2016 公益社団法人日本水産学会/The Japanese Society of Fisheries Science Tel81972322125.Fax81972322293.Emailyukinoris@aŠrc.go.jp †2016 年 4 月 1 日「国立研究開発法人(国研)水産総合研究センター」より組織名改称
a These authors contributed equally to this work.
ブリ卵の小型容器を用いた高塩分孵化管理方法の開発
嶋 田 幸 典,
1a石 川
卓,
1a名古屋博之,
1薄
浩 則,
1堀 田 卓 朗,
2吉 田 一 範,
2藤浪祐一郎,
2岡 本 裕 之
1(2016 年 1 月 6 日受付,2016 年 4 月 13 日受理,2016 年 6 月 30 日 JSTAGE 早期公開)
1(国研)水産研究教育機構†増養殖研究所,2同 西海区水産研究所
Development of an incubation method using a small container with high-salinity water for yellowtail YUKINORISHIMADA,1aTAKASHIISHIKAWA,1aHIROYUKINAGOYA,1HIRONORIUSUKI,1
TAKUROUHOTTA,2KAZUNORIYOSHIDA,2YUICHIROFUJINAMI2AND HIROYUKIOKAMOTO1
1Kamiura Laboratory, National Research Institute of Aquaculture, Japan Fisheries Research and Education
Agency, Saiki, Oita 8722602,2Goto Laboratory, Seikai National Fisheries Research Institute, Japan Fisheries
Research and Education Agency, Goto, Nagasaki 8530508, Japan
Our goal is to develop a strain of yellowtail Seriola quinqueradiata that exhibits economically desirable traits using genome editing. To prevent deaths caused by sinking of yellowtail eggs before hatching and develop a small container incubation method to rear the small number of eggs obtained by microinjection, we ˆrst determined the speciˆc gravity of eggs and identiˆed appropriate salinities for hatching. Then, we measured the hatching rates of eggs microinjected with GFP-mRNA and incubated at the appropriate salinities. The speciˆc gravity of eggs in-creased during development, exceeding that of sea-water 5 h before hatching. Normal hatching rates (i.e., exclud-ing abnormal larvae) ranged from 094.8 at salinities of 3060 psu, and were >80 at those of 3540 psu. The normal hatching rate of microinjected eggs kept at 40 psu was signiˆcantly lower (7.4) than that at 35 psu (56.7 ). The normal hatching rate of microinjected eggs at 35 psu was 80 similar to that of non-injected eggs at 35 psu. Thus, we can control hatching rates of microinjected yellowtail eggs in small containers using sea-water ad-justed to 35 psu. キーワード塩分,受精卵,比重,孵化率,ブリ ブリ Seriola quinqueradiata は親魚養成技術や種苗生 産技術の向上に伴い,育種による系統開発が試みられて いる。13)特に最近,著者らは優良経済形質を持つ系統 を作出するために,いくつかの候補遺伝子を標的にゲノ ム編集魚の作出のための基礎的研究に着手している。一 般にゲノム編集を行うには,受精卵に顕微注入する遺伝 子特異的に働く人工ヌクレアーゼの種類や濃度の検討か ら始められ,次に人工ヌクレアーゼのゲノム編集効率を もとに最適条件の検討が繰り返し行われる。4,5)ゲノム編 集の成否の評価には,ゲノム量が多くなる孵化後数日の 仔魚から DNA を抽出することが望ましい。しかし,ブ リの受精卵を小型容器で孵化管理する場合,未注入の受 精卵でも孵化前に著しい沈降現象が生じて死滅するた め,68)サンプルの安定確保やその後の飼育が容易では ない。これまでに 500 mL 容量の容器を用いた小型容器 孵化管理方法が開発されてきたが,6,7)同法では孵化率が 安定しない場合も多い。実際,著者らによる複数回にわ たる顕微注入試験における小型容器孵化管理において, 受精後 24 時間の受精卵は少なくとも 50 以上の正常 発生率を示してきたが,それらの孵化率は未注入卵でも 数パーセントまで低下した(著者ら,未発表データ)。 大型水槽による孵化管理では一般に通気による水の攪拌 で沈降を抑えるが,小型容器では攪拌強度の調節が難し く,8)安定して高い孵化率を得ることが困難である。ま
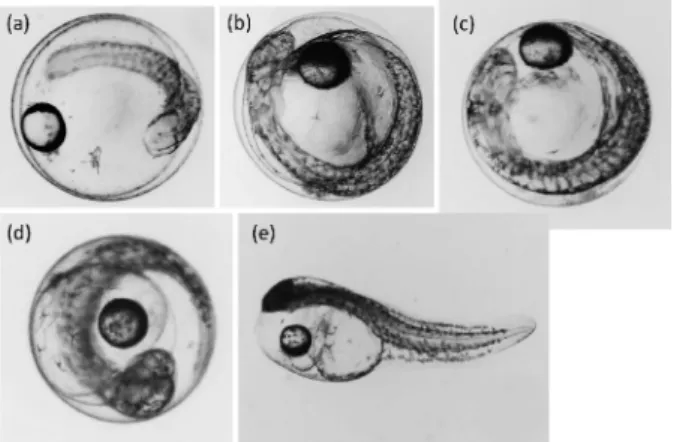
Fig. 1 Developmental stages of yellowtail eggs and lar-vae: 30 (a), 10 (b), and 5 h (c) before hatching (d) and (e) represent the egg and larval stages immediate-ly before and after hatching (0 h).
た,大型水槽を用いても無通気条件での孵化管理では, 卵が孵化前にネット底部に堆積し,まったく孵化しない ことが報告されている。8)さらに現在,魚類のゲノム編 集技術に用いられている顕微注入法では卵 1 粒ずつ 1 細胞期の胚盤内へ RNA を注入することから,一度に数 千から数万の大量の顕微注入卵を確保して大型水槽によ る孵化管理を行うことは難しい。したがって,ゲノム編 集を利用してブリ育種を適切に進めていくには,ブリ受 精卵における安定的な小型容器孵化管理方法の開発が急 務である。 いくつかの魚種では卵発生を適切に進行させて,養殖 用種苗として良質な孵化仔魚を得るために,受精卵に最 適な環境水の比重や卵比重の検討が行われてきた。例え ば,スズキ Lateolabrax japonicus の卵発生に伴う卵比重 は , 2 細 胞 期 か ら ク ッ パ ー 胞 の 出 現 ま で は 1.0200 1.0206 g/cm3と安定し,その後,孵化開始時に 1.0222 g/cm3と最大になるが海水の比重(1.0248 g/cm3)より 軽いため,孵化開始時でも卵が沈降することはない。9) 一方,大西洋サバ Scomber scombrus では卵発生に伴い 比重が受精直後の 1.0245 から 1.0205 g/cm3へと軽くな るが,10)スズキと同様に卵比重が海水の比重より軽いた めに,卵が沈降することはない。また,海水の比重は塩 分と密接に関連することから,中立浮上塩分(NBS, neutral buoyancy salinity)を指標にして卵比重が調べ られた報告もある。例えば,バルト海に生息するカレイ 目魚類(Limanda limanda, Pleuronectes platessa および Pleuronectes ‰esus)や大西洋タラ Gadus morhua では塩 分 14.520.8 psu で卵の中立浮上が保たれることが報告 されている。11,12)シロギス Silago japonica の NBS は, 産卵期間を通して多少の変動があるものの,塩分 30 psu 以下を示す。13)このように,多くの魚種では卵比重 が最も高くなる発生段階であっても平均的な海水の塩分 より低く,受精卵の浮上が観察されてきた。一方,ブリ やシマアジ Pseudocaranx dentex の受精卵は受精後 1 ま たは 2 日までは他の魚種と同様に海水表面に浮上して いるが,孵化のおよそ 10 時間前(ブリ水温 20°C で 受精後約 50 時間,シマアジ水温 22°C で受精後 36 時 間)には卵の沈降が始まる。68,14)ブリ受精卵の卵比重 は,孵化 12 時間前に卵比重が海水の比重(1.0252 g/ cm3)を 上回り,孵化 6 時間前には 1.02631.0274 g/ cm3を示すことが報告されている。8)本種では種苗生産 のための孵化管理は,通気による水の攪拌で卵の沈降を 防止できる大型水槽を用いて行われる。8)大型水槽と同 等の高い孵化率を小型容器で実現するには,卵の沈降を 防止する新たな技術を開発する必要がある。 筆者らは顕微注入を行ったブリ受精卵において,環境 水の比重を卵比重より高くすることで沈降を抑制する技 術の開発を試みた。本研究では,ブリ受精卵の発生に 伴う卵比重の変化を確認すること,6 穴マイクロプ レートを用いてブリ受精卵の孵化管理における最適な塩 分を検討すること(孵化塩分試験),実際に顕微注入 した受精卵を異なる塩分の海水に収容し,塩分が顕微注 入した受精卵の発生に及ぼす影響を明らかにすること (顕微注入卵による孵化塩分試験)を目的とした。 材料と方法 卵 比 重 の 測 定 ブ リ 受 精 卵 の 卵 比 重 の 測 定 に は , 2015 年 10 月 29 日に水産総合研究センター西海区水産 研究所五島庁舎(以下,五島庁舎)で人工授精により得 られた受精卵を使用した。人工授精は,成熟調査により 卵径 700 mm 以上を示した雌個体および腹部圧迫により 排 精 が 確 認 さ れ た 雄 個 体 に HCG ( human chorionic gonadotropin,ヒト絨毛性性腺刺激ホルモン,あすか製 薬株社製)を魚体重 1 kg あたり 600 IU 投与し,48 時 間後に得られた卵および精子を用いて行った。8)受精卵 は受精後 1 日に同センター増養殖研究所上浦庁舎(以 下,上浦庁舎)に輸送した。到着した受精卵は約 10 L の容器を用いて塩分 33.3 psu(海水の比重1.0248 g/ cm3),20°C の微通気条件で卵管理し,適時死卵を除去 した。正常発生している卵の 50 以上が孵化した 10 月 31 日の午後 8 時を孵化 0 時間とした。この時間から 逆算し,孵化 0 時間,5 時間前,10 時間前および 30 時 間前の受精卵(Fig. 1)の中立浮上塩分(NBS, neutral buoyancy salinity)を以下の方法で測定した。予め 200 mL メスシリンダーに十数粒のブリ受精卵と 100 mL の 海水を入れ,蒸留水または塩分 60 psu に調整した人工 海水(Instant Ocean, Aquarium Systems 社製)を加え ることにより,受精卵すべてが中立浮上を保つ塩分を測 定した。いずれも 2 回の測定を繰り返し,数値が大き
測定した塩分と比重の換算表を用いて算出した。孵化 0 時間と 5 時間前には完全に卵が浮上する塩分(positive buoyancy salinity)を調べた。なお,飼育に用いた海水 の塩分は,電導度計(EC300, YSI 社製)により測定し た。 孵化塩分試験 孵化塩分試験は 2015 年 10 月 29 日と 12 月 3 日に五島庁舎で卵比重の測定で使用した卵と同 様の方法で人工採卵した受精卵を用いた。予め塩分 30 と 60 psu の人工海水を作製し,それぞれ比率を変えて 混合することで異なる塩分の海水を作製した。1 回目の 試験は,塩分 35 から 60 psu まで塩分 5 psu ごとに 6 段 階の塩分区を作製した。2 回目の試験では,塩分 30, 35, 37.5, 40お よ び 45 psu の 5 段 階 と し た 。 抗 生 物 質 (ペニシリンストレプトマイシン溶液,和光純薬工業 株社製)を 1 添加した異なる塩分の海水を準備して, 6 穴マイクロプレートに 7 mL ずつ注入し,受精卵(受 精 36 時間後,胚体形成期)を 20 粒ずつ収容した。な お,抗生物質の投入前後の塩分に変化はなかった。その 後,孵化数および正常孵化数を測定した。1 塩分区あた り 3 回の繰り返しを行った。 異なる塩分における生残日数の変化と無給餌生残指数 (SAI, survival activity index)の算出 孵化率の高い 塩分環境が明らかになっても,孵化仔魚の生残率が著し く低下するなら小型容器孵化管理法の開発にはならな い。そこで,孵化後の仔魚に対する環境水中の塩分の影 響を調べるために,1 回目の孵化塩分試験で正常孵化仔 魚が得られた塩分 35, 40 および 45 psu の試験区につい ては孵化後,飼育を実施した。各塩分の孵化仔魚を毎日 観察して,生残数と死亡数を計数した。一般に,無給餌 生残指数(SAI, survival activity index)は生残日数に 重 み づ け を し て 表 し た 卵 質 評 価 の た め の 指 標で あ る が,8)本研究では試験に供した受精卵の卵質をこれまで の報告と比較すること,および異なる塩分間での孵化仔 魚に対する塩分の影響を評価するための指標として用い た。なお,無給餌生残指数は新間と辻ヶ堂15)の下記に 示した計算式にしたがって算出した。 SAI=1 N k
∑
i=1 (N-hi)×i N試験開始時の孵化仔魚数 hii 日目の累積斃死尾数 k生残尾数が 0 となるまでの日数 顕微注入卵による孵化塩分試験 受精卵への顕微注入 は,2015 年 12 月 3 日に五島庁舎で人工採卵された孵 化塩分試験と異なるバッチの受精卵を用いて行った。本 研究では顕微注入の成否を判断するため,GFP タンパ クの緑色蛍光を指標とした。顕微注入には,細胞に注入 されるとリボソーム内で GFP タンパク質が合成されて 緑色蛍光を発する mRNA-GFP を用いた。mRNA-GFP は,pCS2mt-GFP(Michael Klymkowsky 氏の好意によ り,Addgene から研究用として配布されているプラス ミド,#15681)から mMESSAGE mMACHINE SP6 Transcription Kit (Life Technologies Japan 社製)を用 いて合成し,150 ng/mL の濃度に調節した。次に,受精 卵への顕微注入は,2(w/v)アガロースゲルに付け た溝で受精卵を固定し,実体顕微鏡下でガラス針(GD-1または G-1, Narishige 社製)を取り付けたマイクロイ ンジェクター(IM-9B, Narishige 社製)を接続したマ イクロマニピュレーター(UMM-3FC, Narishige 社製) を操作し,1 細胞期の卵の胚盤内へ mRNA-GFP を油球 径の半分程度になるように注入した。およそ 200 粒の 受精卵に注入し,翌日(受精後 24 時間)に 136 粒の原 腸胚期の卵を得た。これらを 2 群に分け,塩分 35 と 40 psu の海水を入れた 500 mL 容器に 68 粒ずつ収容し, 上浦庁舎に輸送した。上浦庁舎到着時の受精卵は胚体形 成期の発生段階で,塩分 35 と 40 psu のそれぞれで 68 粒と 54 粒(計 122 粒)が生存していた。すべての卵は 実体顕微鏡下で観察し,白色に濁って水槽底面に沈んだ 卵または浮上していても発生が止まり白色に濁っている 卵を死卵とし,浮上して胚体を形成している卵を生残卵 とした。生残卵は塩分ごとに 6 穴マイクロプレートへ 1520 粒ずつ収容し,20°C で静置した。未注入卵は, 顕微注入卵と同様に原腸胚期の卵を塩分 35,または 40 psuに調節した海水を入れた 500 mL 容器に 48 粒ずつ 収容して上浦庁舎に輸送した。なお,未注入卵はすべて の受精卵が生存し,それらは塩分ごとに 6 穴マイクロ プレートへ 1416 粒ずつ収容し,20°C で 静置した。 mRNA-GFP を顕微注入した卵の一部は,受精 36 時間 後にデジタルカメラ(DS-Ri2, Nikon 社製)を接続した 蛍光実体顕微鏡(SMZ25, Nikon 社製)下で観察および 撮影を行った。なお,顕微注入卵と未注入卵ともに 1 塩分区あたり 3 回の繰り返しを行った。 統計処理 孵化率,正常孵化率および無給餌生残指数 (SAI)を塩分間および実験区間で比較するために,一 元配置の分散分析(one-way ANOVA)を行った後,有 意水準 5 で TukeyKramer の多重検定を行った。 結 果 ブリ受精卵の発生に伴う卵比重の変化 試験開始時の ブリ受精卵(孵化 30 時間前)は,胚体が卵黄球の約半 分を取り巻き,その翌日には胚体が卵黄のほぼ全周を取 り巻くにまで発生が進行した(孵化 10 時間前)。その 後,孵化まで外見上の大きな変化は観察されなかった (Fig. 1)。ブリ受精卵の卵比重と中立浮上塩分(NBS) は,発 生に 伴い 大き く上 昇した (Fig. 2)。卵比 重と NBS は , 孵 化 30 時 間 前 で そ れ ぞ れ 1.0217 g / cm3と 29.4 psu を,10 時間前でそれぞれ 1.0232 g/cm3と 31.3Fig. 2 Developmental changes in speciˆc gravity of yel-lowtail eggs. Closed and open circles depict neutral and positive buoyancy eggs, respectively. Solid lines are salinities at 25, 30, 35, and 40 psu. The dashed line is the salinity of the original sea-water (33.3 psu) in this study.
Fig. 3 Total hatching (a, c) and normal hatching rates (b, d) of yellowtail eggs at diŠerent salinities: a, b (tri-al 1); c, d (tri(tri-al 2). Symbols with diŠerent letters are signiˆcantly diŠerent at p<0.05 with TukeyKramer's multiple comparison test after one-way ANOVA.
Fig. 4 The survival rate of yellowtail larvae at diŠerent salinities. Yellowtail larvae were incubated in 6-well microplates at diŠerent salinities of 35 (black circle), 40 (white circle), and 45 (gray circle) psu. The num-ber of surviving larvae was observed daily under a microscope. psu を示し,海水の比重 1.0248 g/cm3(塩分 33.3 psu) よりも低い値を示した。一方,孵化 5 時間前にはそれ ぞれ 1.0251 g/cm3と 33.6 psu を示し,33.3 psu の環境 水中では表層から下層への受精卵の沈降が観察された。 さらに,孵化時(孵化 0 時間)の受精卵ではそれぞれ 1.0275 g/cm3および 36.6 psu を示し,顕著な沈降が観 察された。また,孵化 5 時間前および 0 時間の受精卵 を表層へ移動させるのに必要な塩分は,それぞれ 36.6 と 40.0 psu であった。 孵化塩分試験 2 回の孵化塩分試験では孵化の約 30 時間前に受精卵を各塩分へ移槽した。高塩分域に条件を 設定した 1 回目の試験では塩分が 50 psu 以上で孵化率 の低下が見られた( p<0.05, TukeyKramer's multiple comparison test with one-way ANOVA; F5,12=21.421,
p<0.0001, Fig. 3a)。低塩分域も設定した 2 回目の試験 では塩分 30 psu で有意に孵化率が低下した( p<0.05, TukeyKramer's multiple comparison test with one-way ANOVA; F4,10=9.112, p=0.0023, Fig. 3c)。なお,塩分
30 psu で管理した受精卵は,移槽直後に沈降が観察さ れた。塩分 35 から 40 psu までの正常孵化率は,2 回の 試 験 と も に 80 以 上 の 高 い 値 を 示 し た ( p < 0.05, TukeyKramer's multiple comparison test with one-way ANOVA, p<0.01, Fig. 3b, d)。
異なる塩分による孵化仔魚の生残率の変化と無給餌生 残指数(SAI, survival activity index) 孵化率の高い 塩分環境が明らかになっても,孵化仔魚の生残率が著し く低下するなら小型容器孵化管理法の開発にはならな い。そこで,孵化仔魚の塩分に対する影響を検討するた め,塩分 35, 40 および 45 psu で得られた正常孵化仔魚 の生残率の推移を観察し,1 日に 1 回測定した生残数お よび死亡数をもとに無給餌生残指数(SAI)を算出した。 塩分 35 と 40 psu の生残率は孵化後 7 日齢で大きく減少 し,8 日齢に全滅した。一方,塩分 45 の生残率は孵化 後 3 日齢で大きく減少し,4 日齢に全滅した(Fig. 4)。 塩 分 35, 40 お よ び 45 psu の SAI は , そ れ ぞ れ 19.9 ± 1.2(平均値±標準誤差),19.0±2.2 および 4.2±0.1 で あった。SAI は塩分 45 psu で最も低かった( p<0.05, TukeyKramer's multiple comparison test with one-way ANOVA; F2,6=36.800, p=0.0004)。
顕微注入卵による孵化塩分試験 mRNA-GFP を顕微
Fig. 5 Success of microinjection was ascertained from observation of GFP ‰uorescence in embryos. (a) and (b) show a bright and dark ˆeld with GFP ‰uores-cence, respectively. Embryos successfully microinject-ed with GFP-mRNA depictmicroinject-ed by arrowheads.
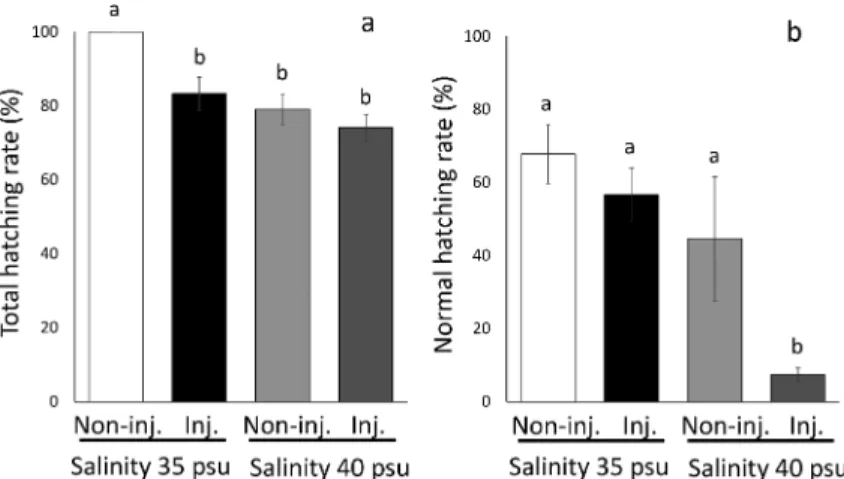
Fig. 6 Total hatching (a) and normal hatching rate (b) of microinjected (Inj.) and non-injected eggs (Non-inj.) at 35 and 40 psu. Symbols with diŠerent letters are signiˆcantly diŠerent at p<0.05 with TukeyKramer's multiple comparison test after one-way ANOVA.
形成期)に GFP の発現を観察し,75 の割合で導入に 成功した(Fig. 5)。孵化率は塩分 35 と 40 psu の環境 で顕微注入卵および未注入卵ともに 70 以上を示した が(Fig. 6a),塩分 35 psu の未注入卵で他の試験区よ り高かった( p<0.05, TukeyKramer's multiple com-parison test with one-way ANOVA; F3,8=10.026, p=
0.0044)。一方,正常孵化率は塩分 40 psu で低下する傾 向が見られ,特に顕微注入卵は 7.4±1.9 を示し,他
の試験区よりも低かった( p<0.05, TukeyKramer's
multiple comparison test with one-way ANOVA; F3,8=
6,706, p=0.0142, Fig. 6b)。塩分 35 psu で管理した顕 微注入卵の正常孵化率は 56.7±7.3 を示し,塩分 40 psuで管理した未注入卵の 44.6±17.0 より高い傾向 があった(Fig. 6b)。 考 察 孵化直前のブリの受精卵は,シマアジと同様に,14)多 くの海産魚の卵比重や中立浮上塩分(NBS)913)と比較 して高く,海水の比重を超える値を示した。また,ブリ 卵の発生に伴う沈降現象はこれまでの報告と同様に,8) 卵比重が環境水中の比重を上回ることにより生じること が確認された。このように,発生に伴い卵比重が増大す ると卵が沈降し,水槽や容器の底部に接触し自重の負荷 によって卵発生が停止する8,14)のはシマアジやブリに見 られる特有の現象と考えられる。この点において,大型 水槽だけでなく小型容器でも受精後から孵化まで比較的 安定した条件で管理できる他魚種1618)とは異なる。ま た,ブリは通気の強度を変えるタイミングや調節に習熟 が必要であるため,孵化管理が難しい魚種といえる。こ れまで著者らによって複数回にわたる予備試験で示され た数程度の低い孵化率は,通気強度を変えるタイミン グや調節が厳密に行われなかったことに起因したと考え られた。本研究ではこのように孵化管理の難しい魚種で あるブリにおいて,環境水中の塩分を調節することによ り,容易に小型容器で孵化管理できる方法を開発した。 以下により詳細な説明を行う。 孵化塩分試験において,2 回の試験ともに塩分 35 か ら 40 psu までは正常孵化率が高く,2 回目の塩分 37.5 psu で最も高い正常孵化率を示した。この範囲の塩分は ブリ受精卵に対して環境ストレス要因にならないことが 考えられた。また,天然のブリの産卵場は,比較的高塩 分環境とされている。19)このことからも,ブリ受精卵の 孵化管理は高塩分環境が適していると推測された。一 方,塩分 30 psu で孵化管理した受精卵は孵化のおよそ 24 時間前に容器底面に沈降し,その孵化率は顕著に低 かったのに対し,塩分 35 psu の環境水で孵化管理した 受精卵は孵化 5 時間前から沈降が観察されたが,孵化 率および正常孵化率のどちらも高い値を示した。したが って,ブリ受精卵の孵化率は沈降後の時間の長さに依存 して低下し,たとえ水槽底に沈降してもおよそ 5 時間
以内であれば,正常孵化率の低下は生じないと考えられ た。 塩分 35 と 40 psu の孵化後の生残率は類似した変化を 示したが,塩分 45 psu は孵化後 3 日齢から急激な生残 率の低下を示した。本研究ではブリの孵化仔魚が孵化後 3 日齢でほぼ卵黄を吸収し,それと同時に開口や目の色 素沈着が観察された。多くの魚種において,孵化直後の 仔魚は様々な環境ストレス要因に対して,高い耐性を示 すことが知られている。2025)孵化直後の仔魚は口および 鰓蓋が未発達であるため,外部環境の影響を受けにくい と考えられるが,卵黄吸収後の仔魚は開口し,鰓が機能 し始めるために,環境ストレスに弱くなると考えられて いる。25)本研究においても孵化後 3 日齢に塩分 45 psu で顕著な生残率の低下が観察されたため,ブリの仔魚に は塩分 45 psu が大きな環境ストレスになり,生残率の 顕著な低下が生じたと考えられた。一方,塩分 40 psu は 35 psu と 同 様 の 生 残 率 の 変 化 や 無 給 餌 生 残 指 数 (SAI)を示し,それらは虫明ら26,27)の報告と類似した 値を示した。したがって,正常孵化率と同様に,この範 囲の塩分は,ブリ孵化仔魚に対して環境ストレスや生存 低下の大きな要因にならないと考えられた。 mRNA-GFP を注入した受精卵は,塩分 35 psu で孵 化管理することにより 56.7 の正常孵化率を示した。 これまでの予備試験で得られた数程度の低い正常孵化 率と比較すると,56.7 は高い正常孵化率と言える。 これは,これまで問題となってきた小型容器孵化管理が 大幅に改善されたことを意味し,ブリのゲノム編集を含 めた顕微注入に関する研究の基礎が整備されたと考えて いる。しかしながら,塩分 40 psu での正常孵化率は, 塩分 35 psu でのそれより有意に低下した(結果顕微 注入卵による孵化塩分試験を参照)。これは未注入卵に おいても類似した傾向が観察された。孵化塩分試験での 正常孵化率は,塩分 35 から 40 psu の間に有意な違いは 認められなかったが(結果孵化塩分試験を参照),顕 微注入卵による孵化塩分試験では顕微注入卵だけでな く,未注入卵においても塩分 40 psu で有意な正常孵化 率の低下が観察された。孵化塩分試験では,受精後およ そ 36 時間の胚体形成期に異なる塩分に移槽したが(材 料と方法孵化塩分試験を参照),顕微注入による孵化 塩分試験では受精後およそ 24 時間の原腸胚期で移槽し た(材料と方法顕微注入による孵化塩分試験を参照)。 これまでに,ブリ受精卵は桑実胚期以前は極めてハンド リングに弱いことが経験的に知られてきたが,8)その他 の発生段階における知見はない。一般に,魚類の原腸胚 期は様々な器官形成が開始され,活発に細胞分裂が行わ れる時期であり,いくつかの魚種ではこの時期に環境ス トレスが加えられると細胞分裂の停止,体節数の減少や 体節の萎縮が生じるとされている。28)塩分 40 psu に移 槽した顕微注入卵は孵化率が高いのに対して正常孵化率 が著しく低下し,それらの正常孵化を示さなかった奇形 個体は主に矮小個体だった。これらのことから,本研究 では原腸胚期と胚体形成期では環境ストレスに対する感 受性が異なり,特に,原腸胚期に高塩分のような環境ス トレスを加えたことで奇形個体が顕著に増加したことが 考えられた。この点に関しては,今後,異なる発生段階 の受精卵を用いて,高塩分への適切な移槽時期を検証す る必要がある。 本研究により,塩分を調節することで小型容器を用い てブリ顕微注入卵の孵化管理ができるようになった。し かし,ゲノム編集魚の飼育は少ない仔魚数で飼育を開始 することが予想される。特に,海産魚の種苗生産では小 型水槽になるほど飼育が困難になる。また,ブリでは種 苗生産初期の油膜除去に失敗すると,ほとんどの個体の 鰾が開腔せず,未成熟魚での脊椎骨上彎症の原因とな る。7)このように,ブリにおいてゲノム編集による系統 作出を達成するには,小型水槽においても高い鰾の開腔 率を示すような飼育方法の確立が不可欠になるだろう。 謝 辞 本研究は農林水産技術会議の平成 27 年度「ゲノム情 報を利用したブリ類の短期育種技術の開発」により実施 したものである。実験を実施するにあたり,水産総合研 究センター西海区水産研究所五島庁舎の職員各位に受精 卵提供に関して多大なるご援助を頂いた。ここに記して 関係各位に感謝の意を表す。また,実験を遂行しまとめ るにあたり,有意義なご助言を頂いた水産総合研究セン ター増養殖研究所上浦庁舎の職員各位に厚くお礼申し上 げる。 文 献
1) Ozaki A, Fuji K, Yoshida K, Kubota S, Kai W, Suzuki J, Akita K, Koyama T, Aoki J, Nakagawa M, Hotta T, Tsuzaki T, Araki K, Okamoto N, Sakamoto T. Genetic breeding for parasite resistance (Benedenia seriolae) using genomics information in yellowtail (Seriola quinqueradia-ta). Proceedings of the 40th U.S.Japan aquaculture panel symposium, 2013; 314.
2) Yoshida K, Ozaki A, Nakagawa M, Hotta T, Aoki J, Koyama T, Araki K, Okamoto N, Sakamoto T, Tsuzaki T. Characteristic evaluation method for Benedenia dis-ease in yellowtail (Seriola quinqueradiata). Proceedings of the 40th U.S.Japan aquaculture panel symposium, 2013; 9699.
3) Ozaki A, Yoshida K, Fuji K, Kubota S, Kai W, Aoki J, Kawabata Y, Suzuki J, Akita K, Koyama T, Nakagawa M, Hotta T, Tsuzaki T, Okamoto N, Araki K, Sakamoto T. PLoS ONE 2013; 8: e64987.
4) Nakayama T, Fish MB, Fisher M, Oomen-Hajagos J, Thomsen GH, Grainger RM. Simple and e‹cient CRISPR/Cas9-mediated targeted mutagenesis in Xeno-pus tropicalis. Genesis2013;51: 835843.
5) Edvardsen RB, Leininger S, Kleppe L, Skaftnesmo KO, Wargelius A. Targeted mutagenesis in Atlantic salmon (Salmo salar L.)using the CRISPR/Cas9 system induces complete knockout individuals in the F0 generation. PLoS ONE 2014; 9: e108622. 6) 中田 久,中尾貴尋,荒川敏久,松山倫也.ブリの成 熟排卵誘導における HCG 投与時の卵径と排卵時間, 卵 量 お よ び 卵 質 と の 関 係 . 日 本 水 産 学 会 誌 2005;71: 942946. 7) 水産総合研究センター.「栽培漁業技術シリーズ No. 12 ブリの種苗生産技術開発」日昇印刷,東京.2006; 183. 8) 日本栽培漁業協会.「栽培漁業技術シリーズNo. 5 ブリの 親魚養成技術開発」(日本栽培漁業協会企画調査室編)日 昇印刷,東京.1999; 172. 9) 牧野 直,松丸 豊,田中 克.スズキ卵の発生に及ぼ す水温と塩分の影響.水産増殖 2003;51: 5564. 10) Coombs SH. A density-gradient column for determining
the speciˆc gravity of ˆsh eggs, with particular reference to eggs of the mackerel Scomber scombrus. Mar. Biol. 1981; 63: 101106.
11) Nissling A, Kryvi H, Vallin L. Variation in egg buoyancy of Baltic cod Gadus morhua and its implications for egg survival in prevailing conditions in the Baltic Sea. Mar. Ecol. Prog. Ser. 1994;110: 6774.
12) Nissling A, Westin L, Hjerne O. Reproductive success in relation to salinity for three ‰atˆsh species, dab (Limanda limanda), plaice (Pleuronectes platessa), and ‰ounder (Pleuronectes ‰esus), in the brackish water Baltic Sea. ICES J. Mar. Sci. 2002;59: 93108.
13) 近藤茂則,吉岡 基,柏木正章.シロギス卵の卵質評価 に関する研究.日本水産学会誌 2001;67: 801806. 14) 水産総合研究センター.「栽培漁業技術シリーズNo. 11 シマアジ親魚養成に関する技術開発成果」日昇印刷,東 京.2005; 161. 15) 新間脩子,辻ヶ堂 諦.カサゴ親魚の生化学的性状と仔 魚の活力について.養殖研究所研究報告 1981;2: 1120. 16) Kaji T, Tanaka M, Oka M, Takeuchi H, Ohsumi S, Teruya K, Hirokawa J. Growth and morphological de-velopment of laboratory-reared yellowˆn tuna Thunnus albacareslarvae and early juveniles, with special emphasis
of the digestive system. Fish. Sci. 1999; 65: 700707. 17) Kaji T, Kodama M, Arai H, Tanaka M, Tagawa M.
Prevention of surface death of marine ˆsh larvae by the addition of egg white into rearing water. Aquaculture 2003;224: 313322.
18) Tagawa M, Kaji T, Kinoshita M, Tanaka M. EŠect of stocking density and addition of proteins on larval sur-vival in Japanese ‰ounder, Paralichthys olivaceus. Aqua-culture 2004; 230: 517525.
19) 水産庁調査研究部.ブリ漁業資源.「日本近海主要漁業 資源」(水産庁調査研究部編),東京.1973; 165174. 20) 青海忠久.イシダイ卵仔稚に対する有機リン系殺虫剤の
急性毒性.日本水産学会誌 1982;48: 599603.
21) Hiroi J, Sakakura Y, Tagawa M, Seikai T, Tanaka M. De-velopmental changes in low-salinity tolerance and responses of prolactin, cortisol and thyroid hormones to low-salinity environment in larvae and juveniles of Japanese ‰ounder, Paralichthys olivaceus. Zool. Sci. 1997; 14: 987992.
22) Shikano T, Fujio Y. Successful propagation in seawater of the guppy Poecilia reticulate with reference to high salinity tolerance at birth. Fish. Sci. 1997; 63: 573575. 23) Shikano T, Fujio Y. Decline in salinity tolerance during
larval stage of chum salmon Oncorhynchus keta. Fish. Sci. 1999; 65: 661662. 24 ) 石 橋 泰 典 , 小 澤 勝 , 平 田 八 郎 , 熊 井 英 水 . マ ダ イ Pagrus major 仔稚魚の発育に伴う各種環境ストレス耐性 の変化.日本水産学会誌 2003;69: 3643. 25 ) 嶋 田 幸 典 , 鹿 野 隆 人 , 青 海 忠 久 . ヒ ラ メParalichthys olivaceus の発育初期における環境ストレスに対する耐性 の変化.日本水産学会誌 2006;72: 169173. 26 ) 虫 明 敬 一 , 新 井 茂 , 松 本 淳 , 新 間 脩 子 , 長 谷 川 泉.モイストペレットで飼育した養殖ブリ 2 年魚の人工 採卵.日本水産学会誌 1993;59: 17211726. 27) 虫明敬一,河野一利,Verakunpiriya W,渡邉 武,長 谷川 泉.市販ソフトドライペレットを給餌したブリの 採卵結果.日本水産学会誌 1995;61: 540546. 28) 藪 健史.魚類胚におけるストレス誘導性アポトーシス の 分 子 機 構 に 関 す る 研 究 . 日 本 水 産 学 会 誌 2006;72: 819822.