バ イ オ イ メ ジ ン グ に よ る 筋 細 胞 内 カ ル シ ウ ム イ オ ン 動 態 の 評 価
曽野 部 崇
二
〇
〇 九 年
バイオイメージングによる筋細胞内カルシウムイオン動態の評価
曽 野 部 崇
電気通信大学
2009 年 3 月
バイオイメージングによる筋細胞内カルシウムイオン動態の評価
曽 野 部 崇
電気通信大学大学院 電気通信学研究科 博士(理学)の学位申請論文
2009 年 3 月
バイオイメージングによる筋細胞内カルシウムイオン動態の評価
博士論文審査委員会
主査 狩野 豊 准教授
委員 丹羽 治樹 教授
委員 山田 幸生 教授
委員 中村 整 教授
委員 白川 英樹 准教授
著作権所有者
曽 野 部 崇
2009
Evaluation of intracellular calcium ion in skeletal muscle using in vivo Bio-imaging
Takashi Sonobe
Abstract
The present study describes that to establish the methods of measurement intramyocyte Ca2+ level under in vivo condition. I used rat spinotrapezius muscle with fluorescence imaging technique, and succeeded to observe in vivo skeletal muscle Ca2+
level. Because of the difficulty in measuring [Ca2+]i under in vivo conditions, almost all investigations of [Ca2+]i have been performed under in vitro conditions.
Bioimaging techniques can visualize intracellular ions directly, and, whereas measurements of [Ca2+]i are potentially feasible under intravital conditions routinely used for microcirculation studies, to date, because skeletal muscle was considered to be too thick for microscopy, most studies of [Ca2+]i have used isolated or cultured single myocytes. Unfortunately, such isolated or cultured cells have quite a different environment than in vivo skeletal muscle with respect to their absence of a microcirculation, different oxygen and substrate availabilities, and metabolism, among other considerations. The purpose of present investigation was to test the original challenges in the spinotrapezius muscle, and I demonstrated following results.
1) Compared with the surgically excised in vitro spinotrapezius, the in vivo preparation (i.e., exteriorized, as for intravital miscroscopy) prolonged (90 min) observation at rest did not elevate [Ca2+]i. 2) An extended series of eccentric contractions elevated
[Ca2+]i to a greater extent than isometric contractions. Moreover, the elevated [Ca2+]i accompanying ECC contractions was prevented or substantially reduced by the SAC blockers. 3) Females maintain [Ca2+]i homeostasis following novel eccentric contractions whereas Males do not which is consistent with a role for elevated [Ca2+]i in eccentric exercise-induced muscle damage. 4) Diabetes muscles show vulnerability of [Ca2+]i homeostasis for increase in oxygen demand during isometric contraction.
バイオイメージングによる筋細胞内カルシウムイオン動態の評価
曽 野 部 崇
概 要
骨格筋は,細胞内カルシウムイオン(Ca2+)濃度変化によって収縮・弛緩が制御され ている.弛緩状態の筋細胞内Ca2+濃度は0.1µM以下に保たれているが,活動電位に よって細胞内 Ca2+が一時的に増加することで筋収縮が可能となる.ところが,筋細胞 内 Ca2+濃度の恒常性が崩れると筋疲労や筋損傷が誘発されると考えられている.筋 疲労や筋損傷の機序を考える場合,細胞内の Ca2+動態を制御する機構の解明が重 要である.しかしながら,筋細胞内 Ca2+動態を調べた先行研究のほとんどは単離細胞 や培養細胞を用いた生体外(in vitro)の実験である.生体内(in vivo)でのCa2+動態は 血流変動にともなう酸素供給レベルや筋収縮様式によって大きな影響を受けると考え られる.そこで本研究では,生体内環境下(in vivo)での Ca2+動態を筋細胞レベルで 評価するリアルタイムバイオイメージング技法を確立し,筋収縮時の筋細胞内 Ca2+動 態の制御機構を明らかにすることを目的として,以下の研究課題を遂行した.
本論文は以下の8章から構成されている.第1章から第2章では,筋細胞内におけ る Ca2+による細胞制御メカニズムについて述べ,これまでに行われてきた細胞内 Ca2+
測定に関する研究,および近年のバイオイメージング技術の発展とその応用について 文献研究を行った.さらに,第3章においてin vitro研究における問題点を指摘した上 で解決のための課題を設定し,同時に,これまでの研究において解明されていない問 題に対する課題を設定した.第 4章から第6 章では,ラットを用いたin vivoにおける 筋細胞内 Ca2+観察の評価法の検討(第4章),筋収縮時に生じる筋細胞内Ca2+変化 の検討と,その変化に寄与する経路の解明(第 5 章),筋モデルの違いが筋収縮時に
生じる筋細胞内Ca2+変化に及ぼす影響(第6章)の大きく3つの研究課題について記 した.以下にその概要を示す.
研究課題1:生体内環境下(in vivo)において筋細胞内Ca2+を測定することを目的とし て,ラット脊柱僧帽筋を観察対象としたバイオイメージング法を用いて,筋細胞内 Ca2+
変化を検討した.その結果,in vitroでは時間経過と共に細胞内Ca2+増加が観察され たが,血流を維持した in vivo においては細胞内 Ca2+は安静レベルを維持していた.
一方で,薬理的刺激によるCa2+変化はin vivo条件においても可能であり,さらにこの 動態を細胞レベルで評価することに成功した.このことから,in vivoでの細胞内Ca2+評 価モデルは,筋細胞内 Ca2+の恒常性を維持できること,さらに,細胞内 Ca2+変動を筋 線維毎に測定できることが明らかになった.
研究課題2(1):in vivoにおける筋細胞内Ca2+観察が可能であったことを受け,脊柱僧
帽筋に対して筋収縮刺激を負荷し,その後の細胞内Ca2+変化を測定することを目的と した.収縮様式の異なるアイソメトリックおよびエキセントリック収縮を行い,細胞内の Ca2+変化を測定した.その結果,エキセントリック収縮においてはアイソメトリック収縮時 と比べて細胞内 Ca2+の有意な増加を示した.このことは,増加した細胞内 Ca2+が,エ キセントリック収縮によって引き起こされる筋損傷に関与するという知見を示唆するもの であった.
研究課題2(2):研究課題2(1)を受け,ここではエキセントリック収縮後のCa2+増加経路
を検討することを目的とした.細胞伸張に応答を示すストレッチ感受性イオンチャネル
(SAC)に着目し,その阻害薬を用いることでエキセントリック収縮時に生じる Ca2+蓄積 へのSACの関与を検討した.その結果,SAC阻害薬はエキセントリック収縮後のCa2+
蓄積を有意に抑制することが明らかになった.この結果は,エキセントリック収縮には SACを介したCa2+蓄積が生じていることを示すものであった.
研究課題 3:研究課題1ならびに2の実験モデルを発展させ,ここでは筋収縮による細 胞内 Ca2+応答についての性差および糖尿病の影響を調べた.その結果,オスと比較 してメスではエキセントリック収縮に対して Ca2+変化が少ないことが明らかになった.ま た,糖尿病の筋ではアイソメトリックに対する細胞内 Ca2+蓄積が大きいことが明らかに なった.
第7章および第8章では,本研究モデルの利点と方法論的な問題点を明らかにす るとともに,各研究課題によって得られた成果を検討し,in vivoの細胞内におけるCa2+
変化と筋収縮との関連性について総括した.さらに今後の課題として,脊柱僧帽筋を 対象としたin vivo観察の空間的,時間的解像度の向上の必要性について言及した.
最後に,本研究の結論として,in vivoバイオイメージングにおける筋細胞内 Ca2+測 定が,生体内環境下での様々な細胞内現象を観察することに非常に有効な手法であ ることを示した.
目次
I. 緒言 ・・・1
II. 文献研究 ・・・4
III. 研究目的および研究課題 ・・・21
IV. 生体内(in vivo)環境下における
骨格筋細胞内Ca2+-kineticsの可視化(研究課題1) ・・・23
V. 筋収縮様式の違いに着目した
骨格筋細胞内Ca2+-kineticsの差異(研究課題2)
(1)様式の異なる筋収縮負荷後の筋細胞内Ca2+-kineticsの測定 ・・・43
(2)伸張性筋収縮で生じるCa2+蓄積に寄与するCa2+増加経路の検討
・・・57
VI. 骨格筋in vivoモデルを用いた細胞内Ca2+測定の発展的応用(研究課題3)
(1)性差によって生じる筋機能の違いと細胞内Ca2+動態の関連について
・・・67
(2)糖尿病がもたらす筋機能の低下と細胞内Ca2+動態の関連について
・・・77
VII. 討論 ・・・86
VIII. 総括 ・・・91
謝辞 ・・・93
参考文献 ・・・94
本論文は,以下の論文および未発表の実験結果を加筆,修正して構成したも のである.
1. Takashi Sonobe, Tadakatsu Inagaki, David C. Poole, and Yutaka Kano.
Intracellular calcium accumulation following eccentric contractions in rat skeletal muscle in vivo: role of stretch-activated channels. Am J Physiol Regul Integr Comp Physiol. 294(4): 1329-1337 (2008)
I. 緒言
1. 細胞内におけるカルシウムイオン制御機構
生物の細胞質内では,多様な細胞機能を実現するための情報伝達が常に行わ れている.情報伝達経路において化学物質がメッセンジャーとして機能してい ることはよく知られており,その中でも,2価の陽イオンであるカルシウムイオ ン(Ca2+)を情報伝達の要として用いている制御機構は数多く存在している.骨 格筋において,その重要な収縮機構の信号は電気的興奮によって伝えられ,最 終的には Ca2+による情報伝達を受けて機能する.骨格筋収縮の過程では,筋を 構成するタンパク質(トロポニン)と Ca2+との結合が張力発生の起点であるこ とがわかっており,Ca2+は筋収縮のトリガーであるといえる.骨格筋の収縮には,
神経から筋膜上を伝播してきた電気的情報を受けた筋小胞体からの Ca2+放出に よる細胞内 Ca2+濃度の瞬間的な上昇が不可欠であり,これらの細胞内 Ca2+上昇 によってトロポニンが Ca2+と結合し,はじめてエネルギーを利用した筋の張力 発揮が起こる.通常,細胞内のCa2+濃度は,筋小胞体によるCa2+再取り込みや,
細胞外へ Ca2+を排出するイオン輸送体(イオン交換体)の働きにより,細胞外 よりも低い状態(0.1 µΜ以下,Armstrong et al. 1991)に厳密に維持されている.
これはわずかな濃度変化にも応答できるような Ca2+に対する細胞内の感受性を 非常に高めるとともに,イオンチャンネルが開いたときにイオン濃度勾配によ る細胞外からのイオン流入を容易にしている.このような細胞内の一時的な Ca2+濃度増加と減少を繰り返すことによって,筋は収縮-弛緩することができる.
このように骨格筋においては筋細胞内 Ca2+濃度の変化がその機能に大きく関与 しており,筋機能を検討する際には細胞内の Ca2+動態を知ることが非常に重要 である.
2. 細胞内Ca2+の測定における課題と問題点
細胞内情報伝達経路で重要な役割を担っている Ca2+の詳細な振る舞いを知る ために,細胞内 Ca2+を測定する様々な手法が考えられてきた.細胞内のイオン 濃度測定は,イオン選択的電極を用いて溶液中のイオン濃度を測定する手法や 細胞懸濁液の吸光度を測定する手法などによって行われてきたが,それらは細 胞情報の 1 場面を切り出してきたものにすぎず,非常に限定的な情報しか得る ことができなかった.近年になって,細胞内の Ca2+動態を検討する場合に広く 用いられるようになった手法として,蛍光光度測定法を用いたバイオイメージ ング法があげられる.これは,対象となる細胞内に Ca2+と結合することによっ て特定の波長での蛍光強度が特異的に変化する蛍光 Ca2+指示薬を負荷すること で,細胞内 Ca2+濃度変化の測定が可能となるものである.細胞内イオン動態の 研究と共に急速に発展してきたバイオイメージング技法は,目的とするイオン に応じた様々な指示薬が開発され,生物学の分野において広く利用されている ことから,測定対象となる細胞組織やイオン種によっては,細胞内の情報伝達 を直接的に観察することのできる非常に有効な手段である.これらの研究に用 いられてきたのは単離細胞や培養細胞などがほとんどであった.骨格筋組織を 対象として直接的な顕微鏡観察を行う場合,筋組織自体の厚さに起因する空間 的分解能の低下が生じ,鮮明な画像を得ることができない.このため,バイオ イメージングの対象となったのは顕微鏡観察が容易な単離単一筋細胞や培養細 胞などであった.しかしながら,そのような生体を離れた細胞では本来の生体 内における環境とは当然異なることが考えられ,とりわけ代謝環境の違いが大 きいと考えられる.これらの制限は筋の機能的側面を検討するにあたって大き な問題点となっていた.
そこで本研究ではこれを解決する手法として,それまで主に血管形態や血流 動態に関連した研究において用いられてきた薄膜状筋である脊柱僧帽筋をイメ ージングの対象とする手法を考案し,生体内環境下にある骨格筋細胞内のイオ ン濃度変化を直接的に観察可能とすることを目的とした.この筋は,最薄部位 で筋線維数本程度の厚さしか持たず,環状な血管ネットワークが筋の剥離後も 保持されるという特徴を持つため,血流や神経活動が維持されている状態での 顕微鏡観察が可能である.脊柱僧帽筋を用いることで,生体から切り出された 細胞では不可能な生体筋組織からの情報が得られる可能性が考えられ,これま で生体外の実験において観察されてきた細胞内現象と,生体内で実際に起こっ ているであろう現象を結びつける役割を担うと考えられる.また,この手法を 用いることで,様々なストレス環境下における筋細胞内 Ca2+応答を検討するこ とを課題として設定した.
II. 文献研究
1. はじめに
生物が活動するにあたり,生体内では様々な種類の情報の伝達が行われてい る.生体が行っている情報伝達の経路として,化学物質が関与していることは よく知られており,イオン濃度勾配によって発生する電気的信号なども情報伝 達経路としてあげられる.身体運動に関わる骨格筋においても,信号は電気的 および化学的な情報伝達を介して伝達し,収縮が起こる.骨格筋収縮の過程に は筋を構成するタンパク質とCa2+との関係が非常に重要となり,Ca2+は筋収縮の トリガーといえる.しかしながら,疲労や収縮負荷の増大に伴い,筋細胞内に 収縮のトリガーとなる Ca2+が蓄積した状態になると,筋の弛緩が困難となり収 縮機能が低下する.これまで筋収縮と Ca2+の関係については様々なアプローチ による研究がなされており,筋細胞内 Ca2+の濃度変化が筋にとって様々な影響 を与えることがわかってきた.また,筋に一定レベル以上の大きな負荷が加わ った場合,筋は損傷という過程にいたる.筋の損傷は,高負荷による断裂など 外的に加えられる物理的ストレス以外にも,膜タンパク質の消滅による筋細胞 の崩壊など,内因的要素にも非常に大きく影響される.そのような内因的要素 のトリガーとして筋細胞内の Ca2+が筋の損傷にも関与していると考えられてい る.このように,骨格筋においては,筋細胞内 Ca2+濃度の変化が大きく関与し ている.筋機能を検討する際には細胞内の Ca2+動態を知ることが非常に重要で あり,本章では,筋と Ca2+の関係や,近年の細胞内 Ca2+測定技術,また様々な ストレス条件下での細胞内 Ca2+の動態について,最近の知見を踏まえて記述し た.
2. 骨格筋の収縮
Ca2+は,生体内の情報伝達機構において重要な役割を果たしている物質である.
Ca2+によって制御される生体機能が非常に多く存在することが古くから知られ ており,筋の制御もその典型的な例の一つである.1950年代後半から1960年代 初頭にかけて,江橋らによって骨格筋収縮は Ca2+によって制御されていること が明らかになった(Kumagai et al. 1955).また,Ca2+は筋収縮に関与するだけで はなく,細胞内に存在する様々な制御タンパク質と結合することにより,情報 伝達の調節因子としても機能していることが知られている.
1)筋収縮の情報伝達
身体を構成する骨格筋の場合,神経から電気的刺激を受けることで機能する.
筋線維に神経からの信号が伝わり,それが筋線維内部の筋原線維に到達し,筋 節が短縮することで収縮となる.これが,神経支配を受けているすべての筋線 維について発生する.このような,神経と筋の連動による筋収縮の一連のシス テムを興奮収縮連関(excitation-contraction coupling: E-C coupling)と呼ぶ.
筋を機能させるのは,主に中枢から発生する電気的信号である.電気的情報 は神経終末まで伝達し,神経筋接合部位の隙間で化学的情報伝達物質であるア セチルコリンが放出される.これによって電気的情報は化学的情報伝達を介し て筋線維表面の細胞膜へと伝達する.筋細胞膜ではアセチルコリンからの情報 を受けてNa+チャネルが開き,Na+の細胞内への流入が起こる.Na+流入による脱 分極で再び電気的信号に変換された情報は,膜状に存在する横行小管(transverse tubule)まで到達し,Ca2+の貯蔵庫である筋小胞体(sarcoplasmic reticulum)から Ca2+が細胞内に放出される.このことによって細胞内のCa2+濃度が上昇すると筋
が収縮を起こす.通常,一時的に上昇した細胞内の Ca2+濃度は,筋小胞体への 回収や Ca2+ポンプによる細胞外への排出によって,直に低い値に維持される.
Ca2+濃度が低下すると筋が弛緩状態に戻る.
2)筋収縮の分子機構
・アクチン・ミオシン
一本の筋線維は多数の筋原線維が集まったものである(Fig.1).筋原線維はア クチンからなる細いフィラメントとミオシンからなる太いフィラメントが規則 的に並ぶように構成されている.ミオシン頭部にはアクチンとの結合部位と ATP 分解酵素の働きがある.はじめミオシン頭部は,こぶしをつくって腕を曲 げたような形でアクチンと結合している.ATP 存在下ではミオシン頭部は立体 構造が変わるため,アクチンとの結合が外れ,こぶしを握ったまま腕を伸ばす ようにフィラメントに沿って移動する.ATP が分解されると,ミオシン頭部は 移動した位置でアクチンと結合しリン酸を放出してさらに強くアクチンと結合 する.リン酸を放出した結果,ミオシン頭部はADPを放して,もとの腕を曲げ た姿勢に戻り,このときアクチンフィラメントをたぐりよせるように張力が発 生する.こうしてミオシンは,はじめの姿勢に戻り,次のサイクルが始められ る.この運動を繰り返すことにより,筋節は短くなり,すなわち収縮がおこる.
・トロポニン
実際にどのように収縮が開始されるかというと,Ca2+が筋を構成するタンパク 質のひとつであるトロポニンと結合することが一連のトリガーとなっている
(Fig.1B).トロポニンは,筋の収縮が Ca2+をトリガーとしているという説が唱 えられだした 1960 年代に Ebashiらが発見したタンパク質であり(Ebashi et al.
1967),詳細な立体構造が解明したのは最近のことである(Takeda et al. 2003).
Fig.1A. 骨格筋の構造(Bloom and Fawcett. A Textbook of histology, 9th ed, Saunders, 1968.)
Fig.1B. トロポニンによる筋収縮制御(Ganong WF. Review of Medical Physiology, 14th ed. Appleton & Lange, 1989.)
トロポニンは Ca2+と結合し,その立体構造を変えることで,アクチンに存在す るミオシン結合部位をブロックしているトロポミオシンを引き離す.そのため,
ATP と結合し,アクチンとの結合が可能となっているミオシン頭部のATPが分 解され,上に述べたようなミオシンとアクチンの相互作用が始まるのである.
このタンパク質が,本来,常に収縮しようとしているアクチン・ミオシン間の 強い相互作用に抑制をかけ,Ca2+との結合時にはその抑制を解除するように構造 が変化することがわかっている.
3)骨格筋の収縮様式
筋収縮の様式を大別すると,静的収縮と動的収縮に分類される.静的収縮に は,筋の長さが変化せずに力を発揮する収縮様式である等尺性(アイソメトリ ック)収縮が属している.また,筋の長さが変化する収縮様式である動的収縮 には,いくつかの収縮様式が含まれる.等張性(アイソトニック)収縮は,筋 力が対抗する負荷よりも大きいと筋が短縮する.これを短縮性(コンセントリ ック)収縮という.一方,筋力が対抗する負荷よりも小さい時,自分が力を発 揮しようとする方向とは逆の方向に力が働く(筋の伸張).この収縮を伸張性(エ キセントリック)収縮と呼ぶ.また筋が一定の速度で収縮する場合を等速性(ア イソカイネティック)収縮と呼ぶが,実際の運動中にはほとんど見られない.
以上のような筋収縮様式は走,跳,投といったヒトの運動中において単独で見 られることは少なく,ほとんどの場合はエキセントリック収縮に続いてコンセ ントリック収縮が生じる.これらの収縮様式についてFig.2にまとめた.
3. バイオイメージング技法
!"#$
%"&$
'()*+,-./0
!"#$
%"&$
1*)*+,-./0 2345+,-./0
%"&$
!"#$678 96:;<=>?@AB
96:;<CDEFG 96:;<HIG
Fig.2. 骨格筋収縮様式の種類と収縮の模式図
近年の蛍光顕微鏡およびレーザー顕微鏡などの光学技術の急激な発達は,形 態学的な顕微鏡実験手法にも飛躍的な発達をもたらした.それまでの光学顕微 鏡に比べてより微細構造の可視化が可能となり,細胞構造の詳細などが明らか になってきた.しかしながら,観察対象となる生体組織や細胞を化学固定ある いは凍結して得られた画像情報には非常に限定的な情報しか含まれておらず,
生体内における生理的現象を明確に伝えたものではなかった.
1)細胞内イオンの可視化
細胞機能を考える際,細胞内のイオン濃度変化,特に Ca2+の変動が重要なフ ァクターとなっていることは前項でも述べてきたが,これまでの形態学的な手 法ではイオンそのものを可視化することができなかった.一方,近年になって 特定のイオンと特異的に結合する蛍光プローブの開発が急速に発展してきたこ とにより,細胞内のイオン動態を経時的に連続画像として取得することが可能 になってきた.今日,細胞内 Ca2+濃度の測定は,細胞のダイナミズムを可視化 する一つの手法として確立されてきている(Saino and Satoh. 2004).
・イオン特異的蛍光プローブの開発
初期の非侵襲的な細胞内イオン測定では,Ca2+と結合すると微弱な光を発する イクオリン(aequorin:オワンクラゲ由来のタンパク質)を,細胞内に注入する 方法が行われた(Allen and Blinks. 1978).この手法は細胞研究にとって非常に 有用であったが,方法論的な問題から,組織を対象とした研究には適用が困難 であった.その後,Ca2+キレート剤(tetracarboxylate)を基にQuin2,Fura2,Indo1, Fluo3などのCa2+プローブが開発された(Grynkiewicz et al. 1985, Tsien. 1980).
これらは,プローブ試薬のカルボキシル基にアセトキシメチル
(acetoxymethyl:AM)基をエステル結合させて脂溶性としたAM体とすることが
できる.AM体自身はイオンとの結合力がなく,細胞膜透過能を有しているため,
AM体を含んだ溶液に細胞を浸しておくことで,AM体は細胞外のイオンと結び 付くこと無しに細胞内に入ることができる.細胞内に入ると,AM基はエステラ ーゼにより切り離され,遊離イオンとの結合能を持ったプローブとなる.また,
水溶性となるため生体膜を透過できなくなり,細胞質内に留まるといった性質 をもっている.最近では二種の蛍光タンパク質,例えば青色蛍光タンパク質(blue fluorescent protein :BFP)と緑色蛍光タンパク質(green fluorescent protein :GFP) などを組み合わせて蛍光共鳴エネルギー移動(fluorescence resonance energy
transfer :FRET)を利用した分子生物学的なCa2+濃度測定方法が開発されている
(Rudolf et al. 2003).また,イクオリンなどのCa2+感受性発光タンパク質を特 定の細胞,もしくは細胞内小器官に遺伝子発現させて,部位特異的なCa2+測定が 試みられている.これらは全く新しい蛍光プローブであり,蛍光性の機能性タ ンパク質を細胞自身につくらせ,細胞の反応性をイメージングするという方法 は,Ca2+測定以外にも応用される可能性が高く,新たなプローブとして注目を集 めている.代表的なものに,Miyawakiらが開発したcameleonなどがある
(Miyawaki et al. 1997).以下には代表的な蛍光プローブの性質を示した.
・Quin2 化学名:
8-Amino-2-[(2-amino-5-methylphenoxy)methyl]-6-methoxyquinoline-N,N,N',N'- tetraacetic acid, tetrapotassium salt
Tsienら(Tsien et al. 1982)によって合成されたCa2+に特異的な初期の蛍光指示
薬.Ca2+と錯形成することによって強い蛍光を示し,Mg2+との反応は無視できる.
Ca2+錯体の蛍光励起波長は339 nm,蛍光波長は492 nmである.
・Fura2
化学名:
1-[6-Amino-2-(5-carboxy-2-oxazolyl)-5-benzofuranyloxy]-2-(2-amino-5-methylphenox y)
ethane-N,N,N',N'-tetraacetic acid, pentapotassium salt
Quin2の後に開発された蛍光プローブ(Grynkiewicz et al. 1985)であり,Quin2 よりもやや解離定数が大きく(Kd = 224 nM),1 µΜ付近までのCa2+濃度を計測 できる.また,吸収も大きく蛍光の量子収率も高いので,細胞内に入る蛍光色
素の量をQuin2 の場合の1/30に減らすことができる.もう一つの大きな特長と
してCa2+結合により励起波長のピークが顕著にブルーシフト(362 nm→335 nm) することで,335 nm付近で励起した場合にはCa2+濃度の上昇に伴い蛍光強度が 増大するのに対して,370〜380 nm付近で励起した時は逆に蛍光強度が減少する.
したがって適当な二波長(340,380 nm)を選択して励起し,蛍光強度の比をと ることによって,それが色素の濃度,光源の強度,細胞の大きさ,厚さ等に関 係なく Ca2+濃度と対応づけられる.この手法をレシオメトリーというが,これ に近い特徴をもつプローブにIndo1がある.
・Fluo3 化学名:
1-[2-Amino-5-(2,7-dichloro-6-hydroxy-3-oxo-9-xanthenyl)phenoxy]-2-(2-amino-5-met hylphenoxy)ethane-N,N,N',N'-tetraacetic acid
蛍光特性が励起波長480〜500 nm,蛍光波長530 nmの可視光のため,蛍光顕 微鏡やフローサイトメーターにとって使い易い.また励起光による細胞の損傷,
細胞由来の蛍光バックグランドなども軽減できる.Fluo3はCa2+結合による蛍光 強度の増加が大きく,約 40 倍にも達する.Ca2+親和力はFura2 よりも若干弱い
ため(Kd = 0.4 µM),Fura2よりも高いレベルのµMオーダーから低濃度に渡って,
Ca2+濃度を高感度にモニターできる.Fluo3の改良型にFluo4がある.
2)生体組織への応用
このような蛍光プローブを用いて行われたCa2+動態に関する多くの研究は,単 離単一細胞や,培養細胞を対象として用いており,組織としての細胞内イオン 動態を観察した例はきわめて少ない.その原因の一つとして顕微鏡の空間的解 像度の低さがあげられる.通常の顕微鏡システムでは,組織としての厚さによ って焦点面を定めにくく,空間的解像度が低下する.それゆえ,細胞内部のイ オン動態を解析するためには,どうしても細胞を単離する必要があった.とこ ろが,リアルタイムで連続した画像を高速に取得できる共焦点レーザー顕微鏡 が開発されたことから,生きた組織標本でCa2+動態を観察することも可能になっ てきた(Saino and Satoh. 2004).また,近年になって,通常の顕微鏡システム を用いた研究においても,共焦点顕微鏡とは別のアプローチによる骨格筋を対 象とした生体組織標本の観察が行われた.それらを以下に紹介する.
・骨格筋組織を対象とした蛍光プローブによる研究
筋細胞内のイオン動態を研究する場合,これまでに単離単一細胞や培養細胞 が主に扱われてきたのは,筋という厚みを持った組織を対象とした場合,顕微 鏡の空間的解像度の問題が原因の一つであった.これを解決する手段として,
対象とする組織を観察可能なものとすることが考えられた.Tothらは,ラット脊 柱僧帽筋(spinotrapezius muscle)を対象とし,Ca2+感受性蛍光プローブIndo1を 用いて,生きた生体組織での筋細胞内Ca2+動態を検討した(Toth et al. 1998).筋 組織の研究例としては他に,Teradaらが摘出した滑車上筋(epitrochlearis muscle) を対象に,Fura2を用いた筋組織内Ca2+変化を検討している(Terada, et al. 2003).
・生体内Ca2+動態の検討
脊柱僧帽筋は背部に位置する筋で,1973年にGrayが報告してから,主に血管 や血流に関連した研究の対象として用いられてきた(Gray. 1973).骨格筋とし ては非常に薄く,筋線維数本分しか厚みがない.主となる血管を傷つけないよ うに剥離することによって,神経活動や毛細血管血流を維持した状態での筋の 透過光顕微鏡観察が可能となる.速筋線維,遅筋線維の両方を含んでおり,骨 格筋として比較的良い研究対象となりうる.Tothら(1998)は,この脊柱僧帽筋 に対してCa2+感受性蛍光プローブIndo1を負荷し,虚血-再灌流および細胞内Ca2+
導入剤イオノマイシン(ionomycin)による薬理学的刺激を与えた時のCa2+増減 を検討した(Ivanics et al. 2000, Toth et al. 1998).同グループは,1991年にTyml が報告した筋標本を参考に,ラット長指伸筋(extensor digitorum longus :EDL)
とヒラメ筋(soleus :SOL)の筋表面部位を露出し,Indo1を用いてカフェインに よる薬理学的刺激時のCa2+増減を検討している(Batkai et al. 1999, Tyml et al.
1991).また,遺伝子導入技術を用いることによって,ラット前脛骨筋(tibialis
anterior :TA)にCa2+感受性を持つ蛍光タンパク質(cameleon)を発現させ,電気
刺激によるCa2+変化を検討したという報告がある(Rudolf et al. 2004).これは,
近年になって開発されてきた蛍光タンパク質を対象とした筋に直接発現させる ことによって,部位特異的にCa2+動態を追うことができる手法である(Dona et al.
2003).
これらのように,バイオイメージング法を用いて生体組織を対象とするCa2+
研究はここ数年のうちに急速な発展を見せている.生体の外で観察される組織 情報ではなく,生体内環境が維持された細胞内現象を知ることは,生体研究を 深く理解する上でも非常に重要な情報となりうる.
4. 筋細胞内 Ca2+の変化と筋機能変化
冒頭でも述べたように,筋を対象とした研究において,その収縮機構の制御 を担っている細胞内Ca2+動態を考慮することは非常に重要である.それゆえ,こ れまでにも非常に多くの研究者が,様々な条件下での筋細胞内Ca2+の様子を研究 対象としてきた.現在,筋収縮に関わるCa2+とタンパク質の情報伝達,収縮の分 子メカニズム,Ca2+増加による影響,細胞内へのCa2+流入経路など,多くの筋と Ca2+に関連した情報が明らかになってきている.しかしながら,未だ明白になっ ていない部分が多く残されていることも事実であり,近年では技術の発達とと もに,生体内環境下でのCa2+現象を検討することも可能となってきている.
1)細胞内Ca2+の増加
筋について研究するうえで,収縮を制御する Ca2+の存在が非常に重要である ことを述べてきた.細胞内では,情報刺激に対する応答の感受性を高めるため,
一般的な哺乳類細胞におけるCa2+濃度が,細胞外で1-2 mMであるのに対して,
細胞内では 10-4 mM と,非常に低い値に厳密に管理されている.筋小胞体によ る取り込みや細胞膜のイオンポンプなどが ATPのエネルギーを利用してこのよ うな環境を作り出している.しかしながら,収縮の繰り返しや持続的な最大張 力発揮などによって筋機能に過負荷が生じた場合は,筋小胞体の Ca2+取り込み 能力の低下や,細胞膜イオン透過性の変化による細胞外からの Ca2+流入を誘発 し,細胞内 Ca2+濃度の恒常性を保てなくなることがある.このような要因によ る細胞内 Ca2+恒常性の破綻は細胞の崩壊,ひいては筋損傷を引き起こすと考え られている.
・筋の収縮に伴うCa2+の上昇
筋の収縮には細胞内の Ca2+濃度上昇が不可欠である.通常,情報の伝達によ り刺激された筋小胞体から Ca2+が放出され,筋の収縮が起こった後は Ca2+濃度
は速やかにもとの低いレベルまで下げられる.しかしながら,筋収縮負荷など によって,筋細胞内の Ca2+濃度が平常時より高く維持されるという現象がわか ってきた.このことは,筋細胞の Ca2+に対する感受性を低下させるとともに,
細胞骨格の崩壊を導く要因となっているのではないかと考えられている.筋収 縮に関連する Ca2+動態については,非常に多くの研究がなされているが,その うち最新の知見をいくつか紹介する.
Hoganらのグループは,アフリカツメガエル(Xenopus)の単離単一筋細胞に
対して Ca2+感受性蛍光プローブを負荷し,電気刺激を与えたときの Ca2+動態を 観察している(Kohin et al. 2001, Kindig et al. 2005).彼らは,単一細胞が異なる 酸素環境下におかれたとき,収縮中のCa2+ピークと安静レベルを測定した(Stary
and Hogan. 2000).酸素環境による有意差は見られなかったが,筋発揮張力が最
大時の半分となった時点(疲労状態)における最大 Ca2+放出量の低下と,安静 状態のCa2+濃度上昇が生じたことを報告した.
Allenらは,伸張性(エキセントリック)収縮による機械的ストレスが,細胞
膜表面の機械受容チャネル(stretch-activated channels:SACs)を刺激することで 細胞内への Ca2+の流入が起こり,上昇した細胞内 Ca2+が筋を損傷へと導く要因 となっていると報告している(Yeung and Allen. 2004, see review Allen et al. 2005). 彼らは単離したマウスの筋線維を対象とし,電気刺激によるエキセントリック 収縮負荷を与えたときの細胞内の Ca2+を測定している.また,SAC のブロッカ ーであるガドリニウムイオン(Gd3+)や,ストレプトマイシンなどを負荷してお くことで,エキセントリック収縮誘発性の Ca2+流入が抑えられたと報告してい る(Yeung et al. 2005).また,Yeungらの報告によればSACの特異的ブロッカ
ーである GsMTx4 とパッチクランプ法を用いた電気生理学的な実験から,筋ジ
ストロフィーモデル動物であるmdxマウスではSACが開口しやすいことが示さ れた.これらSACを介したCa2+流入に関しては,mdxマウスを用いた実験から 筋ジストロフィー患者の筋状態と比較的近いものと考えられており,Ca2+上昇に
よる筋細胞崩壊の抑制などにその応用が考えられている.
また,下り坂運動をするときに,筋は伸張的に動員されることが知られてい る(Lindstedt et al. 2001).Lynchらは,マウスに長時間(24 h, 48 h)の下り坂運 動をさせた後の筋細胞内Ca2+を測定したところ,48時間運動後の長指伸筋では,
コントロールに比べて有意に Ca2+が増加していたと報告している(Lynch et al.
1997).長時間の収縮については,Carrollらが,蛍光Ca2+プローブを負荷した単
離筋細胞を対象とし,慢性的低周波刺激(chronic low-frequency stimulation:CLFS) 後の細胞内Ca2+を測定した(Carroll et al. 1999).10 Hzの電気刺激を最大で10 日間負荷し続け,安静時の Ca2+が 2 時間後から有意に上昇していることを報告 した.Gisselらも長時間の低周波刺激について報告をしている(Gissel. 2000).
45Caを用いた測定手法によって,1 Hzの収縮刺激を240分与えつづけた場合の 筋組織内Ca2+量を測定したところ,有意に差があることを報告した.
このように,筋の機能である収縮と,細胞の崩壊に直接的に関連する Ca2+に ついての研究は様々な条件,手法によって,多くの研究者が取り組んでいる.
その一方で,過度な収縮負荷によって Ca2+が上昇しているということが明らか になってきているが,実際の生きた生体環境下での現象を検討した例はまだ少 なく,議論の余地を残している.
・筋の虚血-再灌流に伴うCa2+の上昇
筋の収縮という生理学的な要因とは別に,細胞内の Ca2+を上昇させる要因と して虚血-再灌流負荷がある.虚血-再灌流障害は,様々な組織においてCa2+を有 意に上昇させることが古くから知られているストレスである.細胞内の Ca2+濃 度上昇が,Ca2+依存性プロテアーゼや各種酵素を活性化し,結果として障害を誘 発するのではないかと考えられている.心筋では,虚血-再灌流についての研究 が比較的多くなされており,虚血時には細胞内 Ca2+濃度が有意に増加するとい う報告をしている研究者も多い(Steenbergen et al. 1987, Kihara et al. 1989).一方,
骨格筋においては,虚血時には細胞内 Ca2+濃度はほとんど変わらないか,多少 上昇する程度で,再灌流時に急激に上昇を示すという報告がある(Forbes et al.
1996).また,骨格筋組織は相対的に虚血-再灌流障害には非感受性であり,筋組 織での不可逆的な細胞死が発生するまでには,約3-6時間の虚血状態を要するこ とが報告されている(Batkai et al. 1999).これを受けてIvanicsらは,虚血時間 の違いによる血流を維持する生体組織(骨格筋)でのCa2+動態を測定した(Ivanics
et al. 2000).その結果,短時間(~30 min)の虚血では,虚血状態に置かれた筋
組織のCa2+濃度変化に有意差は見られなかった.ところが60-120分の虚血状態 では,筋組織内の Ca2+は約 2 倍程度までに上昇し,続く再灌流後に安静時のレ ベルに近い値まで低下した.彼らは,虚血による低酸素状態が Ca2+増加に寄与 している要因の一つではないかと考察している.虚血-再灌流についての研究は,
特に灌流した心臓(心筋)を対象としてものが多く行われてきた.一方で,骨 格筋に関する虚血-再灌流とCa2+の関係は,未だ明確でない部分が多い.
5. まとめ
骨格筋とCa2+の関係について,主に,最近の生体組織でのCa2+研究にスポット をあてて紹介してきた.身体活動には筋の運動が不可欠であり,筋は様々な情 報伝達を介した末にCa2+の変化に応答し,収縮することができる.筋がCa2+によ って制御されていることがわかってから,約半世紀が過ぎたことになるが,Ca2+
の振る舞いについては未だ研究の余地が残されている.生体外におけるCa2+研究 から,単一細胞,組織,さらには生きた生体組織において,現在も様々な研究 が続けられているが,今後は,より生体内での現象を再現,もしくは生体内そ のものの測定系が重要視されてくると思われる.実際に生体内で起こっている 現象を直接的に観察することは,我々の身体自身に対するより発展的な知見を
もたらし,医療やスポーツ科学において,様々な恩恵をもたらすことが期待さ れる.
III. 研究目的および研究課題
骨格筋は,細胞内カルシウムイオン(Ca2+)濃度変化によって制御されている が,Ca2+濃度のバランスが乱れると筋機能低下,すなわち筋疲労や筋損傷を惹起 すると考えられている.その機序を考える場合,細胞内 Ca2+動態を知ることが 非常に重要である.そこで本研究では,生体内環境下での顕微鏡観察が可能な 膜状の骨格筋である脊柱僧帽筋に,蛍光 Ca2+指示薬を用いたバイオイメージン グを適用し,様々なストレスに対する筋細胞内の Ca2+応答を測定することを目 的として,以下の研究課題について検討した.
研究課題1.
生体内(in vivo)環境下における骨格筋細胞内 Ca2+-kinetics の可視化 生体内環境の整った条件(in vivo)において筋細胞内Ca2+を測定することを目 的として,ラット脊柱僧帽筋を観察対象としたバイオイメージング法を用いて,
in vitro,in vivo環境下にある筋細胞内Ca2+変化を比較,検討する.
研究課題2.
筋収縮様式の違いに着目した骨格筋細胞内 Ca2+-kineticsの差異
(1)様式の異なる筋収縮負荷後の筋細胞内Ca2+-kineticsの測定
in vivo における筋細胞内 Ca2+観察が可能であったことを受け,脊柱僧帽筋に
対して筋収縮刺激を負荷し,その後の Ca2+変化を測定することを目的とする.
収縮様式の異なるアイソメトリック,およびエキセントリック収縮を行い,細 胞内のCa2+変化を比較,検討する.
(2)伸張性筋収縮特異的に生じるCa2+蓄積に寄与するCa2+増加経路の検討
研究課題 2(1)を受け,エキセントリック収縮後の Ca2+増加経路を検討するこ
とを目的とする.細胞伸張に応答を示すストレッチ感受性イオンチャネル
(SAC)に着目し,その阻害薬であるストレプトマイシンとガドリニウムイオ ンを用いることでエキセントリック収縮時に生じるCa2+蓄積へのSACの関与を 検討する.
研究課題3.
骨格筋in vivoモデルを用いた細胞内Ca2+測定の発展的応用
(1)性差によって生じる筋機能の違いと細胞内Ca2+動態の関連について 研究課題 2 では筋細胞内の Ca2+レベルが筋収縮によって変化し,その後の筋 機能に影響を与えることが示唆された.研究課題3(1)は状態の異なる筋モデルの 例としてメス動物の筋を対象とし,収縮ストレスに対する細胞内 Ca2+応答を比 較,検討することを目的とする.
(2)糖尿病がもたらす筋機能の低下と細胞内Ca2+動態の関連について
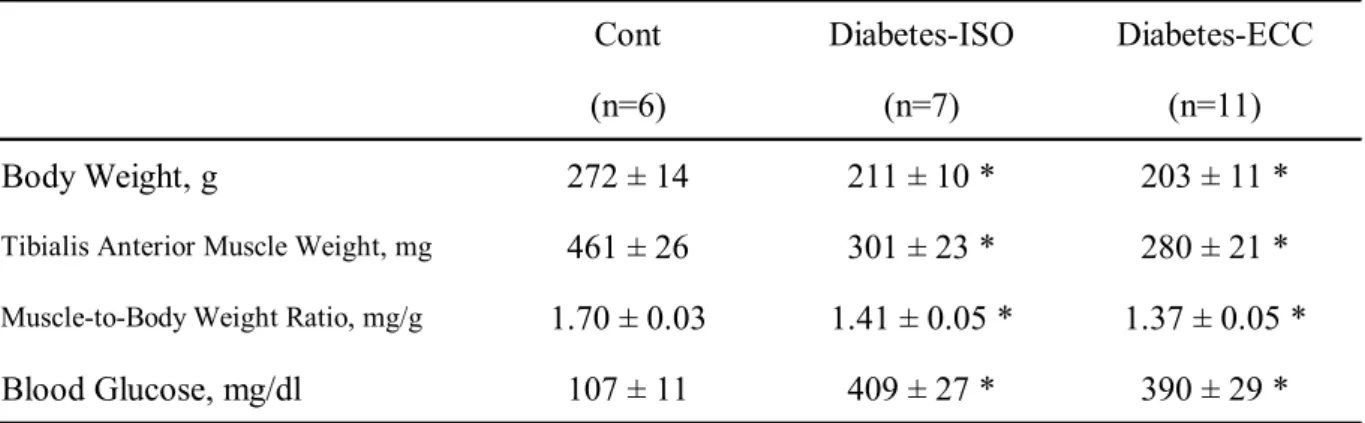
研究課題3(2)は状態の異なる筋モデルの例として,筋の脆弱性や循環機能の低 下を示す糖尿病モデル動物の骨格筋を対象とし,収縮ストレスに対する細胞内 Ca2+応答を比較,検討することを目的とする.
IV. 生体内( in vivo )環境下における骨格筋細胞内 Ca
2+-kinetics の可視化(研究課題 1 )
1. 背景と目的
筋線維の収縮-弛緩サイクルは,筋細胞内カルシウムイオン(Ca2+)濃度の変 化によって制御されている.弛緩状態の筋細胞内Ca2+濃度は0.1 µM以下に保た れているが(Armstrong et al. 1991),筋収縮時には一時的に濃度が増加し,筋収 縮のトリガーとしての役割を担っている.収縮後は Ca2+濃度が低下し,筋は弛 緩状態に戻る.筋の収縮機構のみならず,細胞内 Ca2+は多くのシグナル伝達経 路においてセカンドメッセンジャーとして機能しており,筋細胞内においても Ca2+濃度の変化をトリガーとして活性化する因子が多く存在している.このよう に,身体活動に貢献する筋の機能について考える場合,細胞内の Ca2+動態を知 ることが非常に重要である.これまでに多くの研究者が細胞内の Ca2+について の研究を報告してきたが,ほとんどは,主に生体外(in vitro)で行われた研究が 多く,“生きた”生体内(in vivo)Ca2+の振る舞いを追うことは技術的にも困難 であった.細胞内 Ca2+を測定するための初期の研究においては,イオン選択的 電極を用いた手法や細胞を磨り潰した懸濁液の成分を分析する手法などが行わ れてきたが,それらは非常に限定的な情報しか得ることができず,細胞情報の1 場面を切り出してきたものに限定されていた.
ところが近年になって発展してきた,特異的に蛍光を発するタンパク質や特 定のイオンと結合することで構造変化する蛍光物質を用いるバイオイメージン グ(蛍光イメージング)技法によって,細胞内の様々なイオンを直接的に可視 化する手法が確立されてきた(Tsien et al. 1982, Tsien et al. 1985, Cobbold and Rink.
1987).このことによって,in vitroで行われてきたCa2+の研究を,より生体内に 近い環境で測定することが可能になってきた.細胞内イオン動態の研究と共に 急速に発展してきたバイオイメージング技法は,目的とするイオンに応じた 様々な指示薬が開発され,生物学の分野において広く利用されていることから,
測定対象によっては非常に有効である.しかしながら,これらの研究手法に用 いられてきたのは主に単離細胞や培養細胞などといった生体から切り取られた 組織である.しかしながら,単一細胞や培養細胞では生体内における本来の環 境とは当然異なることが考えられ,筋細胞においては,発達した毛細血管血流 による酸素および代謝環境が特に大きく異なっていると考えられる(Stary and Hogan. 2000, Terada et al. 2003).筋の機能的な側面を検討するにあたり,より生 体内現象が統合された環境における実験系が必要であると考えられた.
これを解決する手法として,それまで主に血管形態や血流動態に関連した研 究において用いられることの多かった背部位の薄膜状筋である脊柱僧帽筋
(Fig.2)をイメージングの対象とする手法が考えられた(Toth et al. 1992, Toth et al. 1998, Ivanics et al. 2000).この筋は,最薄部位で筋線維数本分程度の厚さ
(300~500 µm)しか持たず,特徴的な形状による血管ネットワークが剥離後も
保持されるという特徴を持つ(Musch and Poole. 1996).したがって血流や神経 活動が維持されている状態においての顕微鏡観察が可能である(Gray. 1973).
そこで本研究では,筋細胞内 Ca2+の変化をより生体内に近い環境において検 討するため,in vivo での顕微鏡観察が可能な脊柱僧帽筋に蛍光カルシウム指示 薬を用いたバイオイメージング法を適用し,筋細胞内 Ca2+動態をリアルタイム に捉えることを目的とした実験系の確立を目指した.
2. 方法
・被験動物
本実験では,オスのWistar系ラット(日本SLC),体重200-300 g,8-12週齢 を用いた.全てのラットは室温20-24 ℃で12時間の明暗サイクルに管理された 飼育室において,餌と水をそれぞれ自由摂取できる状態で飼育した.全ての実 験は,電気通信大学動物実験指針に則って行われた.ラットは,ペントバルビ タールソディウム(大日本製薬,70 mg/kg i.p.)の腹腔内注入により麻酔した.
必要に応じて,麻酔は適時追加した.処置中のラットは,体温保持のために37 ℃ に維持されたホットプレート上に静置した.麻酔下において,右脊柱僧帽筋を,
組織に傷付けないよう慎重に露出し,表面を覆う結合組織なども可能な限り取 り除いた.脊柱僧帽筋に特徴的なアーチ状の細動静脈システムを維持するため に,筋の近位部において主となる供給動脈を残したまま筋を剥離した.筋は,
剥離する過程で筋と同等の大きさの馬蹄形針金リングに,安静時の筋長を維持 するよう縫合針と糸(Sigma)を用いて固定した(Fig.3筋標本写真参照).
・試薬調整
筋 は , 表 面 の 乾 燥 や 細 胞 の 崩 壊 を 避 け る た め に 常 に バ ッ フ ァ ー
(Krebs-Henseleit Buffer:KHB)によって保湿した.バッファーの組成は,132 NaCl, 4.7 KCl, 21.8 NaHCO3, 2 MgSO4, 2 CaCl2(mM)であった.バッファーは,95%
N2-5% CO2混合ガスによってpH 7.4に平衡化され,37 ℃に管理された恒温槽に
よって保温した.実験で用いた他の全ての薬品はKHBに溶かして用いた.蛍光 カルシウムプローブFura2-AM(Fig.4. Dojindo)は5 mMのストック溶液となる
ようにDimethyl Sulfoxide(DMSO: wako)に溶かし,細胞膜内へのプローブの浸
透を高めるため導入補助薬であるPluronic F-127(Molecular Probes)を最終濃度
Lash. J Appl Physiol. 1994
Fig.3. 脊柱僧帽筋の模式図と筋標本の様子(Lash. J Appl Physiol. 1994一部改変)
1-[6-Amino-2-(5-carboxy-2-oxazolyl)-5-benzofuranyloxy]-2-(2-amino-5- methylphenoxy)ethane-N,N,N', N'-tetraacetic acid, pentapotassium salt
C
29H
22K
5N
3O
14=831.99
A: Ca
2+結合飽和状態 B:Ca
2+-free
Fig.4. Fura2の分子式と波長特性
が0.1%以下になるよう加えた.蛍光プローブ溶液は,最終濃度が20 µMとなる ようにKHBに加えた.
・試薬負荷
露 出 し た 脊 柱 僧 帽 筋 を イ ン キ ュ ベ ー シ ョ ン チ ャ ン バ ー 内 に 静 置 し ,
Fura2-AM/KHB 溶 液で 30 分 イン キ ュベ ート した.細胞内に取り込まれた
Fura2-AM がその他の小器官に入り込んでしまう前にエステラーゼによってAM
体と切断されるよう,インキュベーションチャンバーは37 ℃のホットプレート 上に置いた.30 分のインキュベーション終了後,負荷されずに細胞外に残った
余分なFura2-AMを除くためKHBで数回,組織をリンスした.インキュベーシ
ョン後は少なくとも15分以内に顕微鏡観察を行った.
・顕微鏡観察
Fig.5 に観察の様子を模式図として示した.蛍光プローブを負荷した筋は,ラ
ットごと37 ℃に保たれたガラスホットプレート(Kitazato Supply)に固定し,
脊柱僧帽筋部分は,呼吸・拍動以外の要因による観察部位のズレを防ぐために ガラスプレート上で固定した.固定された筋組織を,ガラスプレート上で落射 型蛍光顕微鏡(Nikon),10 倍レンズを用いて観察した.はじめに透過光によっ て血流の有無,組織の状態を観察し,組織の端の部分は観察の対象外とした.
観察部位は,剥離後の脊柱僧帽筋血流を維持する供給血管(feeding artery)から 少なくとも1-2本目の分岐血管を視野内に含む部位から無作為に選択した.血管 をランドマークとして,筋線維を認識できる部位を対象とし,明視野の画像を 取得した.その後340 nm,380 nmの2種類の励起波長によって,蛍光を観察し た.蛍光観察の光源にはキセノンランプを用い,340 nmと380 nmのフィルター によって2種類の励起光を得た.測光は510 nmのフィルターを通して観察され
た.蛍光Ca2+指示薬Fura2は,340 nm付近で励起した場合にはCa2+濃度の
Fig.5. 実験装置の概要
上昇に伴い蛍光強度が増大するのに対して,380 nm付近で励起した時は逆に蛍 光強度が減少する.したがって 2 種類の波長によって励起し,蛍光強度の比を とることによって,色素の濃度,光源強度,細胞の大きさ,厚さ等に関係なく Ca2+濃度と対応づける手法をレシオメトリーという.透過光の観察により測定部 位を決定した後,手動で蛍光フィルタブロックを切り替えることにより,340 nm,
380 nm励起による蛍光画像を得た.ブロックの切り替えは約1秒以内で行い,
実験中は,これら2枚の画像を一組として経時的に取得した.
・画像記録とその解析
透過光,蛍光共に,画像の取得には高感度CCDデジタルカメラ(DP70,オリ ンパス)と,画像取得ソフトウェア(DP Control, オリンパス)を用いた.画 像は,解像度 1360 1024,ISO200(or 400)の状態で取得した.透過光像は自 動設定で画像取得し,蛍光画像の取得には,5秒の露光時間を設定し,実験中は 最初に設定した露光時間の変更はしなかった.データに示さないが,KHB のみ でインキュベートした蛍光指示薬を負荷しない場合の筋においては,5秒の露光 時間では観察可能な蛍光を取得できないことを事前に観察した.取得した画像 は,画像管理ソフト(DP Manager,オリンパス)によって一時的に保存し,画 像解析ソフトImageJ(National Institutes of Health)を用いて340 nm励起時の蛍 光画像(蛍光強度:F340)を380 nm励起時の蛍光画像(蛍光強度:F380)で除算 処理 し (image calculator ‒ divide mode),32-bit レシ オ画 像へ と 変換 した
(F340/F380:R).連続的に得られた経時的なレシオ画像を,式(gray = 0.299red + 0.587green + 0.114blue)を用いて各ピクセルをグレースケールに変換して数値化
(analyze - measure)し,初期値(R0)からの変化率をグラフとしてプロットし
た(R/R0).R値が高くなればCa2+濃度が上昇したと判断した.本実験で用いた 脊柱僧帽筋は,単一の細胞ではなく筋組織として顕微鏡観察されるため,一枚 の画像内に多くの筋線維を含んでいる.本実験では,数値化の選択範囲として
ROI(Region of Interest)を一枚の画像から得られる広範囲のエリア(約0.6 mm2) を選択し,これらの動態について検討した.
実験は,以下の2つの条件下において行った.
・条件1.in vitro
はじめに,in vitroの筋組織にて蛍光観察を行った.Fura2によるインキュベー ションが終了した筋を肩甲骨付近で切除し,顕微鏡下に置かれた37 ℃に保たれ たガラスホットプレート上に固定した.筋の乾燥を防ぐために,画像取得に影 響しないタイミングで常にKHBを与えた.画像の取得は最初の取得開始時から 5分おきに 90分間,計19セット行った.得られた19セットの画像を解析し,
グラフ化してCa2+動態として表した.
・条件2.in vivo
次に,血流を維持したin vivoの筋組織にて蛍光観察を行った.試薬を負荷し た後の筋をラット本体とともにガラスプレート上に静置,固定した.ガラスプ レートごと顕微鏡下に移動し,明視野にて組織の血流状態を観察し,撮影部位 を決定した後,条件 1 と同様に 90分間の画像取得を 19 セット行った.画像解 析結果をグラフ化し,Ca2+動態として表した.
各条件での測定後に薬理学的負荷として,筋小胞体リアノジン受容体に作用 してCa2+放出を引き起こすカフェイン溶液(5 mM Caffeine)と,Fura2に由来す る蛍光に対して消光作用を示すマンガンイオン溶液(5 mM MnCl2)の負荷を行 い,得られた蛍光がCa2+と結合したFura2に起因したものであることを確認した.
3. 結果
外科的に露出した脊柱僧帽筋の明視野像をFig.6に示した.対象動物が生きた 状態のまま顕微鏡観察が可能な点が,本実験で用いた脊柱僧帽筋の最大の利点 であるが,組織を構成する筋線維や毛細血管等も詳細に識別することができた.
測定中はラットの様態,血流を常に観察し,これらの血管が,in vivo の実験系 において測定終了時まで十分な血流を維持していることを確認した.明視野に て画像取得部位を決定後,手動の切り替えによる340 nm,380 nm各波長の励起 フィルターによって組織を蛍光励起し,蛍光観察した.各励起波長により,色 相の異なる2枚の蛍光画像を得た(Fig.7. 340 nm, 380 nm).これら2枚の蛍光画 像からレシオ画像を計算し,レシオ画像の様子から,筋細胞内での Ca2+の様子 を視覚的に観察することがきた(Fig.7 Ratio).
Fig.8は,ラットから切除したin vitro条件下での脊柱僧帽筋を,バッファー灌
流下で90分間静置した時の測定結果をグラフ化したものである.in vitro環境下 においてはCa2+濃度変化を表すレシオ値が漸進的に増加し,測定開始から55分 で初期値に対して有意な増加を示し(P < 0.05),その後も90分間の測定中は増 加傾向にあり,90 分後での値は初期値から 47.8 ± 11.2%増加した(Fig.8B).
Fig.8Aは,Fig.8測定時の340,380 nm各波長励起により得られた蛍光画像を,
画像全体の蛍光強度として数値化し,各初期値を 1 とした変化量で表したグラ フである.一方でin vivoにおける測定では,in vitroでの測定に対して90分間の 測定中,有意なレシオ値の増加が観察されなかった(Fig.9B).in vivoでの測定
中340,380 nm各波長励起による蛍光強度は低下する傾向にあった(Fig.9A).
さらにin vivoでの測定後,カフェイン溶液を負荷したところ,レシオ値の増加
が観察された(data not shown).
Fig.6. A typical bright-field image of rat spinotrapezius muscle. Most of all vessels and capillaries keep blood flow. Captured image area was including at least 1st or 2nd branching vessels from main feeding artery and that were used as landmark. This image was captured under in vivo condition. Scale bar is 100 µm.
Fig.7. Fluorescence images were captured from same area. Pairs of serial images were captured at 340, 380 nm excitation light. Exposure time was set to 5sec. Ratio (340/380) image was calculated (divided) by ImageJ (National Institutes of Health) software and measured mean gray-scale value.
These images were captured under in vitro control condition. A) 0 min, B) 30 min, C) 60 min and D) 90 min after dissection.
![Fig. 10. Two distinctly different profiles of [Ca 2+ ] i increase during the in vitro protocol](https://thumb-ap.123doks.com/thumbv2/123deta/7736080.1711820/51.892.171.728.104.867/fig-distinctly-different-profiles-ca-increase-vitro-protocol.webp)
![Fig. 12. Effect of 10 sets of isometric (ISO) and eccentric (ECC) contractions in spinotrapezius muscles in vivo on [Ca 2+ ] i](https://thumb-ap.123doks.com/thumbv2/123deta/7736080.1711820/62.892.170.758.144.594/fig-effect-sets-isometric-eccentric-contractions-spinotrapezius-muscles.webp)

![Fig. 14. Hypercontracted muscle fiber and [Ca 2+ ] i increase after the in vivo ECC](https://thumb-ap.123doks.com/thumbv2/123deta/7736080.1711820/64.892.246.640.77.859/fig-hypercontracted-muscle-fiber-ca-increase-vivo-ecc.webp)
![Fig. 16. Effect of stretch-activated ion channel blocker streptomycin and gadolinium (Gd 3+ ) following eccentric (ECC) contractions in spinotrapezius muscles in vivo on [Ca 2+ ] i](https://thumb-ap.123doks.com/thumbv2/123deta/7736080.1711820/75.892.172.776.138.598/effect-activated-streptomycin-gadolinium-following-eccentric-contractions-spinotrapezius.webp)

![Fig. 18. (A) Effect of sex difference following isometric (ISO) contractions in spinotrapezius muscles in vivo on [Ca 2+ ] i](https://thumb-ap.123doks.com/thumbv2/123deta/7736080.1711820/84.892.177.661.119.799/fig-effect-difference-following-isometric-contractions-spinotrapezius-muscles.webp)
![Fig. 19. (A) Effect of sex difference following eccentric (ECC) contractions in spinotrapezius muscles in vivo on [Ca 2+ ] i](https://thumb-ap.123doks.com/thumbv2/123deta/7736080.1711820/85.892.179.650.120.808/fig-effect-difference-following-eccentric-contractions-spinotrapezius-muscles.webp)