痒みに関与する新規酸化リン脂質受容体の同定と機
能解析
著者
岸 貴之
学位授与機関
Tohoku University
学位授与番号
11301甲第19203号
URL
http://hdl.handle.net/10097/00129258
博⼠論⽂
痒みに関与する新規酸化リン脂質
受容体の同定と機能解析
令和元年度
東北⼤学⼤学院薬学研究科
⽣命薬科学専攻
岸 貴之
略語、略記号⼀覧
AP: alkaline phosphatase BSA: bovine serum albumin cDNA: complementary DNA
CRISPR: clustered regularly interspaced short palindromic repeat DLPA: di-linoleoyl phosphatidic acid
DLPC: di-linoleoyl phosphatidylcholine DLPE: di-linoleoyl phosphatidylethanolamine DLPG: di-linoleoyl phosphatidylglycerol DLPL: di-linoleoyl phospholipid
DLPS: di-linoleoyl phosphatidylserine
DMEM: Dulbeccoʼs modified Eagleʼs medium DMSO: dimethyl sulfoxide
DNFB: 2,4-dinitro-fluorobenzene
dNTP: deoxyribonucleoside 5ʼ-triphosphates DRG: dorsal root ganglion
ET-1: endothelin-1 FCS: fetal calf serum
GAPDH: glyceraldehyde 3-phosphate dehydrogenase GPCR: G protein-coupled receptor
HBSS: Hank's buffered salt solution H&E: hematoxylin-eosin
HEK: human embryonic kidney
HODE: hydroxyl-octadecadienoic acid
HPLC: high performance liquid chromatography HpODE: hydroperoxy-octadecadienoic acid KO: knockout
LA: linoleic acid
LC: liquid chromatography
Mrgprs (MRG): Mas-related G protein-coupled receptors mRNA: messenger ribonucleic acid
MS: mass spectrometry NGF: nerve growth factor
oxDLPA: oxidized di-linoleoyl phosphatidic acid oxDLPC: oxidized di-linoleoyl phosphatidylcholine oxDLPE: oxidized di-linoleoyl phosphatidylethanolamine oxDLPG: oxidized di-linoleoyl phosphatidylglycerol oxDLPL: oxidized di-linoleoyl phospholipid
oxDLPS: oxidized di-linoleoyl phosphatidylserine oxLA: oxidized linoleic acid
PUFA: poly unsaturated fatty acid LysoPG: lysophosphatidylglycerol LysoPS: lysophosphatidylserine LysoPT: lysophosphatidylthreonine PBS: phosphate-buffered saline PCR: polymerase chain reaction PFA: paraformaldehyde
PLA1: phospholipase A1
PSG: penicillin, streptomycin and glutamine RT: reverse transcription
sgRNA: single-guide RNA sn: stereo specific numbering SPF: specific pathogen free
TGFa transforming growth factor alpha WT: wild-type
⽬次
1. 序論 5 2. 実験材料と⽅法 16 3. 結果 32 4. 考察 43 5. 引⽤⽂献 87 6. 発表論⽂リスト 92 7. 謝辞 931. 序論
痒みは主に⽪膚表層に⽣じる、掻きたくなるような衝動を起こさせる不快な 感覚と定義される(1)。⽣体は、痒みを知覚することで、寄⽣⾍や刺激物の侵⼊ に対する除去⾏動を取ることができる。⼀⽅で、制御できない痒みは耐え難い苦 痛を伴う。実際に、痒みは様々な⽪膚疾患(アトピー性⽪膚炎、乾癬、接触性⽪ 膚炎、蕁⿇疹、アレルギー性疾患など)の主要な症状であり、患者の⽣活の質に 重⼤な影響を与えている(2)。さらに、痒みによる掻き⾏動は⽪膚刺激による⽪ 膚炎症の悪化に伴い、内因性の起掻因⼦の増加を介したさらなる痒みの増強と いう悪循環が起こる(1, 3)。従って、慢性掻痒性⽪膚疾患の治療には炎症抑制に 加えて、痒みを阻害するアプローチが有効である。後根神経節(DRG: dorsal root ganglion)に細胞体を有する求⼼性感覚神経(以 下、DRG 神経)の活性化は、痒みの誘導に必須である(4)。⽪膚を感覚⽀配する DRG 神経は脊髄後⾓に投射し、⽪膚において⽣じた痒み刺激を中枢に伝達する。 痒み刺激は、DRG 神経上の特異的な受容体を活性化することで、DRG 神経の 発⽕を誘導する。最も代表的な起掻因⼦としてヒスタミンが挙げられる(1, 5)。 ヒスタミンは特定の DRG 神経に発現する、特異的な G タンパク質共役型受容 体(GPCR)を介して痒みを誘導する(Fig. 1)。抗ヒスタミン薬(ヒスタミン受 容体拮抗薬)はヒトにおいて、ヒスタミン依存的な痒みを抑制することが知られ ており、痒みに対する第⼀選択薬として⽤いられている。しかし、ほとんどの慢 性掻痒性⽪膚疾患は、抗ヒスタミン薬では奏功しない(6)。また、ヒスタミン⾮ 依存的な痒み機構の全容は不明である。従って、新たなヒスタミン⾮依存的な痒 み機構を解明すること、さらに、その機構が関与する慢性掻痒性⽪膚疾患を同定 することは、新規痒み発症機序の解明、治療薬の開発に重要である。
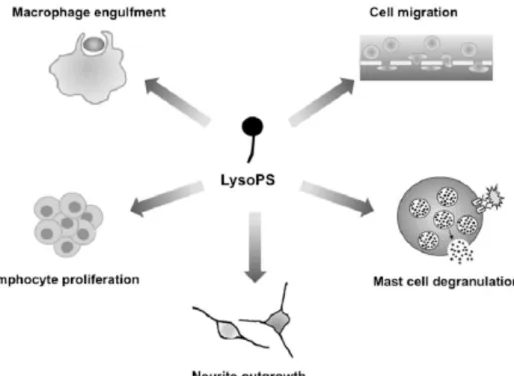
ヒスタミンはマスト細胞と呼ばれる免疫細胞が脱顆粒することにより産⽣さ れる。マスト細胞はヒスタミン以外にも、セロトニン、トリプターゼなどの起掻 因⼦を放出する、痒みの主要な原因細胞である(7)。すなわち、マスト細胞の脱 顆粒を抑制することは、抗ヒスタミン薬よりも幅広く痒みを抑制することがで きる。そこで、私はまず、マスト細胞の新たな脱顆粒機構を明らかにすることを ⽬指し、新規痒み治療薬の開発を試みた。 当研究室では、Lysophosphatidylserine (LysoPS)という、極性頭部にセリン残 基を有するリゾリン脂質に着⽬している。LysoPS は神経細胞の突起伸⻑などの 多様な薬理作⽤に加え、マスト細胞の脱顆粒を促進することが⽰されている (Fig. 2)(8, 9)。マスト細胞の脱顆粒促進作⽤は他のリゾリン脂質や、LysoPS の極性頭部のセリン残基を天然型の L-セリンから D-セリンに置換した⾮天然 型の LysoPS では認められないことから、マスト細胞上には LysoPS の構造を厳 密に認識する LysoPS 特異的受容体(LPSX)の存在が想定される(10)。これまで
に、LysoPS 受容体として、LPS1/GPR34、LPS2/P2Y10、LPS2L/A630033H20Rik
(ヒトの LPS2L相当遺伝⼦は偽遺伝⼦)、LPS3/GPR174 が同定されている(11, 12)。当研究室では、東京⼤学薬学系研究科⼤和⽥智彦教授との共同研究により、 LysoPS の構造類似体(LysoPS アナログ)約 300 種類を保有しており、その中 から LPS1、LPS2/2L、LPS3各受容体特異的アゴニストを⾒出している(10, 13, 14)。 私はこれらのマスト細胞脱顆粒作⽤を検証したが、LPS1、LPS2/2L、LPS3各受容 体特異的アゴニストはマスト細胞脱顆粒を誘導しなかった。従って、マスト細胞 上には、既知の LysoPS 受容体以外の LysoPS 受容体(LPSX)の存在が強く⽰唆 された(Fig. 3)。そこで、私は LPSXの同定を⽬的とし、LPSX同定に向けたツー ルを作製した。当研究室の巻出久美⼦元助教らは、LysoPS アナログの中で、極 性頭部にスレオニン残基を有する Lysophosphatidylthreonine (LysoPT)が in
vitro、in vivo において LysoPS の約 10 倍強⼒にマスト細胞脱顆粒を誘導するこ とを⾒出していた(10)。私は、LysoPT を上回る LPSX活性を有するアゴニスト の創製を⽬指し、上記 300 種類の LysoPS アナログのマスト細胞脱顆粒を評価 し、構造活性相関を検討した。その結果予測された活性モジュールを有機合成に より導⼊することで、LysoPS よりも約 100 倍強⼒なマスト細胞脱顆粒活性を有 する LPSX特異的アゴニスト(2-deoxy-1-C3-ph-p-O-C11-LysoPT)を創製した (Fig. 4)(15)。さらに、LysoPS を⽪下投与したマウスでは顕著な痒み⾏動が認 められ、2-deoxy-1-C3-ph-p-O-C11-LysoPT を投与したマウスではより強⼒な 痒み⾏動が認められた(Fig. 5)。従って、LPSXアゴニストは新規マスト細胞脱 顆粒機構、およびそれに伴う痒みの解析ツールとなりうると⽰唆された。 ⼀⽅で、マスト細胞を介さない痒み機構も知られている。マスト細胞以外の起 掻因⼦の産⽣細胞として、表⽪細胞(ケラチノサイト)が報告されている。表⽪ 細胞からは、神経成⻑因⼦(NGF)やエンドセリン-1(ET-1)が産⽣され、DRG 神経上の受容体に作⽤することで痒みが惹起される(4)。ところで、リノール酸 (炭素数 18、⼆重結合数 2、18:2)は表⽪において最も豊富に存在する多価不飽 和脂肪酸(PUFA: poly unsaturated fatty acid)である(16)。リノール酸はアラキ ドン酸(20:4)やドコサヘキサエン酸(22:6)などの他の PUFA と同様に、ラジ カル反応や酵素反応により、多様な酸化リノール酸分⼦種に代謝される(Fig. 6) (17)。興味深いことに、複数の酸化リノール酸分⼦種はヒト、および齧⻭類にお いて、⽪膚炎病態部で増加すること、マウスにおいて痒みを誘導することが報告 されている(18)。しかし、酸化リノール酸による痒みの誘導メカニズムは明らか にされていない。⽣体において、リノール酸はアシルセラミドやリン脂質に取り 込まれた形で存在する。実際に、表⽪において、リノール酸はセラミドに導⼊さ れ、アシルセラミドとして⽪膚バリア機構にはたらく(19)。また、酸化リノール
酸もリノール酸と同様に、表⽪のリン脂質に取り込まれることが知られている (20)。しかし、酸化リノール酸含有リン脂質の痒みにおける機能は分かっていな い。 私は本研究において、酸化リノール酸により誘導される痒み作⽤の⼀部が酸 化リノール酸含有リン脂質を介していると仮定し、研究に着⼿した(Fig. 7)。と ころが、予想に反し、特定の酸化リノール酸含有リン脂質は酸化リノール酸とは 異なる作⽤機構で、酸化リノール酸よりも強⼒な痒みを誘導した。また、本研究 では、同標的分⼦の⽣化学的な性質の検討、並びに慢性掻痒性⽪膚疾患における 機能解析を⾏い、新規ヒスタミン⾮依存的な痒み機構の解明に迫った。
DRG
Fig. 1.
DRG: dorsal root ganglion DRG
DRG DRG
Fig. 2. LysoPS
LysoPS
LysoPS LysoPS
Fig. 3.
LysoPS
LPS
X A: LysoPS LPS1(GPR34) LPS2 (P2Y10) LPS2L(A630033H20Rik) LPS3(GPR174) GPCR B: LysoPS LPSX LPSXLysoPS
HO O O NH2 P OH O O O OH O LPS1/ GPR34 LPS2 / P2Y10 LPS2L / A630033H20Rik LPS3 / GPR174LysoPS
LPSx LPS1-3 LysoPSA
B
Fig. 4. LPS
X A: LysoPS 300 LysoPS 2-deoxy-1-C3-ph-p-O-C11-LysoPT B, C: 2-deoxy-1-C3-ph-p-O-C11-LysoPT LPSX LysoPS 100 B C 0 20 40 60 34 36 38時間 (min)
直腸温度
(
℃
)
10-9 10-8 10-7 10-6 0 25 50 75 100 化合物濃度 (M) ヒスタミン放出量 ( % )in vitro
in vivo
LPSx LysoPS LPSx LysoPSA
B
C
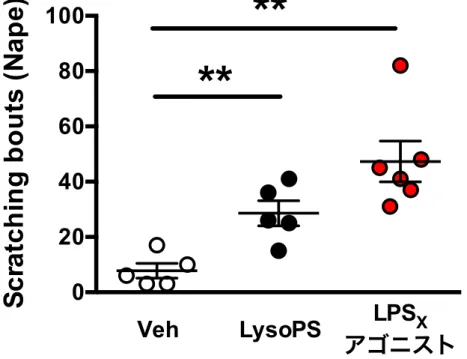
Fig. 5. LPS
XLysoPS
LysoPS 1 mM 50 µL
2-deoxy-1-C3-ph-p-O-C11-LysoPT 100 µM LysoPS
Data are expressed as mean ± s.e.m., **P < 0.01 by two-tailed unpaired Student’s t-test.
0
20
40
60
80
100
Sc
ra
tc
h
in
g
b
o
u
ts
(
N
ap
e)
Veh
LysoPS
LPS
Xアゴニスト
**
**
Fig. 6.
A: 2
1 9-HpODE 13-HpODE
B: 9-HpODE 13-HpODE 31
Peter Spiteller et al., 2001
A
Fig. 7.
2. 実験材料と⽅法
2-1. 実験動物 マウスは SPF 環境下、⾃由飲⽔、及び、9:00-21:00 明期、21:00-9:00 暗期の 明暗周期で飼育した。また、動物の取り扱い東北⼤学⼤学院薬学研究科動物実験 委員会が作製した「実験動物の取り扱いに関する指針」に基づいて⾏った。 野⽣型(WT)C57BL/6J マウス、マスト細胞⽋損マウス(WBB6F1-W/Wv)、 及び、対照群マウス(WBB6F1-WT)は⽇本クレアから購⼊した。 ・CRISPR/Cas9 法による MrgB5 KO マウスの作製 MrgB5 の N 末側に 3 種類の sgRNA(sgRNA1-3)を設計し、そのうち 2 種類 の sgRNA mixture と Cas9 タンパク質を C57BL/6J マウス受精卵へインジェク ションした(インジェクションは兵庫医科⼤学の⼤村⾕昌樹先⽣に実施してい ただいた)。上記受精卵を偽妊娠マウスに移植し、F0 個体を得た。F0 個体の尻 尾ゲノムを抽出し、sgRNA 標的領域付近を PCR 増幅し、ラージデリーション 変異を確認した個体を選別した。次に、上記 F0 個体と野⽣型マウスと交配し、 F1 個体を得たのち、変異の遺伝を確認した。変異が遺伝した F1 個体(Mutant #1 および#2)の MrgB5 配列をクローニングし、シーケンス解析により MrgB5 配列を解析したところ、N 末に約 300 塩基に及ぶラージデリーション変異を確 認した。Mutant #1 および#2 から交配を繰り返し、MrgB5 KO マウスライン(KO #1、#2)を確⽴した。KO #1: sgRNA 1 と sgRNA 2 の mixture を使⽤ KO #2: sgRNA 1 と sgRNA 3 の mixture を使⽤
sgRNA 1: 5ʼ-GGTATTGAACTTGCTGACACAAGG-3ʼ sgRNA 2: 5ʼ-CAGGAAATGCCATAGTCCTATGG-3ʼ sgRNA 3: 5ʼ-TAGAGAGCGCTCAACACTGATGG-3ʼ マウスの genotyping は以下のように⾏った。マウスの尾の先端を PCR tube に採取し、50 mM NaOH 180 µL を加え、サーマルサイクラー(TAKARA)を ⽤いて、95℃で 50 分間反応させ、ゲノム DNA を抽出した。その後、1 M Tris-HCl (pH 8.0)を 20 µL 加えて中和したものを PCR templete として⽤いた。得ら れた PCR 産物は 3% agarose gel で泳動した。KO マウスを⽤いる際は、littermate の WT マウスを対照群として使⽤した。
使⽤したプライマーの配列と PCR の条件は以下の通りである。 ・プライマーの配列
Forward primer: 5ʼ-CTAACCACTCCAGCCTGGAAC-3ʼ Reverse primer: 5ʼ-CTTGGTCTTTGGCAGTGATACC-3ʼ ・PCR 反応液
10 x PCR buffer 1 µL
2.5 mM dNTP 0.8 µL Primer mixture (each 2 µM) 0.6 µL
PCR templete 1.5 µL rTaq 0.55 µL 25 mM MgCl2 0.225 µL MilliQ water 5.825 µL ・反応条件 {95℃, 15 秒→55℃, 30 秒→72℃, 30 秒}(35 cycle)→10℃, ∞
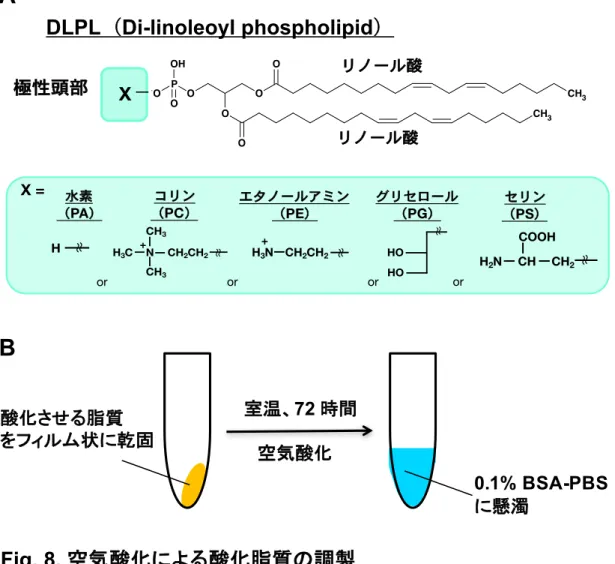
2-2. 脂質
リン脂質は Avanti Polar Lipids より購⼊した。クロロホルムに溶解している 各 脂 質 を マ イ ク ロ シ リ ン ジ で 必 要 量 を コ ー テ ィ ン グ 済 み 試 験 管 (ASAHI TECHNO GLASS) に取り、窒素ガスで乾燥固化した後、0.1% BSA(SERVA) 含有 PBS を加えて bath sonicator での超⾳波処理とボルテックスによる撹拌を 2 分間ずつ 2 回⾏い、1 mM に調製した。PBS は後述(2-6)する 20 x PBS を MilliQ water で 20 倍希釈したものを使⽤した。 酸化脂質の調製は、特に表記がない限り、脂質を乾燥固化後、72 時間暗所で 放置した後、上記のように 0.1% BSA 含有 PBS に溶解した。本研究では、酸化 脂質の濃度は基質となる脂質濃度として表記した。本研究において、個体に投与 するすべての実験で 1 mM 相当を投与した。 リノール酸と酸化リノール酸(9-HODE、13-HODE、9-HpODE、13-HpODE) は Cayman Chemical から購⼊した。
oxLysoPG (18:2)は DLPG から PLA1反応により調製した LysoPG (18:2)を上
記⽅法で空気酸化したものを⽤いた(2-13: PLA1反応の項を参照)。 2-3. 痒み⾏動の測定 痒み⾏動の評価実験は午前 9:00-12:00 の間に実施した。また、事前にマウス をケージに 30 分以上馴化させた。実験⽇の 24 時間以上前に、シェーバーでマ ウスのうなじ部分、または頬の剃⽑を⾏った。イソフルラン⿇酔したマウスに、 29G のマイジェクターを⽤いて、各試験化合物をうなじ部分(50 µL)、または 頬(20 µL)に⽪下投与した。その後、30 分間の後脚による掻き⾏動の数を⽬視 によりカウントした。WT と KO マウスを⽐較する場合は、genotype について ブラインド下で⾏い、実験後、前述の⽅法で genotyping を⾏った。阻害剤を前
処置する場合は、阻害剤 50 µL をうなじ部分に⽪下投与し、30 分後、同じ部位 に痒み物質を 50 µL ⽪下投与した。なお、阻害剤は⽣理⾷塩⽔で下記の濃度に 調製したものを⽤いた。
・YM-254890(Gq/11 阻害剤)(Wako):10 µM in 0.5% DMSO
・capsazepine(TRPV1 阻害剤)(Wako):100 µM in 0.5% DMSO ・U73122(PLC 阻害剤)(Cayman Chemical):10 µM in 0.5% DMSO
2-4. In situ hybridization
下記の⽅法で作製した切⽚を RNA ScopeTM Detection Reagent Kit-RED
(ACDBio)(c-fos, TRPV1 mRNA)、または BaseScopeTM Detection Reagent
Kit-RED(ACDBio)(MrgB5 mRNA)を⽤いて添付⽂書に従って染⾊した。カウンタ ー染⾊としてヘマトキシリン染⾊を⾏った。
・DRG 神経活性化の評価
本実験では、脊髄後⾓の最外層にあたる laminaeⅠ-Ⅱにおける c-fos mRNA 発現量をもって DRG 神経の活性化の指標とした(21)。剃⽑したマウスうなじ部 分に試験化合物を⽪下投与した。30 分後、⿇酔により安楽死させ、うなじ部位 を感覚⽀配する DRG 神経の投射先である頚椎(C3-C5)を丁寧に採取した。頚 椎は 4% (w/v) PFA-PBS 中に浸して 25℃、12 時間固定し、10%→20% (w/v)ス クロース PBS 溶液に置換後、O.C.T. compound (Sakura Finetek)に浸し、液体 窒素を⽤いて凍結包埋した。クライオスタット(Leica)を⽤いて 10 µm 厚の凍 結切⽚を作製し、Mas-coated glass slides (Matsunami Glass)に貼り付けた。 ・DRG における受容体発現解析
を取り除いた後、DRG を採取した。DRG は頚椎同様に凍結包埋し、クライオ スタット(Leica)を⽤いて 7 µm 厚の凍結切⽚を作製し、Mas-coated glass slides (Matsunami Glass)に貼り付けた。MrgB5 mRNA 発現細胞と TRPV1 mRNA 発 現細胞について解析する際は、連続切⽚を⽤いた。 4% (w/v) PFA-PBS の組成は以下の通り調製した。 ・4% (w/v) PFA-PBS paraformaldehyde (Wako) 20 g MilliQ water 450 mL 溶解後、3N NaOH 50 µL を添加し、ホットスターラー上で 60℃に加温しなが ら撹拌した。PFA 溶解後、放冷し、10 x PBS を 50 mL 加えた。10 x PBS は後 述(2-6)する 20 x PBS を MilliQ water で 10 倍希釈したものを使⽤した。 2-5. TRPV1 陽性神経の除神経 TRPV1 陽性神経の除神経は以前の報告を参考に⾏った(22)。マウスに 3 種混 合⿇酔(M/M/B: 0.3 mg/kg medetomidine, 4.0 mg/kg midazolam, and 5.0 mg/kg butorphanol)を腹腔内投与し、⿇酔したのち、25 mg/kg、50 mg/kg、50 mg/kg のカプサイシン(Wako)を 1 ⽇ 1 回 3 ⽇間にわたって背部に 200 µL ⽪下投与 した。カプサイシンは下記の組成の溶媒に懸濁して使⽤した。14 ⽇後、TRPV1 陽性神経の消失を確認するため、1 mM カプサイシン 4 µL をマウスに点眼し、 wiping ⾏動を確認した。実験には、wiping ⾏動が⾒られない個体を使⽤した。 ・カプサイシン溶媒 エタノール 100 µL Tween 20 (Wako) 100 µL ⽣理⾷塩⽔ 800 µL
2-6. 細胞培養
細胞は HEK293A 細胞(Thermo Fisher Scientific)を使⽤した。G タンパク質 ⽋損 HEK293A 細胞は当研究室の井上⾶⿃准教授らが樹⽴した Gq/11 DKO、G12/13
DKO、Gq/11/12/13 QKO HEK293 細胞を⽤いた (23, 24)。細胞の継代・培養には
10% FCS、PSG 含有の DMEM と PBS を⽤い 2 ⽇おきに⾏い、培養は 5% CO2 存在下の CO2 インキュベーター(37℃)にて⾏った。 細胞の維持に⽤いた試薬は以下の通り調製した。 ・DMEM DMEM (⽇⽔製薬) 4.75 g MilliQ water 500 mL 溶解後、オートクレーブ滅菌 (121˚C、15 分) し、以下の試薬を加えた。 100 x PSG 5 mL 10% (w/v) NaHCO3 7.5 mL ⾮働化 FCS (GIBCO) 50 mL ・100 x PSG
Penicillin G sodium salt (SIGMA) 3 g Streptomycin sulfate (GIBCO) 5 g L-Glutamine (GIBCO) 14.6 g
MilliQ water 500 mL
溶解後、Poresize 0.22 µm の 595 Bottle Top Filters (NALGENEL abware) で濾過滅菌し、-20℃で保存した。
・10 % (w/v) NaHCO3 NaHCO3 (Wako) 10 g MilliQ water 100 mL オートクレーブ滅菌 (121˚C、15 分) 後、使⽤した。 ・20 x PBS NaCl (Wako) 80 g KCl (Wako) 2 g Na2HPO4・12H2O (Wako) 29 g KH2PO4 (Wako) 2 g MilliQ water 500 mL PBS は 20 x PBS を MilliQ water で 20 倍希釈してオートクレーブ滅菌 (121˚C、15 分) 後、使⽤した。 2-7. TGFa 切断アッセイ
HEK293A 細胞(Parent 株、または各種 G タンパク質⽋損 HEK293 細胞)を 2.0 x 105 cells/mL で培養⽤ dish (greiner bio-one)に播種し、5% CO
2 存在
下 37˚C で 24 時間培養した。以下、培養液 10 mL(10 cm dish に播種した際 に相当)の場合の操作を記述する。24 時間培養後、Opti-MEM(Invitrogen) 500 µL 中に各種遺伝⼦をコードしたプラスミド(AP-TGFa, 2.5 µg; GPCR, 1 µg) を添加した。別のチューブで Opti-MEM 475 µL と 25 µL のポリエチレンイミ ン(PEI: Polyethylenimine “Max”(Polysciences)、1 mg/mL)を混合し、5 分 後、両溶液を 500 µL ずつ混合し、さらに 20 分後、全量(1 mL)を培養⽫に滴 下することでトランスフェクションを⾏った。全ての遺伝⼦はベクターとして
pCAGGS を⽤いた。本ベクターは、CAG プロモーターを有し哺乳類細胞で強⼒ な発現を誘導する。また、AP-TGFa(AP 標識 TGFa)は愛媛⼤学⼤学院医学 系研究科東⼭繁樹教授により作製されたものを使⽤した。N 末端に human placental alkaline phosphatase が標識された pro-TGFaが細胞膜上に発現するよ う設計がされている。G タンパク質のレスキュー実験の際は、トランスフェク ションの際に GqΔC、Gq、または G11をコードするプラスミド 500 ng を
AP-TGFa、GPCR とともに導⼊した。GqΔC は Gq タンパク質の、GPCR との共
役に重要であるとされる C 末端 7 アミノ酸を⽋失した変異体である。
トランスフェクション 24 時間後、細胞を 2.0 x 104 cells/well となるように
HBSS-HEPES に懸濁し、96-well plate(greiner bio-one)に播種した。30 分間 starvation をした後、各濃度の化合物を各 well に添加し、5% CO2存在下 37˚C
で 1 時間刺激した。
刺激後、96-well plate を plate spin(KUBOTA)を⽤いて 190 x g で 2 分間 遠⼼し、細胞を沈殿させた。その後、上清 80 µL を ASSAY PLATE(IWAKI,3881-096)に分取し、2 x p-NPP solution を上清及び細胞に 80 µL 加え、ただちに 405 nm での吸光度をマイクロプレートリーダー(Spectramax, Molecular Devices) で測定し、バックグラウンドとした。その後、室温で 1 時間インキュベートし た後に 405 nm での吸光度をマイクロプレートリーダーで測定した。 TGFa 切断量(%)は 1 時間インキュベート後の吸光度からバックグラウン ドを引き、以下の式にて算出した。 × 100 さらに、無刺激群(vehicle 刺激群)の TGFα 切断量を差し引いて評価した。 TGFa 切断量
=
OD405(上清)+ OD405(細胞) OD405(上清)TGFa 切断アッセイに⽤いた試薬は以下の通り調製した。 ・HBSS/5 mM HEPES 20 mM HEPES(pH 7.4) 10 mL 1 N KOH 120 µL MilliQ water 790 mL オートクレーブ滅菌(121℃, 15 分)後、以下の液を加え使⽤した。 HBSS stock solution.1 100 mL HBSS stock solution.2 100 mL ・HBSS stock solution.1 CaCl2・2H2O(Wako) 1.85 g MgCl2・6H2O(Wako) 1 g MgSO4・7H2O(Wako) 1 g MilliQ water 1000 mL ・HBSS stock solution.2 KCl(Wako) 4 g KH2PO4 (Wako) 0.6 g NaCl(Wako) 80 g Na2HPO4・12H2O(Wako) 1.2 g D-Glucose(Wako) 10 g MilliQ water 1000 mL HBSS stock solution は調整後、オートクレーブ滅菌し、4 ℃で保存した。
・2 x p-nitrophenyl phosphate (p-NPP) buffer 1 M Tris (pH9.5) 6 mL 5 M NaCl 1 mL 1 M MgCl2 1 mL イオン交換⽔ 92 mL 使⽤時、2x p-NPP buffer 100 mL に対し、1mM p-NPP を 1 mL 加え、2 x p-NPP solution とした。
・NaCl(Wako)、MgCl2(Wako) 及びp-NPP(Wako)はイオン交換⽔に溶解
して⽤いた。
2-8. RNA 抽出、逆転写反応、定量 PCR
Total RNA の抽出には GenEluteTM Mammalian Total RNA Miniprep Kit
(Sigma Aldrich)を⽤いて⾏った。C57BL/6J マウスから各組織を摘出し、ミン スした。キット付属の Lysis buffer 500 µL を分注した tube(TOMY)に、各組 織 30 mg ずつを回収し、MS-100R(TOMY)を⽤いてビーズ破砕した後、添付 ⽂書に従い抽出を⾏った。
逆転写反応は High capacity cDNA RT kit(Applied Biosystems)を⽤いて添 付⽂書に従い⾏った。
・1 x premixture の組成
10 x RT Buffer 1 µL
25 x dNTP 0.4 µL
10 x random primer 1 µL Reverse Transcriptase(50 U/mL) 0.5 µL
RNase inhibitor 0.5 µL RNase free water 1.6 µL 100 ng/mL に調製した RNA 10 µL を PCR plate に分注し、1 x premixture を 10 µL 加え、サーマルサイクラー(TAKARA)により、25℃, 10 分→ 37℃, 120 分→ 85℃, 5 分にて逆転写反応を⾏った。
cDNA 溶液は MilliQ water を⽤いて適宜希釈したものを使⽤した。その後、 SYBR Premix Ex TaqⅡ(TAKARA)を⽤いて添付⽂書に従い、Applied Biosystems 7300 Real Time PCR system (Applied Biosystems)を使⽤して定量 RT-PCR を ⾏った。融解曲線作製により、⽬的の遺伝⼦特異的な PCR 増幅が起きているこ とを確認した。検量線は standard plasmid を⽤いて作製し、cDNA 溶液中の転 写産物を定量した。また、1 サンプルにつき、同⼀条件で 2 well の測定を⾏い、 その平均値を定量値とした。得られたデータは、Gapdh のコピー数で補正した。
反応液の組成及び、プライマー配列は以下の通りである。 ・反応液の組成
2 x SYBR Premix Ex Taq II 5 µL 50 x ROX Reference Dye 0.2 µL Primer mixture (each 1 µL) 2 µL cDNA 溶液(10 倍希釈) 3 µL ・プライマーの配列(final 100 nM)
・Mrgprb5
Forward primer: 5ʼ-GCCAATCCCATCATTTACTTTCTTG-3ʼ
Reverse primer: 5ʼ-AGTTCCCCAGACCTTTGTGAG-3ʼ ・Gapdh
Forward primer: 5ʼ-AGGAGCGAGACCCCACTAAC-3ʼ Reverse primer: 5ʼ-CGGAGATGATGACCCTTTTG-3ʼ ・Ifn-g
Forward primer: 5ʼ-GAGGAACTGGCAAAAGGATGG-3ʼ Reverse primer: 5ʼ-GTTGCTGATGGCCTGATTGTC -3ʼ
2-9. 乾⽪症モデル 乾⽪症モデルマウスの作製は以前の報告を参考に⾏った(25)。イソフルラン ⿇酔下でマウスのうなじ部分を剃⽑した。アセトン、ジエチルエーテルの等量混 合液に浸した脱脂綿をマウスうなじ部分に 15 秒貼付した。その後、直ちに⽔に 浸した脱脂綿を同⼀箇所に 15 秒貼付した。上記操作を 12 時間おきに 6 ⽇連続 継続した。最終の操作から 24 時間後、60 分間の後脚による掻き⾏動の数を⽬視 によりカウントした。 2-10. アレルギー⽪膚炎モデル アレルギー⽪膚炎モデルマウスの作製は以前の報告を参考に⾏った (25)。イ ソフルラン⿇酔下でマウスのうなじ部分を剃⽑した。 29G マイジェクターを⽤ いて、下記の組成で調製した OVA 溶液をマウスに OVA が 50 µg/body となる ように 10 ⽇おきに 2 回腹腔内投与した。2 度⽬の投与から 7 ⽇後、29G マイジ ェクターを⽤いて、⽣理⾷塩⽔に溶解した OVA を 50 µg/body の⽤量でマウス うなじ部分に⽪下投与した。その後、30 分間の後脚による掻き⾏動の数を⽬視 によりカウントした。
・OVA 溶液の組成
Alhydrogel adjuvant 2% (Invitrogen) 100 µL ⽣理⾷塩⽔ 400 µL
2-11. 接触性⽪膚炎モデル
接触性⽪膚炎モデルマウスの作製は以前の報告を参考に⾏った(25)。イソフ ルラン⿇酔下でマウスの腹部、および、うなじ部分を剃⽑した。下記に組成を⽰ す 2,4-dinitro-fluorobenzene (DNFB)(Sigma Aldrich) 溶液を腹部に 25 µL ずつ、 1 ⽇おきに 3 ⽇連続塗布した(Day 1-3)。さらに 5 ⽇後、DNFB 溶液をうなじ 部分に 25 µL ずつ、1 ⽇おきに 3 ⽇連続塗布した(Day 8-10)。各タイムポイン トにおいて、30 分間の後脚による掻き⾏動の数を⽬視によりカウントした(Day 8-12)。なお、測定は DNFB 溶液を塗布する直前に実施した。 ・DNFB 溶液の組成 アセトン 1.2 mL オリーブオイル 0.3 mL DNFB 5 µL ⽪膚炎症の評価は、以下のように⾏った。 ・⽪膚厚の測定 マウスを PBS により全⾝灌流したのち、4% (w/v) PFA-PBS で灌流固定した。 その後、うなじ部分の⽪膚(表⽪、真⽪)を回収し、4% (w/v) PFA-PBS で 25℃、 24 時間固定した。PBS で洗浄し(10 分 x 3)、50% エタノール(1 時間 x 1)→ 70% エタノール(1 時間 x 2)→ 90% エタノール(1 時間 x 2)→ 95% エタ ノール(1 時間 x 2)→ 100% エタノール(30 分 x 3)→キシレン(30 分 x 3) →パラフィン(3 時間以上 x 3)の順に浸した。その後、パラフィンブロックに 加⼯し、ミクロトームを⽤いて 10 µm 厚の切⽚を切り出した。湯浴中で⼗分伸
展したのち、Mas-coated glass slides (Matsunami Glass)に貼り付けた。その後、 ホットプレート上(45℃)で、乾燥させた。スライドガラスをキシレン(3 分 x 3)に漬けパラフィンを溶解した後、100% エタノール(3 分 x 3)→90% エタ ノール→70% エタノールに各 3 分浸した。流⽔洗浄後(2 分)、ヘマトキシリン に 10 分間浸した。再び流⽔洗浄した後(2 分)、エオジンに 2 分間浸した。⽔→ 70% エタノール→90% エタノールに各 2 秒浸し、100% エタノール→キシレ ンに各 3 分間浸し、Entellan new (MERCK)を⽤い、カバーガラスを乗せ、封⼊ した。その後、各個体につき、10 切⽚の⽪膚厚を測定し、平均値を算出した。 ・炎症マーカー(IFNg)mRNA 測定 マウスを PBS により全⾝灌流したのち、うなじ部分の⽪膚(表⽪、真⽪)を 回収し、細かくミンスした。30 mg を秤量し、前述(2-8)した⽅法の通りに qRT-PCR を⾏い、IFNg mRNA 量を測定した。 抗酸化剤の投与は、以下のように⾏った。 ・N-アセチルシステイン(NAC)
Day 1-12 にわたり、NAC (Wako)を 2 g/L の⽤量で溶解したものを⾃由飲⽔ により継続投与した。2 ⽇ごとに飲⽔量を測定するとともに、新しく調製したも のと交換した。なお、飲⽔量は通常の⽔を飲⽔した群とほぼ同等であった。 ・a-トコフェロール(a-Toc)
Day 1-12 にわたり、1 ⽇おきに、a-Toc (Wako)を⽣理⾷塩⽔で懸濁し、250 mg/kg の⽤量で経⼝投与した。
2-12. ⽪膚脂質の Folch 抽出
⾏った。マウスから採取した⽪膚 50 mg をメタノール 0.33 mL、クロロホルム 0.66 mL を分注した tube(TOMY)に回収し、MS-100R(TOMY)を⽤いてビ ーズ破砕した。その後、⽪膚懸濁溶媒をガラス試験管(ASAHI TECHNO GLASS) に移し、45 分間撹拌した。⽣理⾷塩⽔ 0.198 mL を添加し、5 分間撹拌し、21,500 x g、5 分間遠⼼分離した。遠⼼後、⽔溶性成分として上層を、脂溶性成分として 下層を回収した。上層、下層の溶媒をエバポレーションにより除去した。より抽 出効率、および純度を⾼めるため、溶媒を除去した上層、下層に再度、メタノー ル 0.33 mL、クロロホルム 0.66 mL を添加し、撹拌後、⽣理⾷塩⽔ 0.198 mL を添加し、上記の操作を繰り返した(*)。本研究では(*)操作を 2 度繰り返し たサンプルを⽤いた。各画分について、抗酸化物質 2,6-di-t-butyl-p-cresol (BHT) (ナカライテスク)を 1 mM 含むメタノール 500 µL に懸濁し、-80℃で保存し た。アッセイに⽤いる際は、エバポレーションによる溶媒除去を⾏ったのち、 0.01% BSA 含有 HBSS に再懸濁したものを⽤いた。 2-13. PLA1反応 PLA1反応は当研究室川名裕⼰特任助教らが構築した⽅法に基づいて⾏った (26)。DLPG を基質として、78 mM リン酸 buffer (pH 7.4)/10% ジエチルエー テル/0.05% TritonX100/2 mM CaCl2条件下において、Rhizomucor miehei 由来
リパーゼ(Sigma Aldrich)を⽤いて(基質 1 nmol に対して 1 unit)37℃、80 分 間反応させた。9 倍量の酸性化メタノール(1M ギ酸アンモニウム 1 mL とメタ ノール 99 mL にギ酸を添加し、pH 4 に調製)を⽤いて反応を停⽌した。4℃下、 21,500 x g、5 分間遠⼼後の上清を固相抽出に供した。Bond Elut C18(Agilent Technologies)を⽔系溶媒 A: 5 mM ギ酸アンモニウム⽔溶液(pH 4)と有機系溶 媒 B: 5 mM ギ酸アンモニウム 99.5%メタノール(pH 4)を⽤いてコンディショ
ニングし、サンプルをロードした。50% B 溶液でカラムを洗浄し、その後、90% B 溶液で溶出した。上記操作により、PLA1反応の残存ジアシルリン脂質を除去
し、LysoPG(18:2)をできる。アッセイに⽤いる際は、エバポレーションによる 溶媒除去を⾏ったのち、0.01% BSA 含有 HBSS に再懸濁したものを⽤いた。
3. 結果
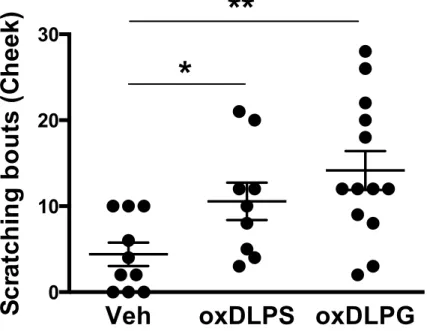
3-1. 酸化リノール酸含有リン脂質の痒み誘導作⽤ 乾癬などの痒み病態時の⽪膚では、複数の酸化リノール酸分⼦種が検出さ れ、その中でも特定の分⼦種は痒みを誘導することが知られている。⽪膚にお いてリノール酸(18:2)、酸化リノール酸は容易にリン脂質に取り込まれるた め、その多くはリン脂質体として存在する。そこで、私は酸化リノール酸の⼀ 部はリン脂質体として痒みを誘導するのではないかと仮説をたて、検討を⾏っ た。まず、酸化リノール酸を含有するリン脂質がマウスの痒み知覚を惹起する かどうか検討した。酸化リン脂質として、2 本のリノール酸を含むリン脂質 (Di-linoleoyl phospholipid、DLPL)の空気酸化物(以下、oxDLPL)を調製 した(Fig. 8)。極性基の異なる 5 種類の DLPL をマウスうなじ部分に⽪下投与 し、30 分間における、後脚による掻き⾏動を測定した。極性頭部に⽔素、コリ ン、エタノールアミンをそれぞれ有する oxDLPA、oxDLPC、oxDLPE を投与 した群では、有意な掻き⾏動の増加は認められなかった。⼀⽅、グリセロー ル、セリンをそれぞれ有する oxDLPG、oxDLPS 投与群では、顕著な掻き⾏動 の増加が観察された。また、空気酸化処理を⾏っていない未酸化の DLPG は 有意な掻き⾏動を誘導しなかった(Fig. 9)。 ⼀般に、うなじ部分への薬物投与依存的な後脚による掻き⾏動は、「痒みと 痛み」の両⽅を反映するとされている。そこで、oxDLPG、oxDLPS をマウス の頬に投与することで、これら薬物による掻き⾏動が「痒み」と「痛み」のど ちらに起因するかどうか検討した。薬物の頬への投与では、前脚による wiping ⾏動は痛みを、後脚による掻き⾏動は痒みをそれぞれ表すことが知られている (27)。興味深いことに、oxDLPG、oxDLPS を頬に投与した群では、wiping ⾏ 動はほとんど惹起されず、掻き⾏動が有意に増加した(Fig. 10)。したがって、oxDLPG、oxDLPS はマウスに痒み知覚を誘導することが⽰された。以下 の実験では、痒み作⽤の評価は実験の簡便性から、うなじへの薬物投与により 評価している。 前述の通り、ある種の酸化リノール酸はマウスに痒みを誘導することが⽰さ れている。そこで、酸化リノール酸含有リン脂質と酸化リノール酸の痒み誘導作 ⽤を⽐較した。酸化リノール酸としては、リノール酸を空気酸化処理したもの (oxLA)、⼊⼿可能であった 9-HODE、13-HODE、9-HpODE、13-HpODE を ⽤いた。各酸化リノール酸はマウスに掻き⾏動を誘導した(Fig. 11)。DLPG を 酸化処理したサンプルでは、未酸化の DLPG が 90%以上残存していたことか ら、酸化効率は少なくとも 10%以下である(data not shown)。このことを考慮 すると、各酸化リノール酸の痒み作⽤は oxDLPG と⽐較して弱いと考えられた。 したがって、oxDLPG は酸化リノール酸よりも極めて強⼒な痒み物質であるこ とが⽰唆された。 3-2. PG 酸化物の痒み誘導作⽤に関する、脂肪酸特異性の解析 次に、様々な脂肪酸を有する PG 酸化物の痒み誘導作⽤に関する特異性につ いて検討した。PG 分⼦種の脂肪酸としては、飽和脂肪酸を 2 本有する PG(di-16:0 PG)、飽和脂肪酸とリノール酸を 1 本ずつ有する PG(18:0/18:2 PG)、酸 化を受けにくい単価不飽和脂肪酸を 2 本有する PG(di-18:1 PG)、さらにはリ ノール酸以外の多価不飽和脂肪酸(PUFA)を 2 本有する PG(di-18:3 PG、di-20:4 PG、di-22:6 PG)の空気酸化物を⽤いた。酸化修飾を受けにくい di-16:0 PG、di-18:1 PG の空気酸化物は痒み作⽤を⽰さなかった。⼀⽅、di-18:2 PG (DLPG)、di-18:3 PG、di-20:4 PG、di-22:6 PG といった PUFA を 2 本含有す る PG 酸化物投与群では有意な痒み⾏動の増加が認められた。意外なことに、
リノール酸を 1 本有する 18:0/18:2 PG 酸化物は痒みを惹起しなかった(Fig. 12)。PS 酸化物の分⼦種については詳細な検討を⾏っておらず、今後解析は必 要ではあるが、上記の結果から、PG 酸化物による痒み作⽤には、PUFA を 2 本 含有することが必要であることが⽰唆された。 3-3. 新規掻痒性酸化リン脂質の標的細胞の解析 主要な痒み物質として知られるヒスタミン、セロトニンなどはマスト細胞が 活性化、脱顆粒することで産⽣され、⽪膚を⽀配する感覚神経上の受容体を介し て痒み知覚をもたらす (5, 6, 7)。oxDLPG、oxDLPS による痒み作⽤がマスト細 胞を介した種々の痒み物質産⽣に起因するかどうか検討するため、マスト細胞 ⽋損マウス(W/Wv マウス)うなじ部位へ oxDLPG、oxDLPS を⽪下投与した 後の掻き⾏動を評価した(28)。W/Wv マウスにおいて、oxDLPG 投与群では野 ⽣型マウスとほぼ同等な掻き⾏動が認められた。⼀⽅、oxDLPS 投与群では、野 ⽣型マウスと⽐較し、W/Wv マウスで有意な差ではないが、わずかな掻き⾏動 の減弱が認められた(Fig. 13)。PS は⾼濃度においてマスト細胞の脱顆粒する 作⽤が報告されており、我々の結果と併せ、oxDLPS ⽪下投与による痒み作⽤の ⼀部はマスト細胞を介していることが⽰唆された。しかしながら、oxDLPG、 oxDLPS による痒み作⽤の⼤部分はマスト細胞⾮依存的であることが⽰された。 後根神経節(DRG)に細胞体を有する感覚神経、以下 DRG 神経は⽪膚にお ける痒みや痛みの知覚に必須である。DRG 神経の中で、C 繊維と呼ばれる細い 神経軸索を有する神経上にはカチオンチャネルの⼀種である TRPV1(Transient receptor potential V1)が発現する。マウスに TRPV1 の強⼒なアゴニストを投 与することで TRPV1 陽性 DRG 神経を除神経すると、アトピー性⽪膚炎や接触 性⽪膚炎時の痒みが減弱することが報告されている(29, 30)。そこで、oxDLPG、
oxDLPS 依存的な痒みが TRPV1 陽性 DRG 神経を介するかどうか検討した。 TRPV1 アゴニストであるカプサイシンの段階的な投与により、除神経処置を実 施した(22)。TRPV1 陽性神経の除神経により、対照群で認められた oxDLPG、 oxDLPS 依存的な痒み⾏動が有意に減弱した(Fig. 14)。
⽪膚において DRG 神経が活性化すると、その投射先である脊髄後⾓の最外 層、laminae I-Ⅱ(⻘枠部位)において、神経活性化マーカーである c-fos mRNA 発現が上昇する(21)。そこで、oxDLPG ⽪下投与 30 分後、脊髄切⽚を作製し、 laminae I-Ⅱにおける c-fos mRNA の発現を in situ hybridization 法を⽤いて調 べた。対照群では c-fos mRNA はほとんど検出されなかったが、その⼀⽅、 oxDLPG 投与群では顕著な c-fos mRNA 発現誘導が認められた(Fig. 15)。 以上の結果から、oxDLPG、oxDLPS は TRPV1 陽性 DRG 神経の活性化を介 して痒みを誘導することが⽰唆された。 3-4. 新規掻痒性酸化リン脂質の痒みシグナルの解析 次に、oxDLPG、oxDLPS の標的分⼦について検討するため、シグナル分⼦の 阻害剤を⽤いた解析を⾏った。TRPV1 は TRPV1 陽性 DRG 神経上に発現 す るある種の GPCR 下流シグナル伝達に重要であることが報告されている(21, 31)。神経上の GPCR に作⽤して痒みを誘導する薬物(ヒスタミンなど)は主に Gq/11タンパク質を介し、ホスホリパーゼ C(PLC)の活性化を誘導する。その 後、TRPV1 などのチャネル細胞内リン酸化部位をリン酸化し、活性化を誘導す ることで神経活動を促進する。そこで、Gq/11、PLC、TRPV1 阻害剤処置 30 分 後の oxDLPG 依存的な痒み作⽤を評価した。上記の阻害剤の本系での有効性は、 ヒスタミン依存的な痒みに対する阻害作⽤を指標に確認した。oxDLPG 依存的 な痒みはこれら阻害剤により有意に減弱した(Fig. 16)。他の分⼦を介する可能
性も考えられるが、少なくとも、oxDLPG は Gq/11共役型 GPCR に作⽤し、下 流で PLC、TRPV1 を活性化することで DRG 神経の活性化を誘導することが⽰ 唆された。 3-5. 新規掻痒性酸化リン脂質の受容体の探索 先の解析から、oxDLPG(および oxDLPS)は DRG 神経上に発現する GPCR に 作⽤すると想定した。約 20 年前、DRG に⾼発現するリガンド未知(オーファ ン)GPCR 集団として、Mas-related G protein-coupled receptors(Mrgprs)が 同定された(32)。Mrgprs はヒト 8 種類、マウス 24 種類の遺伝⼦から構成され ており、現在までに DRG 神経上のヒト MRGX1、MRGX4、およびマウス MrgA1、 MrgA3、MrgC11、MrgD が起搔物質による痒みに関与することが報告されてい る(Fig. 17)(33, 34, 35, 36, 37)。そこで、oxDLPG、oxDLPS に応答しうるマ ウス Mrgprs を GPCR 活性化検出法である TGFa切断アッセイにより探索した。 すなわち、20 種のマウス Mrgprs を個々に HEK293 細胞に過剰発現し、oxDLPG または oxDLPS 刺激を加えた際に切り出された TGFa量を評価した。興味深い ことに、20 種のマウス Mrgprs の中で、MrgB5 のみが oxDLPG、oxDLPS 依存 的に活性化した(Fig. 18)。 次に、先に検証した様々な酸化リン脂質による痒み作⽤が MrgB5 を介するも のであるかを簡易的に検証するため、これらの MrgB5 に対する活性を⽐較した。 5 種の極性基をそれぞれ有する oxDLPL の中で、oxDLPG、oxDLPS は濃度依 存的に MrgB5 活性化を誘導した⼀⽅、痒み誘導が検出されなかった oxDLPA、 oxDLPC、oxDLPE は MrgB5 活性化を誘導しなかった。さらに、様々な脂肪酸 を有する PG 酸化物について同様の検討を⾏ったところ、di-18:2 PG(DLPG)、 di-18:3 PG、di-20:4 PG、di-22:6 PG 酸化物のみが MrgB5 を濃度依存的に活性
化した。また、空気酸化処理を⾏っていない DLPG、酸化リノール酸(9-HODE、 13-HODE)、酸化リノール酸含有 LysoPG(oxLysoPG (18:2))は MrgB5 をほ とんど活性化しなかった(Fig. 19)。 oxDLPG は Gq/11タンパク質依存的に痒みを誘導する。TGFa切断アッセイは Gq/11 および G12/13共役型受容体の活性化を検出することが可能である(12)。⾔ い換えると、TGFaの切断は Gq/11および G12/13タンパク質の活性化に依存する。 当研究室では、Gq/11⽋損 HEK293 細胞、G12/13⽋損 HEK293 細胞、Gq/11/12/13⽋ 損細胞を所有している。そこで、これら⽋損細胞に MrgB5 を過剰発現し、 oxDLPG 刺激後の TGFa切断を評価した。興味深いことに、野⽣型細胞(Parent) で認められた oxDLPG 依存的な MrgB5 活性化は、G12/13⽋損細胞(G12/13 KO) において同等に検出されたが、Gq/11⽋損細胞(Gq/11 KO)、および Gq/11/12/13⽋損 細胞(Gq/11/12/13 KO)では、顕著な減弱を認めた。また、Gq/11/12/13⽋損細胞に Gq または G11タンパク質を過剰発現すると oxDLPG 依存的な MrgB5 活性化が野 ⽣型と同等に検出された。以上より、MrgB5 は oxDLPG により活性化し、Gq/11 シグナルを伝達しうることが⽰唆された(Fig. 20)。 以上の結果から、特定の酸化リン脂質は MrgB5 活性化を介し、Gq/11シグナル を誘導することが明らかになり、酸化リン脂質依存的な痒み作⽤の標的候補と して MrgB5 が挙げられた。 3-6. MrgB5 発現細胞の解析 マウス 15 臓器における MrgB5 mRNA 発現分布を qRT-PCR 法により確認し た。すでに報告されているように、MrgB5 mRNA は DRG や迷⾛神経節(Nodose ganglion)のような感覚神経節に検出された(32, 38)。また、意外なことに、複 数の免疫組織においても MrgB5 mRNA 発現が認められた(Fig. 21)。
次に、DRG 神経において、MrgB5 mRNA 発現細胞をin situ hybridization 法 により調べた。MrgB5 mRNA は、⽐較的多くの細胞体において検出された。さ らに、連続切⽚を⽤い、TRPV1 mRNA 陽性細胞において MrgB5 mRNA が検出 されるかどうか調べたところ、おそらく同⼀細胞と思われる細胞体上(⻘⽮頭で 表記)に MrgB5、TRPV1 mRNA が検出された(Fig. 22)。今後、同⼀切⽚上で の多重染⾊などにより、より詳細な解析を進めたい。 以上より、MrgB5 は oxDLPG の作⽤標的細胞であると想定される TRPV1 陽 性 DRG 神経に⾼発現することが強く⽰唆された。 3-7. MrgB5 KO マウスの作製 個体レベルにおける MrgB5 の機能について解析するため、MrgB5 KO マウス の作製を⾏った。MrgB5 KO は CRISPR-Cas9 法を⽤いて作製した。変異個体の MrgB5 機能消失を確認するため、変異個体ゲノムから MrgB5 配列をクローニ ングし、HEK293 細胞に発現させ、oxDLPG 応答性を TGFa 切断アッセイで評 価した。2 種類の変異体由来 MrgB5 配列(KO #1、#2 と記載)はいずれも野⽣ 型マウス由来 MrgB5 で認められた oxDLPG 応答性が完全に消失することが確 認された(Fig. 23)。なお、以降の解析には基本的に KO #1 ラインを⽤いた。 3-8. MrgB5 の oxDLPG 依存的痒み・DRG 神経活性化に及ぼす影響 oxDLPG 依存的な痒みが MrgB5 を介するかどうか検証した。すなわち、野⽣ 型、MrgB5 KO マウスうなじ部分に oxDLPG を⽪下投与した後 30 分間の後脚 による掻き⾏動を評価した。MrgB5 KO マウスでは、野⽣型で認められる掻き ⾏動が有意に減弱した。なお、本結果は、KO #2 でも確認された(Fig. 24)。 また、oxDLPG 依存的 DRG 活性化に MrgB5 が関与するかどうか調べた。野
⽣型、および MrgB5 KO マウスに oxDLPG ⽪下投与 30 分後、脊髄切⽚を作製 し、laminae I-Ⅱにおける c-fos mRNA の発現を in situ hybridization 法を⽤い て調べた。MrgB5 KO マウスでは、野⽣型で認められた oxDLPG 投与による顕 著な c-fos mRNA 発現誘導が減弱した(Fig. 25)。
以上の結果から、oxDLPG 依存的な痒み⾏動や、DRG 神経の活性化は、MrgB5 を介した現象である可能性が強く⽰唆された。 3-9. MrgB5 のマスト細胞依存的痒みに及ぼす影響 本研究で作製した MrgB5 KO マウスにおいて、ヒスタミンやマスト細胞性の 分泌物依存的な痒み作⽤を解析した。野⽣型、および MrgB5 KO マウスにヒス タミン、または、マスト細胞脱顆粒誘導剤である compound 48/80 を⽪下投与 し、痒み作⽤を解析した。いずれの物質を投与した際も、野⽣型、MrgB5 KO マ ウス共に同等の掻き⾏動が認められた(Fig. 26)。したがって、MrgB5 はヒスタ ミン依存的な痒み経路には関与しないことが⽰唆された。 3-10. ⽪膚炎依存的痒みにおける MrgB5 の機能解析 2009 年、Dong らは Mrgprs の機能を明らかにすることを⽬的として、MrgB5 を含む 12 種の Mrgprs 遺伝⼦(MrgA1、A2、A3、A4、A10、A12、A14、A16、 A19、B4、B5、C11)を同時に KO した MRG cluster KO マウスを作製した(34)。 興味深いことに、MRG cluster KO マウスは 3 つの⽪膚掻痒モデル(乾⽪症、 OVA 誘導性アレルギー⽪膚炎、接触性⽪膚炎)において、痒み⾏動の減弱が報 告された(25)。現在のところ、単独の Mrgprs 遺伝⼦の⽋損で MRG cluster KO マウスの表現型が確認された報告はない。そこで、MrgB5 が上記病態時に機能 するかどうか検討を⾏った。
野⽣型マウスと MrgB5 KO マウスに乾⽪症、OVA 誘導性アレルギー⽪膚炎 を発症させ、⼀定時間内における掻き⾏動を評価したところ、両群に有意な差は 認められなかった(Fig. 27)。ところが、野⽣型マウスと MrgB5 KO マウスに DNFB 誘導性の接触性⽪膚炎を発症させたところ、MrgB5 KO マウスは野⽣型 マウスと⽐較し、痒み症状が顕著に減弱した(Fig. 28)。MrgB5 KO で認められ た痒み症状が炎症に付随した現象であるかどうか検討した。⽪膚炎を惹起して から継時的に病態部⽪膚をサンプリングした。採取した⽪膚について、炎症マー カーである IFNg mRNA 量を qRT-PCR 法、および組織切⽚を作成し、⽪膚の肥 厚を測定した。その結果、ほぼ同等の炎症応答が確認された(Fig. 29)。したが って、MrgB5 は炎症ではなく、痒みそのものに寄与することが⽰唆された。な お、接触性⽪膚炎病態時の DRG における MrgB5 mRNA 発現解析を⾏ったとこ ろ、病態未発症時と同等の発現量が認められた(Fig. 30)。 3-11. 接触性⽪膚炎時における酸化抑制による痒み作⽤解析 先の解析から、MrgB5 が接触性⽪膚炎時に何らかの内在性リガンドを認識し、 痒みを誘導することが⽰唆された。これまでに検証した限り、in vitro において MrgB5 はリン脂質酸化物のみに応答するため、⽪膚炎時にも掻痒性酸化物が産 ⽣され、MrgB5 を活性化しているのではないかと仮定した。そこで、2 種類の 抗酸化剤、N-アセチルシステイン(NAC)、a-Tocopherol(a-Toc)の継続的投 与を⾏い、接触性⽪膚炎時の痒み症状を評価した。興味深いことに、これら抗酸 化剤を投与した群では、接触性⽪膚炎において、痒みの減弱傾向が認められた (Fig. 31)。抗酸化剤は炎症も抑制することが報告されているため、痒み特異的 な影響ではないと考えられるが、少なくとも接触性⽪膚炎時には酸化反応が起 こり、掻痒性酸化物が産⽣されていることが⽰唆された。
3-12. 接触性⽪膚炎部位の抽出物の MrgB5 活性解析 MrgB5 KO マウスで痒み症状が認められる際に、病態部に MrgB5 内在性リガ ンドが存在するかどうか検討した。⽪膚炎を惹起し、経時的に病態部⽪膚をサン プリングした。採取した⽪膚から古典的な脂質抽出法である Folch 法を⽤いて、 ⽔溶性(メタノール/⽔に可溶)、脂溶性成分(メタノール/クロロホルムに可溶) を抽出した。抽出物の溶媒を留去し、0.01% BSA 含有 HBSS 液に懸濁し、TGFa 切断アッセイにより、MrgB5 に対する活性を評価した。⽔溶性画分には MrgB5 活性が認められなかった⼀⽅で、脂溶性画分では、惹起後⽪膚炎の進⾏に伴って MrgB5 活性が増加した(Fig. 32, 33)。このことから、接触性⽪膚炎時には、MrgB5 を活性化しうる脂質分⼦が産⽣されていることが明らかになった。今後、カラム クロマトグラフィーなどの分画法により、接触性⽪膚炎抽出物を細かく分画、 MrgB5 活性評価・質量分析を⾏い、MrgB5 内在性リガンドの構造同定、⽣体中 での検出を⽬指したい。 3-13. 接触性⽪膚炎で機能しうる Mrgprs の探索 Mrgprs はヒトには 8 種類しか存在しない⼀⽅で、マウスには 24 種もの遺伝 ⼦が存在する。ヒト 8 種類のうち、マウスにオルソログを有する遺伝⼦は MRGD、 MRGE、MRGF、MRGG の 4 種類であり、MRGX1、MRGX2、MRGX3、MRGX4 にはマウスにオルソログが存在しない(35)。また、MrgB5 は齧⻭類にのみ保存 された遺伝⼦であり、ヒトにオルソログが存在しない。本研究において、接触性 ⽪膚炎時に MrgB5 内在性リガンドが産⽣され、痒みを誘導することが⽰唆され たが、ヒトにおいて同様の機能を有する Mrgprs は不明である。ヒト患者の⽪膚 病変部脂質を⼊⼿することは困難であるため、先の解析で使⽤したマウス接触
性⽪膚炎抽出物に対して反応するヒト Mrgprs を探索した。8 種類のヒト Mrgprs を個々に HEK293 細胞に過剰発現し、接触性⽪膚炎抽出物で刺激した際の応答 性を TGFa切断アッセイで評価した。MRGD、MRGE、MRGF、MRGG、MRGX1 は応答性が認められなかった⼀⽅、MRGX2、MRGX3、MRGX4 について、接 触性⽪膚炎抽出物応答性が認められた(Fig. 34)。MRGX2 はマスト細胞上に発 現することが知られており、ポリアミン類やペプチド性のリガンドを有すると 報告されている(39)。MRGX3 はリガンド報告が全くなく、MRGX4 は胆汁酸や 胆汁⾊素であるビリルビンに応答することが報告されている(36, 37)。これら受 容体が接触性⽪膚炎病態に関わるという報告はないが、MrgB5 と類似の機能を 有している可能性が⽰唆された。なお、MrgB5 が既知の MRGX2 リガンド (cortistatin-14、compound 48/80)、および MRGX4 リガンド(胆汁酸)に応 答しないことは確認済みである(data not shown)。
マウス Mrgprs は配列相同性が⾼く、機能的に相補しあう可能性が考えられ る。先に述べた MRG cluster KO マウスは接触性⽪膚炎時の痒み⾏動が野⽣型 マウスと⽐較して減弱するが、痒み⾏動が完全には消失しない。このことから、 上記マウスで KO されていないマウス Mrgprs が接触性⽪膚炎時の痒みに関与 する可能性が考えられた。そこで、ヒト同様の⼿法でマウス Mrgprs の接触性⽪ 膚炎抽出物応答性を検証した。その結果、新たに MrgA7 と MrgB3、MrgG が接 触性⽪膚炎抽出物によって活性化することが明らかになった(Fig. 35)。これら 受容体はいずれも機能に関する報告はないが、MRG cluster KO マウスでは⽋損 しておらず、接触性⽪膚炎時に MRG cluster KO マウスに残存する痒み⾏動を 説明できる可能性がある。
4. 考察
リノール酸は表⽪において最も豊富な PUFA であり、遊離脂肪酸として以外 にも、リン脂質に取り込まれた状態で存在する(16)。リノール酸が酸化反応を受 けることにより産⽣される多様な酸化リノール酸のうち、特定の分⼦種はマウ ス個体に痒みを誘導することが⽰されているが、その分⼦機構は不明である (18)。私は、酸化リノール酸による痒みの⼀部は酸化リノール酸を取り込んだリ ン脂質によるものであると仮定し、酸化リノール酸含有リン脂質の痒み作⽤を 検証した。本研究において、私は、特定の酸化リン脂質が⼀次求⼼性感覚神経で ある TRPV1 陽性 DRG 神経を介して、酸化リノール酸よりも強⼒な痒み知覚を 惹起するという、新規の痒み機構を明らかにした。さらにその標的分⼦として、 Mas-related G protein-coupled receptors(Mrgprs)に属する機能未知受容体 MrgB5 を同定した。酸化リノール酸を投与した MrgB5 KO マウスは野⽣型マウ スと同等の掻き⾏動を⽰したことから、当初の想定と異なり、私が同定した酸化 リン脂質による痒み機構は酸化リノール酸依存的な痒みとは異なるものである と考えられた(Fig. 36)。 Mrgprs は 2001 年に Dong らによって、DRG 神経に⾼発現する、ヒト 8 種、 マウス 24 種からなるオーファン GPCR ファミリーとして同定された(32)。そ のうち⼀部のメンバーについてはある種の神経ペプチド、ポリアミド、⼩分⼦化 合物をリガンドとすることが知られていた(34, 35, 39, 40)。これまでに、Mrgprs がリン脂質をリガンドとするという報告はなされていない。興味深い点として、 MrgB5 は未酸化状態のリン脂質を認識せず、酸化されたリン脂質を特異的に認 識した。酸化脂質は掻痒性疾患を含む様々な病態で検出されている(41)。実際に、 リノール酸を含めた様々な PUFA の酸化物が乾癬患者の⽪膚のうち、痒みを感じる部位で顕著に増加する(18)。また、最近、アラキドン酸の酸化物である、5-oxoeicosatetraenoic acid(5-oxoETE)が Mrgprs のメンバーである MrgD のリ ガンドとして、過敏性腸疾患(IBS)時の痛覚過敏に関わることが報告された(42)。 本研究と併せ、Mrgprs のリガンドとして、酸化脂質が有望な候補となりうるこ とが⽰唆された。⼀⽅で、酸化リン脂質は⽣体において、極めて微量であり、特 定の分⼦種を除き、解析された例がほとんどない。今後、質量分析技術の進歩に より、掻痒性⽪膚疾患部位における酸化リン脂質の網羅的な解析が⾏われ、掻痒 関連酸化リン脂質が明らかになることが期待される。 マウス Mrgprs のうち、MrgA1、MrgA3、MrgC11、MrgD の 4 種類は既に DRG 神経上で痒み誘導に関与すると知られている。各メンバーのリガンドが MrgB5 に対して活性を持たないことは私の検討を含め、既に報告されている (data not shown)。結果の項でも述べたように、Mrgprs の 12 個の遺伝⼦(MrgA1、 A2、A3、A4、A10、A12、A14、A16、A19、B4、B5、C11)を⽋失した MRG cluster KO マウスは乾⽪症、アレルギー性⽪膚炎、接触性⽪膚炎病態時に発症 する痒みが野⽣型マウスと⽐較して顕著に減弱する(25)。しかしながら、12 個 の Mrgprs 遺伝⼦のうち、どの遺伝⼦が上記の表現型の責任因⼦であるかについ ては全く不明であった。MrgB5 の単独 KO マウスは乾⽪症、アレルギー性⽪膚 炎において野⽣型マウスと同等の痒み⾏動を⽰した。⼀⽅で、接触性⽪膚炎病態 時、MrgB5 KO マウスは MRG cluster KO マウスと同様に、有意な痒み⾏動の 減少が認められた。MrgB5 KO マウスの表現型がなぜ接触性⽪膚炎時にのみ認 められるのかどうかは定かではないが、私は各掻痒性⽪膚疾患における痒みの 知覚は異なる Mrgprs メンバーとそのリガンドの組み合わせによって制御され ていると推測している。乾⽪症では、免疫細胞がほとんど表⽪に浸潤しないが、 アレルギー性掻痒と接触性⽪膚炎では多数の免疫細胞の浸潤が認められる(43)。
また、アレルギー性掻痒では、主にマスト細胞、好酸球に起因する Th2 型優位 な免疫応答が起こる⼀⽅で、接触性⽪膚炎では、主に Th1 型の免疫応答が優位 である(44, 45)。従って、MrgB5 リガンドは Th1 型の免疫細胞そのもの、ある いは Th1 型の免疫細胞と表⽪細胞の何らかの相互作⽤により産⽣されると想定 される。 本研究で検討した酸化リン脂質のうち、リノール酸(18:2)、a-リノレン酸 (18:3)、アラキドン酸(20:4)、ドコサヘキサエン酸(22:6)を分⼦内に 2 本有 するホスファチジルグリセロール(PG)酸化物はいずれもほぼ同等、かつ最も 強⼒な痒み誘導活性と MrgB5 活性を有した。DRG 神経上の MrgB5 が機能する 部位は⽪膚であること、また、⽪膚(特に表⽪)に存在する PUFA のほとんど がリノール酸であること、接触性⽪膚炎を発症した⽪膚中の脂質を LC-MS/MS に よ り 測 定 す る と 、 リ ノ ー ル 酸 を 2 本 有 す る PG ( DLPG; di-linoleoyl phosphatidylglycerol)が顕著(かつ特異的)に増加した(Fig. 37)ことから、検 討したものの中で、oxDLPG が MrgB5 の内在性リガンドである可能性が⾼いと 考えられた。本研究で⽤いた oxDLPG は DLPG を空気酸化処理したものであ り、様々な酸化様式の oxDLPG からなる混合物である。そこで、当研究室の吉 ⽥美沙紀学⼠を中⼼に、oxDLPG 中の MrgB5 活性本体の構造同定、および、接 触性⽪膚炎病態部における検出を試みた(Fig. 38)。その結果、DLPG の 2 本の リノール酸のうち、⽚⽅は未酸化状態であり、もう⽚⽅がヒドロキシ基、ケトン 基、エポキシ基、ハイドロキシペルオキシド基修飾のいずれかを受けた酸化リノ ール酸を有することが⽰唆された(Fig. 39)。しかし、これまでの解析では、接 触性⽪膚炎病態部において上記分⼦は検出されていない。 上記のように、酸化リン脂質が⽣体内で MrgB5 リガンドとして痒みを誘導す るかどうかは未だ不明である。興味深いことに、NAC や a-Toc といった抗酸
化剤を接触性⽪膚炎マウスに投与しておくと、痒み⾏動が減弱した。従って、接 触性⽪膚炎時には何かしらの掻痒性酸化物が産⽣されることが⽰唆された。さ らに、直接的に MrgB5 リガンドの性状に迫るためのアプローチとして、⽪膚炎 部位からの抽出物の MrgB5 活性を評価する実験系を構築した。古典的脂質抽出 法である Folch 法により、⽔溶性、脂溶性画分を回収し、MrgB5 活性を評価す ると、⽪膚炎未発症時には⽔溶性、脂溶性画分の両⽅で、ほとんど活性は検出さ れなかった。⼀⽅で、接触性⽪膚炎部位からの抽出物では、脂溶性画分で顕著な MrgB5 活性が認められた。従って、MrgB5 リガンドは何らかの脂質成分である ことが強く⽰唆された。今後、MrgB5 リガンドが酸化依存的に産⽣されるかど うか検討するため、抗酸化剤を投与した接触性⽪膚炎発症マウスからの抽出物 の MrgB5 活性について評価する必要がある。また、本抽出物を液体クロマトグ ラフィーなどにより分画し、活性画分を絞り込み、質量分析器を⽤いて構造を特 定することで、MrgB5 の内在性リガンドの同定が期待される。 Mrgprs 研究における問題点として、ヒトとマウス遺伝⼦の相同性の低さに起 因する種間の実験結果の translation の難しさが挙げられる。実際に、ヒトにお いて、MrgB5 に相当する遺伝⼦は報告されていない。ヒト遺伝⼦ 8 種のうち、 MRGD、MRGE、MRGF、MRGG の 4 種のみが配列上極めて相同性の⾼いマウ スオルソログを有する。⼀⽅、ヒト MRGX1、MRGX2、MRGX3、MRGX4 には マウスオルソログが存在しない(35)。しかし、興味深いことに、Dong らは、リ ガンド活性を指標とした評価により、MRGX1 がマウス MrgA3、および、MrgC11 と、MRGX2 がマウス MrgB2 と、また、MRGX4 がマウス MrgA1 と同⼀のリ ガンドを認識することを明らかにした(Fig. 40)(34, 36, 39)。このように、Mrgprs は配列の相同性が低い、異なる遺伝⼦間であっても、機能的ホモログとして、同 ⼀のリガンドを認識し、機能する可能性が想定される。そこで、本研究では接触
性⽪膚炎抽出物を⽤いて、⽪膚炎時に MrgB5 と同様に機能しうるヒト Mrgprs の候補分⼦を探索した。その結果、MRGX2、MRGX3、MRGX4 が⽪膚炎時の 抽出物依存的に活性化することが明らかになった。さらなる検証が必要ではあ るが、接触性⽪膚炎時には、これらヒト Mrgprs が関与する可能性が⽰唆された。 今後、接触性⽪膚炎などのヒト慢性掻痒性⽪膚疾患患者の⽪膚サンプルから抽 出した脂質を⽤いることで、これらヒト Mrgprs がヒト病態時にも機能しうるか どうか検討が可能となると期待している。また、オーファン GPCR がほとんど である Mrgprs は当然ながら阻害剤を持たないメンバーがほとんどである。今 後、疾患関連 Mrgprs 遺伝⼦が同定された場合、本研究で構築した⼿法を応⽤す ることで、仮に内在性のリガンドが不明である場合でも鎮搔薬のシード化合物 の探索が可能となるかもしれない。ヒト⽪膚炎患者サンプルを⼿術などの侵襲 的⽅法により採取することは困難であるが、共同研究ベースで、テープストリッ ピングにより、⽐較的⾮侵襲的に患者⽪膚サンプルが回収できるようになって いる。このようにして回収した脂質は微量であることが想定され、今後、TGFa 切断アッセイのスケールダウン(384 well plate などの利⽤)を含め、系の改善 が必要となる。 当研究室の⽯⿊純修⼠により、ヒト MRGX4 は極めて酸化反応が進んだ短鎖 型の酸化リン脂質に強く応答することが⽰されていた(46)。この結果は、MrgB5 が好む酸化リン脂質構造(軽度の酸化を受けた⻑鎖型の酸化脂肪酸を有するリ ン脂質)とは異なっていた。⼀⽅で、空気酸化により調製した oxDLPG は MRGX4 を活性化することが明らかになった(Fig. 41)。以上から、MrgB5 と MRGX4 のリガンド認識は必ずしも⼀致していないが、類似のリガンドを認識 する可能性がある。結果の項でも述べたが、最近、MRGX4 が胆汁うっ滞時の痒 みに関わることが⽰された。すなわち、胆汁うっ滞時に⾎中濃度が⾶躍的に上昇