2
クロマチン免疫沈降法
3
New SimpleChIP® Plus Enzymatic Chromatin IP Kit
4
CST
社キットを用いたマウス組織の
ChIP
アッセイ
5
ヒト大腸がん細胞株
HCT116
を用いた
p53
の
ChIP
解析
6
酵素によるクロマチン断片化の最適化
7
よくあるお問い合わせ
クロマチン免疫沈降
(
ChIP
)
SimpleChIP® Plus Enzymatic Chromatin IP Kit
一般的に、ChIPで得られたDNAはスタンダードまたは定量的PCRで分析します。その他、ChIPアッセ イを、シークエンシング(ChIP-seq)、ゲノムタイリングマイクロアレイ(ChIP-on-chip)法、クロー ニング法などと組み合わせ、タンパク質–DNA相互作用やヒストン修飾をゲノムレベルで解析すること もできます(5–8)。 可溶化後、クロマチンを酵素/ ソニケーションで断片化します。 ヒストンや非ヒストンタンパク質特異的抗体と
Protein G Agarose/Magnetic Beadsを
用いて免疫沈降します。 ヒストンや非ヒストンタンパク質とDNAを クロスリンクするために、細胞や分離組織を ホルムアルデヒドで固定します。 脱クロスリンク後、DNAを 分析用に精製します。
PCR primers are specific for the transcriptionally active GAPDH and c-Myc genes and the inactive Myt1 gene. Tri-Methyl-Histone H3 (Lys4)(C42D8) Rabbit mAb #9751 Acetyl-Histone H3 (Lys9/Lys14) Antibody #9677 Rpb1 CTD (4H8) Mouse mAb #2629 Normal Rabbit IgG #2729
signal relative to input
30 25 20 15 10 5 0
signal relative to input
0.8 0 1 0.9 0.7 0.6 0.5 0.4 0.3 0.2 0.1 TCF4 (C48H11)
Rabbit mAb #2569 β-Catenin Antibody#9562
Normal Rabbit IgG #2729
PCR primers are specific for known TCF4 binding sites in the CAMK2D and c-Myc genes, and a region of the Myt1 gene that does not contain a TCF4 binding site. GAPDH SimpleChIP® GAPDH Competitor c-Myc SimpleChIP® c-Myc Competitor Myt1 SimpleChIP® Myt1 Competitor CAMK2D SimpleChIP® CAMK2D Competitor 1500 1000 700 500 400 300 200 100 1 2 base pairs
クロマチン免疫沈降法
酵素消化
vs.
ソニケーション処理
クロマチン免疫沈降(
ChIP
)法は、細胞のクロマチン中のタンパク質
–DNA
相互作用を調べる、強力かつ汎用性の高い手法で
す
(1,2)。この手法は、特定のゲノム領域と関係している複数のタンパク質の同定や、また逆に、ある特定のタンパク質に関係
している多くのゲノム領域の同定に使用されます
(3-6)。また、
ChIP
アッセイは、特定のタンパク質
–DNA
相互作用の空間的、
経時的関係を明確にするのにも使用されます。例えば、様々なタンパク質因子を遺伝子プロモーターに動員する順序の確定
や遺伝子の活性化における遺伝子座全体の特定のヒストン修飾の相対量を「測定」するのに使用できます
(3,4)。ヒストンタン
パク質の他、転写因子および転写補因子、
DNA
複製因子、
DNA
修復タンパク質の結合の解析にも使用可能です。
Background References1. Orlando V (2000) Mapping chromosomal proteins in vivo by formaldehyde-crosslinked-chromatin immunoprecipitation. Trends Biochem. Sci. 25(3), 99–104. 2. Kuo MH. et al. (1999) In vivo cross-linking and immunoprecipitation for studying dynamic Protein:DNA associations in a chromatin environment. Methods 19(3), 425–33. 3. Agalioti T. et al. (2000) Ordered recruitment of chromatin modifying and general transcription factors to the IFN-beta promoter. Cell 103(4), 667–78.
4. Soutoglou E. et al. (2002) Coordination of PIC assembly and chromatin remodeling during differentiation-induced gene activation. Science 295(5561), 1901–4. 5. Mikkelsen TS. et al. (2007) Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 448(7153), 553–60.
6. Lee TI. et al. (2006) Control of developmental regulators by Polycomb in human embryonic stem cells. Cell 125(2), 301–13.
7. Weinmann AS. et al. (2002) Identification of unknown target genes of human transcription factors using chromatin immunoprecipitation. Methods 26(1), 37–47. 8. Wells J. et al. (2002) Characterizing transcription factor binding sites using formaldehyde crosslinking and immunoprecipitation. Methods 26(1), 48–56.
ChIP
アッセイでは、細胞内のタンパク質
–DNA
相互作
用を固定、あるいは「保持」する可逆的なタンパク質
–
DNA
架橋剤であるホルムアルデヒドで細胞をまず固定
します(左図参照)
(1,2)。次に、細胞をすすぎ、ソニケ
ーションか酵素消化によりクロマチンを断片化して、
特定のタンパク質あるいはヒストン修飾に特異的な抗
体を用いてクロマチンを免疫沈降します。目的のタン
パク質やヒストン修飾に関連する
DNA
配列は、クロス
リンクしたクロマチン複合体の一部として共沈降し、
この過程で
DNA
配列の相対量が増加します。免疫沈降
後にタンパク質
–DNA
を脱クロスリンクし、
DNA
を精
製します。この
DNA
配列の濃縮物をさまざまな手法で
解析することができます。
酵素消化は、ソニケーションより穏やかにクロマチンを断片化しますので、クロマチンとエピトープを傷つけず、免疫沈降の効
率を向上させます。
免疫沈降効率の向上により、
DNA
に結合したタンパク質の検出能が高くなります。
標的
DNA
の検出効率の向上により、試料、時間、試薬の無駄を省けます。
酵素消化は、転写因子や補因子と結合した
DNA
を検出するのに不可欠な、低いバックグラウンドで高い感度を実現します。
酵素消化(SimpleChIP®
)したクロマチンを用いると、 ソニケーション処理(Competitor
)だけのクロマチンよりも良好な結果が得られます。 酵素消化(Lane 1
)とソニケーション法 (Lane 2
)を採用しているChIP
キットを 用いて、クロマチンを断片化しました。種交差性 希望納入価格
#9004S SimpleChIP
® Plus Enzymatic
Chromatin IP Kit (Agarose Beads) H, M,
R, Mk 101,000円 #9005S SimpleChIP
® Plus Enzymatic
Chromatin IP Kit (Magnetic Beads) H, M,
R, Mk 114,000円
SimpleChIP® Plus Enzymatic Chromatin IP Kit (Agarose Beads) #9004: Mouse
brain and mouse liver tissue were prepared and disaggregated into a single-cell suspension using a Dounce homogenizer and tissue disaggregator, respectively. The cells were then formaldehyde-crosslinked and chromatin was prepared and digested. DNA was purified and separated by electrophoresis on a 1% agarose gel and stained with ethidium bromide. The majority of chromatin from both brain (lane 1) and liver (lane 2) was digested to 1 to 5 nucleosomes in length (150 to 900 bp). 200 100 300 400 500 600 700 800 900 1000 1200 1500 1 2
組織サンプルでの競合比較
免疫沈降して得られた
DNA
のインプットクロマチンに対する値をグラフにしました。
#9005 SimpleChIP® Plus Enzymatic
Chromatin IP Kit (Magnetic Beads)
が、競合製品に比べてシグナルの増幅が顕著かつ高感度であることが実証されました。
CST
の新しい
SimpleChIP® Plus Enzymatic Chromatin IP
キットは、培
養細胞や組織サンプルの内在性レベルのタンパク質
–DNA
相互作用を検
出するのに最適です。
CST
はクロマチンの断片化に酵素消化を採用して
おり、キットには酵素消化
–
クロマチン免疫沈降
(
ChIP
)実験を、細胞や
組織サンプルで迅速かつ簡便に行うために必要な全ての試薬が含まれて
います(
ChIP
アッセイ
30
回分)
。また、ポジティブとネガティブコント
ロールも含まれ、プロテイン
G
アガロースビーズ(
#9004
)およびプロテ
イン
G
磁気ビーズ(
#9005
)のキットをご用意しています。
0.1 0.05 0.15 0.2 0.25Signal relative to input
Histone H3 (D2B12) XP® Rabbit mAb (ChIP Formulated) #4620 Tri-Methyl-Histone H3 (Lys4) (C42D8) Rabbit mAb #9751 Tri-Methyl-Histone H3 (Lys27) (C36B11) Rabbit mAb #9733 Acetyl-Histone H3 (Lys9) (C5B11) Rabbit mAb #9649 Rpb1 CTD (4H8) Mouse mAb #2629 Normal Rabbit IgG #2729
GAPDH RPL30 HoxA1 HoxD10
0 0.004 0.002 0.006 0.008 0.01 0.12
Signal relative to input
Ezh2 (D2C9) XP® Rabbit mAb #5246 SUZ12 (D39F6) XP®
Rabbit mAb #3737 RING1B (D22F2) XP® Rabbit mAb #5694 Normal Rabbit IgG #2729
GAPDH RPL30 HoxA1 HoxD10
0
SimpleChIP® Plus Enzymatic Chromatin IP Kit (Agarose Beads) #9004: Mouse
brain was prepared and disaggregated into a single-cell suspension using a Dounce homogenizer. The cells were then cross-linked with formaldehyde, and chromatin was prepared and digested. Chromatin immunoprecipitations were performed using the indicated ChIP-validated antibodies. Purified DNA was analyzed by quantitative real-time PCR using SimpleChIP® Mouse GAPDH Intron 2 Primers #8986, SimpleChIP® Mouse
RPL30 Intron 2 Primers #7015, SimpleChIP® Mouse HoxA1 Promoter Primers #7341, and
SimpleChIP® Mouse HoxD10 Intron 1 Primers #7429. The amount of immunoprecipitated
DNA in each sample is represented as signal relative to the total amount of input chromatin (equivalent to 1).
SimpleChIP® Plus Enzymatic Chromatin IP Kit (Magnetic beads) #9005 (A) compared to a competitor’s sonication-based Tissue Chromatin IP kit (B). Mouse liver was prepared and disaggregated
into a single-cell suspension using a tissue disaggregator. The cells were then cross-linked with formaldehyde, and chromatin was prepared and digested according to manufacturer’s recommendations. Chromatin immunoprecipitations were performed using the indicated ChIP-validated antibodies. Purified DNA was analyzed by quantitative real-time PCR using SimpleChIP® Mouse GAPDH Intron 2 Primers #8986, SimpleChIP® Mouse
AFM Intron 2 Primers #7269, SimpleChIP® Mouse HoxA1 Promoter Primers #7341, and SimpleChIP® Mouse HoxD10 Exon 1 Primers #7429. The amount of immunoprecipitated DNA in each sample is represented as
signal relative to the total amount of input chromatin (equivalent to 1).
0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4
Signal relative to input
GAPDH AFM HoxA1 HoxD10
0
Histone H3 (D2B12) XP®
Rabbit mAb (ChIP Formulated) #4620 Tri-Methyl-Histone H3 (Lys4) (C42D8) Rabbit mAb #9751 Tri-Methyl-Histone H3 (Lys27) (C36B11) Rabbit mAb #9733 Acetyl-Histone H3 (Lys9) (C5B11) Rabbit mAb #9649 Rpb1 CTD (4H8) Mouse mAb #2629 Normal Rabbit IgG #2729
0.005 0.01 0.015 0.02 0.025 0.03
Signal relative to input
Ezh2 (D2C9) XP® Rabbit mAb #5246 SUZ12 (D39F6) XP® Rabbit mAb #3737 RING1B (D22F2) XP® Rabbit mAb #5694 PCAF (C14G9) Rabbit mAb #3378 Normal Rabbit IgG #2729
GAPDH AFM HoxA1 HoxD10
0 0 0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4
GAPDH AFM HoxA1 HoxD10
Signal relative to input
Histone H3 (D2B12) XP®
Rabbit mAb (ChIP Formulated) #4620 Tri-Methyl-Histone H3 (Lys4) (C42D8) Rabbit mAb #9751 Tri-Methyl-Histone H3 (Lys27) (C36B11) Rabbit mAb #9733 Acetyl-Histone H3 (Lys9) (C5B11) Rabbit mAb #9649 Rpb1 CTD (4H8) Mouse mAb #2629 Normal Rabbit IgG #2729
0 0.005 0.01 0.015 0.02 0.025 0.03
GAPDH AFM HoxA1 HoxD10
Signal relative to input
Ezh2 (D2C9) XP® Rabbit mAb #5246 SUZ12 (D39F6) XP® Rabbit mAb #3737 RING1B (D22F2) XP® Rabbit mAb #5694 PCAF (C14G9) Rabbit mAb #3378 Normal Rabbit IgG #2729
A
SimpleChIP
®Plus Enzymatic Chromatin IP Kit (Magnetic beads) #9005
B
Competitor’s Sonication-based Tissue ChIP Kit
New
SimpleChIP
®
Plus
Enzymatic Chromatin IP Kits
細胞と組織サンプルに最適化した、詳細かつサンプル量に応じて試薬量を簡
単に調整できるプロトコールをご提供しますので、貴重な時間と試薬を無駄
にしません。
全ての試薬と適切な実験コントロールが含まれ、経済的です。
キットは
CST
の
ChIP
用抗体の規格試験で使用しており、最適化する手間を省け
ます。
キット(
#9005
)に含まれる
ChIP
グレードの磁気ビーズ(
#9006
)は、高感度
かつ低バックグラウンドなので、
ChIP-on-chip
や
ChIP-seq
に最適です。
キットを開発・製造している研究者による、的確かつ迅速な技術サポートを
ご提供いたします。
図
1. DNA
断片化の解析K:
腎臓, L1 & L2:
肝臓図
2.
抗体による標的領域の特異的な濃縮
使用製品
試験研究用
#9005
:
SimpleChIP® Plus Enzymatic Chromatin IP Kit
(
Magnetic Beads
)
CST
社キットを用いたマウス組織の
ChIP
アッセイ
結論
CST
社
SimpleChIP® Plus Enzymatic Chromatin IP Kit (Magnetic Beads) (#9005)
を用いて、組織サンプルから安定
したクロマチン断片化を行うことができた。また、
Histone H3
抗体によるゲノム
DNA
濃縮を確認した。キットに添付
の抗体とプライマーによって実験の成否を確認できるため、適切に実験系を構築できたか評価することができた。次回
以降も本キットを使用する予定である。
結論
結
結
資料提供:東京大学大学院 医学系研究科 疾患生命工学センター 健康環境医工学部門 相田 圭子先生、吉岡 亘先生プロトコール
マウス腎臓ならびに肝臓を用いて、
CST
社キット(
#9005
)に付属の抗
Histone H3
抗体とプライマーを用いて
ChIP
アッ
セイを行った。
1.
サンプル
C57BL/6
系統マウス腎臓ならびに肝臓(それぞれ
35 mg
)
2. ChIP
アッセイ
CST
社プロトコールに準じた。
Mince
した組織サンプルを入れる容器に関しては、プロトコール推奨の
15 mL
のコニカルチューブではサンプルの
ロスが大きいという問題が生じたため、
2 mL
チューブを用いた。核膜の破砕は、
Bioruptor
を用いて行い、破砕の
程度を光学顕微鏡で確認した。
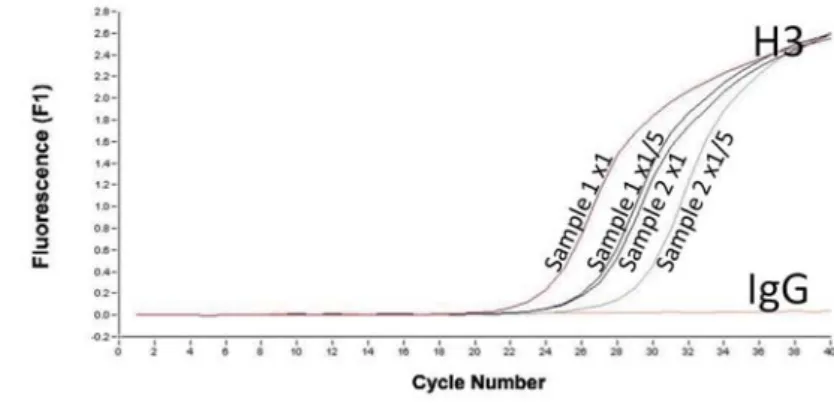
結果
クロマチンの断片化と濃度の分析を行ったところ、
100-500 bp
に断片化され(図
1
)、精製した
DNA
濃度は腎臓が
150
μg/mL
、肝臓は
250 μg/mL
であった。クロマチン断片化の程度はアッセイ間でばらつきが少なかった。
添付の抗体(ポジティブコントロール
Histone H3
とネガティブコントロール
Rabbit IgG
)で免疫沈降し、添付のプライ
マー
RLP30
で
qPCR
を行ったところ、ポジティブコントロール特異的な増幅がみとめられた(図
2
)。
プロ
ロ
プ
プ
結果
結
結
Micrococcal Nuclease
量
ChIP Result;
定量的
PCR
(
n=1
)
p53-binding p21 promoter
RPL30
20 18 16 14 12 10 8 6 4 2 0relative amount of DNA
(target/Input) p53WT p53KO α-p53 α-Histone lgG 20 24 18 16 14 12 10 8 6 4 2 0
relative amount of DNA
(target/Input) p53WT p53KO α-p53 α-Histone lgG HCT116細胞のp53WT細胞とp53KO細胞におけるp21 promoterへのp53のリクルートメントを検討した実験。 p53WT細胞では、p21 promoterへのp53の結合が認められる。p53KO細胞では、p53の結合が引き起こされていない。一方、 p53の結合がないRPL30遺伝子に対しては、どちらの細胞においてもp53の結合は観察されていない。なお、どちらの細胞
(p53WTとp53KO)および標的遺伝子(p21とRPL30)においても、 Histone H3抗体によるHistoneタンパク質の集積が認め
られ(Positive Control)、Normal Mouse IgGによっては、バンドが検出されていない(Negative Control)ことから、実
験はうまくいっていると思われる。 この検討により、DNAの断片化が成功してい ることを確認した。(p53KO-HCT116細胞) M: Size marker 0.5 μL:加えたMicrococcal Nucleaseの量 1000 (bp) M 0.5 μL 500 300 200 100
使用製品
試験研究用
#9005
:
SimpleChIP® Plus Enzymatic Chromatin IP Kit
(
Magnetic Beads
)
ヒト大腸がん細胞株
HCT116
を用いた
p53
の
ChIP
解析
結論
今回、内因性
p53
の標的遺伝子に対する結合活性を
SimpleChIP® Plus Kit
を用いて行った。
p53
を欠損した
p53KO-HCT116
細胞と対比させて検討することで、検出している
PCR
増幅に間違いがないことを確認できた。私たちは、本
Kit
が、
in vivo
サンプルにおいても使用可能であることを確認しており、
in vitro
、
in vivo
の用途を問わない点が優れてい
ると感じた。また、
ChIP
反応は、
Magnetic Beads
を用いて行うため、簡便で操作性もよく、非特異的産物を減少させ
ることができるためよかった。
結論
結
結
資料提供:熊本大学大学院 薬学教育部 遺伝子機能応用学分野 福田 亮介様、首藤 剛先生プロトコール
CST
社
#9005 SimpleChIP® Plus Kit
に添付の推奨プロトコールに従って行った。
※定量的
PCR
の方法、反応組成は異なる
1.
それぞれ
p53
遺伝子を有するまたは欠損するヒト大腸がん細胞株
p53WT-HCT116
または
p53KO-HCT116
から、
8
×
10
6個の細胞を準備し、
Micrococcal Nuclease
溶液
0.5 μL
を加え、
37
℃で
20
分間クロマチンを消化した。
2. DNA
を精製後、アガロースゲル電気泳動によりクロマチン
DNA
のサイズを確認した。
3.
クロマチン調製液(
2.6 μg/ChIP
)と
p53
に対するマウスモノクローナル抗体(
#2524S
)、
Histone H3
に対する抗体
(
Kit
に添付)
(
Positive Control
)および
Normal Mouse IgG
をそれぞれ
2 μg
を用いて
ChIP
を行った(
4
℃、
Rotation
下で、
14 hr
)。
4. p53
の標的遺伝子
p21
のプロモーター領域、
p53
の標的でない
RPL
遺伝子領域(
Negative Control
)に対するプライ
マーを用いて定量的
PCR
解析を行い、得られた結果をもとに、
Relative amount of DNA (Target/Input)
を算出し、
グラフを作成した。
結果
プロ
ロ
プ
プ
結果
結
結
酵素によるクロマチン断片化の最適化
クロスリンクされたクロマチン
DNA
を
150-900 bp
の長さに断片化するための最適条件は、使用する細胞数や組織量に
対する
Micrococcal Nuclease
の量に大きく依存します。下記は、特定の組織や細胞タイプでクロマチン断片化の最適
条件を決定するためのプロトコールです。
1.
CST
推奨プロトコール(
#9005
または
#9004
)の
Section I
、
II
、
III
に従い、組織
125 mg
あるいは
2
×
10
7個の細胞を用いてク
ロスリンクされた細胞核を調製してください。
Section III
のステップ
2.
を行った後、下記に進んでください。
2.
細胞核調製液
100 μL
を
1.5 mL
遠心用チューブ
5
本に分注し、氷上に置いてください。
3.
Micrococcal Nuclease 3 μL
を
1
×
Buffer B + DTT
混合液
27 μL
に加えてください(酵素を
10
倍に希釈します)。
4.
ステップ
2.
の各チューブに、希釈した
Micrococcal Nuclease 0 μL
、
2.5 μL
、
5 μL
、
7.5 μL
、
10 μL
を加え、チューブを数
回転倒混合し、
37
℃でよく混ぜ合わせながら、
20
分間インキュベートしてください。
5.
0.5 M EDTA 10 μL
を加えることで断片化を停止し、チューブを氷上に置いてください。
6.
13,000
rpm
、
4
℃で
1
分間遠心して細胞核を沈澱させ、上清を除去してください。
7.
細胞核を
1
×
ChIP Buffer + Protease Inhibitor Cocktail
混合液
200 μL
に再懸濁し、氷上で
10
分間インキュベートしてくだ
さい。
8.
溶解物を数回超音波処理し、核膜を破砕してください。超音波処理の合間は、サンプルを氷上で
30
秒間インキュベートして
ください。細胞核を完全に溶解するための最適条件は、超音波処理の前後に光学顕微鏡でサンプルを観察することで決定で
きます。
VirTis Virsonic 100 Ultrasonic Homogenizer/Sonicator
を
6
に設定し(
1/8-inch probe
)、
20
秒間の超音波処理
を
3
回行うことで、
HeLa
細胞核は完全に溶解されます。
Dounce homogenizer
で
20
回処理を行っても細胞核は溶解されま
すが、完全な溶解でない可能性があります。
9.
10,000
rpm
、
4
℃で
10
分間遠心し、溶解物を浄化してください。
10.
超音波処理した各溶解物
50 μL
を新しい遠心用チューブに移してください。
11.
各サンプル
50 μL
に、
nuclease-free water 100 μL
、
5 M NaCl 6 μL
、
RNAse A 2 μL
を加えてください。ボルテックスして、
37
℃で
30
分間インキュベートしてください。
12.
RNAse
A
で処理した各サンプルに、
Proteinase K 2 μL
を加えてください。ボルテックスして、
65
℃で
2
時間インキュベート
してください。
13.
100 bp DNA
マーカーとともに、各サンプル
20 μL
を
1%
アガロースゲルで電気泳動し、
DNA
フラグメントサイズを測定し
てください。
14.
DNA
サイズ
150-900 bp
(ヌクレオソーム
1-5
個)が得られた断片化の条件を確認してください。本プロトコールで目的サイ
ズの
DNA
フラグメントを産出した希釈した
Micrococcal Nuclease
の量は、目的サイズの
DNA
フラグメントを得るために
1
回分の免疫沈降調製液(分離組織
25 mg
あるいは
4
×
10
6個の細胞)に加える
Micrococcal Nuclease
ストック量の
10
倍とな
っています。例えば、本プロトコールで希釈した
Micrococcal Nuclease 5 μL
が
150-900 bp
の
DNA
フラグメントを産出し
たとすると、
CST
推奨
ChIP
プロトコールの
Section III
において、
0.5 μL
の
Micrococcal Nuclease
ストックを免疫沈降
1
回分
の調製液に加えてクロマチンを断片化することになります。
15.
目的サイズの
DNA
フラグメントが得られなかった場合は、
Micrococcal Nuclease
の量を調整しながら本プロトコールを繰
り返してください。または、クロマチン断片化の時間を調整することで
DNA
フラグメントサイズを最適化することができま
す。
SimpleChIP® Plus Enzymatic Chromatin IP
キットを
ChIP-seq
に使えますか?
はい、
#9005 SimpleChIP® Plus Enzymatic Chromatin IP Kit (Magnetic
Beads)
は、ChIP-seq
にご使用いただけます。#9005
に含まれるプロテインG
磁気ビーズ(
#9006
)にはsalmon sperm DNA
のようなDNA
ブロッキング試 薬を添加していないので、シークエンシングにおけるコンタミネーションの 心配はありません。なぜ酵素消化はソニケーションによるクロマチン断片化より優れ
ているのですか?
ソニケーションはよく使われているクロマチン断片化の方法ですが、出力の ばらつきやソニケーション中のクロマチンサンプルの乳化など、多くの問題 が確認されています。ソニケーションが不足するとクロマチンの断片化が不 十分ですし、やり過ぎやサンプルの乳化はタンパク質の変性と分解を引き起 こし抗体エピトープが失われてしまいます。CST
では培養細胞を用いて酵素消 化とソニケーションの比較実験を行い、酵素消化がソニケーションより穏や かにクロマチンを断片化し、ChIP
アッセイにおける免疫沈降効率を劇的に向 上させることを確認しました。組織サンプルを用いた検討でも、同様の結果 が得られています。細胞数や組織量によってクロマチン消化に使用する
Micrococcal
Nuclease
の量を調整する必要がありますか?
クロマチン調製に使用する細胞数あるいは組織量を測定することは大変重要 です。CST
では、各クロマチン免疫沈降反応1
回ごとに4
×10
6個の細胞あるい は25 mg
の組織を使用することをお薦めしています。組織サンプルは、酵素 消化前にクロスリンクして単細胞懸濁液にします。クロマチン消化に用いるMicrococcal Nuclease
量と細胞数の比率は、クロマチンを適正サイズ(150-900 bp
)に断片化するのに重要です。この比率は細胞や組織の種類によって 若 干 異 な り ま す が、4
×10
6個 の 細 胞( あ る い は25 mg
の 組 織 )に 対 し、Micrococcal Nuclease 0.5 μL
の割合がクロマチン消化に最適で、かつ再現性 の高いことが社内試験から分かっています。なぜ
Micrococcal Nuclease
によるクロマチン消化に加えソニケ
ーションが必要なのですか?
SimpleChIP® Plus Enzymatic Chromatin IP
キットに含まれるBuffer A
とBuffer B
は、ホルムアルデヒドでクロスリンクした細胞の細胞膜と核膜を完全 に可溶化せず、Micrococcal Nuclease
を細胞に入れてクロマチンを消化する ために細胞の透過性を高めているだけです。従って、クロマチンを溶液に放 出するため、膜を破砕するわずかなソニケーションが必要です。ソニケーシ ョンによってクロマチンをさらに断片化することが目的ではありません。適切に断片化されたクロマチンは、アガロースゲル電気泳動でど
のように観察されますか?
SimpleChIP® Plus Enzymatic Chromatin IP
キットのプロトコール「IV.
クロ マチンの断片化と濃度の分析(推奨ステップ)」に記載の通り、CST
ではクロマ チン消化後、IP
実施前にサンプルを分析しています。下図は、エチジウムブロ マイドで染色した1%
アガロースゲルにおけるクロマチンの理想的な泳動結果 です。レーン1
はDNA
マーカー、レーン2
は精製したクロマチンDNA
です。ク ロマチンDNA
は、モノ-
、ジ-
、トリ-
、テトラ-
、ペンタ-
ヌクレオソーム単位 (150-900 bp
)にせん断され、クロマチンDNA
濃度をOD
260で測定したところ、様々な細胞や組織で大体125-250 μg/mL
になりました。クロマチンの泳動に 問題がある場合は、プロトコールの「Appendix B
: クロマチン断片化の最適化」をご参照ください。*
注:もし150 bp
(モノヌクレオソーム)付近のシン グルバンドのみ観察される場合、クロマチンが過剰に 消化されています。細胞数あるいは組織量に対しヌク レアーゼ量が過剰なので、ヌクレアーゼ量を減らす か、細胞数あるいは組織量を増やしてください(6
ペ ージ参照)。各
IP
にどれくらいのクロマチンが必要ですか?
CST
では、全ての標的タンパク質に対し、IP
反応1
回につき4
×10
6個の細胞あ るいは25 mg
の組織で社内試験を始めています。これは10-20 μg
のクロマチ ンに相当します。ただし、ヒストンのIP
では、1
×10
6個相当の細胞、あるい は2.5-5 μg
のクロマチンでも可能です。一般的に、IP
を準備するとき、IP
反応 液500 μL
を調製するためにクロマチンを1
×ChIP Buffer
で希釈し、抗体と一 晩反応させます。しかし、CST
のプロトコールでは必ずしもクロマチンを希釈 する必要がなく、希釈していないクロマチンをIP
にご使用いただけます。組織サンプルで予想されるクロマチン産生量は?
組織サンプルからクロスリンクされたクロマチンを採取する場合、クロマチ ンの産生量は組織タイプによってかなり異なります。下記テーブルに、各組 織25 mg
(一番下のHeLa
細胞は4
×10
6個)から得られると予想されるクロマ チン産生量と予想されるDNA
濃度を掲載しました。各組織タイプにおいて、Medimachine (BD Biosciences)
とDounce Homogenizer
を用いた分離方 法でクロマチン産生量に大きな違いはありませんでしたが、組織分離にMedimachine
を用いた場合、Dounce Homogenizer
に比べてIP
効率が高く なりました。脳組織の場合、Medimachine
では適切に単細胞懸濁液にできな いので、Dounce Homogenizer
での分離を強くお薦めします。また、最適なChIP
の結果を得るため、各免疫沈降に5-10 μg
のクロスリンク、断片化され たクロマチンを使用することをお薦めします。 組織/細胞 総クロマチン産生量 予想されるDNA濃度 脾臓 組織25 mgにつき20-30 μg 200-300 μg/mL 肝臓 組織25 mgにつき10-15 μg 100-150 μg/mL 腎臓 組織25 mgにつき8-10 μg 80-100 μg/mL 脳 組織25 mgにつき2-5 μg 20-50 μg/mL 心臓 組織25 mgにつき2-5 μg 20-50 μg/mL HeLa 4×106細胞につき10-15 μg 100-150 μg/mLChIP
に適した抗体とは?
CST
でChIP
用に検証済みの抗体のデータシートには、IP
に必要な量が掲載さ れています。SimpleChIP® Plus Enzymatic Chromatin IP
キットと共に使用 される場合の抗体希釈率は、Recommended Antibody Dilutions
の下にご ざいます。また、社内試験で使用した抗体の正確な容量をChIP
データの説明 文に記載しています。CST
の社内試験では、4
×10
6個の細胞(10-20 μg
のク ロマチン)を用いて最適な抗体量を決定しています。 もし抗体がCST
でChIP
用に検証済みでない場合、ChIP
アッセイでの抗体性能 を保証できません。未検証の抗体をChIP
アッセイで試されたい場合、通常のIP
で検証済みの抗体を使用されることをお薦めします。また、ChIP
反応1
回に つき、2-5 μg
の抗体で始めることをお薦めします。磁気ビーズとアガロースビーズではどちらが良いですか?
SimpleChIP® Plus Enzymatic Chromatin IP
キットにはプロテインG
アガロ ースビーズ(#9004
)およびプロテインG
磁気ビーズ(#9005
)のキットをご用 意しています。両ビーズの比較実験では、どちらも低バックグラウンドで特 異的結合を示しています。アガロースビーズは従来よりIP
で用いられるビーズ ですが、磁気ビーズはより利便性の高いビーズです。磁気ビーズは磁石側の チューブ側面に付着するので、洗浄バッファーの吸引除去時にロスがなく、 遠心分離も必要ありません。ただし、#7017 6-Tube Magnetic Separation
Rack
などが必要です。磁気ビーズは
DNA
でブロッキングしていないため、ChIP-seq
にご使用いただ けます。アガロースビーズは、バックグラウンドを軽減するためにソニケー ションしたsalmon sperm DNA
でブロッキングしているため、ハイスループ ットシークエンシングにおいてブロッキングDNA
が配列読取りに影響を与え ます。 1500bp 1 2 1000bp 700bp 500bp 400bp 300bp 200bp 100bpよくあるお問い合わせ
SimpleChIP
®組織サンプルでも培養細胞でも!
酵素消化を採用した新
ChIP
キット
Over 140 ChIP Validated Antibodies
CST
は、他のアプリケーションと同様にChIP
用抗体にも厳格な品質基準を定め、厳しい規格試験を実施しています。最新の
ChIP
用抗体リストは、ウェブでご確認ください。
www.cstj.co.jp/technologies/chip.php
SimpleChIP® Control PCR Primers
2
種類のコントロールプライマーのミックスで、ChIP
アッセイで得 られるDNA
の増幅にご使用いただけます。これらのプライマーは、ChIP
アッセイに用いた抗体で検出した標的タンパク質の既知の結 合サイトを含む、ポジティブコントロールDNA
配列を増幅します。 また、抗体の感度を確認するために、ネガティブコントロールとし てもご使用いただけます。各抗体に対するポジティブとネガティブ コントロールプライマーの一覧は、ウェブでご確認ください。SimpleChIP® Assay Kits
複数の
ChIP
用抗体とコントロールPCR
プライマーミックスのセット で、多能性やエピジェネティック研究にご使用いただけます。キッ トには、厳格な品質管理と社内試験において、品質、バリデーショ ン、ロット間差について最高水準の基準を満たす抗体が含まれてい ます。さらに、ポジティブとネガティブのプライマーセットも含まれ ていますので、お客様の実験系で適切なコントロールとしてご使用 いただけます。最新の製品リストは、ウェブでご確認ください。 本パンフレット中の価格は2012年11月現在のもので、消費税は含まれておりません。製品の仕様、包装、価格等につきましては、予告なく変更される場合がありますのでご了承ください。 © 2012 Cell Signaling Technology, Inc. XP®, SimpleChIP®, and Cell Signaling Technology® are trademarks of Cell Signaling Technology, Inc.希望納入価格 (円)
#9004S SimpleChIP® Plus Enzymatic Chromatin IP Kit (Agarose Beads) 101,000
#9005S SimpleChIP® Plus Enzymatic Chromatin IP Kit (Magnetic Beads) 114,000
希望納入価格 (円)
#8980S SimpleChIP® Stem Cell Master Regulator Assay Kit 126,000
#8982S SimpleChIP® Human Bivalent Promoter Assay Kit 105,000
#8981S SimpleChIP® Mouse Bivalent Promoter Assay Kit 105,000
ChIP
関連の技術的問題は、製品の開発担当者が的確かつ迅速にサポートいたします。
New
SimpleChIP
®Plus
Enzymatic Chromatin IP Kits
■取扱店 ■輸入販売元