ATR-X 症候群モデルマウスにおけるスパイン形成異
常に関する神経薬理学的研究
著者
山口 航矢
学位授与機関
Tohoku University
学位授与番号
11301甲第18625号
URL
http://hdl.handle.net/10097/00125884
ATR-X
症候群モデルマウスにおける
スパイン形成異常に関する神経薬理学的研究
東北大学大学院 薬学研究科
薬理学分野 山口航矢
- 1 -
目次
要旨
--- 2
第一章 諸言
--- 3
第二章 方法
--- 7
第三章 結果
--- 18
第四章 考察
--- 31
謝辞
--- 35
参考文献
--- 36
発表論文リスト
--- 50
- 3 -
要旨
X 連鎖 α サラセミア精神遅滞症候群(ATR-X 症候群)は ATRX 遺伝子の変異 により発症する疾患である。Atrx 遺伝子のエクソン 2 を欠損させた ATR-X 症 候群のモデルマウスでは、認知機能障害や異常な樹状突起スパイン形成を含む ヒト知的障害患者でみられる症状に似たフェノタイプを示す。今回私たちは、 脳由来神経栄養因子(BDNF)などの神経栄養因子の活性化を促進することで 神経保護や神経新生作用を示す、シグマ 1 受容体(Sig-1R)の活性化を標的と した。私たちは Sig-1R の強力な活性化剤である SA4503 を処置することで、 ATR-X 症候群モデルマウス由来の培養皮質ニューロンでみられた軸索伸長や樹 状突起スパインの異常が改善することを示した。さらに SA4503 の投与は ATR-X 症候群モデルマウスで見られた認知機能の低下も改善した。私たちは ATR-ATR-X 症候群モデルマウスの内側前頭前野(mPFC)において BDNF のタンパク質量 が顕著に減少していることを発見した。さらには、SA4503 の投与により ATR-X 症候群モデルマウスで見られた BDNF タンパク質量の減少は回復した。これ らの結果から Sig-1R の活性化を介した樹状突起スパイン異常の改善が ATR-X 症候群の診断後治療に有効である可能性が示唆された。- 4 -
第一章 緒言
X 連鎖 α サラセミア精神遅滞症候群 (ATR-X 症候群) は ATRX 遺伝子の変 異により男性のみで発症する疾患である (Gibbons et al., 1992, 1995)。ATR-X 症 候群は重度の精神遅滞、α サラセミア (HbH 病)、特異な顔貌、外性器異常、骨 格異常、独特の行動・姿勢異常、てんかん発作などを含む様々な臨床症状により 定義されている (Gibbons et al., 2008)。日本国内では、約 100 症例が診断されて おり世界では日本の症例を含め約 500 症例以上が診断されている。発症頻度は 出生男児4−5 万人に 1 例、日本国内では年間 10 名前後の患者が発症していると 推定されている (Gibbons et al., 2008)。
ATRX 遺伝子は SWI/SNF (switch/sucrose nonfermentable) 様のクロマチンリモ
デリングタンパク質である ATRX をコードしており、ATRX タンパク質は二つ の特徴的な配列を有している。1 つは ATRX-DNMT3-DNMT3L (ADD) ドメイン と呼ばれる植物ホメオドメイン (plant homeodomain : PHD) であり、このドメイ ンはヒストン 3 (H3) 尾部、特に 4 番目のリシン残基がメチル化されておらず、 9 番目のリシン残基がジメチル化あるいはトリメチル化されているという修飾 部位 (H3K4me0K9me2/3) に結合する (Argentaro et al., 2007 ; Dhayalan et al., 2011 ; Iwase et al., 2011)。もう 1 つは 7 つのヘリカーゼサブドメインを含む小領域であ り、ATPase 活性を有する (Picketts et al., 1996; Mitson et al., 2011)。ATR-X 症候 群の患者に見出されてきた 80 以上の変異はそのほとんどが N 末端側にある ADD ドメインもしくは C 末端側にあるヘリカーゼ小領域に集中していること が知られている (Gibbons et al., 2006)。しかしながら ATRX タンパク質の機能に 関してはいまだ不明な点が多いのが現状である。
- 5 -
(Picketts et al., 1996 ; Gibbons et al., 1997 ; Villard et al., 1999)。エクソン 2 における 37 番目のアルギニンがストップコドンに変異している (R37X) ATR-X 症候群患 者では、リンパ芽球様細胞における ATRX タンパク質の発現量低下に付随して 比較的経度の知的障害が見られる (Howard et al., 2004; Abidi et al., 2005)。私たち は以前、エクソン 2 を欠損させた Atrx 変異マウス (AtrxΔE2 マウス) において、
ヒト ATR-X 症候群患者で見られる R37X 変異に対応する変異タンパク質が発 現していることを報告した (Nogami et al., 2011)。さらに AtrxΔE2 マウスでは 27
人の ATR-X 症候群患者で見られた様に (Mitson et al., 2011)、ATRX タンパク質 量が約 80% 減少していた (Nogami et al., 2011 ; Shioda et al., 2011, 2018)。また、 AtrxΔE2 マウスでは認知機能の低下や内側前頭前野 (medial prefrontal cortex : mPFC) における樹状突起スパインの形態異常が見られた (Shioda et al., 2011)。 知的障害で見られる特徴と一致して (Kaufmann and Moser, 2000 ; Levenga and Willemsen, 2012)、AtrxΔE2 マウスにおいても WT マウスに比べ長く細い樹状突 起スパインが見られた (Shioda et al., 2011)。 スパインは脳の神経細胞において興奮性シナプス入力を受信している部位で あり、刺激に応じたスパインの形態変化は、カルシウム動態・シナプス可塑性の 違いを反映しており、学習・記憶とも密接に関与している。これまでスパイン形 態とシナプス機能や可塑性が密接に関わっているということが数多く報告され ている。例えば、スパインのヘッドの大きさはシナプス後膜肥厚の領域やプレシ ナプスの活性領域へのシナプス小胞の結合数、ポストシナプスにおける受容体 数やそれに伴うシナプス電流の大きさ、シナプス強度に直接比例している (Matsuzaki et al., 2001 ; Holtmaat, et al., 2005 ; Noguchi et al., 2005 ; Kasai et al., 2010)。 樹状突起スパインの形態や機能のこれらの異常は、アルツハイマー病や統合失 調症、知的障害を含む認知機能障害と関係する神経疾患で報告されている (Fiala
- 6 -
et al., 2002 ; Kaufmann and Moser, 2000 ; Levenga and Willemsen, 2012 ; Forrest et al., 2018)。
Sigma-1 受容体 (Sig-1R) は、小胞体膜に局在する 2 回膜貫通型の非オピオ イド型の受容体である。小胞体の中でも特にミトコンドリアに接する領域 [mitochondrial-associated endoplasmic reticulum (ER) membrane (MAM)] に 豊富に存在し、IP3R と結合することで、ミトコンドリアへの Ca2+ 輸送を調節
する (Hayashi T and Su TP et al., 2007) などシャペロンとしての機能もあり、 様々なタンパク質の発現や機能に関与していることが報告されている (Su et al., 2010)。当研究室ではこれまで、Sig-1R のアゴニストである SA4503 が IP3
受容体を介したミトコンドリアへのCa2+ 流入、それに伴う ATP 産生を増加さ
せることを報告した (Hirano et al., 2014)。それだけではなく、siRNA による Sig-1R のノックダウンにより、ラット海馬培養ニューロンでは樹状突起スパイ ンの形態異常が生じること(Tsai et al., 2009)、SA4503 はラット海馬において BDNF タンパク質の発現増加を誘導することなど(Kikuchi-Utsumi and Nakaki, 2008)、Sig-1R がスパイン形成に関わることが報告されている。
脳由来神経栄養因子 (brain-derived neurotrophic factor : BDNF) は長期増強 (LTP) やそれに関連する樹状突起スパインの形態変化など、様々な型のニューロ ン可塑性に関わることが報告されている (Hedrick et al., 2016, Harward et al., 2016)。 今回私たちは、BDNF を含む神経栄養因子の活性化を促進することで、神経保 護や神経新生作用を示す Sig-1R の活性化を標的とした (Kourrich et al., 2012; Su et al., 2016)。BDNF は神経発達やシナプスの成熟、神経可塑性に重要な新経営用 因子である (Thoenen et al., 1995, Lewin et al., 1996, Horch et al., 2004)。BDNF 経 路の活性化はレット症候群や脆弱性 X 症候群などの知的障害モデルマウスに おける学習記憶能力の向上に有効である(Banerjee et al., 2012, Lauterborn et al.,
- 7 -
2007)。強力な Sig-1R の活性化剤である SA4503 の処置により (Matsuno et al., 1996)、AtrxΔE2 マウス由来の培養皮質ニューロンにおける軸索の発達異常、樹状 突起スパインの形態異常は改善した。これらの結果に一致して、AtrxΔE2 マウス
における認知機能障害は SA4503 の投与により回復した。神経発達やシナプス 成熟、シナプス可塑性と関係する重要な神経栄養因子である BDNF レベルが WT マウスに比べ AtrxΔE2 マウスの mPFC では減少していた。さらには、 AtrxΔE2 マウスへの SA4503 の投与は BDNF タンパク質レベルも顕著に改善 した。これらの結果から Sig-1R の活性化を介した樹状突起スパイン異常の改善 が ATR-X 症候群における認知機能障害への治療標的になる可能性が示唆され た。
- 8 -
第二章 方法
1. 実験動物
マウス (C57BL/6J) は標準的な餌と水を自由に与えられながら、温度調節下で 12 時間照明のもと飼育した。実験は国立大学法人東北大学動物実験等に関する 規程に則り行った。また AtrxΔE2 マウスは以前記載した (Nogami et al., 2011) 通
りに作製した。簡単に説明すると、まず Atrx ターゲッティングベクターを作製 するために Atrx 遺伝子の 2 kb EcoRV-SpeI 遺伝子断片と、5.5kb XhoI-BamHI 遺 伝子断片をそれぞれ短い相同領域、長い相同領域として使用した。スプライシン グアクセプター部位を有する IRES-beta-geo カセットを用いて、exon 2 を含む SpeI-XhoI 断片を置換した。ターゲティングベクターを J1 ES 細胞にトランス フェクトし、G418 耐性クローンを選択した。相同組換え体をサザンブロット解 析により同定した。AtrxΔE2 マウスを C57BL/6J 上で 6 世代戻し交配した。RT-PCR やイムノブロット、行動解析を行うために雄性マウスは 10-12 週齢で使用 した。 2. 初代培養 2.1 カバースリップの準備 カバースリップ 100 枚を 50 ml チューブに入れ、1N HNO3 を加えて 15 分間 超音波処理した。milliQ で 4 回洗浄した後、再び 15 分間超音波処理し milliQ で 1 回洗浄した。オートクレーブし、冷ました後 milliQ を捨てエタノール (99.5%) を加えた。エタノールごとシャーレに移し、カバースリップを一枚ずつアルミホ イル上で乾燥した。オートクレーブをかけ溶かしたパラフィン (Psraplast plus 88895002004) を、パスツールピペットを用いてカバースリップに 4 か所スポッ
- 9 -
トした。UV ランプで 1 時間滅菌した後、アルミホイルで包み保存した。 使用する前に 5 枚/60mm ディッシュに分け、それぞれのカバースリップに Poly-L-lysine 溶液 [Poly-L-lysine (SIGMA), 0.1M pH8.5 Borate Buffer)] (1mg/ml) 150 μl を重層した。一晩室温でインキュベートした後、Poly-L-lysine 溶液を除き 滅菌水 4ml を加え UV 下で 30 分間インキュベートした。滅菌水を除き、再び 滅菌水 4ml を加え UV 下で 3 時間インキュベートした。その後滅菌水を除き、 Neural-plating-MEM [Minimum Essintial Medium (Invitrogen) Penicillin-streptomycin (100units/100μg/ml), 10% FBS, 0.6% Glucose (Wako, Osaka, Japan), 1mM Pyruvic acid (Sigma-Aldrich, St. Louis, MO, USA)] を 4ml 加え使う日までインキュベーターで 保存した。
2.2 グリア培養
生後 1 日目 (P1) のラットの頭を切り、頭部から脳を取り出し、HBSS [1×Hank’s balanced salt solution (490ml), 1.0M HEPES buffer solution (5ml), Penicillin-streptomycin (100units/100μg/ml)] 入りシャーレに集めた。実体顕微鏡下で脳の裏 側から膜を除き、内側の膜と共に海馬も取り除いた。大脳皮質を取り出し HBSS 入り 60mm シャーレに集めた。HBSS を取り除き眼科用ハサミで大脳皮質を切 り刻み、HBSS を加えて全量 12ml になるように回収した。トリプシン (10×) 、 DNase1 を 1.5ml 加え、転倒混和し 37℃ の湯浴で 12 分間温めた。パスツール ピペットで 10 回程度ピペッティングし 3 分インキュベート後、上層をとり 5ml の FBS 入りの 50ml チューブにフィルターを通し集めた。残りの沈殿組織にト リプシン 1.5ml 、HBSS を加え 15ml に合わせた。37℃の湯浴で 12 分間インキ ュベート後、再びパスツールピペットで 10 回程度ピペッティングし 3 分インキ ュベート後、3ml の FBS 入り 50ml チューブにフィルターを通し集めた。それ
- 10 -
ぞれで集めた液を 1 本にまとめ、2 本の 15ml チューブに分けた。800g で 5 分 間遠心し、上清を除いた。適当量の Glial-MEM [Minimum Essential Medium (Invitrogen), Glucose (5g), Pyruvic acid (110mg), NaHCO3 (2.20g), 10mM HEPES buffer solution, Penicillin-streptomycin (100units/100μg/ml)] で底にたまった血のみ を懸濁し、取り除いた。2 匹/1 フラスコとなるように細胞を Glial-MEM で懸濁 し、20ml の Glial-MEM が入ったフラスコにまき培養した。DIV 1 に HBSS で 一度洗い、メディウムを交換した。DIV 4 に HBSS で一度洗った後、もう一度 HBSS を加えフラスコをたたき HBSS でさらに 2 回洗浄し、メディウムを交換 した。その後、週に 2 回メディウムを変え、2 週間ほど培養し実験に用いた。 2.3 神経細胞培養
胎生 18 日目 (E18) の wild-type (WT) マウスまたは Atrx マウスから大脳皮質 を取り出し、冷 HBSS 入りの 35mm ディッシュにうつした。実体顕微鏡を用い て脳の裏側から膜をはがした後、海馬を取り除き大脳皮質を分離した。15ml チ ューブに組織を集め、4.5ml になるように HBSS を加えた。トリプシンを 0.5ml 加え、転倒混和し 37℃の湯浴で 15 分間インキュベートした。上清を取り除き、 HBSS を 10ml になるように加え 5 分間インキュベートした。再び上清を取り除 き、HBSS を 10ml になるように加え 5 分間インキュベートした。上清を除き、 HBSS を 2ml 程加えパスツールピペットで 10 回程度ピペッティングした。その 後、先を熱して細くしたパスツールピペットを用いてもう一度 10 回程度ピペッ ティングした。少し静置し、上層の細胞数をカウントした。その細胞 100μl に 発現ベクターを 5μg NEPA21 Type II (NEPAGENE Co., Ltd. Chiba, Japan) を用い てトランスフェクションした。(条件としては、Poring Pulse… Voltage : 275V 、 Pulse Length : 0.5msec 、Pulse Interval : 50msec 、Number of Pulse : 2 回、Decay
- 11 -
Rate : 10% 、Polarity : + Transfer Pulse… Voltage ; 20V 、Pilse Length : 50msec 、 Pulse Interval : 50msec 、Number of Pulse : 5 回、Decay Rate : 40% 、Polarity : +/- で行った。) これを 4ml の Neural-plating-MEM が入った 35mm ディッシュ内 の Poly-L-lysine 処理済みのカバースリップに滴下した。3 時間後、カバースリ ッ プ を Neural-basal-MEM [Neurobasal Medium (Invitrogen), 1% Gluta MAX (Invitrogen), 2% B27 supplement (Invitrogen)] 4ml 入りの、グリアを培養していた 60mm ディッシュに移した。その 3 日後、グリアの成長を止めるために 5 μM になるように cytosine β-D-arabino-furanoside (Ara-C) (Sigma-Aldrich) を加え、そ れから週 2 回 Neural-basal-MEM でメディウムチェンジし、インキュベーター内 で培養した。皮質ニューロンは DIV5 または DIV24 に固定した。これらのタイ ムポイントはそれぞれ発達段階で軸索形成や樹状突起伸長が起こるステージ 4、 成熟段階でシナプス形成が起こるステージ 5 に分類される。軸索は樹状突起よ り 5-10 倍早く伸長することから、DIV3 以降軸索と樹上突起の同定が可能であ り、今回はより長い神経突起を軸索と同定した。 3. 薬物
SA4503 は過去に記載された方法 (Fujimura et al., 1997) で合成されたものを 使用した (in the Laboratory of Medicinal Chemistry, Zhejiang University)。SA4503 は 滅菌水により溶解させ、終濃度が 1μM になるようにそれぞれ DIV5, DIV24 ま で 48 時間培養皮質ニューロンに処置した。マウスでは WT または AtrxΔE2 マウ
スに生後 70 日から 84 日までの 2 週間 SA4503 (1mg/kg) または saline を腹腔内 に一日一回投与し、その後記憶関連行動を評価した。げっ歯類における学習記憶 に関する研究から、SA4503 は 1.0mg/kg またはそれ以下の濃度で効果的である ことが示されている (Maurice et al., 1997, Zou et al., 2000)。実験スケジュールの
- 12 -
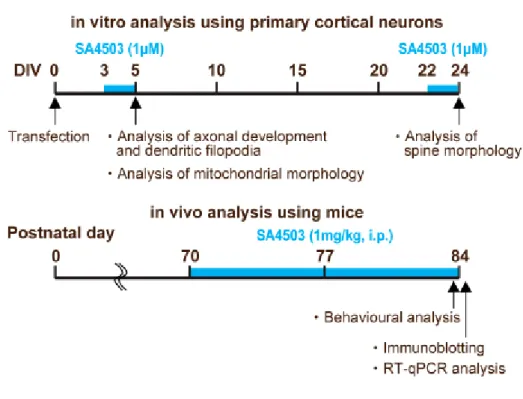
図 (図 1) を以下に記載した。
図 1. Diagrams of the experimental schedule. In in vitro analysis using primary cortical neurons, SA4503 (1 µM, dissolved in distilled water) was treated for 48 h in cultured cortical neurons in both DIV5 and DIV24. In in vivo analysis using mice, we administered SA4503 over the course of two weeks (1 mg/kg, intraperitoneally (i.p.) daily from postnatal day (P) 70 to P84) to AtrxΔE2 mice and subsequently assessed memory-related behaviors and biological studies.
4. 抗体とペプチド
免疫ブロット解析では次に示す 1 次抗体を使用した。
anti-BDNF antibody (1:1000, AB1779SP, Millipore, Burlington, MA, USA); anti-Sig-1R antibody (1:1000, ab53852, Abcam, Cambridge, UK);anti--Tubulin antibody (1:1000, T0198, Sigma-Aldrich).
- 13 -
また、免疫ブロットでは次に示す 2 次抗体を使用した。
goat anti-mouse IgG (H + L) and human ads-HRP (1:5000, 1031-05, SouthernBiotech, Birmingham, AL, USA); goat anti-rabbit IgG (H + L); and mouse/human ads-HRP (1:5000, 4050-05, SouthernBiotech).
免疫細胞化学では次に示す 1 次抗体を使用した。
living colors®Full-Length GFP polyclonal antibody (anti-GFP) (1:300, 632592, Clontech, Mountain View, CA, USA)
また、免疫細胞化学では次に示す 2 次抗体を使用した。
donkey anti-Rabbit IgG (H + L) highly cross-adsorbed secondary antibody; and Alexa Fluor 488 (1:500, A21206, Invitrogen)
5. 免疫ブロット
免疫ブロットは以前記載した (Shioda et al., 2010) 通りに行った。簡潔に説明 すると、まず組織をホモジナイズバッファー [50 mM Tris-HCl (pH 7.5), 0.5% Triton X-100, 0.15 M NaCl, 4 mM EDTA, 4 mM EGTA, 1 mM Na3VO4, 50 mM NaF, 1 mM DTT, 2 μg/ml pepstatin A, 1 μg/ml leupeptin, 100 nM calyculin A] 内で粉砕した。 等量のタンパク質を SDS-PAGE により電気泳動し、イモビロンフッ化ポリビニ リデンメンブレンにトランスファーした。5%の脱脂粉乳を含んだ TTBS 溶液 [50 mM Tris–HCl (pH 7.5), 150 mM NaCl, and 0.1% Tween 20] によりメンブレンを 室温で 1 時間ブロッキングをした後、メンブレンを 1 次抗体と共に 4℃で一晩イ ンキュベートした。メンブレンを洗浄した後、HRP 標識された適切な 2 次抗体 を 溶 解 し た TTBS 溶 液 と 共 に イ ン キ ュ ベ ー ト し た 。 バ ン ド は ECL immunoblotting detection system (Amersham Biosciences, NJ, USA)により増幅され、 luminescent image analyzer LAS-4000 (Fuji Film, Tokyo, Japan) で免疫活性バンドと
- 14 -
して可視化した後、Image Gauge version 3.41 (Fuji Film) を使って定量した。
6. 免疫細胞化学的、免疫組織化学的解析 免疫細胞化学的、免疫組織化学的解析は以前記載した (Shioda et al., 2010) 通 りに行った。簡潔に説明すると、まず 4% のパラホルムアルデヒド (PFA) を溶 かしたリン酸緩衝液 (PB) により固定した細胞または脳スライスを、0.1% の Triton X-100 を含んだ PBS で 10 分間処理した。サンプルを 1 次抗体 (EGFP antibody) と共に 4℃で一晩インキュベートした。翌日に PBS でサンプルを洗浄 した後、適切な 2 次抗体 [Alexa 488-conjugated donkey anti-rabbit (1:500; A-21206, Invitrogen)]. と 共 に イ ン キ ュ ベ ー ト し た 。 核 の 染 色 に は 4, 6-diamidino-2-phenylindole (DAPI) (Sigma-Aldrich) を 用 い た 。 蛍 光 画 像 は 共 焦 点 顕 微 鏡 (LSM700, Carl Zeiss,Oberkochen, Germany) を用いて解析した。
7. スパイン形態解析 スパインの頭の直径は、スパインの首に垂直な軸に沿って、スパインの頭のほ ぼ中央で測定した。スパインの長さは、スパインの頭の先端から樹状突起シャフ トまでの距離として計算した。スパインの直径は、スパイン頭部の最大寸法に対 応していた。スパイン頭部が不規則な形態を示した場合、またはスパイン頭部の 最大直径が首に対して垂直ではなかった場合、最大頭部直径を測定した。スパイ ン形態は以前に記載されている方法 (Irwin et al., 2002) に従って、未成熟型から 成熟型に分類した。測定は Image J (National Institutes of Health freeware) を用い て行った。
- 15 -
8. ミトコンドリア形態解析
ミトコンドリア形態解析は以前に記載されたとおりに行った (Wiemerslage et al., 2016) 。 簡潔 に 説明 す ると 、 細胞 に 終濃 度 350nM の MitoTracker™ Red CMXRos (M7512, Invitrogen) を 15 分間処置した後、4% PFA で細胞を固定した。 共焦点レーザー顕微鏡 (LSM700, Carl Zeiss, Oberkochen, Germany) を用いて画像 を取得した後、Image J software による解析した。Image J は各シグナルを測定 し、同時に各シグナルのピクセル領域、周囲長、最大長を測定した。これらの測 定 値 か ら roundness index を 次 の 式 で 計 算 し た [roundness index = 4π× (area/perimeter2)]。
9. RNA の抽出
RNA の抽出は RNeay Mini Kit (Qiagen) 用いて行った。WT マウスから摘出し、 -80℃で保存しておいた脳組織 (75mg) に buffer RLT (buffer RLT : 1ml + β-mercaptoethanol : 10μl) を 600μl 加えホモジナイズした。ホモジナイザーで 30 秒 間粉砕した後、20℃、15000rpm で 3 分間遠心し、上清を回収した。上清に同量 の 70% エタノール (DEPC 水で作成) を加えピペッティングした後、2ml コレ クションチューブ (添付) の中にセットした RNeasy カラムにアプライした。カ ラムの蓋を閉め、20℃、13000rpm で 15 秒間遠心した後ろ液を捨て、700μl の buffer RW1 を RNeasy スピンカラムに添加した。カラムの蓋を閉め、20℃、 13000rpm で 15 秒間遠心した後ろ液を捨て、500μl の buffer RPE [buffer RPE : エ タノール (96~100%) =1 : 4] を RNeasy スピンカラムに添加した。カラムの蓋 を閉め、20℃、13000rpm で 15 秒間遠心した後ろ液を捨て、500μl の buffer RPE を RNeasy スピンカラムに添加した。カラムの蓋を閉め、20℃、13000rpm で 2 分間遠心した後、カラムを新しい 2ml コレクションチューブに移した。蓋を閉
- 16 - め、20℃、15000rpm で 1 分間遠心し、RNeasy スピンカラムを新しい 1.5ml コ レクションチューブ (添付) にセットした。50μl の RNase フリー水をスピンカ ラム・メンブレンに直接添加し、蓋を閉め、20℃、15000rpm で 1 分間遠心した。 RNase フリー水の代わりに溶出液を用いてこの操作をもう一度行い、この溶出 液を RNA サンプルとし-80℃で保存した。 10. 逆転写 PCR (RT-PCR) と定量リアルタイム PCR (qPCR) 解析
Total RNA は RNeasy Mini Kit (Qiagen,Hilden, Germany) を用いてマウス脳か ら精製した。RNA は oligo (dT) primer (Promega) と Moloney murine leukemia virus-reverse transcriptase (Invitrogen) を用いて一本鎖 cDNA に逆転写し、定量 リアルタイム PCR (qPCR) を行った。また qPCR は iQ SYBR Green Supermix 2× (Bio-Rad)、48-well plates (Mini Opticon real-time PCR system, Bio-Rad) を用いて以 前記載した通り (Shioda et al., 2012) に行った。遺伝子発現は GAPDH を用いて 標準化した後、比較 Ct 法により評価した。また、コントロールとの比は 2-ΔΔCt により算出した。下に示した遺伝子特異的プライマーを今回は使用した。
< 定量リアルタイム PCR 用プライマー >
Mouse Bdnf in exon IX (FW) (50-AAGGACGCGGACTTGTACAC-30) Mouse Bdnf in exon IX (RV) (50-CGCTAATACTGTCACACACGC-30) Mouse Gapdh (FW) (50-TGTGTCCGTCGTGGATCTGA-30)
- 17 -
11. 行動解析
マウスには Y 字迷路試験 (Y-maze task)、新規物体認識試験 (novel object recognition task)、バーンズ迷路試験 (Barnes maze task) を含む行動試験を行った。 すべての行動分析のためのビデオテープは、遺伝子型と処置方法を知らない訓 練を受けた観察者によって解析された。Y 字迷路試験では黒いプレキシグラス でできた 3 本の同一アーム (50×16×32 cm3) からなる装置を使用し、Y 字型迷路 における自発的交替行動を空間学習記憶の指標として評価した。交替行動は 3 本 のアームすべてに連続して進入することとして定義された。最大交替数は、アー ムに進入した総数から 2 を引いたものとして定義され、交替の割合は実際の交 替数/最大交替数×100 として計算した。セッション中のアームへの総進入回数に つ い て も 測 定 し た 。 新 規 物 体 認 識 試 験 で は オ ー プ ン フ ィ ー ル ド ボ ッ ク ス (35×25×35 cm3) を使用した。獲得試行では同じ材料、形の二つの物体をチャン バーの中心に対称的に置き、マウスを 10 分間探索させた。24 時間後、一つの物 体を新規の物体に置き変え、マウスを 5 分間探索させた。物体への探索行動は、 対象物の上にのぼる、または立ち上がること、1cm 未満の距離で物体のにおいを 嗅いだり鼻を触れること、またはその両方と定義した。空間的新規性の識別は、 新規および既知物体への探索的接触数と両方への合計接触数との差を比較する ことによって評価され、これにより合計探索接触の違いを比較することを可能 にした。バーンズ迷路試験では、周囲に沿って20個の穴 (穴径 : 5cm) を有す る円形プラットフォーム (直径 : 92cm) からなる装置を使用した。各試験の開 始時には、迷路中央においた円筒形の開始チャンバー (7.5cm) 内にマウスを置 いた。10 秒後、チャンバーを取り除き、マウスに迷路上を自由に探索させた。 試験はマウスがゴールの穴に進入する、または 3 分が経過することで終了した。 マウスがゴールボックス内に進入した後すぐに、マウスをボックス内に 1 分間
- 18 - 静かに滞在させた。マウスは 4 日間 1 日に 2 回試験を行った。ゴールボックス へ逃げ込むまでにかかった探索時間とエラー数をテストパラメータとして記録 した。エラー数はゴールボックスにつながっている穴以外の穴に鼻や頭を近づ けた回数として定義した。 12. 統計処理 すべてのデータは、平均値±標準誤差で算出した。2 実験間における比較は独 立 Student’s t-test により検定処理した。多群間における統計上有意な差に関し ては one-way または two-way ANOVA と post-hoc Tukey tests によって検討し た。p<0.05 をもって統計上有意差ありと判定した。
- 19 -
第三章 結果
SA4503 の処置は AtrxΔE2 マウス由来培養皮質ニューロンにおける軸索の発達
異常や樹状突起フィロポディア異常を改善する
私たちはシナプス形態の変化を AtrxΔE2 マウス由来皮質培養ニューロンを用
いて検討した。皮質ニューロンはそれぞれ軸索形成や軸索決定 (ステージ 2-3)、 樹状突起伸長 (ステージ 4) シナプス新生や成熟 (ステージ 5) の神経成長ス テージに対応する DIV 1、DIV 5、DIV 14 以降に回収した (Dotti et al., 1988; van Spronsen et al., 2013)。軸索の成長は樹状突起に比べ 5-10 倍早いため、DIV 3 以 降には軸索と樹状突起の判別が可能であり、より長い神経突起を軸索と同定し た (Ahnert-Hilger et al., 2004)。さらに、マウスにおける樹状突起スパインの形成 は出生後早期に樹状突起フィロポディアから生じると考えられている (Hering and Sheng, 2001; Holtmaat and Svoboda, 2009)。軸索や樹状突起フィロポディアを 解析するために、私たちは DIV 0 に EGFP をトランスフェクションすることで 培養細胞をラベルし、DIV 5 にその形態を解析した。初めに、Atrx 遺伝子変異が 軸索伸長に及ぼす影響を検討した (Fig. 1A, upper panels)。その結果、WT ニュー ロンに比べ AtrxΔE2 マウス由来のニューロンでは軸索の長さが顕著に減少して
いた (Fig. 1B) (The axon length; WT + vehicle, 453.5 ± 13.3 μm; WT + SA4503, 445.5 ± 15.1 μm; AtrxΔE2 + vehicle, 300.1 ± 14.5 μm; AtrxΔE2 + SA4503, 419.6 ± 22.3 μm)。
樹状突起スパインは樹状突起フィロポディアを元とするが、私たちは AtrxΔE2
マウスニューロンではこれらの形態発達に変化があるのではないかと仮説を立 てた。WT マウスに比べ、AtrxΔE2
マウス由来の培養ニューロンではフィロポデ ィア数が有意に増加していたが、SA45034 を処置することでこれらの異常は改 善した (Fig. 1A, middle and lower panels and Fig. 1C) (The number of filopodia per 20
- 20 -
μm dendritic length; WT + vehicle, 2.1 ± 0.3; WT + SA4503, 2.1 ± 0.4; AtrxΔE2 + vehicle,
7.7 ± 0.4; AtrxΔE2 + SA4503, 3.8 ± 0.4)。
SA4503 の処置により AtrxΔE2 マウス由来皮質培養ニューロンにおける樹状突
起スパインの形態異常は改善する
シナプス形成を検討するために、EGFP をトランスフェクションした皮質ニュ ーロンを DIV 21 で回収した。これらのニューロンを固定した後、抗 EGFP 抗 体で染色することでスパイン形態解析を行った (Fig. 2A)。AtrxΔE2 マウス由来の
培養ニューロンでは WT に比べ、異常に細く、長いフィロポディア様のスパイ ン数が顕著に増加していた。AtrxΔE2 マウスニューロンでは長いフィロポディア
様のスパインの割合が増加しているのを示すように、長さの分布が WT に比べ より長い右側にシフトしていた。しかし、SA4503 の処置により AtrxΔE2 マウス
ニューロンで見られたこれらのスパイン異常は改善した (Fig. 2B and 2C) (The spine length; WT + vehicle, 0.68 ± 0.01 μm; WT + SA4503, 0.76 ± 0.01 μm; AtrxΔE2
+ vehicle, 1.02 ± 0.01 μm; AtrxΔE2 + SA4503, 0.56 ± 0.01 μm)。またスパインの頭の大
きさに関しても、AtrxΔE2 マウスでは WT ニューロンに比べ有意に減少してい
た が、SA4503 の処置によりこの異常は改善した (Fig. 2D) (The spine head diameter; WT + vehicle, 0.49 ± 0.01 μm; WT + SA4503, 0.49 ± 0.01 μm; AtrxΔE2 +
vehicle, 0.36 ± 0.01 μm; AtrxΔE2 + SA4503, 0.41 ± 0.01 μm)。予想通り、AtrxΔE2 マウ
スでは WT ニューロンに比べ未成熟スパイン数が増加し、成熟スパイン数が減 少していた。しかしながら、総スパイン数に関しては変化が見られなかった (Fig. 2E) (The number of mature spines per 20 μm dendritic length: WT + vehicle, 14.4 ± 0.5; WT + SA4503, 13.9 ± 0.3; AtrxΔE2 + vehicle, 3.3 ± 0.2; AtrxΔE2 + SA4503, 14.2 ± 0.4), (The number of immature spines per 20 μm dendritic length: WT + vehicle, 3.6 ± 0.4;
- 21 -
WT + SA4503, 4.4 ± 0.3; AtrxΔE2 + vehicle, 15.9 ± 0.4; AtrxΔE2 + SA4503, 4.3 ± 0.4)。
SA4503 の投与により AtrxΔE2 マウスにおける記憶、認知障害は改善する
げっ歯類での学習記憶の研究において、SA4503 は効果的な投与量として 1.0mg/kg またはそれ以下の容量で一般的に使用される (Maurice and Privat, 1997; Zou et al., 2000)。そのため、私たちは AtrxΔE2 マウスに対し SA4503 を 1mg/kg で 2 週間 (生後 70-84 日) 腹腔内投与し、その後記憶関連行動を検討した。まず は新規物体認識試験を行ったが、同じ形の物体を使ったトレーニングではどの マウスにおいても違いは見られなかった (Fig. 3A)。しかし 24 時間の保持期間の 後に違う形の物体を用いて行うテスト試行では WT に比べ AtrxΔE2 マウスで は新規物体への識別指数が有意に減少していた。この AtrxΔE2 マウスにおける 識別能力の低下は、溶媒投与群に比べ SA4503 の投与群で有意に改善していた (Fig. 3B) (The discrimination index in test session: WT + vehicle, 0.21 ± 0.05; WT + SA4503, 0.22 ± 0.03; AtrxΔE2 + vehicle, -0.06 ± 0.06; AtrxΔE2 + SA4503, 0.15 ± 0.04)。 Y 字迷路試験において、AtrxΔE2 マウスではアームへの総進入回数に変化は見ら れなかったが、交替率は WT マウスに比べ有意に減少していた。しかし、SA4503 の投与によりこれらの異常は有意に改善した (Fig. 3C and 3D) (The percentage of alternation behaviors: WT + vehicle, 73.2 ± 3.5; WT + SA4503, 73.7 ± 1.9; AtrxΔE2 + vehicle, 49.1 ± 4.0; AtrxΔE2 + SA4503, 75.9 ± 4.0)。Barnes 迷路試験では、目的の穴 に入るまでの時間を測定するが、WT マウスでは 4 日間の試験により徐々に穴 に入るまでの時間が減少していき、正常に認知行動が向上していることを示し ている。対照的に AtrxΔE2 マウスでは WT に比べ穴に入るまでの時間が遅く、
認知機能が向上していないことがわかった。しかしながら SA4503 の投与によ り、これらは有意に改善した (Fig. 3E) (The latency time in Day 4: WT + vehicle,
- 22 -
20.6 ± 1.0 seconds; WT + SA4503, 25.7 ± 3.1 seconds; AtrxΔE2 + vehicle, 64.8 ± 7.7 seconds; AtrxΔE2 + SA4503, 24.8 ± 2.8 seconds)。
SA4503 の投与は AtrxΔE2 マウスの mPFC における BDNF タンパク質量を
増加させる
Sig-1R はストレスやリガンドに調節されている小胞体シャペロンタンパク質 であり、細胞膜に脂質やタンパク質をシャトルしている (Su et al., 2010)。Sig-1R は神経伝達物質受容体やイオンチャネルを調節することで神経保護や神経可塑 性、神経伝達物質の放出など様々な過程に関与している (Kourrich et al., 2012; Su et al., 2010)。特に Sig-1R の活性化はラット初代培養アストロサイト (Malik et al., 2015) やマウス線条体 (Francardo et al., 2014)、ラット神経芽細胞腫 B104 細胞 (Fujimoto et al., 2012) において BDNF の分泌を促進する。そのため、私た ちは AtrxΔE2 マウスで BDNF の発現量に変化があるのかどうかを検討した。そ
の結果、AtrxΔE2 マウスの mPFC では mRNA 量の変化なしに、BDNF のタンパ
ク質発現量が減少していることがわかった。さらに、SA4503 の投与が AtrxΔE2
マウスの mPFC でみられた BDNF タンパク質の減少を改善することもわかっ た (Fig. 4A and 4B)。しかし Sig-1R の発現量に違いはみられなかった (Fig. 4B)。 Sig-1R はチャイニーズハムスター卵巣細胞 (CHO 細胞) (Hayashi and Su, 2007) や神経芽細胞腫 Neuro 2A 細胞 (Shioda et al., 2012) においてカルシウムをミト コンドリアに輸送し、ATP 合成に働くことが知られており、また心筋細胞では SA4503 の処置によ りミトコンド リア形 態が変化する ことが 知られている (Tagashira et al., 2013)。そこで私たちは、ミトコンドリア特異的なマーカーであ る Mito Tracker Red を用いて DIV 5 の培養皮質ニューロン軸索におけるミト コンドリア形態を解析した。しかしながら、どの群においてもミトコンドリアの
- 23 -
長さ、どれだけ丸いかを表す指標である roundness index に違いは見られなかっ た (Fig. 4D)。これらの結果は、培養皮質ニューロンにおいては Atrx の枯渇また は Sig-1R の刺激がミトコンドリアの分解やミトコンドリア融合の促進に影響 を与えないことを示している。
- 24 -
Figure 1. Treatment with SA4503 reverses abnormality of axonal development and dendritic filopodia in cultured AtrxΔE2 (Atrx mutant mice lacking exon 2) neurons. (A) Representative images of EGFP-transfected cortical neurons at DIV5. Neurons are stained for anti-GFP. Images in the middle and bottom panels are enlarged from the corresponding boxed areas. Scale bars: top panels, 50 µm; middle panels, 10 µm; bottom
- 25 -
panels, 5 µm. (B) Total axonal length. ** p < 0.01, versus vehicle-treated WT (wild type) neurons; ## p < 0.01, versus vehicle-treated AtrxΔE2 neurons by one-way ANOVA with post-hoc Tukey’s test; F (3, 76) = 17.4; n = 20 neurons for each group. The experiments were repeated three times with similar results. (C) Data show the number of filopodia per 20 µm dendritic length. ** p < 0.01, versus vehicle-treated WT neurons; ## p < 0.01, versus vehicle-treated AtrxΔE2 neurons by one-way ANOVA with post hoc Tukey’s test; F (3, 76) = 48.26; n = 20 neurons for each group. The experiments were repeated three times with similar results. Each bar represents the mean ± SEM. Abbreviations: Veh., vehicle; SA, SA4503.
- 26 -
Figure 2. Treatment with SA4503 ameliorates dendritic spine abnormality in cultured AtrxΔE2 neuron. (A) Representative images of EGFP-transfected cortical neurons at DIV21. Neurons are stained for anti-GFP. Images in the bottom panels are enlarged from each dendrite. Scale bars: top panels, 100 µm; bottom panels, 10 µm. (B) Relationship between cumulative percentage of spines and spine length. p < 0.01 in WT versus AtrxΔE2
- 27 -
neurons and in AtrxΔE2 versus SA4503-treated AtrxΔE2 neurons by the Kolmogorov– Smirnov test. (C) Data show the number of spines. ** p < 0.01 versus vehicle-treated WT neurons; ## p < 0.01, versus vehicle-treated AtrxΔE2 neurons by one-way ANOVA with post hoc Tukey’s test; F (3, 1494) = 89.38. (D) Data show the spine-head diameter. ** p < 0.01 versus vehicle-treated WT neurons; ## p < 0.01, versus vehicle-treated AtrxΔE2 neurons by one-way ANOVA with post hoc Tukey’s test; F (3, 76) = 76.38. (E) Spine protrusions per 20 µm dendritic length. ** p < 0.01 versus vehicle-treated WT neurons; ## p < 0.01, versus vehicle-treated AtrxΔE2 neurons by two-way ANOVA with post hoc Bonferroni’s test; F (3, 228) = 2.549, p = 0.0566 (group); F (2, 228) = 1008, p < 0.01 (spine protrusions), F (6, 228) = 243.6, p < 0.01 (interaction between group and spine protrusions); n = 20 neurons for each group. The experiments were repeated three times with similar results. WT, n = 360 spines; SA4503-treated WT, n = 367 spines; AtrxΔE2, n = 384 spines; SA4503-treated AtrxΔE2, n = 370 spines. Each bar represents the mean ± SEM. Abbreviations: Veh., vehicle; SA, SA4503.
- 29 -
Figure 3. SA4503 treatment decreased cognitive deficits in AtrxΔE2 mice based on memory-related tests. (A) Number of total arm entries (left) and alternations (right) in a Y-maze test. ** p < 0.01, versus treated WT mice; ## p < 0.01, versus vehicle-treated AtrxΔE2 mice by one-way ANOVA with post hoc Tukey’s test; F (3, 18) = 12.98. (B) Novel object recognition task. Discrimination index of object exploration during the sample phase (left), and while exploring familiar and new objects in the test phase after 24 h (right). ** p < 0.01, versus treated WT mice; # p < 0.05, versus vehicle-treated AtrxΔE2 mice by one-way ANOVA with post hoc Tukey’s test; F (3, 30) = 8.898. (C) Barnes maze test measurement of total latency (s) to reach the target hole. * p < 0.05 by two-way ANOVA with post hoc Bonferroni’s test; F (3, 120) = 6.535, p < 0.01 (group); F (3, 120) = 25.16, p < 0.01 (day), F (9, 120) = 1.184, p = 0.3112 (interaction between group and day). Each bar represents the mean ± SEM. Abbreviations: Veh., vehicle; SA, SA4503. WT + Vehicle: n = 9 mice, WT + SA4503: n = 9 mice, AtrxΔE2 + Vehicle: n = 8 mice, AtrxΔE2 + SA4503: n = 8 mice.
- 30 -
Figure 4. Treatment with SA4503 increases BDNF protein level in AtrxΔE2 mouse brain. (A) Representative immunoblot of mouse medial prefrontal cortex lysates probed with the indicated antibodies (top panel). Densitometry analysis of indicated proteins to β-tubulin (arbitrary units; (A.U.)) (bottom panel). * p < 0.05, versus vehicle-treated WT mice; ## p < 0.01, versus vehicle-treated AtrxΔE2 mice by one-way ANOVA with post hoc Tukey’s test; F (3, 16) = 13.7 in BDNF; n = 5 mice each. (B) Real-time RT-qPCR showing
- 31 -
total BDNF mRNA (using exon IX primers) expression in mouse medial prefrontal cortex lysates. BDNF mRNA were determined relative to Gapdh mRNA. n = 5 mice for each group. The experiments were repeated two times with similar results. (C) Representative images of cultured cortical neurons at DIV5 with the mitochondrial-specific marker, MitoTracker Red. Images in the right panels are enlarged from the corresponding boxed areas. Scale bars: left panels, 50 µm; right panels, 3 µm. (D) Data show the mitochondrial length (left) and the roundness index (right); n = 20 cells each. Abbreviations: Veh., vehicle; SA, SA4503.
- 32 -
第四章 考察
この論文で私たちは AtrxΔE2 マウスでシナプス形態や行動レベルで起こる知 的障害に関した異常が、生後 Sig-1R を活性化させることで改善することを示し た。さらに私たちは AtrxΔE2 マウスの mPFC における BDNF タンパク質レベ ルについても Sig-1R の活性化で増加することを明らかにした。これらの結果か ら、大人の ATR-X 症候群モデルマウスにおけるシナプス形態や行動異常に対 する薬理学的な改善効果が明らかとなり、さらには生後の Sig-1R の活性化が ATR-X 症候群を含む知的障害の様々な衰弱症状に対する治療戦略になり得る 可能性が示唆された。 樹状突起スパイン形態はシナプス伝達 (Matsuzaki et al., 2001) やシナプス形成 (Holtmaat et al., 2005)、スパイン頭部から樹状突起へのカルシウム拡散に関与し ていることが報告されている (Noguchi et al., 2005)。さらに、スパイン頭部の大 きさとシナプス強度の間には正の相関があると共に、スパインの形態変化は学 習記憶と関係している (Kasai et al., 2010)。樹状突起スパインは様々な形態をと ることが知られており、その構造は filopodia、thin、stubby、masuroom 型を基に 分類される (Hering and Sheng, 2001, Holtmaat and Svoboda, 2009)。マウス脳にお いて微小なスパイン (fulopodhia、thin) は神経活動時に迅速に構造変化し、消失 す るか 大きなス パインへと 成 長する (Maletic-Savatic et al., 1999, Engert and Bonhoeffer et al., 1999, Trachtenberg et al., 2002, Grutzendler et al., 2002)。微小なス パインは主に短命であり、脆弱なシナプスや機能していないサイレントシナプ スでよく見られる (Rochefort and Konnerth et al., 2012)。これらの知見は微小スパ インの形態変化が適応過程や学習過程の根底あることを支持している (Kasai et al., 2003)。これとは逆に、マウス皮質では巨大なスパイン (stubby、mashroom) は- 33 -
一ヶ月以上 (Trachtenberg et al., 2002) または一年 (Grutzendler et al., 2002) とい う長い期間安定で生存することから、記憶は脳において構造形成という形で長 い期間維持されていると考えられている。微小スパインが増加するというスパ インの異常と知的障害との関係性はダウン症や脆弱性 X 症候群、レット症候群 などでこれまで報告されている (Kaufmann and Moser et al., 2010, Levenga and Willemse et al., 2012)。今回、私たちは DIV 5 の AtrxΔE2 マウス由来培養皮質ニ ューロンで軸索伸長の異常や樹状突起フィロポディアの増加が起きていること を明らかにした (Fig. 1)。さらに私たちは、DIV 21 の AtrxΔE2 マウス由来皮質ニ
ューロンが WT に比べ長く、細い樹状突起スパインを持つことを示した。この 結果は in vivo における結果と一致しており (Shioda et al., 2011)、これらの結果 は AtrxΔE2 マウスにおける樹状突起スパインの未成熟形成が生後の脳の発達過
程で生じるものであることを示唆している。興味深いことに、短期間 (48 時間) SA4503 を処置することで、AtrxΔE2 マウス由来培養皮質ニューロンで見られた 軸索異常や樹状突起スパインの未成熟化が DIV 5、DIV 21 のどちらでも改善し たのである (Fig. 1 and Fig. 2)。また、in vivo における SA4503 の短期間投与 (生 後 70 日から 84 日) でも AtrxΔE2 マウスで見られた認知機能の低下が改善した (Fig. 3)。これらの結果は樹状突起の成熟が ATR-X 症候群における診断後治療 に有効である可能性を示唆しており、成熟したニューロンでもスパインはその 大きさや形態が可塑的で変化しうることを示唆している。この樹状突起スパイ ンの可塑的な性質がおそらく知的障害における認知機能治療の鍵となる重要な 要素だと考えられる。 Sig-1R は樹状突起スパインの発達や安定性に重要な役割を果たしている。 siRNA による Sig-1R のノックダウンにより、ラット海馬培養ニューロンでは 樹状突起スパインの形態異常が生じる (Tsai et al., 2009)。加えて SA4503 はラッ
- 34 -
ト海馬において BDNF タンパク質の発現増加を誘導する (Kikuchi-Utsumi and Nakaki, 2008)。神経芽細胞腫 B104 細胞において SA4503 は単に BDNF の総量 を増やすのではなく、タンパク質分泌にかかわる BDNF タンパク質の翻訳後過 程を促進する (Fujiwara et al., 2012)。さらに Sig-1R のノックダウンは成熟 BDNF の 放 出 を 減 少 さ せ る が 、 BDNF の 合 成 に 関 し て は 影 響 を 与 え な い (Fujiwara et al., 2012)。BDNF は樹状突起や軸索の伸長、樹状突起スパイン密度 の増加、シナプス可塑性の増強を促進することがよく知られている (Thoenen et al., 1995, Lewin and Barde et al., 1996, Horch et al., 2004)。ヒト BDNF 遺伝子にお ける一塩基多型には 66 番目のバリンがメチオニンに置換したもの (Val66Met) が存在するが、この変異により BDNF の輸送や活性依存的放出に障害が生じる ことで認知機能障害が引き起こされる (Egan et al., 2003)。さらには、Val66Met ノックインマウスの海馬歯状回顆粒細胞において樹状突起の複雑性が減少する ことが報告されている (Chen et al., 2006) ことから、BDNF の適切な分泌輸送が 樹状突起スパインの成長や可塑性に重要であると考えられている。今回私たち は、AtrxΔE2 マウスの mPFC では BDNF タンパク質量が減少していること、そ して SA4503 を投与することで mRNA レベルの変化無しに BDNF タンパク 質量が回復することを明らかにした。AtrxΔE2 マウスの脳内において BDNF タ ンパク質量の減少が生じる分子メカニズムはいまだ不明であるが、今回の結果 から Sig-1R の活性化による BDNF の放出増加が AtrxΔE2 マウスで生じるシ
ナプス形態や行動異常を回復させることが明らかになった。
Sig-1R のアゴニストは Sig-1R をミトコンドリア関連小胞体膜 (MAM) から 解離させることで細胞内の様々な場所に再分布させることが知られている (Su et al., 2016)。このアゴニスト誘導性の Sig-1R の再分布は、ラットの脳卒中モデ ル (Ruscher et al., 2011) やパーキンソン病モデルマウスである 6-ヒドロキシド
- 35 -
パミン障害マウスにおいて BDNF を含む生体分子の細胞内輸送の増加に関わ っている (Francardo et al., 2014)。興味深いことに、坑うつ薬であるイミプラミン による Sig-1R の活性化によって、BDNF 誘導性の PLCγ/IP3/Ca2+ 経路の活性化
が起こり、皮質ニューロンからのグルタミン酸の放出が引き起こされるのであ る (Yagasaki et al., 2006)。その他の Sig-1R 選択的アゴニストである PRE084 [2-(4-morpholinethyl)1 phenylcyclohexanecarboxylate] は神経剥離損傷ラットにおい てアストロサイトによる GDNF の発現を増加させる (Penas et al., 2011)。Sig-1R アゴニストの原型であるペンタゾシンは Sig-1R 活性化坑うつ薬であるイミプ ラミンやフルボキサミンと同様、PC 12 細胞において神経栄養因子 (nerve growth factor : NGF) 誘導性の神経突起の出芽を促進する (Takebayashi et al., 2002) 。また PC 12 細胞において、Sig-1R は上皮増殖因子 (epidermal growth factor : EGF) シ グ ナ ル を 活 性 化 す る こ と で 神 経 突 起 生 成 を 促 進 す る (Takebayashi et al., 2014)。このような Sig-1R を介した様々な栄養因子の相乗効 果が、AtrxΔE2 マウスにおける樹状突起スパインや認知機能の障害の回復と関係 しているのかもしれない。 ATR-X 症候群と BDNF との相互作用における適切な分子メカニズムを特定 するためにさらなる研究が必要であるが、結論として今回の結果から Sig-1R の 活性化が ATR-X 症候群における様々な神経症状や行動異常に対し、改善効果 を発揮することが示された。顕著な治療効果に加え、これらの結果は知的障害に おけるスパインの形態異常の主要な、そして潜在的な原因になり得るという根 拠を支持している。知的障害におけるスパインとシナプス異常に関係した臨床 での表現系を考えることで、私たちは介入治療への形成異常の原因解明や治療 ターゲットの同定ができるだろう。
- 36 -
謝辞
本研究を進めるにあたり、終始ご甚大なるご指導、ご鞭撻を賜りました当研究 室教授 福永浩司先生、熊本大学 発生医学研究所 ゲノム神経学分野 准教 授 塩田倫史先生に謹んで感謝の意を表します。 また、有益なご助言と貴重なご協力を頂きました薬理学分野 講師 森口茂 樹先生と同分野 助教 矢吹悌先生、篠田康晴先生、これまでの研究でご協力い ただきました当教室の皆様に心から御礼申し上げます。 最後に、本論文を御高閲賜りました東北大学大学院 薬学研究科 生活習慣 病治療薬学分野 教授 平澤典保先生、衛生化学分野 准教授 野口拓也先生 に深く感謝致します。- 37 -
参考文献
Gibbons RJ, Suthers GK, Wilkie AO, Buckle VJ, Higgs DR.
X-linked alpha-thalassemia/mental retardation (ATR-X) syndrome: localization to Xq12-q21.31 by X inactivation and linkage analysis.
Am J Hum Genet. 51, 1136-1149. (1992)
Gibbons RJ, Picketts DJ, Villard L, Higgs DR.
Mutations in a putative global transcriptional regulator cause X-linked mental retardation with alpha-thalassemia (ATR-X syndrome).
Cell. 80, 837-845. (1995)
Gibbons RJ, Wada T, Fisher CA, Malik N, Mitson MJ, Steensma DP, Fryer A, Goudie DR, Krantz ID, Traeger-Synodinos J.
Mutations in the chromatin-associated protein ATRX. Hum Mutat. 29, 796-802. (2008)
Argentaro A, Yang JC, Chapman L, Kowalczyk MS, Gibbons RJ, Higgs DR, Neuhaus D, Rhodes D.
Structural consequences of disease-causing mutations in the ATRX-DNMT3-DNMT3L (ADD) domain of the chromatin-associated protein ATRX.
- 38 -
Dhayalan A, Tamas R, Bock I, Tattermusch A, Dimitrova E, Kudithipudi S, Ragozin S, Jeltsch A.
The ATRX–ADD domain binds to H3 tail peptides and reads the combined methylation state of K4 and K9.
Hum Mol Genet, 20, 2195-2203. (2011)
Iwase S, Xiang B, Ghosh S, Ren T, Lewis PW, Cochrane JC, Allis CD, Picketts DJ, Patel DJ, Li H, Shi Y.
ATRX ADD domain links an atypical histone methylation recognition mechanism to human mental-retardation syndrome.
Nat Struct Mol Biol, 18, 769-776. (2011)
Picketts DJ, Higgs DR, Bachoo S, Blake DJ, Quarrell OW, Gibbons RJ.
ATRX encodes a novel member of the SNF2 family of proteins: mutations point a common mechanism underlying the ATR-X syndrome.
Hum Mol Genet. 5, 1899-1907. (1996)
Mitson M, Kelley LA, Sternberg MJ, Higgs DR, Gibbons RJ. Functional significance of mutations in the Snf2 domain of ATRX. Hum Mol Genet. 20, 2603-2610. (2011)
- 39 -
Gibbons RJ, Bachoo S, Picketts DJ, Aftimos S, Asenbauer B, Bergoffen J, Berry SA, Dahl N,Fryer A, Keppler K, Kurosawa K, Levin ML, Masuno M, Neri G, Pierpont ME, Slaney SF, Higgs DR.
Mutations in transcriptional regulator ATRX establish the functional significance of a PHD-like domain.
Nat Genet 17, 146–148. (1997)
Villard L, Bonino MC, Abidi F, Ragusa A, Belougne J, Lossi AM, Seaver L, Bonnefont JP, Romano C, Fichera M, Lacombe D, Hanauer A, Philip N, Schwartz C, Fontés M. Evaluation of a mutation screening strategy for sporadic cases of ATR-X syndrome. J Med Genet 36, 183–186. (1999)
Howard MT, Malik N, Anderson CB, Voskuil JL, Atkins JF, Gibbons RJ.
Attenuation of an amino-terminal premature stop codon mutation in the ATRX gene by an alternative mode of translational initiation.
J Med Genet 41, 951-956. (2004)
Abidi FE, Cardoso C, Lossi AM, Lowry RB, Depetris D, Mattéi MG, Lubs HA, Stevenson RE, Fontes M, Chudley AE, Schwartz CE.
Mutation in the 5’ alternatively spliced region of the XNP/ATR-X gene causes Chudley-Lowry syndrome.
- 40 -
Nogami T, Beppu H, Tokoro T, Moriguchi S, Shioda N, Fukunaga K, Ohtsuka T, Ishii Y, Sasahara M, Shimada Y, Nishijo H, Li E, Kitajima I.
Reduced expression of the ATRX gene, a chromatin-remodeling factor, causes hippocampal dysfunction in mice.
Hippocampus. 21, 678-687. (2011)
Shioda N, Beppu H, Fukuda T, Li E, Kitajima I, Fukunaga K.
Aberrant calcium/calmodulin-dependent protein kinase II (CaMKII) activity is associated with abnormal dendritic spine morphology in the ATRX mutant mouse brain.
J Neurosci. 31, 346-358. (2011)
Shioda N, Yabuki Y, Yamaguchi K, Onozato M, Li Y, Kurosawa K, Tanabe H, Okamoto N, Era T, Sugiyama H, Wada T, Fukunaga K.
Targeting G-quadruplex DNA as cognitive function therapy for ATR-X syndrome. Nat Med. 24, 802–813. (2018)
Kaufmann WE, Moser HW.
Dendritic anomalies in disorders associated with mental retardation. Cereb Cortex 10,981–991. (2000)
Levenga J, Willemsen R.
Perturbation of dendritic protrusions in intellectual disability. Prog. Brain Res. 197,153–168. (2012)
- 41 -
Matsuzaki M, Ellis-Davies GCR, Nemoto T, Miyashita Y, Iino M, Kasai H.
Dendritic spine geometry is critical for AMPA receptor expression in hippocampal CA1 pyramidal neurons.
Nat. Neurosci. 4, 1086–1092. (2001)
Holtmaat AJGD, Trachtenberg JT, Wilbrecht L, Shepherd GM, Zhang X, Knott GW. Transient and persistent dendritic spines in the neocortex in vivo.
Neuron. 45, 279–291. (2005)
Noguchi J, Masanori M, Ellis-Davies GCR, Kasai H.
Spine-neck geometry determines NMDA receptor-dependent Ca2+ signaling in dendrites. Neuron. 46, 609–622. (2005)
Kasai H, Fukuda M, Watanabe S, Hayashi-Takagi A, Noguchi J. Structural dynamics of dendritic spines in memory and cognition. Trends Neurosci. 33, 121-129. (2010)
Fiala JC, Spacek J, Harris KM.
Dendritic spine pathology: cause or consequence of neurological disorders? Brain Res Brain Res Rev. 39, 29-54. (2002).
Forrest MP, Parnell E, Penzes P.
Dendritic structural plasticity and neuropsychiatric disease. Nat Rev Neurosci. 19, 215-234. (2018)
- 42 -
Kourrich S, Su TP, Fujimoto M, Bonci A.
The sigma-1 receptor: roles in neuronal plasticity and disease. Trends Neurosci. 35, 762-771. (2012)
Hayashi T and Su TP.
Sigma-1 receptor chaperones at the ER-mitochondrion interface regulate Ca2+ signaling and cell survival.
Cell. 131, 596-610. (2007)
Su TP, Hayashi T, Maurice T, Buch S, and Ruoho AE.
The sigma-1 receptor chaperone as inter-organelle signaling modulator. Trends Pharmacol Sci. Elsevier Ltd. 31, 557–566. (2010)
Hirano K, Tagashira H, Fukunaga K.
Cardioprotective effect of the selective sigma-1 receptor agonist, SA4503. Yakugaku Zasshi. 134, 707-13. (2014)
Tsai S-Y, Hayashi T, Harvey BK, Wang Y, Wu WW, Shen R-F, et al.
Sigma-1 receptors regulate hippocampal dendritic spine formation via a free radical-sensitive mechanism involving Rac1{middle dot}GTP pathway.
- 43 -
Kikuchi-Utsumi K, Nakaki T.
Chronic treatment with a selective ligand for the sigma-1 receptor chaperone, SA4503, up-regulates BDNF protein levels in the rat hippocampus.
Neurosci Lett. 440, 19-22. (2008)
Hedrick NG, Harward SC, Hall CE, Murakoshi H, McNamara JO, Yasuda R.
Rho GTPase complementation underlies BDNF-dependent homo- and heterosynaptic plasticity.
Nature. 538, 104-108. (2016)
Harward SC, Hedrick NG, Hall CE, Parra-Bueno P, Milner TA, Pan E, Laviv T, Hempstead BL, Yasuda R, McNamara JO.
Autocrine BDNF-TrkB signalling within a single dendritic spine. Nature. 538, 99-103. (2016)
Su TP, Su TC, Nakamura Y, Tsai SY.
The Sigma-1 Receptor as a Pluripotent Modulator in Living Systems. Trends Pharmacol Sci. 37, 262-278. (2016)
Thoenen H.
Neurotrophins and neuronal plasticity. Science 270, 593–598. (1995)
- 44 -
Lewin GR, Barde YA.
Physiology of the neurotrophins.
Annu. Rev. Neurosci. 19, 289–317 (1996)
Horch HW.
Local effects of BDNF on dendritic growth. Rev Neurosci 15: 117–129. (2004)
Banerjee A, Castro J, Sur M.
Rett syndrome: genes, synapses, circuits, and therapeutics. Front Psychiatry. 3, 34. (2012)
Lauterborn JC, Rex CS, Kramár E, Chen LY, Pandyarajan V, Lynch G, Gall CM.
Brain-derived neurotrophic factor rescues synaptic plasticity in a mouse model of fragile X syndrome.
J Neurosci. 27, 10685-94. (2007)
Matsuno K, Nakazawa M, Okamoto K, Kawashima Y, Mita S.
Binding properties of SA4503, a novel and selective sigma 1 receptor agonist. Eur J Pharmacol. 306, 271-279. (1996)
- 45 -
Fujimura K, Matsumoto J, Niwa M, Kobayashi T, Kawashima Y, In Y, Ishida, T.
Synthesis, structure and quantitative structure-activity relationships of sigma receptor ligands, 1-[2-(3,4-dimethoxyphenyl)ethyl]-4-(3-phenylpropyl) piperazines.
Bioorg Med Chem. 5, 1675-83. (1997)
Maurice T, Privat A.
SA4503, a novel cognitive enhancer with sigma1 receptor agonist properties, facilitates NMDA receptor-dependent learning in mice.
Eur J Pharmacol. 328, 9-18. (1997)
Zou LB, Yamada K, Sasa M, Nakata Y, Nabeshima T.
Effects of sigma(1) receptor agonist SA4503 and neuroactive steroids on performance in a radial arm maze task in rats.
Neuropharmacology. 39, 1617-27. (2000)
Irwin SA, Idupulapati M, Gilbert ME, Harris JB, Chakravarti AB, Rogers EJ, Crisostomo RA, Larsen BP, Mehta A, Alcantara CJ, Patel B, Swain RA, Weiler IJ, Oostra BA, Greenough WT.
Dendritic spine and dendritic field characteristics of layer V pyramidal neurons in the visual cortex of fragile-X knockout mice.
- 46 -
Wiemerslage L, Lee D.
Quantification of mitochondrial morphology in neurites of dopaminergic neurons using multiple parameters.
J Neurosci Methods. 262, 56-65. (2016)
Dotti CG, Sullivan CA, Banker GA.
The establishment of polarity by hippocampal neurons in culture. J Neurosci. 8, 1454-1468. (1988)
van Spronsen M, van Battum EY, Kuijpers M, Vangoor VR, Rietman ML, Pothof J, Gumy LF, van Ijcken WF, Akhmanova A, Pasterkamp RJ, Hoogenraad CC.
Developmental and activity-dependent miRNA expression profiling in primary hippocampal neuron cultures.
PLoS One. 8, e74907. (2013)
Ahnert-Hilger G, Höltje M, Grosse G, Pickert G, Mucke C, Nixdorf-Bergweiler B, Boquet P, Hofmann F, Just I.
Differential effects of Rho GTPases on axonal and dendritic development in hippocampal neurons.
J Neurochem. 90, 9-18. (2004)
Hering H, and Sheng M.
Dendritic spines: structure, dynamics and regulation. Nat. Rev. Neurosci. 2, 880–888. (2001)
- 47 -
Holtmaat A, Svoboda K.
Experience-dependent structural synaptic plasticity in the mammalian brain. Nat. Rev. Neurosci. 10, 647–658. (2009)
Malik M, Rangel-Barajas C, Sumien N, Su C, Singh M, Chen Z, Huang RQ, Meunier J, Maurice T, Mach RH, Luedtke RR.
The effects of sigma (σ1) receptor-selective ligands on muscarinic receptor antagonist-induced cognitive deficits in mice.
Br J Pharmacol. 172, 2519-2531. (2015)
Francardo V, Bez F, Wieloch T, Nissbrandt H, Ruscher K, Cenci MA.
Pharmacological stimulation of sigma-1 receptors has neurorestorative effects in experimental parkinsonism.
Brain. 137, 1998-2014. (2014)
Fujimoto M., Hayashi T., Urfer R., Mita S., Su T.P.
Sigma-1 receptor chaperones regulate the secretion of brain-derived neurotrophic factor. Synapse, 66, 630–639. (2012)
Shioda N, Ishikawa K, Tagashira H, Ishizuka T, Yawo H, Fukunaga K.
Expression of a truncated form of the endoplasmic reticulum chaperone protein, σ1 receptor, promotes mitochondrial energy depletion and apoptosis.
- 48 -
Tagashira H, Zhang C, Lu YM, Hasegawa H, Kanai H, Han F, Fukunaga K.
Stimulation of σ1-receptor restores abnormal mitochondrial Ca²⁺ mobilization and ATP production following cardiac hypertrophy.
Biochim Biophys Acta. 1830, 3082-3094. (2013) Maletic-Savatic M, Malinow R, Svoboda K.
Rapid dendritic morphogenesis in CA1 hippocampal dendrites induced by synaptic activity.
Science 283, 1923–1927. (1999)
Engert, F. and Bonhoeffer, T.
Dendritic spine changes associated with hippocampal long-term synaptic plasticity. Nature 399, 66–70. (1999)
Trachtenberg JT, Chen BE, Knott GW, Feng G, Sanes JR, Welker E, Svoboda K. Long-term in vivo imaging of experience-dependent synaptic plasticity in adult cortex. Nature 420, 788–794. (2002)
Grutzendler J, Kasthuri N, Gan WB.
Long-term dendritic spine stability in the adult cortex. Nature 420, 812–816. (2002)
Rochefort NL, Konnerth A.
Dendritic spines: from structure to in vivo function. EMBO Rep. 13, 699–708. (2012)
- 49 -
Kasai H, Matsuzaki M, Noguchi J, Yasumatsu N, Nakahara H. Structure-stability-function relationships of dendritic spines. Trends Neurosci. 26 360–368. (2003
Egan MF, Kojima M, Callicott JH, et al.
The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function.
Cell. 112, 257–269. (2003)
Chen ZY, Jing D, Bath KG, Ieraci A, Khan T, Siao CJ, Herrera DG, Toth M, Yang C, McEwen BS, Hempstead BL, Lee FS.
Genetic variant BDNF (Val66Met) polymorphism alters anxiety-related behavior. Science 314, 140–143. (2006)
Ruscher K, Shamloo M, Rickhag M, Ladunga I, Soriano L, Gisselsson L, Toresson H, Ruslim-Litrus L, Oksenberg D, Urfer R, Johansson BB, Nikolich K, Wieloch T.
The sigma-1 receptor enhances brain plasticity and functional recovery after experimental stroke.
Brain 134, 732-746. (2011)
Yagasaki Y, Numakawa T, Kumamaru E, Hayashi T, Su TP, Kunugi H.
Chronic antidepressants potentiate via sigma-1 receptors the brain-derived neurotrophic factor-induced signaling for glutamate release.
- 50 -
Penas C, Pascual-Font A, Mancuso R, Forés J, Casas C, Navarro X.
Sigma receptor agonist 2-(4-morpholinethyl)1 phenylcyclohexanecarboxylate (Pre084) increases GDNF and BiP expression and promotes neuroprotection after root avulsion injury.
J Neurotrauma. 28, 831-840. (2011)
Takebayashi M, Hayashi T, Su TP.
Nerve growth factor-induced neurite sprouting in PC12 cells involves sigma-1 receptors: implications for antidepressants.
J Pharmacol Exp Ther. 303, 1227-1237. (2002)
Takebayashi M, Hayashi T, Su TP.
Sigma-1 receptors potentiate epidermal growth factor signaling towards neuritogenesis in PC12 cells: potential relation to lipid raft reconstitution.
- 51 -
発表論文リスト
1. SA4503, A Potent Sigma-1 Receptor Ligand, Ameliorates Synaptic Abnormalities and Cognitive Dysfunction in a Mouse Model of ATR-X Syndrome.
Yamaguchi K, Shioda N, Yabuki Y, Zhang C, Han F, Fukunaga K Int J Mol Sci. 2018 Sep 18;19(9). 2811.