東京農業大学
博士論文
マウスにおける始原生殖細胞の
性特異的運命決定の分子機構
平成 28 年度修了
農学研究科バイオサイエンス専攻
坂下 陽彦
マ
ウ
ス
に
お
け
る
始
原
生
殖
細
胞
の
性
特
異
的
運
命
決
定
の
分
子
機
構
バ イ オ サ イ エ ン ス 専 攻坂
下
陽
彦
○
○
マウスにおける始原生殖細胞の
性特異的運命決定の分子機構
東京農業大学大学院農学研究科
バイオサイエンス専攻
平成 28 年度 博士課程修了
坂下 陽彦
指導教授 河野 友宏
1 論文題目
マウスにおける始原生殖細胞の性特異的運命決定の分子機構
博士論文 目次 第一章 緒論・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 4 付表および付図 第二章 マウス雌雄始原生殖細胞における性特異的発現遺伝子群の同定 第一節 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・16 第二節 材料および方法・・・・・・・・・・・・・・・・・・・・・・・18 付表および付図 第三節 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・25 1. FACS 法による E13.5 雌雄始原生殖細胞の回収率および純度 2. E13.5 雌雄始原生殖細胞の遺伝子発現プロファイルと性特異的発現遺伝子 群の抽出 3. 性特異的発現遺伝子群の生物学的意義の検証 付表および付図 第四節 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・44 第三章 シングルセルトランスクリプトーム解析によるマウス雌雄始原生殖細 胞集団における転写不均質性の証明 第一節 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・48 第二節 材料および方法・・・・・・・・・・・・・・・・・・・・・・・50 第三節 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・54 1. E13.5 雌雄始原生殖細胞における単一細胞レベルでの遺伝子発現プロファ イルと性特異的分化 2. E13.5 雌雄始原生殖細胞の遺伝子発現状態における変動遺伝子群と非変動 遺伝子群の同定 3. E13.5 雌雄生殖細胞におけるアポトーシス関連遺伝子群および細胞周期関 連遺伝子群の転写不均質性2 付表および付図 第四節 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・68 第四章 Sry 欠損性転換雌マウスの表現型解析 第一節 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・71 付表および付図 第二節 材料および方法・・・・・・・・・・・・・・・・・・・・・・・74 付表および付図 第三節 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・84 1. Sry ターゲティング CRISPR/Cas9 ベクターの構築 2. Sry 変異性転換雌マウスの変異アレル解析 3. Sry 変異性転換雌マウスにおける妊孕性の検証と卵巣組織の病理的解析 付表および付図 第四節 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・98 第五章 Sry 欠損性転換雌マウス雌性生殖細胞における網羅的転写産物および エピゲノム解析 第一節 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・100 第二節 材料および方・・・・・・・・・・・・・・・・・・・・・・・・102 付表および付図 第三節 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・107 1. Sry 変異雌性生殖細胞および生殖巣支持細胞の遺伝子発現プロファイルと 発現変動遺伝子群の抽出 2. 発現変動遺伝子群の生物学的意義の検証 3. Sry 変異雌性生殖細胞における生殖細胞崩壊機構の解明 付表および付図 第四節 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・131 第六章 総合考察・・・・・・・・・・・・・・・・・・・・・・・・・・136 第七章 総括・・・・・・・・・・・・・・・・・・・・・・・・・・・・144
3
Summary・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・152
謝辞・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・160 参考文献・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・161
4 第一章 緒論 遺伝的性に依存した性分化機構 自然界における生命の繁殖方法は、雌雄それぞれの生殖細胞の融合によって 成立する有性生殖 (Sexual reproduction) と単独個体(細胞)が出芽や分裂によ って次世代を作る無性生殖 (Asexual reproduction) に大別される。無性生殖に よって生じた次世代の個体群は遺伝的に同一のクローンであるため、劇的な環 境変化や病原体の感染拡大によって絶滅の危機に瀕する可能性を秘めている一 方、高頻度の遺伝的変異を獲得する能力を備えている。一方、多くの高等動物 が採用している有性生殖では、配偶子形成過程または接合時に遺伝子の組み換 えが生じるため、遺伝的多様性を獲得することが可能であったと考えられてい る。これにより有性生殖によって生じた次世代の個体集団は、親世代よりも多 様な環境に適応できる表現型を獲得できると考えられている。加えて、この有 性生殖における雌雄ゲノムの接合は、先カンブリア時代の地層からもその痕跡 が確認されている。約 5 億 6500 万年前に生息していた Fractofusus と呼ばれる 海洋生物は、以前まで生物が採用していたクローン増殖と珠芽による接合の 2 つの繁殖方法を行っていたことが推測されており (Mitchell et al., 2015)、その直 後にカンブリア大爆発などの生物の遺伝的多様性の獲得に深く関与する重要な 現象が生じている (Fox, 2016)。このことから、有性生殖は生物進化にも極めて 重要な働きをみせたことが窺える。 この遺伝的多様性を生み出す有性生殖において雌雄それぞれの機能的な差異
5 を形成する「性決定機構」は、生命の継続を支える極めて重要な仕組みといえ る。性決定には動物種ごとに多彩な方法が採用されていることが知られている が、「遺伝的性決定」と「環境性決定」に大別され、以下の 4 種の機構に分類さ れる(:性染色体の構成、染色体の倍数性、温度、生育環境)(Bachtrog et al., 2014)。 遺伝的に性が決定される多くの脊椎動物においては、性染色体が重要な役割を もつことが報告されており、哺乳類および魚類では、XX (雌)、XY (雄)が性染色 体として存在し、胎仔期の生殖巣支持細胞において Y 染色体上から発現する SRY (Sex-determining region Y) ま た は DMY (DM-domain gene on the Y chromosome) が雄性性決定因子として同定されている (Bachtrog et al., 2014; Gubbay et al., 1990; Matsuda et al., 2002; Sinclair et al., 1990)。一方、Z ならび に W 染色体を性染色体として保有する鳥類や一部の爬虫類では、ZW をもつ個 体が雌へ、ZZ をもつ個体が雄へ分化することが知られている(Bachtrog et al., 2014)。これらの Z および W 染色体に依存した性決定を採用している生物種で は、SRY や DMY のような明確な性決定のマスター遺伝子は同定されていない が 、 W 染 色 体 上 の FET1 (Female expressed transcript 1) 、 ASW (Avian sex-specific, W-linked) 遺伝子および Z 染色体上の DMRT1 (Double sex and mab-3 related transcription factor 1) の存在量が性決定に重要であることが現在 までの解析によって示唆されている (Chue and Smith, 2011; O'Neill et al., 2000; Reed and Sinclair, 2002; Smith et al., 2007)。上述の性染色体上のマスタ ー遺伝子の働きにより、下流の性決定関連因子の逐次的な発現が制御され、遺 伝的性に基づいた性分化が起こることが明らかにされている。また、遺伝的性
6
決定機構の内、染色体の倍数性を採用しているハチやアリなどの膜翅目では、 半数体の個体が雄型に、2 倍体の個体が雌型へ分化することが報告されている (Bachtrog et al., 2014; Cho et al., 2007)。気温や水温、pH などの環境要因によ って性が決定される多くの爬虫類や一部の魚類では、温度依存的に性ホルモン の転換を行うアロマターゼなどが性決定因子として働いていることが知られて お り 、 外 部 環 境 に 合 わ せ て 容 易 に 性 転 換 を 起 こ す こ と が 報 告 さ れ て い る (Bachtrog et al., 2014; Desvages and Pieau, 1992; Navarro-Martin et al., 2011)。
本研究で対象とする哺乳動物のモデル生物であるマウス (Mus musculus) の 性決定は、XX または XY の遺伝的性に支配され、胎齢 (E) 11.5 の生殖巣におい て性分化の初動が観察される (Larney et al., 2014) (Figure 1-1)。性決定以前の 生殖巣における SRY の発現は、プロモーター上に転写抑制マーカーである H3K9me2 (Histone H3 dimethylated Lysine 9) が存在することにより抑制され ている。この H3K9me2 による SRY の転写抑制は、E11.0 (10―12 尾体節期) に おいて 6 番染色体から発現されるヒストン脱メチル化酵素である JMJD1A (Jumonji domain-containing 1a) によって解除され、14―22 尾体節期ステージで 性決定および雄化のマスター遺伝子である SRY の発現が GATA4 (GATA binding protein 4) 陽性を示す生殖巣支持細胞内で惹起される (Kuroki et al., 2013; Tanaka and Nishinakamura, 2014; Tevosian et al., 2002; Warr et al., 2012)。これ により雄性生殖巣内の支持細胞は、セルトリ細胞 (Sertoli cells) へと分化するこ とが知られている。SRY の発現を起点としてセルトリ細胞内で下流の雄性性決 定因子である Sox9 (Sex-determining region Y-box 9) や Fgf9 (Fibroblast growth
7
factor 9) の発現が誘起され、Fgf9 の正のフィードバック作用により Sox9 は精 巣内で常に高い発現を示すようになる (Jakob and Lovell-Badge, 2011; Kim et al., 2007)。この Sox9 の高い発現が Amh (Anti-Mullerian hormone) 遺伝子の発 現を促し、雌性生殖管であるミューラー管を退行させることで、雌型への分化 が抑制される (Munsterberg and Lovell-Badge, 1991)。さらに、セルトリ細胞に おける Sox9 の発現は周辺の体細胞にも影響を与え、E12.5 の生殖巣ではライデ ィッヒ細胞 (Leydig cells) が形成される (Chen and Liu, 2016; Clark et al., 2000; Yao et al., 2002)。このライディッヒ細胞内の転写因子 Sf1 (Nr5a1) の働 きによりテストステロンの合成が開始され (Barsoum and Yao, 2010)、パラクリ ンを通じてウォルフ管の精巣上体、精輸管および精嚢への分化が生じる (Gupta and Bentlejewski, 1992; Higgins and Burchell, 1978)。また、テストステロンは、 脳、筋肉および骨格にも作用し、生体全体の雄化を促すことが知られている (Chen et al., 2004; Griggs et al., 1989; Zuloaga et al., 2008)。これらの体細胞の 分化に伴い、精巣に到達した始原生殖細胞 (Primordial germ cells: PGCs) も E14.5 まで有糸分裂を続け、セルトリ細胞が分泌する TGFβ (Transforming growth factor beta) の 影 響 を 受 け て 細 胞 周 期 を 停 止 さ せ た 前 精 原 細 胞 (Gonocyte/Pro-spermatogonial stem cells ) が形成される (Moreno et al., 2010)。 この精原細胞が精子形成における幹細胞となり、出生後もセルトリ細胞から分 泌される GDNF (Glial-cell derived neurotrophic factor) および SCF (Stem cell factor) に応答して自己複製を続ける (Chen and Liu, 2015; Mei et al., 2015; Zhang et al., 2014)。その後、6 週齢までに減数分裂を経て成熟精子へと分化す
8
る。一方、雌型 (XX) の生殖巣では Y 染色体が存在しないため SRY による卵巣 決定因子 (Wnt4、Rspo1) の発現抑制が生じず、これらが雌の生殖巣で高い発現 を示す (Chassot et al., 2012; Chassot et al., 2014; Vainio et al., 1999)。これに よりウォルフ管は退行し、ミューラー管が卵管および子宮へ分化することが報 告されている。また、卵巣内でも、Wnt4 (Wingless-type MMTV integration site family, member 4)および Rspo1 (R-spondin 1) のシグナルを受けて、Foxl2 (Forkhead box L2) 陽性を示す髄質顆粒層細胞 (Medullary granulosa cells)、
Lgr5 陽性を示す皮質顆粒層細胞 (Cortical granulosa cells) および Nr2f2 陽性を
示す間質細胞 (Interstitial cells) が出現し、生殖巣内の体細胞系譜が確立される (Biason-Lauber, 2012; Rastetter et al., 2014) (Figure 1-1) 。この卵巣内の体細胞 環境に支持され、雌性 PGCs は、中腎から供給されるレチノイン酸に応答して 減数分裂期に移行する(Niederreither and Dolle, 2008)。
生殖細胞と生殖巣体細胞環境の相互作用による雌雄生殖細胞形成 これらの雌雄生殖細胞の発達を支持する生殖巣の体細胞環境は、生殖細胞の 性特異的な分化と維持に極めて重要であり、現在までに様々な検証が行われて きた。第一に、Foxl2 の顆粒層細胞特異的ノックアウトによって性成熟後の成体 の卵巣においても SOX9 陽性を示す精細管様の構造体が生じることが明らかに されている(Uhlenhaut et al., 2009)。また、雄マウス精巣内においても Dmrt1 の セルトリ細胞特異的ノックアウトによって FOXL2 陽性を示す顆粒層細胞および 莢膜細胞が出現することが報告されている(Matson et al., 2011)。これらのマウ
9
スは、Foxl2 もしくは Drmt1 の薬剤誘導性欠損と同時に生殖細胞が退行し、不妊 の表現型を示す (Matson et al., 2011; Uhlenhaut et al., 2009)。これらの研究に よって、一次性決定だけでなく性成熟後の性特異的な生殖細胞形成過程におい ても、周辺の体細胞環境とその性特異的な特性を維持する転写因子の働きが重 要であることが明らかになった。 第二に、Mclaren らの生殖細胞移植実験により、移動期の XX 雌性 PGCs を雄 型 (XY) の生殖巣環境におくことで、精子形成が促さられることが明らかにな った。また、XY 雄性 PGCs を XX の体細胞環境に置いたときも同様に、卵子形 成が誘導されることが示された (Adams and McLaren, 2002)。これにより、 E12.5 までに移植された PGCs は、自身の性染色体の構成ではなく、移植先の 体細胞の性に依存して卵子または精子に分化することが明らかにされた。加え て、XX 型および XY 型双方の体細胞環境をもつ XX↔XY 生殖巣キメラマウスで は、XY 体細胞が分泌する PTGDS (Prostaglandin-H2 D-isomerase) がパラクラ イン因子として働き、周辺の XX 体細胞も精巣構成細胞である精細管周囲筋様細 胞 (Peritubular myoid cells: PMD)、ライディッヒ細胞あるいはセルトリ細胞の いずれかへ分化させることが示されている (Adams and McLaren, 2002; Moniot et al., 2009)。これによりほとんどのキメラ個体が雄の生殖巣である精巣を形成 することが報告されている(Adams and McLaren, 2002; Palmer and Burgoyne, 1991)。興味深いことに、XX↔XY 精巣中の XX 生殖細胞は精子形成へと誘導され、 ゲノム中には父方インプリントが刷り込まれることが明らかにされている (Isotani et al., 2005)。
10
最後に、突然変異もしくは人為的遺伝子欠損によって生じる性転換マウス では、自身のもつ性染色体の構成とは異なる生殖巣内の体細胞環境をもつこと が知られている (Table 1-1) 。例えば、一般的な XY 性転換雌マウスでは、Y 染 色体を保有しているのにも関わらず、Sry 遺伝子の機能欠損により雌型の生殖巣 環境へ分化することが報告されている (Gubbay et al., 1990; Gubbay et al., 1992; Kato et al., 2013; Lovell-Badge and Robertson, 1990)。これにより、生殖 細胞は体細胞の性に依存して卵子様に分化することが古くから知られていた。 これらの知見から、生殖細胞の性特異的な運命決定は、自身の性染色体の構成 に加え、体細胞 (生殖巣環境) の性決定に大きく依存することが明らかにされて いる。しかしながら、現在までの研究によって正常な性分化が起こったように 見える性転換マウスにおいて様々な生殖および繁殖能力の異常が確認されてい る (Burgoyne and Baker, 1981; Cattanach et al., 1971; Mahadevaiah et al., 1993; Probst et al., 2008; Vernet et al., 2011; Vernet et al., 2014)。これらの性転 換マウスの異常な表現型は、ヒトのターナー症候群 (XO woman)、スワイヤー 症候群 (XY woman) ならびにクラインフェルター症候群 (XXY or XYY man) などの性分化疾患の臨床所見とも類似している (Ma et al., 2000; Massanyi et al., 2013; Smyth and Bremner, 1998; Sybert and McCauley, 2004)。
Y 染色体の欠失によって単一の X 染色体のみを保有する XO 雌マウスでは、 野生型雌マウスと比較して大幅な卵胞数の低下による繁殖期間の短縮が報告さ れている (Burgoyne and Baker, 1981; Probst et al., 2008)。また、ヒトにおいて
11
はさらに重篤な症状が観察されており、卵巣組織が索状痕跡様の構造を示し、 生殖細胞および卵胞は、思春期を迎える前に全てが退行してしまう (Sybert and
McCauley, 2004)。加えて、Sry 遺伝子が欠失した XYTdym1もしくは XYsry-雌マウ
スの生殖細胞も大部分が第一減数分裂過程で性染色体の不対合を起こし、消失 してしまうことが報告されている(Burgoyne et al., 2009; Mahadevaiah et al., 1993)。また、出生後に成長する卵母細胞も透明帯の形成異常および卵子サイズ の低下などの表現型異常が示されている(Vanderhyden et al., 1997)。ターナー症 候群と同様に、ヒトにおけるこの核型も卵巣組織の索状化と生殖細胞の退行が 観察されている(Massanyi et al., 2013)。加えて、Sry の下流に位置する他の雄性 性 決 定 因 子 の 欠 損 マ ウ ス も 類 似 し た 表 現 型 を も つ こ と が 報 告 さ れ て い る (Bagheri-Fam et al., 2008; Colvin et al., 2001; Lavery et al., 2011; Siggers et al., 2014)。興味深い例として、Molosinus 型の Y 染色体をもつ C57BL/6 雄マウスの Y 染色体を Domesticus 型に置換することで、一定の確率でコンソミック系統の XY 雌 マ ウ ス が 誕 生 す る こ と が 報 告 さ れ て い る (Antonio-Rubio and Moreno-Mendoza, 2012; Correa et al., 2012; Nagamine et al., 1999; Whitney et al., 2000)。これらの卵子も減数分裂過程で X 染色体不活性化および紡錘糸の形 成不全によって消失し、不妊の表現型を呈する (Amleh et al., 1996; Amleh and Taketo, 1998; Correa et al., 2012; Merchant-Larios et al., 1994; Taketo and Naumova, 2013; Villemure et al., 2007)。また、ほとんどの XX 雄性転換マウス も同様に、精子形成過程のいずれかのステージで生殖細胞が退行してしまい、 妊孕性の低下もしくは消失が確認されている (Ma et al., 2000; Vernet et al.,
12 2011)。このように、見かけ上は正常に雌型もしくは雄型に性分化した性転換マ ウ ス で さ え 、 生 殖 細 胞 形 成 過 程 に お い て 様 々 な 異 常 が 検 出 さ れ て い る (Table1-1)。 残された解決すべき課題 これまでに生殖細胞の性特異的な分化を支持する SRY を含む多様な体細胞側 の因子は同定されており (Figure 1-1)、これらの体細胞の性に依存して生殖細胞 は、有糸分裂の停止、減数分裂への移行および性特異的なゲノムインプリント の確立など性特異的な細胞運命を獲得することが明らかにされている(Barlow and Bartolomei, 2014; Moreno et al., 2010; Saba et al., 2014)。しかしながら、 機能的な雌雄生殖細胞形成を理解する上で、残された課題も少なくない。例え ば、(1) 発生のどの時期から生殖細胞に明確な性特異的な分化が生じるか? また、 どのような制御因子およびネットワークが関与し生殖細胞の性特異的な運命を 規定するか理解されていない。当然、そのプロセスも全く不明といえる。(2) Y 染色体には性決定遺伝子 Sry の他、多数のタンパク質コード遺伝子が存在する が、それらは性特異的生殖細胞形成においてどのような機能を保有しているか ほとんど知られていない。(3) 性染色体の構成と生殖巣環境の性の不一致によっ て、完全な雌型の生殖巣環境をもつ性転換雌マウスの卵子形成が、何故崩壊し てしまうのか、興味深いが分子生物学的な理解は全く進んでいない。さらに、(4) 生殖細胞の性特異的運命決定を支配する機構には、DNA メチル化やヒストン修 飾を含むエピゲノム修飾によるは制御機構の関与が当然重要な働きをしている
13 と考えられるが、その実態についてもほとんど解明されていない。 本研究の目的 本研究は、マウス生殖細胞の性特異的運命決定のプロセスを制御する分子機 構の解明を進め、生殖細胞形成やそれらの機能獲得に対する理解を深めること を目的に、前述の残された解明すべき研究課題のうち、特に (1) から (3) に注 目し、以下の一連の実験を実施した。(1) 包括性および定量性に優れた RNA-seq 法を用いて、生殖巣到達直後の E13.5 雌雄 PGCs の網羅的トランスクリプトー ム解析を行った。これにより、この時期の雌雄 PGCs がどのような遺伝子発現 プロファイルを保有しているかを明らかにし、体細胞の性に依存した明確な性 分化がすでに雌雄生殖細胞において生じているかどうか検証を行った。(2) その 後、雌雄 PGCs の遺伝子発現における性差に焦点を当て性特異的発現遺伝子群 を同定し、これらの in silico 解析を介して機能的生殖細胞形成を支持する制御因 子の同定を試みた。(3) さらに、性染色体の構成と雌雄特異的な生殖巣環境が機 能的生殖細胞形成にどのような影響を与えているかをより詳細に解析するため、
ゲノム編集技術である CRISPR/Cas9 システムを用いて Sry 欠損 XYsry-性転換雌
マウスを作出し、XYsry-卵子形成が何故正常に行われないのか追究した。本研究
により得られた知見は、機能的雌雄生殖細胞形成において必須な生殖細胞の性 特 異 的 運 命 獲 得 機 構 の 理 解 に 貢 献 す る 。 ま た 、 ヒ ト に お け る 性 分 化 疾 患 (disorders of sex development, DSDs)の発症機構の理解にも役立つことが期 待される。
14 Figure 1-1. 哺乳動物 (マウス) における生殖腺の性分化機構 生殖巣環境の性は、E11.5 ごろに生殖巣支持細胞内における Sry の発現の有無によって決定される。 これにより雌の生殖巣支持細胞 (顆粒層細胞) では Ovarian genes の発現が、雄の生殖巣支持細胞 (セルトリ細胞) では Testicular genes の発現が活性化し、性特異的な生殖巣形成 (卵巣形成および 精巣形成) が開始される。
15
Strain Sex Fertility Allele Reference
Inbred F Fertile Wild type
-Outbred F Subfertile,
Shorter reproductive life Single X chromosome Decker JFM. 1981
Congenic F
Infertile,
Embryo derived from this female arrest cell cycle at 2-cell stage
X chromosome carries the
X-linked Zfy2 Vernet N. 2014
Outbred F Subfertile,
Occasional offspring
Y chromosome has a 11 kb
deletion, including Sry locus Lovell-Badge R. 1990
Outbred F Subfertile,
Occasional offspring
Y chromosome has a 11 kb deletion, including Sry locus and large deletion in MSYq region
Toure A. 2004
Outbred F Infertile Disrupted Sry locus by TALEN
mediated knockout Kato T. 2013
Consomic F (60%), H (30%),
M (10%) Not reported
Disrupted Jmjd1a locus by
targeting knockout Kuroki S. 2013
Congenic F Subfertile,
Occasional offspring
Disrupted Sox9 locus by
targeting knockout Lavery R. 2011
Congenic F
Not reported
Null mouse die at birth by lung hypoplasia
Disrupted Fgf9 locus by
targeting knockout Colvin JS. 2001
Outbred F Subfertile,
Similar to XO female
Y chromosome has depleted
Rbmy gene cluster. Vernet N. 2014
B6.YAKR Consomic F (60%), H (10%),
M (30%)
F and H individuals are infertile. But, M individual is fertile
Y chromosome from Mus
musculus domestius (AKR) Suto J. 2009
B6.YC3H Consomic F (15%), M (85%) F individual is infertile. But, M
individual is fertile
Y chromosome from Mus
musculus domestius (C3H) Suto J. 2009
B6.YTIR Consomic F (25%), H(25%),
M(50%)
F and H individuals are infertile. But, M individual is fertile
Y chromosome from Mus
musculus domestius (Tirano) Taketo T. 1989
Consomic F (75%), H(25%)
Subfertile,
Occasional offspring (Extremely rare)
Y chromosome from Mus
poschiavinus Eicher EM. 1982
Inbred M Fertile Wild type
-Outbred M
Infertile,
How ever, offspring could be obtained using ROSI.
X chromosome carries the X-linked Eif2s3y. Sry transgene locates autosome.
Vernet N. 2011
Outbred M
Infertile,
How ever, offspring could be obtained using ROSI.
X chromosome carries the X-linked Eif2s3y and Sxr (Sex reversal region)a
Vernet N. 2011
Outbred M
Infertile,
How ever, offspring could be obtained using ROSI.
X chromosome carries the X-linked Eif2s3y and Sxr (Sex reversal region)b w hich is deletion variant of Sxra
Vernet N. 2011
Outbred M
Infertile,
This mouse have no spermatogonial stem cell
Sry transgene Koopman P. 1991
Congenic M
Infertile,
This mouse have hypoplastic testis.
Dmrt1 transgene Zhao L. 2015
Outbred M Fertile
Y chromosome has a 11 kb deletion, including Sry locus, male development is dependent on the autosomally located Sry transgene.
Vernet N. 2011
Outbred M
Infertile,
This mouse have no spermatogonial stem cell
Sex chromosome trisomy Lue Y. 2011
XEif 2s3ySxraO XEif 2s3ySxrbO XYsry - Sry XXY B6.YDOM B6.YPOS XX Dmrt1 XX Sry XYTdy m1qdel XY Sox9 -/-XY Fgf9 -/-XY XEif 2s3yO Sry XYsry -XYC3H Jmjd1a -/-XYd1-d5 Genotype XX XO XZf y 2O XYTdy m1
性別 (sex) の列に含まれる F, H および M は、それぞれ Female (雌)、Hermaphrodite (半陰陽性個体) および Male (雄) を示す。
16 第二章 マウス雌雄始原生殖細胞における性特異的発現遺伝子群の同定 第一節 緒言 有性生殖を行う高等生物において、生殖細胞は受精を介して「全能性」を獲 得する唯一の細胞であり、生殖細胞のみが自身の遺伝情報を次世代に受け継ぐ ことができる。生殖細胞の形成過程では、DNA メチル化やヒストン修飾状態が ゲノム広範囲に再編成され、Blimp1、Fragilis、Stella、Prdm14 などの遺伝子が エピブラストの一部の細胞で発現し、始原生殖細胞 (PGCs) への分化誘導を制 御する (Kurimoto et al., 2008)。生殖細胞の機能を特徴づける分子機構は、「エピ ゲノム修飾」と呼ばれる DNA メチル化修飾やヒストン修飾などの塩基配列に依 存しない遺伝子発現制御機構により成立している (Saitou et al., 2012)。 その後、雌雄 PGCs は定着した生殖巣の性に従って、雌性 PGCs は減数分裂 期に移行し、雄性 PGCs は細胞周期を停止させることが知られており、この PGCs の性特異的な細胞プロセスには、様々な性特異的な生殖巣環境中の遺伝子 制御ネットワークが重要であることが現在までの研究によって明らかにされて いる。例えば、雌雄双方の中腎管において Raldh3 によるレチノイン酸合成は生 じるものの、雄の生殖巣ではセルトリ細胞が CYP26B1 を産生しているため、レ チノイン酸は生殖巣中で分解される。これにより、雄性 PGCs は Stra8 の活性 化を受けず、体細胞分裂期を維持することが報告されている。一方雌性 PGCs 中ではレチノイン酸に応答し Stra8 および Rec8 の発現が生じ、下流の減数分裂 関連遺伝子の発現を活性化させることが示されている。
17 しかしながら、発生のどの時期から生殖細胞自身に明確な性特異的な分化が 生じるか、 また、どのような制御因子およびネットワークが生殖細胞の性特異 的な運命を規定するか未だ明らかにされていない。従って本研究では、雌雄 PGCs における性分化の分子機構を明らかにすることを目的として、性特異的分 化の起点となる E13.5 の雌雄 PGCs を次世代シークエンサーに供試し、RNA-seq 解析によって性分化の起点となる E13.5 雌雄 PGCs の網羅的転写産物解析を行 った。
18
第二節 材料および方法
1. 供試動物
本研究では、性成熟後の 8―12 週齢の C57BL/6NJcl (B6N) 雌マウス (日本ク レア) および Pou5f1 の遠位エンハンサーの制御下で Green fluorescence protein (GFP) を発現する性成熟後の 10―12 週齢の Pou5f1-⊿PE-GFP 雄マウス (B6 background) を供試した (Yoshimizu et al., 1999)。使用したマウスは全て午前 8 時点灯、午後 8 時消灯、水、餌は不断供給の環境下で飼育した。これらの実験 動物を用いて行った実験は全て東京農業大学の「実験動物の管理と使用に関す る指針」に基づいて行った。 2. マウス ES 細胞の培養および回収 本研究では、E13.5 BALB/cA 由来胎仔繊維芽細胞と共培養を行ったマウス ES 細胞 (XY B6J background) を用いた。細胞培養は、1×105 ES cell /ml の濃度 で開始し、LIF (1000 U/ml; Chemicon)および 2i (0.4 μM PD0325901; MEK 阻害 剤 (Stemgent) および 3 μM CHIR99021; GSK3 阻害剤 (Stemgent))添加 ESC 培 地 (15% KSR, 0.055 mM β-merccaptoethanol, 2 mM L-glutamine, 0.1 mM MEM non-essential aminoacids, 5000 U/ml penicillin/streptomycin in KnockOut DMEM) 内で行った。コンフルエントになるまで 2―3 日間培養を行った後、
0.25% trypsin-EDTA (Sigma) 処理により単一細胞に分離し、1×104細胞を 1 プ
19
3. E13.5 雌雄 PGCs の回収
Pou5f1-⊿PE-GFP 雄マウスと交配後 13.5 日目の B6N 雌マウスを頸椎脱臼に より安楽死させ、E13.5 Pou5f1-⊿PE-GFP ヘテロマウス胎仔を 0.4% BSA 添加 PBS 中に回収した。顕微鏡下でこれらの胎仔から生殖隆起を採取し、精索の形 成が確認できたものを雄、確認できなかったものを雌として雌雄判別を行った。 雌雄それぞれの生殖隆起を 1.5 ml チューブに移し、0.1% Collagenase (Wako) 添加 L15 培地を 1 ml 加え 37℃で 30 分間処理した。ピペッティングにより細胞 が解離したことを確認し、遠心分離 (5000 rpm, 4℃, 10 分) 後、上清を除去し た。その後、0.25% trypsin-EDTA を 500 μl 加え 37℃で 15 分間処理し、等量の 10% FBS 添加 DMEM 培地を加えてトリプシン反応を停止した。細胞懸濁液を M2 培地に置換し、FACSAria II (Becton Dickinson) によりセルソーティングに 供試した。
Fluorescence activated cell sorting (FACS) において GFP の蛍光強度が高い 細胞集団を PGCs と判断し、ソーティングゲートを作製した。その後、RNA-seq
に 供 試 す る サ ン プ ル は 、 1 × 104 細 胞 を 1 プ ー ル と し て 75 μl 1%
β-mercaptoethanol 添加 RLT buffer (RNeasy Micro Kit; QIAGEN) 中に、ChIP-seq
に供試するサンプルは、1×105細胞を 1 プールとして 20 μl Nuclei ez lysis buffer
(Sigma) 中に分取し、使用時まで-80℃で保存した。また、一部 (2×104細胞) を
ソーティング精度確認用に 0.4% BSA 添加 PBS 中に回収した。精度確認は、PE Mouse anti-SSEA-1 抗体 (1/50 dilution, 560142, BD Pharmingen) を用いた分取 後細胞の免疫染色により行い、GFP および Phycoerythrin (PE) 双方の蛍光が検
20
出される細胞をカウントし、精製度 (%) を算出した (Figure 2-1)。
4. total RNA 抽出および RNA-seq ライブラリの構築
回収後の雌雄 PGCs およびマウス ES 細胞の total RNA 抽出を RNeasy Micro Kit (QIAGEN) を用いて行った。キットの定法に従い、DNase 処理および total RNA を抽出し、ゲノム DNA が完全に除去されているかは PCR 増幅によって判
定した。各 RNA サンプル 1 μl を Gapdh 遺伝子座特異的プライマー (Forward;
5’-ACCACAGTCCATGCCATCAC-3’, Reverse;
5’-TCCACCACCCTGTTGCTGTA-3’) および Ex Taq (TaKaRa Bio)を用いて 94℃ 10 分、(98℃ 10 秒、55℃ 30 秒、72℃ 30 秒) ×40 サイクル、72℃ 5 分の条 件で PCR 増幅し、電気泳動後に増幅産物が検出されないことを確認した。 精製後の RNA サンプルを 2100 Bioanalyzer RNA 6000 pico kit (Agilent) に供 試し、RIN (RNA integrity number) 値が 0.8 を超えるサンプルのみを RNA-seq ライブラリ構築に使用した。cDNA 合成および増幅は、SMART-Seq v4 Ultra Low Input RNA Kit for Sequencing Kit (Clontech) を用いて行い、実験手順および Long distance-PCR の条件設定は、キット付属のプロトコルに準拠した。各増幅 後 cDNA サンプルを Covaris S2 ソニケーターをにより 200 bp 長に裁断し、 NEBNext Ultra DNA Library Prep Kit for Illumina および NEBNext Multiplex Oligo for Illumina (New England Biolabs) を用いてライブラリ化した。構築した RNA-seq ラ イ ブ ラ リ サ ン プ ル を Agecourt AMPure XP ビ ー ズ (Beckman coulter) を 用 い て 精 製 し 、 各 ラ イ ブ ラ リ の 濃 度 と ク オ リ テ ィ を 、 2100
21
Bioanalyzer High sensitivity DNA kit (Agilent) お よ び KAPA Library Quantification Kit (NIPPON Genetics) により測定した。その後、各ライブラリ 濃度を 10 nM に調整し、Hiseq 2500 次世代シークエンサー (Single end, 100 bp; Illumina) に供試した。
5. RNA-seq リードアライメントおよび定量解析
4. で出力されたシーケンスリードを、CLC Genomic Workbench ver9.5 (CLC Bio) を 用 い て マ ウ ス レ フ ァ レ ン ス 配 列 (mm10, Genome Reference Consortium Mouse Build 38) にアライメントし、mm10 UCSC gene annotation データを使用し、アノテーションを行った。出力された BAM ファイルを Strand NGS (Agilent) に入力し、補正方法として TMM (Trimmed mean of M values) 法 を採用し各転写産物の RPKM 値を算出した。
6. 性特異的発現遺伝子群の同定
各 RNA-seq データセットの遺伝子発現量を可視化するため、R ggplot2 およ び psych パッケージを用いて散布図 (Scatter plot) を作製した。各 2 サンプルの Biological replicate の遺伝子発現量を平均化し、雌雄 PGCs 間で発現量に差があ る 遺 伝 子 群 を 2 fold change お よ び 統 計 検 定 (Moderated t-test with Benjamini-Hochberg FDR, p<0.05) によって同定した。
22
7. GO 解析およびパスウェイ解析による性特異的発現遺伝子群の生物学的意義 の検証
雌 雄 PGCs の 各 性 特 異 的 発 現 遺 伝 子 群 リ ス ト を DAVID web tool (http:/david.abcc.ncifcrf.gov/) に入力し(Huang da et al., 2009)、検出された雌雄 の性特異的発現遺伝子群中にどのような特性をもった遺伝子群が濃縮されてい るか検証を行った。Fisher exact test によって p<0.1 を示す GO term (Biological process) およびパスウェイを有意に濃縮されている term とし、生物学的意義の 考察を行った。 8. インプリント遺伝子群の発現アレル解析 各インプリント遺伝子 (Dlk1, Zdbf2, Snrpn および Ube3a 遺伝子) の発現状態 を BDF1 E13.5 雌雄 PGCs で解析するため、それぞれのサンプルを FACS 後、1 ×104 細胞を 1 プールとして 75 μl 1% β-mercaptoethanol 添加 RLT buffer
(RNeasy Micro Kit; QIAGEN) 中に回収した。Total RNA 抽出は、4. と同様の方 法 を 用 い て 行 い 、 Oligo (dT) プ ラ イ マ ー 及 び SuperScript Ⅲ Reverse Transcriptase (Invitrogen) を用いた逆転写反応により cDNA を合成した。その 後、PCR によって目的の領域を増幅し、電気泳動を行った。ゲル精製後、ダイ レクトシークエンスを行い、シークエンス波形から各インプリント遺伝子の発 現アレルを解析した。使用したプライマーは Table 2-1 を参照。
23
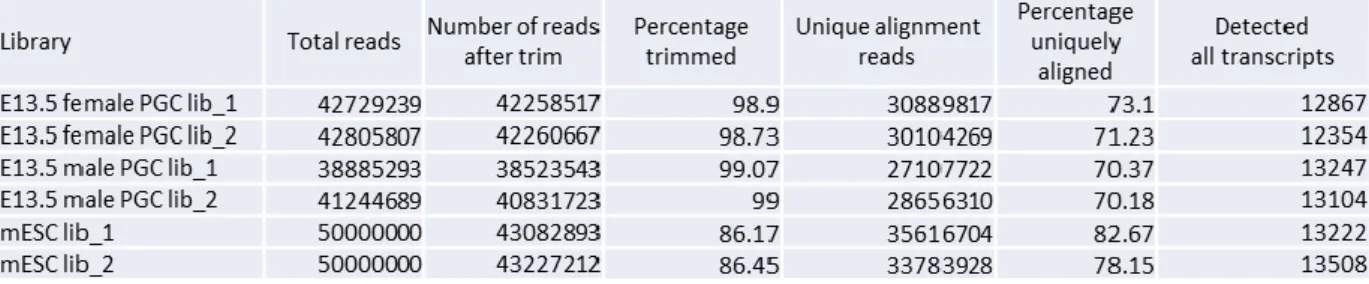
Figure 2-1. FACS 法による E13.5 雌雄 PGCs の採取
(A) FACS 法による E13.5 雌雄 PGCs 採取の略図。雌雄生殖巣を採取後、精索の有無により 雌雄判別を行った。生殖巣細胞懸濁液を FACS AriaII セルソーター (BD Biosceience) に供 試し、RNA-seq に用いる E13.5 雌雄 PGCs サンプル (1×104
cells/ 1 tube) を採取した。(B) FACS 後 E13.5 雌雄 PGCs の免疫染色結果。赤 (PE)が SSEA1 のシグナルを、緑が GFP 蛍 光を示す。FACS の精度確認のため、これらの細胞数を計測した結果、回収した PGCs の 97.5%が GFP および PE 双方のシグナルをもつ細胞であった。
24
Denaturation Annealing Extention Cycle Dlk1_F1 GGACAGGCCATCTGCTTCAC Dlk1_R1 CGGGGAAGATGATATTGACCGC Ube3a_F1 ATTGTGGGCTGGAACAGCAG Ube3a_R1 GCACCACACTGTTTCCTTGACTT Snrpn_F1 ACCGGAATGGCTTGTAAATCTTAGC Snrpn_R1 GTGAAACCAAATGTGTTTCAGTGGT Zdbf2_F1 GACAGTGGCTCATGAACTCCAG Zdbf2_R1 AGCCCCTTGAGGTTGATCATTAG 196 bp 368 bp 220 bp 268 bp
Primer name Sequence Amplicon length PCR condition for Ex Taq HS polymerase
98℃, 10sec 98℃, 10sec 98℃, 10sec 98℃, 10sec 62℃, 30sec 62℃, 30sec 62℃, 30sec 62℃, 30sec 72℃, 30sec 72℃, 30sec 72℃, 30sec 72℃, 30sec 35 35 35 35 Table 2-1. インプリント遺伝子群の発現アレル解析に使用した各プライマー配列と PCR 条件
25 第三節 結果 1. FACS 法による E13.5 雌雄始原生殖細胞の回収率および純度 Pou5f1-⊿PE-GFP 雄マウスと B6N 雌マウスの交配により得られた E13.5 胎 仔から、GFP 陽性を示す細胞を PGCs として FACS 法による回収を試みた。 PGCs を含む生殖巣細胞懸濁液の GFP 強度を FACS により解析した結果、102 ―103にピークをもつ細胞集団と 104―105にピークをもつ細胞集団が観察され た (Figure 2-1)。従って、104―105にピークをもつ細胞集団を PGCs として回 収し、以降の解析に供試した。回収できた PGCs は、雌雄共に一胎仔あたり 4000 ―6000 細胞であり、Anti-SSEA (PE conjugated) 抗体を用いた免疫染色の結果、 回収した PGCs の 97.5%が GFP および PE 双方のシグナルをもつ細胞であるこ とが明らかになった (Figure 2-1)。
2. E13.5 雌雄始原生殖細胞の遺伝子発現プロファイルと性特異的発現遺伝子 群の抽出
SMART 法により作製した E13.5 雌雄 PGCs およびマウス ES 細胞の RNA-seq ライブラリを、Hiseq2500 次世代シークエンサーを用いて Single end, 50 bp の条件でシークエンスしマウスレファレンス配列 (mm10) にマッピング
した結果、高いマッピング効率とともに (70.18―82.67%)、28M―35M の Unique
mapped reads がそれぞれ得られた (Table 2-2) 。また、性染色体の影響を除く ため、Y 染色体は、マッピングから除去した。RPKM 値が 0.1 を超える転写産 物を発現遺伝子として検出した結果、雌性 PGCs (2 biological replicates) で 12,867 および 12,354 transcripts が、雄性 PGCs (2biological replicates) で
26
13,247 および 13,104 transcripts が、マウス ES 細胞 (2 biological replicates) で 13222 および 13508 transcripts がそれぞれ検出された。また、雌雄 PGCs 間の 総転写産物数をカイ二乗検定によって比較した結果、雄性 PGCs の方が有意に
高い転写産物数を保有していることが明らかになった (p value = 1.6×10-22
)。 さらに、mm10 UCSC gene annotation データを使用し、発現している転写産物 の Gene type を検証した結果、E13.5 雌雄 PGCs およびマウス ES 細胞で検出 されたほとんどの遺伝子は、Protein coding 遺伝子であることが明らかになった (Figure 2-2)。加えて、各サンプル間の遺伝子発現プロファイルをもとに、ピア ソン相関係数を算出した結果、雌雄 PGCs 間で r=0.97―0.98、PGC およびマウ ス ES 細胞間で r=0.86 の正の相関が観られた。また、Biological replicate 間の相 関係数も r=0.98―0.99 と高い値を示すことから細胞のもつ遺伝子発現状態を強 く反映した高品質のライブラリであることが確認された (Figure 2-3)。興味深い ことに、既知の PGCs 特異的発現遺伝子群 (Sabour et al., 2011) の発現状態を
RNA-seq データセットから検証した結果、Fkbp6、4930432K21Rik および Pik3r3
は、マウス ES 細胞においても雌雄 PGCs と同程度の発現を示していた (Figure 2-4)。
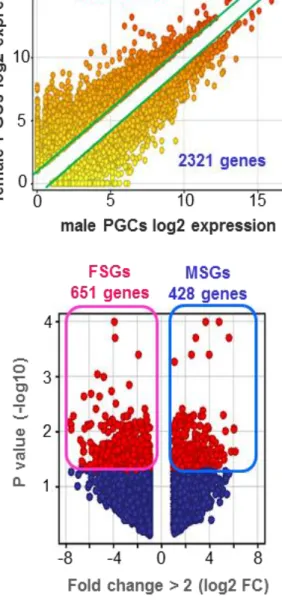
転写産物の性差に焦点をあて、雌雄 PGCs 間で発現量に差がある遺伝子群を 2
fold change により検出した結果、それぞれ 3769 個および 2321 個の性特異的遺 伝子群が抽出された。倍数変化の手法のみでは発現量の低い遺伝子群に多くの 偽陽性遺伝子が含まれてしまうため、これらを除くため、Moderated t-test with Benjamini-Hochberg FDR 法を用いて統計検定を行い、p 値が 0.05 以下のものの
27
みを真の性特異的発現遺伝子として検出した。その結果、651 個の雌特異的発現 遺伝子群 (Female specific expressed genes: FSGs) および 428 個の雄特異的発 現遺伝子群 (Male specific expressed genes: MSGs) の同定に成功した (Figure 2-5)。これにより、この時期 (E13.5) の雌雄 PGCs は、すでに性特異的な運命
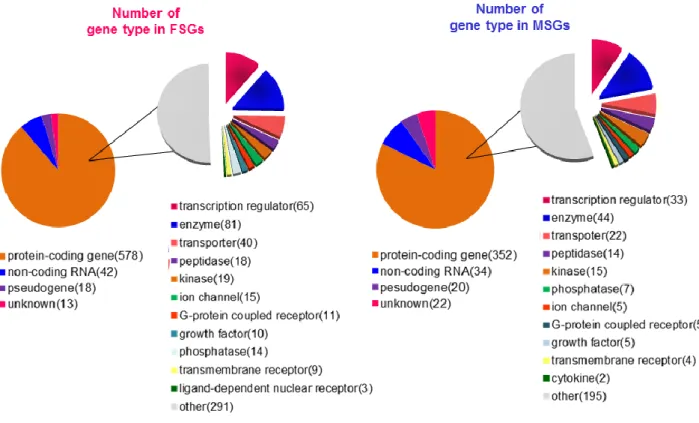
を獲得していることが明らかになった。加えて、UCSC および Ingenuity Pathway
analysis (IPA) のアノテーション情報をもとに、各性特異的遺伝子群の Gene type を検証した結果、雌雄共に多くの転写制御因子群が含まれていることが明 ら か に な っ た (65 in FSGs, 33 in MSGs, Figure 2-6) 。 Mouse Genome Informatics (MGI) データベースのアノテーション情報をもとにこれらの転写制 御因子群を機能別にクラスタリングした結果、C2-H2 type の DNA 結合ドメイン をもつ Zinc finger タンパク質が多く濃縮されていることが明らかになった。さ らに、これらの転写制御因子群の補因子として FSGs 内には、Tgfb1i1、Cited1、
Grip1、Nfkbia ならびに Lmo4 が、MSGs 内には、Epas1、Optn、Actn2 ならび
に Btg1 が検出されたため、これらの補因子がターゲットとなるそれぞれの転写 制御因子の活性を制御することで、雌雄 PGCs の性特異的な運命決定に寄与す る可能性が示唆された。
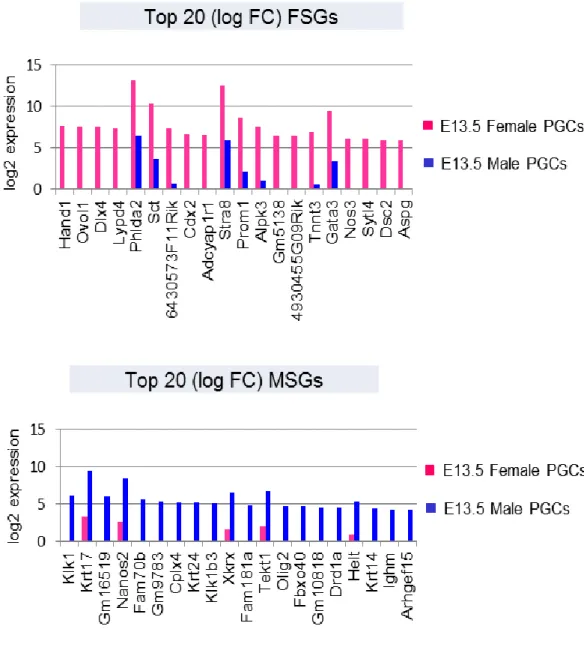
興味深いことに、各性特異的発現遺伝子群のリストを発現変動値 (Log FC value) をもとに降順に並べ替えた結果、FSGs Top 20 の遺伝子群には、Hand1、
Cdx2 および Gata3 のような胚体外組織分化を促進する因子群が、MSGs Top 20
の遺伝子群には、Olig2 および Helt といった神経細胞分化に関与する因子群が含 まれていることが明らかになった (Figure 2-7)。
28
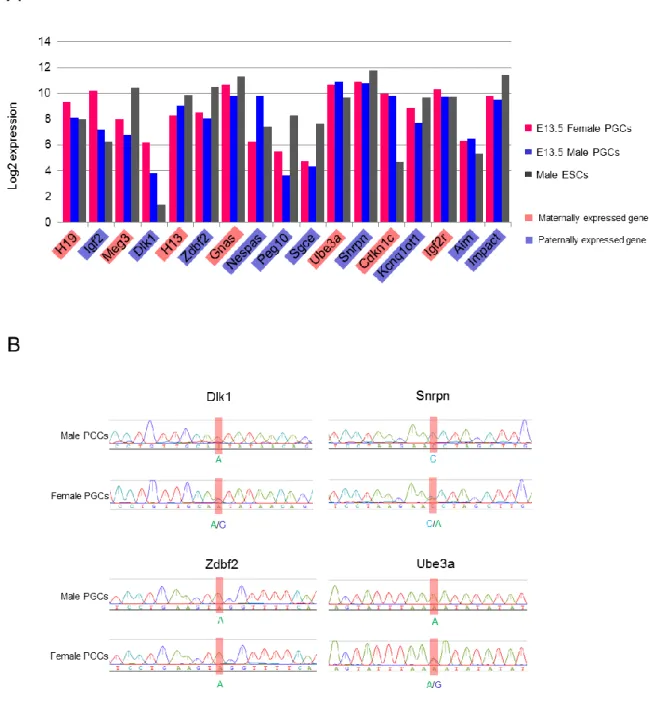
ゲノム中に刷り込まれた DNA メチル化インプリントは、E10.5―11.0 の PGCs において特異的に発現する Tet1 (Ten-eleven translocate 1) が仲介する酸化反応 によって消去されることが報告されている(Yamaguchi et al., 2013b)。従って、 メチル化インプリントが消去された E13.5 雌雄 PGCs 内でインプリント遺伝子 群がどのような発現状態を示すか明らかにするため、RNA-seq データセットを もとに各インプリント遺伝子群の発現解析を行った。興味深いことに、父方発 現インプリント遺伝子である Igf2 および Dlk1 が、雄性 PGCs と比較して雌性 PGCs で有意に高い発現量を示していた。さらに Igf2、Dlk1、Meg3、Peg10、 Sgce ならびに Cdkn1c 遺伝子は、雌雄 PGCs においてマウス ES 細胞と比較し て 2―3 倍の発現量を保有していることが明らかになった (Figure 2-8A)。さら に、各インプリント遺伝子が片アレル性発現を維持しているかどうか明らかに するため、B6N と DBA/2J 間に多型 (Single nucleotide polymorphism: SNP) が 検出された Snrpn、Ube3a、Dlk1 ならびに Zdbf2 遺伝子について発現アレル解 析を行った。Snrpn、Ube3a および Dlk1 遺伝子は、雌性 PGCs においては両ア レル性発現を示す一方、雄性 PGCs において片アレル性発現が検出された。ま た、Zdbf2 遺伝子は雌雄双方の PGCs で片アレル性発現を維持していた (Figure 2-8B)。これらの結果は、DNA メチル化の消去後も、他のエピゲノム修飾因子に よってインプリント遺伝子の片アレル性発現が部分的に維持されていることを 示唆している。 3. 性特異的発現遺伝子群の生物学的意義の検証 2. で同定された FSGs および MSGs の生物学的意義をさらに詳細に解析する
29
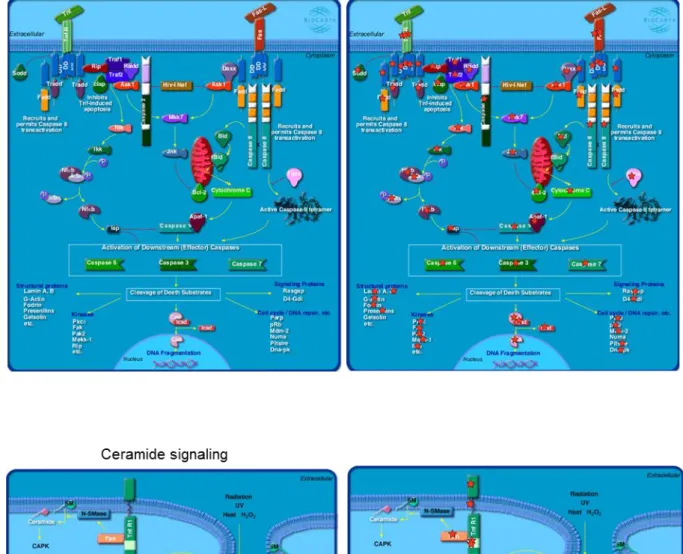
ため、DAVID web tool を用いて(Huang da et al., 2009)、各性特異的発現遺伝子 群の GO 解析およびパスウェイ解析を行った。FSGs において検出された GO term (Biological process) の中で、「regulation of transcription」、「regulation of transcription, DNA dependent」および「regulation of RNA metabolic process」 の term が Top 3 を占めており、転写制御因子群が FSGs 内に最も多く濃縮され ていることが示唆された。一方 MSGs では、「regulation of RNA metabolic process 」、「 regulation of transcription, DNA dependent 」 お よ び 「 neuron differentiation」の term をもつ遺伝子群が多く濃縮されていることが明らかにな った (Figure 2-9)。これにより、FSGs および MSGs 間には、有意な機能的な差 があることが明らかになった。また、雌雄 PGCs 間の発現遺伝子群の差にどの ような生物学的意義が潜在しているかより詳細に解析するため、雌雄 PGCs の RNA-seq データセットにおける全ての発現遺伝子を DAVID の BioCarta プログ ラムに供試し、どのようなパスウェイが濃縮されているか検証を行った。その 結果、雌雄 PGCs においてそれぞれ 16 個および 21 個のパスウェイの濃縮が検 出された (Figure 2-10)。また、これらの内、4 個 (female pathway) および 9 個 (male pathway) のパスウェイが、もう一方の性では検出されない性特異的なパ スウェイであった。雌性 PGCs 特異的パスウェイとしては、「HIVnef Pathway: negative effector of Fas and TNF」、「Angiotensin II mediated activation of JNK pathway via Pky2 dependent signaling」、「Rho cell motility signaling pathway」 および「Telomeres, telomerase, cellular aging and immortality」の外因性アポト
30
雄性 PGCs 特異的パスウェイとして、「Cyclin and cell cycle regulation」、「Control of gene expression by vitamin D receptor 」、「mTOR signaling pathway 」、 「 Ceramide signaling pathway 」、「 p53 signaling pathway 」、「 Role of mitochondria in apoptotic pathway」、「Skeletal muscle hypertrophy is regulated via AKT/mTOR pathway 」、「 Rac1 cell motility signaling 」 お よ び 「 Human cytomegalovirus and MAP kinase pathway」の細胞周期調節、DNA 傷害/ミトコ ンドリア性 (内因性) アポトーシスならびに細胞老化に関与する 9 つのパスウ ェイが濃縮されていた (Figure 2-11)。HIVnef Pathway を構成する 48 遺伝子の うち、Traf1、Nfkbia ならびに Casp6 などの外因性アポトーシスパスウェイの中 核となる遺伝子群が、雌性 PGCs において特異的発現が観察された (Figure
2-11A)。一方雄性 PGCs においては濃縮されていた「Ceramide signaling pathway」
は近年までの研究によって、細胞膜上に局在する Ceramide 分子がセカンドメッ センジャーとして働き、アポトーシス、細胞老化、細胞周期ならびに分化の様々 な細胞プロセスに関与していることが報告されている (Figure 2-11B)。加えて、 雄性 PGCs 特異的なパスウェイとして「p53 signaling pathway」および「Role of mitochondria in apoptotic pathway」も濃縮されていることから、E13.5 雄性 PGCs において内因性のアポトーシスシグナルが活性化していることが示され た。これらの結果より、E13.5 雌雄 PGCs において雌性 PGCs においては外因 性アポトーシスシグナルを、雄性 PGCs では内因性アポトーシスシグナルを介 した生殖細胞削減プログラムがすでに活性化している可能性が示唆された。ま た、体細胞分裂制御に関与する「MAP kinase signaling pathway」および「Cell
31
cycle: G1/S check point」などのパスウェイが、双方の PGCs で強い濃縮が確認 された (Figure 2-10, Figure 2-12A)。このことは、生殖巣に到達した雌雄 PGCs の細胞集団中に、未だ有糸分裂期にある細胞群がある程度存在していることを 示唆している。また、細胞の生存および細胞死双方に関与する因子を包括して いる「Ras pathway」も雌雄 PGCs において検出された (Figure 2-12B)。これに より、雌雄 PGCs 双方の細胞集団中に、生の運命決定を受けた細胞群と死の運 命決定を受けた細胞群の 2 つが混在していることが示された。これらの結果に より、雌雄 PGCs の転写不均質性が示唆されたため、個々の PGC の動態をより 詳細に解析するためにはシングルセル RNA-seq 解析が必須である。
32
33
Figure 2-2. E13.5 雌雄 PGCs および ES 細胞サンプル全転写産物中における 各 Gene type の割合
円グラフ中の%は UCSC Genome Browser のアノテーション情報に由来する各 Gene type (Protein coding gene、non-coding RNA、pseudogene および unknown) の割合を示す。
34
Figure 2-3. E13.5 雌雄 PGCs および ES 細胞サンプル各遺伝子発現プロファイル の比較解析結果
左の散布図は、各サンプル間の遺伝子発現プロファイルを反映したおり、右図に含まれる数値 は、各サンプル間のピアソン相関係数を示す。
35
Figure 2-4. PGCs 特異的発現遺伝子 (Sabour, 2011) の発現解析結果
各遺伝子の発現量は、log2 RPKM 値で表されており、各サンプルの発現量は棒グラフで表示 されている (pink= E13.5 female PGCs, blue= E13.5 male PGCs, grey= male ESCs)。
36
Figure 2-5. E13.5 雌雄 PGCs における性特異的発現遺伝子群の同定
上段の散布図は、2 fold change によって抽出された各性特異的発現遺伝子群を示す (female: 3769, male: 2321)。これらの遺伝子群から偽陽性を除くため統計検定
(Moderated t-test with Benjamini-Hochberg FDR) を行った。下図の Volcano plot 内の Red spheres が真の性特異的発現遺伝子群 (FSGs: 651 genes, MSGs: 428 genes)を示す。
37
Figure 2-6. 各性特異的遺伝子群の遺伝子特性 (Gene type)
左の円グラフは UCSC Genome Browser のアノテーション情報に由来する Gene type であり、 右の円グラフは Protein coding gene に分類された遺伝子を Ingeneuity Pathway Analysis (IPA) のアノテーションをもとにさらに詳細に分類した Gene type と割合を示す。( )内の数値は 遺伝子数を示し、右の 2 つの円グラフは FSGs 内の、左は 2 つの円グラフは MSGs 内の Gene type の割合を示す。
38
Figure 2-7. 各性特異的発現遺伝子 (上位 20 個) の発現解析結果
上段の棒グラフは FSGs の内、最も発現変動値 (log2 FC value) が高い上位 20 個の遺伝子を降順 に示しており、下段の棒グラフは MSGs の内、、最も発現変動値 (log2 FC value) が高い上位 20 個 の遺伝子を降順に示している。各遺伝子の発現量は log2 RPKM 値で表されており、各サンプルの発 現量は棒グラフで表示されている。(pink= E13.5 female PGCs, blue= E13.5 male PGCs)
39
Figure 2-8. 各インプリント遺伝子の発現アレル解析結果
(A) RNA-seq データセットを用いた代表的なインプリント遺伝子の発現解析結果。 各遺伝子の発現量は、 log2 RPKM 値で表され、各サンプルの発現量は棒グラフで表示されている (pink= E13.5 female PGCs, blue= E13.5 male PGCs, grey= male ESCs)。赤でハイライトされた遺伝子は母方発現インプリント遺伝子 であり、青でハイライトされた遺伝子は父方発現インプリント遺伝子である。(B) BDF1 ゲノムを用いたインプ リント遺伝子 (Dlk1、Snrpn、Zdbf2、Ube3a) の発現アレル解析結果。赤でハイライトされた塩基は
40
Figure 2-9. 性特異的発現遺伝子群の GO 解析結果
DAVID web tool による FSGs (上段) および MSGs (下段) の GO 解析の結果、最も濃縮されて いた上位 10 個の GO term (Biological process) を棒グラフで示す (Fisher’s exact test: cut-off < 0.1).。横軸は濃縮されている遺伝子数を示す。
41
Figure 2-10. 雌雄 PGCs 全発現転写産物の BioCarta パスウェイ解析結果
DAVID web tool による雌雄 PGCs 全発現転写産物の BioCarta パスウェイ解析結果。リスト中の「Count」 は遺伝子数を、「p value」はFisher’s exact test により算出された p 値を示す。ハイライトされているパスウェ イ (term) は性特異的パスウェイを示す (pink= female PGCs specific pathway, blue= male PGCs specific pathway)。
42
Figure 2-11. E13.5 雌雄 PGCs における代表的な性特異的 BioCarta パスウェイ
(A)雌性 PGCs 特異的な「HIV-I Nef: negative effector of Fas and TNF」パスウェイ。(B) 雄性 PGCs 特異的な「Ceramide signaling」パスウェイ。右図のスターでハイライトされている遺伝子は、雌雄 PGCs で発現している遺伝子を示す。
43
Figure 2-12. E13.5 雌雄 PGCs における代表的な Common BioCarta パスウェイ
雌雄 PGCs で共通して濃縮されていた MAP kinase pathway (A) および Ras signaling pathway(B)。 右図のスターでハイライトされている遺伝子は、雌雄 PGCs で発現している遺伝子を示す。
44 第四節 考察 本研究における E13.5 雌雄 PGCs の包括的トランスクリプトーム解析によっ て、性特異的な生殖細胞の運命決定を支持する性特異的発現遺伝子群の同定に 成功した。また、これらの解析結果は、生殖巣到達直後の雌雄 PGCs がすでに 性特異的な細胞系譜を獲得していることを示している。 E13.5 までに後の生殖巣となる生殖隆起に到達した PGCs は、Sry の発現の有 無によって構築された体細胞の性によって性特異的な分化を起こすことが報告 されている (Tanaka and Nishinakamura, 2014)。近年の研究によって、雌性 PGC における Stra8 の発現を起点とした減数分裂関連遺伝子の発現上昇および雄性 PGC における Nodal signaling 関連遺伝子の発現上昇など、E13.5 PGCs におけ る雌雄差が部分的に明らかにされてきている (Baltus et al., 2006; Wu et al., 2013; Yokobayashi et al., 2013)。本研究における定量性および包括性に優れた RNA-seq 解析 (deep sequencing) によって性特異的な転写因子群、アポトーシ ス関連遺伝子および細胞発達関連遺伝子群を含む以前の研究に無い、より多く の性特異的遺伝子群 (FSG: 651 genes, MSG: 428 genes) が E13.5 雌雄 PGCs において同定された。実際、以前の Jameson らのマイクロアレイを用いて抽出 された germ cell primed genes の遺伝子リスト (Jameson et al., 2012)と本研究 によって抽出された雌雄それぞれの性特異的発現遺伝子群リストを比較した結 果、一致した遺伝子は非常に少なく FSG で 27 遺伝子、MSG で 42 遺伝子であ った。従って、本研究における様々な発見は、雌雄 PGCs における性特異的な 細胞特性の獲得と維持について新たな洞察を与えた。
45
加えて、本研究において同定された FSGs 中には、「regulation of transcription」
の term も持った遺伝子群が最も多く存在し、興味深いことに、これらの転写制 御関連遺伝子群の中には、Cdx2 や Hand1 などの栄養膜外胚葉系譜の遺伝子が 含まれていた(Scott et al., 2000; Strumpf et al., 2005)。一般的に、胚盤胞内部細 胞塊および ES 細胞において、Cdx2 は Pou5f1 の発現によって抑制されること から(Niwa et al., 2005)、雌性 PGCs において、これらの転写因子は、現在まで に報告されている転写制御機構とは全く異なる方法によって、性特異的な分化 を支持している可能性が考えられる。一方、MSGs に特異的な転写因子群に着 目すると、33 個中 11 個の遺伝子が神経系細胞において増殖や分化を制御するこ とが報告されている転写因子であった。近年の研究によって Sry の発現によっ て生じたセルトリ細胞では、Notch シグナル応答経路が働いていることが明らか にされ、この経路は主に神経系において分化抑制と幹細胞プールの維持に関与 することが報告されている (Garcia and Hofmann, 2013; Grandbarbe et al., 2003)。本研究において観察された雄性 PGCs の神経細胞様の遺伝子発現パター ンは、セルトリ細胞の分化抑制シグナルに PGCs が生殖巣到達後、すぐに応答 していることを示唆している。
また、以前の研究によって E13.5 雌雄 PGCs においてプログラム細胞死様の 機構が働き、生殖細胞の選別が生じることが明らかにされている。FSGs および MSGs の DAVID web tool を用いた BioCarta パスウェイ解析の結果、雌性 PGCs では外因性のアポトーシス経路が、雄性 PGCs では DNA 傷害/ミトコンドリア 性 (内因性) アポトーシス経路が濃縮されていた。このことは、雌雄それぞれの
46 生殖系列において異なるアポトーシスシグナルによる「選別」経路を経ている ことを示している。しかしながら、雌雄 PGCs 双方で濃縮されていた「MAP kinase pathway」では、細胞周期 (G1→S 期→G2 期→M 期) の全ステージの制 御因子が、雌雄 PGCs それぞれの全転写産物内に含まれており、また「Ras signaling pathway」では、生存および細胞死に寄与する因子双方が全転写産物 内に存在することが明らかになった。これにより、E13.5 雌雄 PGCs が転写不 均質性をもった細胞集団であることが示唆されたため、単一細胞レベルで再度 雌雄 PGCs 間の網羅的トランスクリプトーム解析を行い、細胞周期およびアポ トーシス関連遺伝子群に転写的ヘテロ性が観られるかどうか詳細に解析するこ とが必須である。
我々は以前に Post bisulfite adaptor tagging (PBAT) 法を用いた雌雄 PGCs の 全ゲノム包括的な DNA メチル化解析によって、E13.5 の雌雄 PGCs ゲノムが、 一部のレトロトランスポゾン領域を除いて低メチル化状態を示すことを明らか にしている (CpG methylation level female: 3.2%, male: 3.8%)。これと一致して、 共同研究により行った性特異的発現遺伝子群のプロモーター領域 (TSS ±5 kb) も雌雄 PGCs 間で共通して低メチル化状態を示していた (<3%)。このこと は、DNA メチル化状態が性特異的な遺伝子発現制御機構に寄与していないこと を示唆している。 加えて、近年の ChIP-seq 法を用いた研究よって、ヒストンの修飾パターンが マウス PGCs における遺伝子発現を制御することが部分的に明らかにされてい
47
る。実際、転写活性化マーカーである H3K4me3 および H3K27ac の蓄積が、生 殖細胞特異的発現遺伝子である Zic3 や Dazl の TSS もしくはエンハンサー領域 上に検出されており、また PGCs で発現が抑制されている Dlx や Hoxa などの
ホメオボックス遺伝子上には、転写抑制マーカーである H3K4me3 と H3K27me3
が共局在する Bivalent chromatin mark が修飾されていることが報告されている (Sachs et al., 2013)。共同研究における我々の ChIP-seq データも同様な傾向を 示し、さらに性特異的発現遺伝子群のプロモーター領域における H3K4me3 と H3K27me3 の修飾パターンは雌雄それぞれで明らかに異なっていた。従って、 H3K4me3 と活性化シグナルと H3K4me3-H3K27me3 の抑制化シグナルによっ て性特異的遺伝子の発現は部分的に支配されていることが示唆される。しかし ながら、性特異的遺伝子群中には、H3K4me3 および H3K27me3 どちらのピー クも検出されない領域も存在していたので、今後さらに、ATAC-seq や H3K27ac の ChIP-seq など行い、性特異的遺伝子発現パターンにどのようなクロマチン構 造が寄与するか明らかにすることが必須である。
48 第三章 シングルセルトランスクリプトーム解析によるマウス雌雄始原生殖細 胞集団における転写不均質性の証明 第一節 緒言 哺乳動物の発生は単一細胞である受精卵から開始され、その後の各細胞は発 生過程における微小環境からのシグナルもしくは自身がもつエピゲノム修飾変 化に応じて約 200―250 種の細胞種に分化することが知られている。従来より、 体細胞数が 1000 細胞前後である線虫では、個々の細胞の機能解析が行われてき たが(Potts and Cameron, 2011)、数百種類の細胞から構成されているヒトをは じめとした哺乳動物においては個々の細胞の挙動を観察するシングルセル解析 の手段は、フローサイトメトリーや蛍光イメージングのみであり極めて限定的 であった。しかしながら近年、単一細胞の捕捉、微量 DNA の増幅および次世代 シークエンサーによる高出力解析など、各分野で目覚ましい発展が観られ、細 胞の遺伝子発現プロファイルをシングルセルレベルで解析することが可能とな った。 これにより、近年の研究では上記の技術を用いて、解析対象となる細胞集団 の転写不均質性に焦点を当てた研究が多く進められ、実際、腫瘍細胞、ES 細胞 および神経前駆細胞など多くの細胞種で特定の遺伝子発現の「ゆらぎ」が検出 されている。従って、シングルセルレベルの網羅的トランスクリプトーム解析 は、細胞間の生物学的ばらつき、分化過程および周辺細胞との相互作用などバ ルクの解析では得られなかった新たな知見を見出すための非常に有効な手段と
49 なっている。 第二章で述べた雌雄 PGCs の BioCarta パスウェイ解析の結果、本研究で対象 としているマウス PGCs も転写不均質性をもつ細胞集団であることが強く示唆 された。そのため、同時期の雌雄 PGCs を用いてシングルセル RNA-seq 解析を 行い、個々の雌雄 PGCs 間に生物学的ばらつきが観られるかどうか、また細胞 間で高い発現量の差が検出される変動遺伝子群の同定とその生物学的意義の検 証を行った。
50 第二節 材料および方法 1. 供試動物 供試動物および飼育条件は第二章の方法に準じた。 2. E13.5 雌雄 PGCs の回収 第二章の方法に準じて、FACS ソーティング法により 5000 細胞の E13.5 雌 雄 PGCs を 100 μl 0.4%BSA 添加 PBS 中に採取した。その後、採取した PGCs を C1 Single-Cell Auto Prep System (Fludigm) に供試するため、遠心分離 (3000 rpm, 4℃, 30 分) 後上清を除去し、5 μl 0.4%BSA 添加 PBS 中に PGCs を再懸濁 した。
3. 雌雄 Single PGC の捕捉および cDNA 合成
2. で採取した雌雄それぞれの E13.5 PGCs 細胞懸濁液全量 (1000 cell/μl) を、 集積回路 (Medium-sized (10―17 μm cell diameter) Integrated fluidic circuit (IFC), Fludigm) 上 の ウ ェ ル に 添 加 し 、 C1 Single-Cell Auto Prep System (Fludigm) に よ り 単 一細 胞 の 捕 捉 を 行 っ た。 また、 同時に LIVE/DEAD Cell Viability/Cytotoxicity Kit (2 μM Calcein-AM and 4 μM Ethidium homodimer III in C1 Cell Wash Buffer, PromoKine) を用いて、細胞の生死判別も行い、生細胞の みを以降のライブラリ構築におけるサンプルとした。その後、SMARTer Ultra Low Input RNA HV Kit および Advantage 2 PCR Kit (Clontech) の各試薬を、C1
51
Single-Cell Auto Prep System の定法に従い、IFC 上のウェルに添加し、cDNA 合成および Long-distance PCR を行った。
4. シングルセル RNA-seq ライブラリの構築
回収された各 cDNA 合成後サンプル溶液 (3 μl each) を、96-well プレートに 移し、C1 DNA Dilution Reagent を用いて 4 倍希釈を行った。各サンプルの cDNA 増幅が確実に行われているかどうかは定量的 PCR 法によって判定した。各
cDNA サンプル 1 μl を Gapdh 遺伝子座特異的プライマー (TaqMan probe, ID:
Mm99999915_g1, Applied Biosystems) を用いて 50℃ 2 分、95℃ 10 分、(95℃ 15 秒、60℃ 1 分) ×45 サイクルの条件で TaqMan Master mix (Applied Biosystems) による PCR 増幅を行い、増幅曲線が検出されることを確認した。 その後各 cDNA サンプル 1.5 μl を、Nextera XT DNA Library Prep Kit および Nextera XT Index Kit -96 indices (Illumina) を用いてライブラリ化した。構築し た各シングルセル RNA-seq ライブラリサンプル 1 μl を、2100 Bioanalyzer High sensitivity DNA kit (Agilent) に供試し、各ライブラリの濃度を測定した。シー クエンス時に各サンプルで均一なリード数が得られるようにするため、測定濃 度に基づいて、各サンプルの濃度が 5 nM になるように 1 つのチューブにプール した。Agecourt AMPure XP ビーズ (Beckman coulter) を用いて精製し、200― 650 bp の断片のみを Pippin Prep および 2% agarose gel cassette (Sage science) を用いて抽出した。回収したサンプルを NucleoSpin Gel and PCR clean-up Kit (TaKaRa) を 用 い て 精 製 し 、 各 ラ イ ブ ラ リ の 濃 度 と ク オ リ テ ィ を 、 2100