメチル基が 存在したとしてもその方向は結晶面とは反対側で,Ile が Leu に置換するにあたっての立体障害は CaseA に比べて 小さくなると予想される.したがって,Ile が Leu に変わ って置換される可能性が生じる.一旦置換されてしまう と今度は外側を向いたβ位のメチル基がそれ以上に成長 しようとする場合の障害となってしまう.このβ位のメ チル基は,この位置にさらにLeu や DMBS が接近しよう とすると,メチル基との間に立体障害を起こすため,成 長を阻害すると考えられる. 夾雑物としてのIle はこのような機構で成長速度の抑制 に寄与していると考えられ,このような考え方は Tailor-made additive による成長抑制と分類されている 10,12).
Tailor-made additive では今回の Leu と Ile の関係のよう に,ホスト分子に非常によく似た添加物を加えると,そ の添加物の結晶への取り込まれ方が面により異なるため に,その成長速度の抑制にも違いが現れる.そこで,こ の添加物の種類を選定することで,結晶形状の制御への 展開が可能にするというものである13).
5. まとめ
以上の様にアミノ酸の結晶形状は夾雑物によって影響 を受ける.その機構としては結晶表面に夾雑物の分子が 取り込まれ,成長を阻害しているためと考えられる.ま たこれらの機構は結晶構造とも密接な関係を持っている と考えられる.そのため結晶構造と成長速度等の解析を 通じて,夾雑物による成長速度への影響や,その結果生 じる形状変化の推定にもつながるものと考える.参 考 文 献
1) P. M. Hardy: Chemistry and Biochemistry of the Amino Acids, G. C. Barrett ed., Chaoman and Hall, NY, 1985
2) Chiaki Sano, Nobuya Nagashima, Tetsuya Kawakita, and Yoichi Iitaka: Journal of Crystal Growth, 99 (1990) 1070
3) Chiaki Sano, Nobuya Nagashima, Tetsuya Kawakita, and Yoichi Iitaka: Analytical Science Feb., 5 (1989) 121
4) M. M. Harding and R. M. Howiesom: Acta Cryst., B32 (1976) 633
5) K. Torii and Y. Iitaka: Acta Cryst., B27 (1971) 2237
Steps
impurity 不純物がステップに吸着した場合
step advance impurity step advance
Ile/Leu≦3% 成長速度への影響小 3%≦Ile/Leu 成長速度急激に低下 (011) DMBS Leu α β γ crystal surface
crystal surface Leu
α β γ DMBS (011) Case A Case B
Fig. 12 Crystal growth inhibition mechanism by impurities.
Fig. 13 Molecular steric hindrance difference by molecular direction.
CaseA: Ile cannot replace Leu CaseB: Ile can replace Leu
6) K. Torii and Y. Iitaka: Acta Cryst., B26 (1970) 1317
7) D. Nagai: Japan patent No., 40-11373 (1965) 8) K. Hasegawa, K. Ishikawa, R. Kawaoka, C. Sano,
K. Iitani, H. Komatsu, and N. Nagashima: Acta Cryst., C54 (1998) 637-641
9) T. Nakai: Shoseki Kogaku, 1986
10) I. Weissbuch, L. Addadi, M. Lahav, and L. Leiserowits: Science, 253 (1991) 637
11) R. J. Davey and J. W. Mullin: Journal of Crystal Growth, 26 (1974) 45
12) S. N. Black, R. J. Davey, and M. halcrow: Journal of Crystal Growth, 79 (1986) 765
13)
J. L. Wang, Z. Berkovitch-Yellin, and L. Leiserowits: Acta Cryst., B41 (1985) 341