HoneybeeScience(1998)
ローヤルゼ リー タ ンパ ク質 の特徴 と機能
ローヤルゼ リ-は,健康食品 として現在広 く 利 用 され て お り,種 々 の薬 理 作 用 (藤 井, 1995)および栄養生理的作用 (中島,1994)が 報告 されている. このローヤルゼ リーの有効成 分 の研究 は,10-ヒ ドロキ シ-2-デセ ン酸を代表 とす る脂肪酸, ア ミノ酸, ビタ ミンなどの低分 子化合物を中心 になされてきた.一方, ローヤ ルゼ リーには通常10%以上 の タンパ ク質 が含 まれているが, この タンパク質の分子構造や生 理作用 な どに関す る研究 はお くれて い る. ま た, ローヤルゼ リー配合 の ドリンク剤などの製 造時には, アルコール抽 出が行われ, タンパ ク 質 は沈殿物 として分離 されるが, このタンパ ク 質の積極的な有効利用 は図 られていないのが現 状である. そこで著者 らは, ローヤルゼ リータンパ ク質 の分子構造や生理的機能 の解明な らびに ドリン ク剤製造時の副産物であるタンパ ク質の高度利 用 を 目的 と して研究 を行 って きた. したが っ て, ここでは著者 らが ローヤルゼ リーか ら単離 した分子量350,000および 55,000の糖 タンパ ク質を中心 に, その性質,構造な らびに培養動 物細胞 に対す る作用 について述べ る. またロー ヤルゼ リータンパ ク質の うち,唯一全 ア ミノ酸 配列が解明されているロイヤ リシンと呼ばれ る 抗菌性 タンパ ク質 について も触れる. ところで, ローヤルゼ リータンパ ク質 につい ての研究 は, まずゲル電気泳動 による分析 (八 並 ら,1987;Haれesand Simuth,1992)が行 われ,20成分以上 の タ ンパ ク質 甲存在 が報告 されている.他方では,各種溶媒を用 いた分画 (友田 ら,1974),さらにカラムクロマ トグラフ ィーによる分離精製 (Tomoda eta1.,1977;米倉 政実
竹中,1984;竹中 ・越後,1984)が試み られ, 竹中 (1984)は,2種の水溶性 タンパ ク質 (分 子量46,000および 55,000)および 2種 のアル カ リ可溶性 タンパ ク質 (分子量63,000および 30,000)の単離 に成功 した. そ して, これ ら4 種の タンパ ク質 は, いずれ も糖 タンパ ク質であ ること, またそれ らのア ミノ酸組成や等電点を 明 らかに している. しか し,その分子構造や生 理的機能 に関す る研究 はほとん ど行われていな い.Ⅰ.
ローヤルゼ リータ ンパ ク質 の特徴 1. ローヤルゼ リーは高 タンパク質 ローヤルゼ リーの化学成分 については古 くか ら多 くの報告があるが,各成分 の値 は研究者 に よりかな りまちまちであ った.そこで, ある一 定条件 の もとで採取 した ローヤルゼ リーを用 い て分析 した結果 (竹中,1982)によると,その タンパ ク質含量 は,中冨 お よび台湾産 の場合 9.3-12.5%, 日本産 の場合 9.5-14.1%であっ た.高 タンパ ク質で,完全食品 といわれている 采綱田ま, ローヤルゼ リ- と同程度 の水分含量 (約66%)であ り,タンパ ク質 は約 12%含 まれ ている. したが って, ローヤルゼ リーは鶏卵 と 同様高 タンパ ク質の食品 ということがで きる.2.
口-ヤルゼ リーのタンパク質成分 ローヤルゼ リーのタンパ ク質 は,水溶性の も の と水不溶性 の ものが あ り,前者 が全体 の約 75%を,後者 が約 25%をそれぞれ 占めてい る (竹中,1982).また,竹中 (1982)は水溶性 の タンパ ク質をゲル波過法 により分析 した結果, 分子量が約10万,約 9万 および 1万以下 の少・・.一 rLf
A一
一
・
一
磯 煮葦 二.
i
保持 時間ゲル電気泳動
(
SDS-PAGE)
高速ゲル漉過
図1 ローヤルゼリータンパク質のゲル電気泳動および高速ゲル減退による分析 な くとも3種類 の タ ンパ ク質 が含 まれて いる ことを報告 している. 著者 ら(1992)ち,ローヤルゼ リーの全 タン パ ク質を ドデシル硫酸 ナ トリウム-ポ リアク リ ルア ミドゲル電気泳動(
SDS
-
PAGE)
によ り分 析 した結果,分子量約 10万-1万 の タンパ ク 質が約20種類存在す ることがわか った. その 中で最 も量的に多 いのが分子量58,000の タン パ ク質であ り,次 いで多 いのが55,000の タン パ ク質であった (図 1).また高速ゲル臆過法 に よりローヤルゼ リーの水溶性 タンパ ク質を分析 した結果 (図 1),分子量約35万,約 12万,約 6万,約 4万および 1万以下の各 タンパ ク質成 分 が認 め られ,中で も最 も多 い成分 が約35万 のタンパ ク質であることも明 らかにな った.級 述す るよ うに, この分子量35万 の タ ンパ ク質 は,S
DS
-
PAGE
に お い て検 出 され た分 子 量 58,000の成分 と同一であ り,この タンパ ク質6 分子か ら構成 されているオ リゴマータンパ ク質 であることが明 らかにな った. また,分子量約6
万 の タ ンパ ク質 は,S
DS
-
PAGE
にお ける分 子量55,000の タンパ ク質成分 と同一であるこ とがわか った.3.
糖 タンパ ク質が多 いローヤルゼ リーS
DS
-
PAGE
を用 いて ローヤルゼ リー タンパ ク質を分離 したのちに,PAS
(過 ヨウ素酸 -シ ッフ試薬)染色法 により糖 タンパ ク質 の検 出を 試みたところ, ほとんどの タンパ ク質成分が染 色 され, ローヤルゼ リータンパ ク質 の大部分が 糖 タンパ ク質であることが明 らかになった. ま た, この結果 は レクチ ンを用いたアフイニテ ィ クロマ トグラフィーによ って も確認 された. ら ちろん,単純 タンパ ク質 も一部含 まれていた.Ⅱ. 分子量
350,
000
の糖 タ ンパ ク質
(
アピシン)
1
.
ローヤルゼ リータンパク質の分離精製 ローヤルゼ リーの水溶性 タ ンパ ク質 につ い て,その成分 を個 々に分離 し,純粋 な形で取 り 出す ことを,図2に示す方法 によ り行 った.ま ず, 生 ローヤルゼ リーか ら透析 によ り分子量1 万以下の低分子成分を除去 した水溶性 タンパ ク 質を,DEAE-セル ロファイ ンA-500カラムを 生ローヤルゼリー a析(分育分子J的 l万)遠
心
分
■ 水溶性l3-ヤルゼ リータンパ ク文 捨イオ ン交Jtクロマ トグラフィー (DEAE-セル ロ7 7インA-500) Dl(兼述 LJ)ii分 D3(吸暮)百分 「cIMlt'
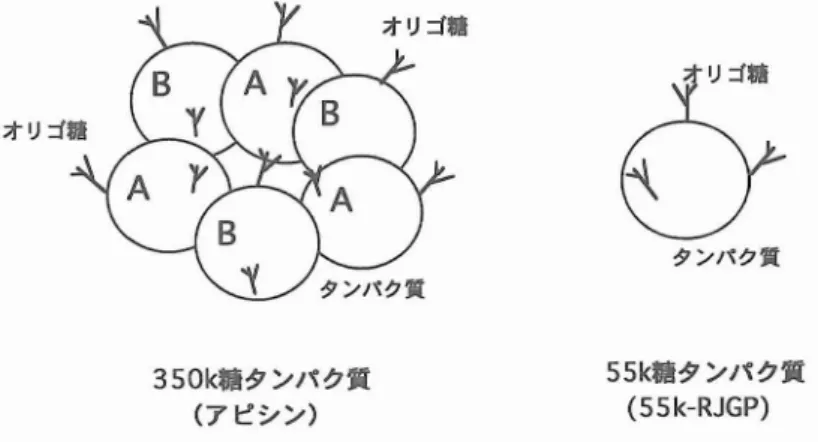
J諾 ,',n71 7,i.言,イ l(:;崇 ヲ ,イン臥 ,0..,0 55k一書タンパク文 350k一書タンパクJt (55k-RJGP) (アビシン) 図2 ローヤルゼリー糖タンパク質の分離精製法350k糖タンパク質 (ア ピシン) 55k楯タ ンパ ク笑 (SSk-RJGP) 図3 ローヤルゼ リーに含まれている350k糖 タンパク質 (アピシン)と55k糖タンパク質 (55k-RJGP)の構造 用 いた陰 イオ ン交換 クロマ トグラフィーによ り 分画 した. 次 に吸着画分 で あ るD3画分 を, セ ル ロ フ ァイ ンGCL1000sfカ ラ ムを用 いた ゲ ル癒過 に供 し,分子量35万 の タ ンパ ク質 を純 品 と して取 り出 した (米倉 ら,1992).なお,ミ ツバ チの学名 (APismellifera L.)にちなん で, この分子量35万 の タ ンパ ク質 を ア ピシ ン (apisin)と命名 した. 他方,前述 のDEAE-セル ロフ ァイ ンA-500 M8n81-2MBna1-6 MAnαl-2M&Jlal-3 カ ラムを用 いた陰 イオ ン交換 クロマ トグラフ ィ ーにおいて, カ ラムに吸着 しなか った素通 り画 分 で あ るDl画分 を,CM-セ ル ロ フ ァイ ン C-500カ ラムを用 い る陽 イオ ン交換 クロマ トグラ フ ィーにか け,分子量55,000の タ ンパ ク質 を 純 品 と して精製 した (伊豆川 ら,1995). これ ら2種 の糖 タ ンパ ク質 が,竹 中(1984) が単 離 した4種 の糖 タ ンパ ク質 の いず れ に該 当す るかを検討 したが, 断定 す るには至 らなか \ hhJlα1-6 / \
M&nβト4GIcNAcP1-4GIcNAc M9A :55% /
MAJln1-2MAnal-2M8JICLl-3
MAJlal-2M&Jla1-6
\ M&Jla1-6
/ \
M8nα1-3 MAAβ1-4GlcNAcβ1-4GIcNAc M8A I.5% /
M8Zlα1-2MJLJlαト2M&Aql-3
M8Jlal-2M■ncL1-6
\ MAna1-6
/ \
M&Jlα1-3 M&nβ1-4GIcNAcβ114GIcNAc M7A :22%
/
MAnal-2M8JICLl-3
M■皿α1-6
\ M8Jlα1-6
/ \
M&JICLI-3 Mwβ1-4GIcNAcβト4GIcNAc M5A :18%
/
MAnaト3
J< rr L r 八 < ・ E i 八 卜 ト ム F rl エ 八 日 t n Y 八 rr や 八 r l E i : 八 日 爪 ト ] ( ; H 卜 八 日 t i f i .< ・ r I 八 < ・ ) ・ 巳 八 < ・ y Ej '. y 八 { ・i Y 山 八 < ・ r l
J7
潜 入 仰 4 4 (J f 八 y 恥 Y < ・ 漣 八 叶 爪 i r Y h 八 日 # q r ト 八 日 爪 ト 図5 ローヤルゼリータンパク質 (アピシンおよび55k-RJGP)と牛乳 タンパク質 αS)-カゼインのアミノ酸組成の比較 った .2
.
ア ピシンの タ ンパ ク質化 学 的性質 と構 造 ア ピシンの分子量 を測定 した結果,高速 ゲル 減退 法 で は350,000,-万 SDS-PAGE法 で は 58,000であ った.したが って,ア ビシンは分子 量58,000のサ ブユニ ッ ト6個 か らな るオ リゴ マー タンパ ク質 であると推定 された. また, こ のサ ブユニ ッ トは, その ア ミノ酸組成分析 およ びN-末端 ア ミノ酸配列分析 の結果,2種類 (A およびBサ ブユニ ッ トとす る)であることがわ か った.よ って,現時点 で は図3に示すよ うに ア ピシ ンはA サ ブユニ ッ ト3個 と Bサ ブユニ ッ ト3個 か らな るへ テ ロオ リゴマ ー タ ンパ ク 質であると著者 ら(1995)は考 えてい る. また, ア ピシ ンの等電点 を求 めた ところ,約 4.4-5.4であ り, ア ピシ ンは酸性 タンパ ク質 で あることが明 らかにな った. 次 に, ア ピシン中のオ リゴ糖含量 を分析 した ところ, 約5.5%であ り, さ らにその糖鎖構造 を1H-NMRによ り解 析 した結 果, マ ンノー ス が5個 (M5A),7個 (M7A),8個 (M8A)な らびに9個 (M9A)それぞれ結合 したオ リゴマ ンノースか らな る典型的 な高 マ ンノース型糖鎖 (図 4) であることが明 らかにな った (Kimur -aeta1.,1995). ア ピシンのア ミノ酸組成 を牛乳 の タンパ ク質 であるαsI-カゼイ ンと比較 した ものが,図5で あるが,大部分 のア ミノ酸 では両 タンパ ク質 に おいてその含量 が類似 してお り, ア ピシ ンはカ ゼイ ンと同様,栄養価 の高 いタンパ ク質である といえ る. ただ し, ア ピシ ンで はアスパ ラギ ン 酸量 が多 く,逆 にカゼイ ンではグル タ ミン酸量 とプロ リン量 が多 く, この3種 のア ミノ酸含量 が両者で大 き く異 な っていた.3.
7 ピ シ ンの 培 養 動 物 細 胞 に対 す る作 用 ア ピシ ンが ミツバチに とって, あ るいは ヒ ト に対 して どのよ うな生理作用 を発揮す るかを解 明す る第一段階 と して, ア ピシ ンの培養動物細 胞 に対 す る作 用 を調 べ た (Watanabeeta1.,1996,1997).すなわち, 細胞 を シャー レの中 で培養 し, この際培地 にア ピシ ンを添加 して, 生 きている細胞 の数 を測定 した. その一例 を図 6に示すが, これは ヒ トの血球系単球細胞株 に ついての結果であ り, ア ピシンを添加す ると, 培養1日後で約 5倍 の細胞数 に増え,培養 日数 とともにさ らに少 しずつ増加 し,培養5日後 に は培 養 開始 時 の約7倍 に増殖 す ることが明 ら かにな った.一方, ア ピシンを添加 しない場合 で も,細 胞 の増殖 は起 こ り,培 養4日後 で約 3.5倍 に増殖 していたが, ア ピシソの有無で紬
( o o o J
oL
X ) 点 盛 男 胡 30 20 0 1 2 3 4 5 培 養 日 数 (日) 図6 ヒト血球系単球細胞の増殖に対するアピシンの影響 胞数 に大差が認め られた. このよ うに, アビシ ンは細胞の増殖 を強 く促進す る作用のあること が明 らかになった. このような培養細胞の増殖 に対す る促進作用 は,他 の ヒ ト単芽球様細胞株や と トー ヒト-イ ブ リドーマ株 などの ヒ ト細胞, チャイニーズハ ムスター細胞 な らびに昆虫 (ハスモ ンヨ トウ) 細 胞 にお いて も認 め られ た (関 島 ら,1995; Watanabeeta1.,1996).ただ し,細胞の種類 によって, ア ピシンの増殖促進作用の程度 には 差異があり,細胞 によってア ピシンに対す る応 答が異 なることがわか った. さらに, シロネズ ミ (ラッ ト)の肝臓細胞を 取 り出 し,同様 にシャー レの中で培養 し, ア ピ シンの添加効果を調べた (Fujiieta1.,1996). すなわち, ネズ ミ肝臓 の初代培養細胞 に対す る 作用を調べた結果,図7に示すように,ア ピシ ソを培地 に添加 した場合,培養1日後で約 2倍 の細胞数 に増 えたのち,20
日間の長期 にわた り,その細胞数の約9割を維持す る作用のある ことが明 らかになった,他方, ア ビシンを添加 しない場合で も,培養1日後 には約 1.8倍 の細 胞数 に増殖 したが,培養2日後か ら次第に細胞 が死滅 し,約1週間後 には培養開始時よりわず かに多 い くらいの細胞数 まで大 きく減少す るこ とがわか った. このことか ら, ア ピシンは肝臓細胞の増殖 を 促進す る作用 はさほど強 くないものの, いった ん増殖 した細胞が死ぬのを抑制 し,その寿命を 延ば していることがわか った.その作用 メカニ ズムについては現在のところ不明であるが,今 後の研究 により解明 されるものと期待 される.Ⅲ.
分 子 量 55,
000
の糖 タ ンパ ク質
(
55k-
RJGP)
1
. 55k-
RJGP
の性質 と構造 Ⅱ. 1. で述べた方法 により,精製 した分子 量55
,000の タンパ ク質 (以下55k-
RJ
GP
と略 す る) について,分子量 を測定 した ところ,高 速 ゲ ル 臆 過 法 お よ びSDS-
PAGE
法 と も に 55,000であった.したが って,本 タンパ ク質 は ア ピシンとは異 な り,サブユニ ッ ト構造を もた ないことが明 らかにな った (図 3).また,その 等電点 は約6.3-7.3のほぼ中性 にあ ることが わか った (伊豆川 ら,1995). また,55k-
RJ
GP
のア ミノ酸組成分析を行 っ た結果,図5に示すように,全体 としてアピシ ンに類似 してお り, α 51-カゼイ ンとはアスパ ラ ギ ン酸, グルタ ミン酸 およびプロ リンにおいて その含量 に差異がみ られたが, ア ミノ酸組成か0 0 2 4 6 8 10 12 14 16 18 20 培 養 日 数 (EI) 図7 ネズ ミ肝臓細胞に対するアビシンおよび55k-RJGPの影響 らみ た55k-RJGPの栄 養 価 はカゼ イ ンと同様 高 い もので あ った.なお,55k-RJGPのN-末端 ア ミノ酸配列 は, ア ピシ ンとは全 く異 な ってお り,本 タ ンパ ク質 が ア ピシ ンとは異 な るタ ンパ ク質 で あ ることを裏付 けた. さ らに,55k-RJGPの糖 含 量 を測 定 した結 果, 約2.8%で あ り, ア ピシ ンよ りも低 い値 で あ っ た. ま た, この オ リゴ糖 の 糖 鎖 構 造 を】 H-NMRお よ びMALDLTOF MSに よ り調 べ た結果,マ ンノース9個 か らな るオ リゴマ ンノ ース型 の糖鎖 (M9A,図 4)で あ り, ア ピシ ン と同様 高 マ ンノ ー ス型 で あ った (Kimuraet a1.,1996).しか し,ア ピシ ンで発見 されたマ ン ノース5個,7個 な らび に 8個 のオ リゴマ ンノ ース型糖鎖 は認 め られなか った. 2. 55k-RJGPの培養動物細胞 に対す る作用 ア ピシ ンと同様 に, ヒ トの血球系単球細胞株 を用 いて,その細胞増殖 に対 す る55k-RJGPの 添加効果 を調 べ た ところ,細胞増殖促進作用 が 認 め られた. しか し,55k-RJGPの場合, - ス モ ンヨ トウの培養細胞 で は細胞増殖促進作用 は ほ とん ど認 め られ なか った.概 して,55k-RJGP の細胞増殖促進作用 はア ピシ ンに比べ,弱 いよ うであ る. また, シロネズ ミの初代培養肝臓細胞 に対 す る55k-RJGPの効果 を調 べた結果 をEg17に示 すが, ア ピシ ンに比 べ若干 弱 い ものの,肝臓細 胞 の死 滅 を防 ぎ,20日間 にわ た って細 胞数 を 維持 す る効果 が認 め られた. このよ うに, ア ピシ ンとは全 く異 な る糖 タ ン パ ク質 で あ る55k-RJGPにつ いて も,ア ピシ ン と同様,培養動物細胞 の増殖 を促 した り,細胞 死 を防 ぎ,長生 きさせ る作 用 のあ ることが明 ら か にな った. しか し, その作用 はいずれ もア ピ シ ンよ りも弱 い もので あ った. Ⅳ .
分子量
5,
500
のタンパク質
(ロイセ リシン)
1
. ロイセ リシンの構造 と性質
ロイヤ リシ ン(royalisin)と名付 け られた分 子 量5,523の タ ンパ ク質 は,Fujiwaraら(1990) によ って発見 され, その全 ア ミノ酸配列 が決定 されて い る (Eg18).ロイヤ リシンは,51個 の ア ミノ酸 か らな る小 さな タ ンパ ク質 で,分子 内 に 3カ所S-S結合 があ り, 架橋 されて いる. ち ょうど ヒ トの イ ンシュ リン (51個 の ア ミノ酸 か らな り, 分子量5,800)とい うホルモ ンとほ ぼ同 じサイ ズの タ ンパ ク質 であ るが, イ ンシュ リンには血糖低下作用 が あ るのに対 して, ロイ ヤ リシ ンは抗菌作用 を もってお り,両者 は全 く 異 な った タ ンパ ク質 であ る. また, ロイヤ リシ ンに はオ リゴ糖 は結合 してお らず,単純 タンパ鞭 遷 51 分子量 ;5,523 図8 抗菌性タンパク質ロイヤリシンのアミノ顧酒己列 ク質である.
2.
ロイセ リシンの抗菌作用 ロイヤ リシ ンは抗菌性 タ ンパ ク質 であ り, Bifidobacterium , Clostridium , Corynebac -terium,Lactobacillus,Leuconostoc,Staph y-lococcus,Streptococcusなどのグラム陽性菌の 増殖 を強 く阻害 した(Fujiwaraeta1.,1990).またその抗菌力 は,菌種 によって は抗生物質 と 同 じくらいの強 さ (1〟M 以下) であることが 報告 されている. しか しなが ら,大腸菌 (Esc -herichiacolt),サルモネラ菌 (Salmonellai n-fantis)などの グラム陰性菌 に対 して は抗菌力 は認 め られなか った.なお, ロイヤ リシンは熱 安定性があ り,100℃で15分間加熱 して も,そ の抗菌力 は失われなか った. ロイヤ リシンは ミ ツバチが細菌の感染か ら身 を守 るための生体防 御物質の1つ と考 え られている . ところで, ローヤルゼ リーにはもともと抗菌 作用のあることが報告 されてお り,その主体 は 10-ヒ ドロキ シ-2-デセ ン酸 な どの脂肪酸 であ る (Yatsunamiand Echigo,1985).デセ ン 酸 は,大腸菌などのグラム陰性菌 に対 して抗菌 作用を示す. このように ローヤルゼ リー中のロ イヤ リシンが グラム陽性菌 に対 して,デセ ン酸 などの脂肪酸が グラム陰性菌 に対 してそれぞれ 抗菌作用を発揮す ることによって, ローヤルゼ リーは強い抗菌力を示す ことがで きるものと考 え られ る.
Ⅴ.
まとめ
ローヤルゼ リーのタンパ ク質 の うち,量的に 多 い2種の成分 を分離精製 し,そのタンパ ク質 化学的性質,構造および培養動物細胞 に対す る 作用を調べた.分子量350,000の糖 タンパ ク質 (アピシン) は, 6個 のサ ブユニ ッ ト (分子量 58,000)か らな るオ リゴマー タンパ ク質 であ り, その糖鎖 は高 マ ンノース型であった. ア ピ シンはヒ トなどの培養細胞 に対 して,強 い細胞 増殖促進作用 を示す一方, シロネズ ミ肝臓の初 代培養細胞では,細胞死 を抑制 し,長生 きさせ る作用があることが明 らか にな った. 分子量55,000の糖 タンパ ク質 (55k-RJGP) は,サブユニ ッ ト構造 を もたない中性 タンパ ク 質 で あ り, その糖鎖 は高 マ ンノース型 で あ っ た.55k-RJGPも培養動物細胞 に対 して増殖促 進作用を示 したが,その作用 はア ピシンよりも 弱 いものであった. また, シロネズ ミ肝臓の初 代培養細胞 に対 してア ピシンと同様 の効果が認 め られたが, ア ピシンよ り若干弱か った. Fujiwaraら(1990)によ って発見 された ロイ ヤ リシンは, グラム陽性菌 に対 して抗菌作用を 示す,分子量5,523の単純 タンパ ク質であった. ここで述べた著者 らの研究 は,茨城大学農学 部食品機能学研究室において,渡辺和彦 をは じ め とす る学生諸氏 とと もにな された もので あ る. また,農林水産省食品総合研究所篠原和毅引用文献
藤井畝 1995.ミツバチ科学 16(3):97-104. Fujii,M.,M.Yonekura,T.Higuchi,K.Mori
-mitsu,I.Yoshino,S, Mukai,T.Aoki,T. Fukunaga,Y.Inoue,M.Sato and J .Kana-eda.1996.FoodS°i.Techno1.,1nt.2( 4):223-225.
Fujiwara,S.,J.Imai,M,Fujiwara,T.Yaeshima, T.Kawashima and K.Kobayashi.1990.∫. Biol.Chem.265 (19):11333-11337.
Haれes,∫.and∫.Simuth.1992.∫.Apic.Res.31 (1):22-26.
伊豆川智美,本多創,藤井信,木村吉伸,金枝純,栄 倉政実.1995.生化学 67(7):615.
Kimura,Y.,N,WashinoandM.Yonekura.1995. Biosci.Biotech.Biochem.59 (3):507-509. Kimura.Y.,S.Kajiyama,J.Kanaeda,T.Izukawa
andM.Yonekura.1996. Biosci.Biotech.Bi o-chem.60(12):2099-2102.
中島将次.1994.New FoodIndustry36( 3):54-64. 関島善行,原敏夫,駕野憲之,米倉故実.1995.農化 69:133. 竹中哲夫.1982.ミツバチ科学 3(2):69-74. 竹中哲夫.1984.玉大農研報 24:10ト149. 竹中哲夫,越後多嘉志.1984. ミツバチ科学 5(1): 7-12. 友田五郎,松山悼,柴内昌子,矢崎恵理子.1974.玉 大農研報 14:86-96.
Tomoda,G.,∫.Matsuyama and M.Matsuka. 1977.J.Apic.Res.16 (3):125-130.
27:31-40.
米倉政実,渡辺和彦,斎藤学志,堤将和,篠原和毅, 木村吉伸,高木茂明.1992.生化学 64(8):816.
YoNEKURA,MASAMI.Characterizationandphys -iologicalfunctionofroyaljellyproteins.Hone y-bee Science(1998) 19(i):15122.Schoolof Agriculture,IbarakiUniverslty,3-21-1,Chuuou, Ami-machi,Ibaraki,300-0393Japan.
We purifled and characterized two glycoprot -einsfrom royaljellyofthehoneybeeApismeL -LiferaL Onewasaglycoprotein which had a molecularweightof350-kDaand consisted of sixsubunitswithamolecularweightof58-kDa. Forthisglycoprotein,we proposed the name apisin,ThestructuresofN-linkedsugarchains ofaplSlnWerefoundtofallintothecategoryof oligomannose-typesugarchains.Apisinstimula -tedthegrowthofhumanlymphocyticcellllneS inserum-freeconditions.Apisin wasalsoeffec
-tive in maintenance the high viability ofthe primary monolayerculture ofrathepatocytes for20daysinaserum-freemedium.
The other glycoprotein (55k-RJGP) was a slnglepolypeptidewlthamolecularmas sof55-kDaanditsル 1inkedsugarchainwasfoundto be a non-processed high mannose typesugar chain.The55k-RJGP alsohad theproliferation stimulating activityforhuman monocytesand maintainedthehighviabilityofratllVerPrima -ryculturedcell.