アワビ消化管由来の海産紅藻スサビノリ細胞壁分解酵素祖
の調製
誌名
誌名
水産増殖 = The aquiculture
ISSN
ISSN
03714217
著者
著者
大塚, 周二
福田, 覚
柴田, 大輔
ほか2名,
巻/号
巻/号
54巻1号
掲載ページ
掲載ページ
p. 83-88
発行年月
発行年月
2006年3月
農林水産省 農林水産技術会議事務局筑波産学連携支援センターTsukuba Business-Academia Cooperation Support Center, Agriculture, Forestry and Fisheries Research Council Secretariat
アワビ消化管由来の海産紅藻スサビノリ細胞壁分解酵素の調製
大塚周二
1・福田 覚
1, 2・柴田大輔
1・嵯峨直恆
1・尾島孝男
1Preparation of Abalone Enzymes Capable of Cell Wall Degradation of a
Marine Red Alga Porphyra yezoensis (Bangiales, Rhodophyta)
Shuuji O
OTSUKA1, Satoru F
UKUDA1, 2, Daisuke S
HIBATA1,
Naotsune S
AGA1and Takao O
JIMA1Abstract: We extracted a xylanase and a mannanase from Ezo abalone Haliotis discus hannai,
and examined the enzyme activity and the ability for cell wall degradation of Porphyra yezoensis. Abalone xylanase was partially purified from the crude enzyme prepared from the digestive fluid by column chromatography on a TOYOPEARL CM-650M. Abalone mannanase was successfully purified from the crude enzyme by column chromatographies on TOYOPEARL CM-650M, hydroxyapatite, and TOYOPEARL HW-50F. The gametophytic thalli of P. yezoensis could not be degraded by the xylanase alone. The purified mannanase could degrade the thalli into cell masses consisting of 10−20 cells; however, it produced practically no protoplasts from the thalli. When the thalli were incubated both with the xylanase and the purified mannanase, some protoplasts could be produced from the thalli. The two enzymes may be useful for the studies on cell engineering and asexual seedling production of this alga.
Key words: Haliotis discus hannai; Porphyra yezoensis; Mannanase; Xylanase
2005年11月21日受付:2005年12月26日受理.
1 北 海 道 大 学 大 学 院 水 産 科 学 研 究 院(Graduate School of Fisheries Sciences, Hokkaido University, Minato 3-1-1, Hakodate, Hokkaido 041-8611, Japan).
2 共和コンクリート工業㈱アルガテックKyowa(Algatech Kyowa, Kyowa Concrete Industry Co., Ltd., Benten 24-13, Hakodate, Hokkaido 040-0054, Japan).
海産紅藻スサビノリ Porphyra yezoensis Ueda は, 紅 色 植 物 門 Rhodophyta, 紅 藻 綱 Rhodophyceae, 原始紅藻亜綱 Protoflorideophycidae,ウシケノリ目 Bangiales,ウ シ ケ ノ リ 科 Bangiaceae, ア マ ノ リ 属 Porphyraに分類される海産の多細胞性藻類である。ま た,日本の海苔養殖業を支える最も代表的な種であり, 水産業においても最も重要な地位を確立している水産 物の一つに数えられる(植木ら 2005)。スサビノリに は,粘質多糖のポルフィランが多量に含まれており, このポルフィランは抗腫瘍活性(Noda et al. 1989)や 血清コレステロールを低下させる効果が知られている (Ren et al. 1994)。また,スサビノリのポルフィラン を分解して得られるオリゴ糖にも同様の作用があるこ とが報告されている(大住ら 1998)。このようにスサ ビノリは,養殖種としての利用以外に機能性食品や医 薬品の素材としての応用研究も行われてきている。 以上のことから,スサビノリでは,より良い養殖 品種の作出や様々な生理作用および諸現象を解明する ために,プロトプラストの単離技術,プロトプラスト の細胞工学的な利用方法,そしてモデル生物としての 基盤技術の研究開発が進められている(藤田 1991; 嵯 峨・田畑 2001)。スサビノリのプロトプラストの研究 においては,アワビの消化管由来の粗酵素(以降アワ ビ粗酵素)が利用される他(Saga et al. 1986),近年 では,アマノリ属の細胞壁構成成分であるβ-1,4-マン ナン,β-1,3-キシランならびにポルフィランの 3 種類 の多糖を分解する酵素,すなわち,β-1,4-マンナナー ゼ,β-1,3-キシラナーゼおよびポルフィラン分解酵素 を産生する細菌由来の粗酵素(以降細菌粗酵素)を 混合することで,スサビノリから大量のプロトプラ ストを単離することに成功している(荒木 1997)。し かしながら,アワビ粗酵素では,細菌粗酵素よりプロ
84 大塚・福田・柴田・嵯峨・尾島 トプラストの単離数が少ない,細菌粗酵素ではプロト プラストの再生において不定形のカルス様藻体を形成 するといった問題点も挙げられている(藤田 1991; 荒 木 1997)。なぜこのようなことが起こるのかについて は良く分かっていないが,どちらも粗酵素の段階で 利用していることから,原因を特定しにくくなって いるものと考えられる。したがって,細胞壁構成成分 を分解する酵素,あるいは,分解に関与する酵素を精 製し,その酵素活性を 1 つ 1 つ確認することが必要で ある。また,実際に市販されている細菌粗酵素でプロ トプラストの再生藻体を利用した選抜育種が行われて きたが,粗酵素が高価であるため一貫した利用が困難 であり,利便性が悪いことが指摘されている(増田ら 1997)。以上のことから,スサビノリにおけるプロト プラストの単離技術の開発については,未だ検討の余 地がある。 そこで,本研究では,エゾアワビ Haliotis discus hannaiの消化管由来の粗酵素からスサビノリ葉状の 配偶体の細胞壁構成成分であるキシランとマンナンを 分解する酵素を抽出し,それぞれの酵素活性と細胞壁 分解能を調べることを目的とした。 材料および方法 エゾアワビの消化管からの粗酵素の抽出 エゾアワビは,北海道函館市内の市場で購入したも のを試料として用いた。殻長約 6 × 8 cm のエゾアワ ビ10個体の消化管から 1 ml のシリンジを用いて吸引 し,消化液を採取した。次に得られた消化液は500 ml の10 mM リン酸ナトリウム緩衝液(pH 7.0)中で,4℃, 6時間透析した。次に100,000×g で30分間遠心分離し た後,得られた上清をエゾアワビ粗酵素とした。 エゾアワビ粗酵素中のマンナナーゼおよびキシラナー ゼの活性測定 マンナナーゼの活性測定では,ローカストビーン ガム(Locust bean gum from Ceratonia siliqua seeds, SIGMA)を 5 mg/ml,キシラナーゼの活性測定では, キシラン(Xylan from Oat Spelts,Wako)を20 mg/ml の濃度になるよう10 mM リン酸ナトリウム緩衝液 (pH 7.0)で溶解し,基質溶液とした。それぞれの基 質溶液 1 ml にエゾアワビ粗酵素液を0.5μl添加し,マ ンナナーゼの活性測定では30℃で30分間,キシラナー ゼの活性測定では30℃で120分間反応させた。その後, 反応液を0.1 ml 採取し,100℃に加熱してある0.5 ml の0.1% SDS に加えることにより反応を停止した。基 質の分解による生成された還元糖量は Park-Johnson 法(Park and Johnson 1949)により定量した。酵素の
1単位(unit)は,1 分間に 1μmolのガラクトースに 相当する還元糖を生成する酵素量と定めた。 マンナナーゼとキシラナーゼの活性画分の溶出 上記の方法で得られたエゾアワビ粗酵素液25 ml を 10 mMリン酸ナトリウム緩衝液(pH 7.0)で平衡化し た TOYOPEARL CM-650M カラム(25 mm×265 mm) に供し,吸着したタンパクを 0−0.3 M NaCl の直線濃 度勾配によって溶出した。溶出液は 1 画分あたり 5 ml ずつ分取した。 マンナナーゼの精製 TOYOPEARL CM-650Mカラムで溶出したマンナ ナーゼ活性画分(Fraction 59-63)を集め,10 mM リ ン酸カリウム緩衝液(pH 7.0)で透析した後,同溶 液に平衡化してあるハイドロキシアパタイトカラム (15 mm×200 mm)に供した。吸着したタンパクは 0.01−0.3 M リン酸カリウム緩衝液(pH 7.0)の直線 濃度勾配によって溶出した。溶出液は 1 画分あたり 5 mlずつ分取した。このクロマトグラフィーで得られ
た高活性画分(Fraction 57-63)を Apollo Centrifugal Concentrator 20 ml(Orbital Biosciences)を用いて約 3 mlに濃縮し,この濃縮物を TOYOPEARL HW-50F カラム(20 mm×900 mm)でゲル濾過した。
精製されたマンナナーゼの活性測定
アガロースを 2 mg/ml,カルボキシメチルセル ロースナトリウム(CMC)(Carboxymethyl cellulose sodium salt, medium viscosity,ICN Biomedicals), グルコマンナン(Glucomannan from Konjac Tuber, Wako),ローカストビーンガム,β -1,4-マンナン(1,4-β-D-Mannan, Megazyme)を 5 mg/ml の濃度になる よう10 mM リン酸ナトリウム緩衝液(pH 7.0)で溶 解し,基質溶液とした。それぞれの基質溶液 1 ml に 精製されたマンナナーゼを0.5 ml 添加し,30℃で反 応させた。反応開始後 5,10,20,30分後にそれぞれ 反応液を0.1 ml 採取し,100℃に加熱してある0.5 ml の0.1% SDS に加えることにより反応を停止した。基 質の分解により生成された還元糖量は Park-Johnson 法(Park and Johnson 1949)により定量した。酵素の 1単位(unit)は,1 分間に 1μmolのガラクトースに 相当する還元糖を生成する酵素量と定めた。
スサビノリの培養条件
本実験では,スサビノリの TU-1 株(Kuwano et al. 1996)の単藻培養株の葉状の配偶体を用いた。本材料 は,15℃,10時間明期(光量80μE/m2/s;白色蛍光灯)・ 14時間暗期の短日条件の下,ESL 培地(Kitade et al.
2002)が入った容量2,000 ml の簡易式培養容器(TU-2, 藤森工業)中で通気(ポアサイズ0.2μmのフィルター でろ過滅菌した空気,250 ml/min)し,環流浮遊状態 で培養された。培地は 7 日ごとに交換した。 スサビノリのエゾアワビ酵素による分解 葉長約15 mm の材料を 1 ml のエゾアワビの酵素液 (5 unit の酵素液,0.7 M ソルビトールを含む人工海 水)に加え,22℃で 4 日間インキュベートした。人工 海水は Sealife(マリンテック)を用いた。酵素液は, マンナナーゼとキシラナーゼ活性のあった画分,キ シラナーゼ活性があった画分および精製したマンナ ナーゼをそれぞれ用いた。この間の材料の分解の様 子は正立顕微鏡および倒立顕微鏡で観察した。細胞壁 の有無は,カルコフローホワイト(Calcofluor White M2R, SIGMA)による藻体の細胞壁を青く染色する方 法(Maeda and Ishida 1967)を用いて確認をした。
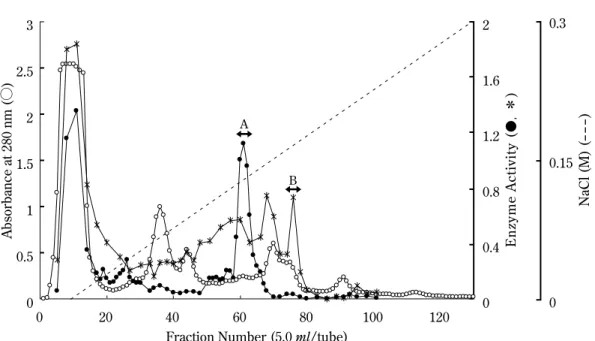
結 果 マンナナーゼとキシラナーゼの活性画分の溶出 エゾアワビ10個体から約20 ml の粗酵素液が得られ た。粗酵素液を TOYOPEARL CM-650M カラムで分 画した結果,マンナナーゼおよびキシラナーゼ活性の あるパス画分が得られた(Fig. 1)。また,マンナナー ゼおよびキシラナーゼ活性のある吸着画分(Fraction 59-63)が得られ,これを画分 A とした(Fig. 1)。また, キシラナーゼ活性のある吸着画分(Fraction 75-77) が得られ,これを画分 B とした。 マンナナーゼの精製 画分 A をハイドロキシアパタイトカラムに供し, さらに TOYOPEARL HW-50F カラムでゲル濾過を行 うことによりマンナナーゼを精製した。その結果, Fraction 75-77にマンナナーゼ活性を示す画分が溶 出 し た。 こ の 画 分 は,SDS-PAGE の 結 果, 分 子 量 約39,000の単一バンドを示したため,これを精製マ ンナナーゼとした。精製マンナナーゼは,比活性が 11.48 U/mg,アワビ粗酵素からの収率は1.59%であ り,ローカストビーンガム,グルコマンナン,β -1,4-マンナンを良く分解するが,アガロース,キシラン, CMCを全く分解しないことからβ-1,4-マンナナーゼ と同定された(Fig. 2)。
Fig. 1. TOYOPEARL CM-650M column chromatography of abalone crude enzyme. ○, absorbance at 280 nm; ●, man-nanase activity; *, xylanase activity. A, fraction A consisting of fraction 59-63; B, fraction B consisting of fraction 75-77.
Fig. 2. Substrate specificity of the purified abalone
man-nanase. Abalone mannanase activity was assayed with locust bean gum (●), β-1,4-mannan (▲), glucomannan (■), agarose (○), xylan (+) and CMC (×).
��� ���� ��� ���� ��� ���� � �������������������� �� �� ��� ��� ��� ��� �� ���������� ��������������� ����������� β����������� ������� ����� ��� 3 2.5 2 1.5 1 0.5 0 Absorbance at 280 nm (○ ) 0 20 40 60 80 100 120
Fraction Number (5.0 ml/tube)
2 1.6 1.2 0.8 0.4 0 0.3 0.15 0 NaCl (M) (- --) E nz ym e A ct iv it y ( ● , * ) � �
86 大塚・福田・柴田・嵯峨・尾島 エゾアワビ酵素のスサビノリTU-1株の葉状の配偶体 に対する分解能 アワビ粗酵素,マンナナーゼおよびキシラナーゼ活 性のあった画分 A,キシラナーゼ活性のあった画分 B ならびに精製マンナナーゼを葉長約15 mm の材料に対 して酵素処理を行った。エゾアワビ粗酵素については, 酵素処理の結果,プロトプラスト化されることが確認 された(Fig. 3B, C)。画分 A については,4 日間の酵 素処理をした結果,藻体の崩壊が起こり,若干のプロ トプラスト化が確認された(Fig. 3D, E)。画分 B につ いては,4 日間の酵素処理をした結果,藻体の分解の 様子は全く変化せず,プロトプラスト化も確認されな かった(データは示さない)。精製マンナナーゼにつ いては,酵素処理の結果,10∼20 細胞から成る細胞 塊が複数生じることが確認された(Fig. 3F, G)。 考 察 本研究により,エゾアワビ粗酵素,画分 A および精 製マンナナーゼの酵素処理の結果からスサビノリのプ Fig. 3. Degradation of thalli from Porphyra yezoensis with abalone enzymes. A, thalli of P. yezoensis. B and C, protoplasts
from thalli of P. yezoensis degraded by abalone crude enzyme. D and E, thalli of P. yezoensis degraded by the fraction A includ-ing mannanase and xylanase. F and G, thalli of P. yezoensis degraded by the purified mannanase. C, E and G, stained with Calcoflour white.
(A)
(B) (C)
(D) (E)
ロトプラスト化を促す酵素がエゾアワビ消化管にも存 在することが確認された。特に,精製マンナナーゼは, 10∼20細胞の細胞塊を生ずる酵素活性が確認されたこ とから,スサビノリのプロトプラスト化に強く関与し ている可能性が示唆された。本材料と同じウシケノリ 科に属するウシケノリ Bangia atropurpurea ではマン ナナーゼ,キシラナーゼおよびポルフィラン分解能 を有するアガラーゼを産生する細菌の粗酵素でプロト プラスト化に成功している。ウシケノリでは,マンナ ナーゼ活性を有する粗酵素のみで酵素処理をしたが, 細胞壁はほとんど分解されなかったと報告されている (Araki et al. 1994)。このことから,酵素処理の条件, 酵素の由来および種の違い等があり,一概には言えな いが,今後,精製マンナナーゼによる種々のプロトプ ラストの単離条件を検討することでスサビノリにおけ るプロトプラスト化に関する新たな知見が得られるか もしれない。画分 A,画分 B および精製マンナナーゼ の酵素処理の結果から,キシラナーゼと精製マンナ ナーゼの組合せによるスサビノリ葉状の配偶体から のプロトプラストの単離技術の発展が期待される。ま た,本実験で得られた精製マンナナーゼのように純化 された酵素の応用として,例えば,精製したマンナナー ゼやキシラナーゼのみで,スサビノリのβ-1,4-マンナ ンとβ-1,3-キシランを分解することにより,効率的に ポルフィランを抽出することに利用できるかもしれな い。また,スサビノリのプロトプラストの単離には, パパインによる前処理が行われてきた(藤田 1991)。 しかしながら,本実験方法では,パパイン処理を行っ ていないが,エゾアワビ粗酵素,画分 A でプロトプラ スト化が見られた。このことから,実験方法が従来の プロトプラスト単離法より簡便になる可能性がある。 また,種々のスサビノリの品種判別に用いる簡便で純 度の高い DNA 抽出にプロトプラスト化した後,DNA を抽出する方法が提案されており(岩渕 2003),将来 的にスサビノリの品種判別の技術向上への貢献が期待 できる。 スサビノリは単相(n)の葉状の配偶体と複相(2n) の糸状の胞子体との両世代が交代する主生活環と,単 胞子(n)を介して葉状の配偶体世代を繰り返す副生 活環を持つ。現在の海苔養殖では,糸状の胞子体から 殻胞子を放出させ採苗し,葉状の配偶体,すなわち海 苔を生産している(中山 2005)。一方,副生活環を利 用した,すなわち単胞子をクローン種苗(単胞子を 1 つ 1 つの種として考える)として利用したり,単胞子 を実験材料として大量調製するために,単胞子の放出 制御(Kitade et al. 1999)や単胞子の誘導(Mizuta et al. 2003)するといった手法が開発されてきている。 しかし,単胞子の放出制御は,海苔養殖に利用できる ほどの大量の単胞子を得ることには不向きであり,単 胞子の誘導では,単胞子を誘導するのに時間を要する ので即効性が期待できない。一方,プロトプラストの 単離技術の利点は,比較的簡便に,早く大量に調製で きることであるが,再生率に難がある。したがって, 再生率の問題が克服できれば上記のように最適な実 験材料系になるとともにクローン種苗調製としても価 値ある技術となると考えられる。そこで,今後は本研 究で得られたエゾアワビ粗酵素とキシラナーゼ・精製 マンナナーゼの混合物で単離されたプロトプラストの 収量および再生率を詳細に比較・検討することが重要 な課題となる。また,キシラナーゼ活性が測定された 画分 B やマンナナーゼ活性およびキシラナーゼ活性 が測定されたパス画分より,各酵素を精製し,スサビ ノリに対する酵素活性を検討することも進めていきた い。 要 約
我々はエゾアワビ Haliotis discus hannai からキシラ ナーゼとマンナナーゼを抽出し,酵素活性とスサビノ リ Porphyra yezoensis の細胞壁分解能を調べた。キシラ ナーゼは,エゾアワビの消化液より調製した粗酵素を TOYOPEARL CM-650Mカラムで分画することによっ て得た。マンナナーゼは,同粗酵素を TOYOPEARL CM-650Mカラムで分画した後,ハイドロキシアパタ イトカラムおよび TOYOPEARL HW-50F カラムで精 製することによって得た。スサビノリ葉状の配偶体は, キシラナーゼ単独では分解されなかった。精製マンナ ナーゼは,同葉状の配偶体を10∼20細胞からなる細胞 塊に分解したが,プロトプラストを生成しなかった。 同葉状の配偶体をキシラナーゼ・精製マンナナーゼの 混合物とインキュベートすると,少量のプロトプラス トを生成した。本研究で調製された 2 種類の酵素は, スサビノリの細胞工学的研究やクローン種苗の研究に 利用できるかもしれない。 謝 辞 本研究の一部は,文部科学省 COE プログラム「海 洋生命統御による革新的食糧生産」の助成により遂行 されたものであり,ここに記してお礼申し上げる。 文 献 荒木利芳(1997)プロトプラストの単離技術.有用海藻の バイオテクノロジー(能登谷正浩編),恒星社厚生閣, 東京,pp. 62-72.
88 大塚・福田・柴田・嵯峨・尾島
Araki, T., M. Hayakawa, Y. Tamaru, K. Yoshimatsu and T. Morishita (1994) Isolation and regeneration of haploid protoplasts from Bangia atropurpurea (Rhodophyta) with marine bacterial enzymes. J. Phycol., 30, 1040-1046. 藤田雄二(1991)海洋植物の細胞操作.ラボマニュアル−
マリンバイオテクノロジー(嵯峨直恆・松永 是編), 裳華房,東京,pp. 69-85.
岩渕光伸(2003)DNA を調べてノリ養殖品種を識別する. 海苔と海藻,66,7-13.
Kitade, Y., S. Fukuda, M. Nakajima, T. Watanabe and N. Saga (2002) Isolation of a cDNA encoding a homologue of actin from Porphyra yezoensis (Rhodophyta). J. Appl. Phycol., 14, 135-141.
Kitade, Y., S. Fukuda and N. Saga (1999) Preliminary study on early development of monospore of Porphyra yezoensis (Bangiales, Rhodophyta). Fish Genet. Breed. Sci., 28, 27-34.
Kuwano, K., Y. Aruga and N. Saga (1996) Cryopreservation of clonal gametophytic thalli of Porphyra (Rhodophyta).
Plant Sci., 116, 117-124.
Maeda, H. and N. Ishida (1967) Specificity of binding of hexo-pyranosyl polysaccharides with a fluorescent brightener.
J. Biochem., 62, 276-278.
増田恵一・谷田圭亮・水田 章(1997)プロトプラスト再 生藻体の選抜育種.有用海藻のバイオテクノロジー(能 登谷正浩編),恒星社厚生閣,東京,pp. 83-89. Mizuta, H., H. Yasui and N. Saga (2003) A simple method to
mass produce monospores in the thallus of Porphyra
yezoensis Ueda. J. Appl. Phycol., 15, 345-349.
中山一郎(2005)海藻類の育種に向けての遺伝子研究−ゲ ノム情報を利用したノリ育種.海洋生物の機能(竹井 祥郎編),東海大学出版会,神奈川,pp. 231-245. Noda, H., H. Amano, K. Arashima, S. Hashimoto and K.
Nishizawa (1989) Antitumour activity of polysaccharides and lipids from marine algae. Nippon Suisan Gakkaishi,
55, 1265-1271.
大住幸寛・川合正允・天野秀臣・野田宏行(1998)スサビ ノリポルフィラン由来オリゴ糖の抗腫瘍活性.日水誌,
64,847-853.
Park, J. T. and M. J. Johnson (1949) A submicrodetermination of glucose. J. Biol. Chem., 181, 149-151.
Ren, D., H. Noda, H. Amano, T. Nishino and K. Nishizawa (1994) Study on antihypertensive and antihyperlipidemic effects of marine algae. Fish. Sci., 60, 83-88.
Saga, N., M. Polne-Fuller and A. Gibor (1986) Protoplasts from seaweeds: production and fusion. Beihefte Zur Nur
Nova Hedwigia, 83, 37-43. 嵯峨直恆・田畑哲之(2001)海のモデル植物スサビノリ. 植物のゲノム研究プロトコール(佐々木卓治・田畑哲之・ 島本 功編),秀潤社,東京,pp. 208-215. 植木知佳・飯塚 治・福田 覚・遠藤博寿・北出幸広・安 井 肇・嵯峨直恆(2005)海産紅藻スサビノリにおけ る産業上重要な形質に関する遺伝解析(予報).水産増 殖,53,231-237.