Title
抗真菌剤による肝肥大及び肝発がんへのconstitutive
androstane receptor(CAR)の関与に関する研究( 本文(Fulltext) )
Author(s)
田村, 圭
Report No.(Doctoral
Degree)
博士(獣医学) 乙第150号
Issue Date
2017-03-13
Type
博士論文
Version
ETD
URL
http://hdl.handle.net/20.500.12099/56188
※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。抗真菌剤による肝肥大及び肝発がんへの
constitutive androstane receptor (CAR)の関与
に関する研究
2016 年
岐阜大学大学院連合獣医学研究科
抗真菌剤による肝肥大及び肝発がんへの
constitutive androstane receptor (CAR)の関与
に関する研究
目次 序論 1 第 1 章 トリアゾール系抗真菌剤シプロコナゾール,テブコナゾール,フルコナゾールによる肝 肥大及び肝発がんへのCAR の関与 緒言 6 実験材料及び実験方法 8 実験結果 12 考察 17 小括 20 第 2 章 トリアゾール系抗真菌剤シプロコナゾール,テブコナゾール,フルコナゾールによる肝 肥大におけるCAR の関与と投与用量の関係 緒言 22 実験材料及び実験方法 24 実験結果 27 考察 30 小括 33 第3 章 イミダゾール系抗真菌剤イマザリルによる肝肥大及び肝発がんへの CAR の関与 緒言 35 実験材料及び実験方法 36 実験結果 40 考察 43 小括 45 結論 46 謝辞 50 引用文献 51
要旨 59
Abstract 62 図表 64
1 序論 現在の化学物質のリスク評価の最終目的は,その化学物質が作業者曝露を含む環境あるい は食品を介した曝露においてヒトの健康に影響を及ぼすかを判断することである。リスク評 価の一端を担う毒性評価における大きな課題として,実験動物で認められた毒性影響のヒト への外挿性が挙げられる。化学物質の毒性評価には,げっ歯類を主とした実験動物を用いる が,往々にして毒性には種差が認められる。実験動物で毒性が認められた化学物質のヒトへ の影響,すなわち外挿性の判断が化学物質のリスク評価に重要であるのだが,そこには種差 をどう考えるべきかという大きな壁がある。近年実験動物に特異的な変化と科学的に証明さ れた毒性については,リスク評価においてヒトへの外挿性なしと判断されているが,外挿性 を科学的に明確にできない場合は,実験動物の毒性がヒトへも当てはまるものとして保守的 にリスク評価を進めざるを得ない。 我が国の死因の 1 位はがんであり,化学物質曝露によりヒトで誘発された場合の社会的影 響は非常に大きい。そのため,発がん性には慎重なリスク評価が必要である。そこで本研究 では,肝発がんの外挿性に着目した。肝臓腫瘍は,げっ歯類においても加齢に伴い好発する 腫瘍であり,ヒトへの曝露量よりも高用量で実施する発がん性試験において,非常に高頻度 に誘発される腫瘍である (49)。そのため,肝発がんのヒトへの外挿性の評価は極めて重要で あり,げっ歯類の肝発がんをどのように考えるべきか多くのアプローチがなされてきた (12)。 その結果,げっ歯類の肝発がんの中には,ヒトに外挿されないものがあることが示唆された。 Constitutive androstane receptor (CAR)を介した肝発がんもその 1 つである。CAR は,活性化さ れることでチトクローム P450 (CYP) のような異物代謝酵素の発現を増加させる核内受容体 である (46, 50)。CAR の代表的な活性化物質であるフェノバルビタールをマウスに長期間投 与すると肝発がんが認められる (53)。一方,CAR ノックアウトマウスにフェノバルビタール を長期的に投与しても肝発がんが認められない (51, 55)。これらの結果から,げっ歯類におけ るフェノバルビタールによる肝発がんはCAR に依存していることが明らかになった。フェノ
2 バルビタールは,ヒトのてんかん治療薬として長期的に使用されている。フェノバルビター ルはヒトのCAR を活性化し,ヒトへの投与により肝肥大を誘発することが知られている (13, 40, 41)。しかし,ヒトの疫学調査では,げっ歯類で肝発がんを誘発するフェノバルビタール の曝露量においても肝臓腫瘍の増加は認められていない (22, 29)。以上から,CAR の活性化 による肝発がんはマウスとヒトで種差があることが示唆され,フェノバルビタールによるげ っ歯類の肝発がんは,ヒトには外挿されないことが明らかとなった。フェノバルビタールに 関するげっ歯類及びヒトの結果から,フェノバルビタールに似た特徴を持った化学物質によ る肝発がんもヒトに外挿されないと考えるmode of Action (MOA) が提唱された (12)。この MOA には 2 つの問題点があると考え,その問題点を明らかにすることを本研究の目的とした。
フェノバルビタールは,マウスにCAR を介した肝肥大を誘発する (51, 55)。フェノバルビ タールによるげっ歯類の肝肥大及び肝発がんがいずれもCAR に依存しているため,あたかも 肝肥大が肝発がんのinitial event のように考えられ,上記の MOA では肝肥大が肝発がんの key event の 1 つとされた。その結果,化学物質の毒性試験で肝肥大及び肝発がんがみられた場合 に,その化学物質による肝発がんがフェノバルビタール様,すなわち肝発がんがヒトに外挿 されないと判断されてしまう懸念があった。しかし,肝肥大は様々な要因によって誘発され るため,肝肥大を肝発がんのkey event と考えるのは適切ではなく,肝肥大と肝発がんはそれ ぞれ独立して誘発されていると考えた。以上が本研究を始めるきっかけとなった上記 MOA の問題点の1 つである。 第2 の問題点として,肝発がんに CAR が関わることが証明されている化学物質がフェノバ ルビタールしかないことが挙げられる。また,肝肥大にCAR が関与している化学物質の情報 も少ない。フェノバルビタールのみのデータから,フェノバルビタール様の化学物質が同じ MOA によって肝発がんがヒトに外挿されないとしているが,実際に他の化学物質による肝発 がんへのCAR の関与を確認した報告がないことから,提唱された MOA の検証は十分になさ れていない。そこで,フェノバルビタールと同様にCAR の活性化が示唆される化学物質によ
3
る肝肥大及び肝発がんに,実際にCAR の関与が認められるのか調べる必要があると考えた。 抗真菌剤は,毒性試験において肝肥大及び肝発がんが頻繁に認められる化学物質群である。 CAR の下流の遺伝子である CYP2B 誘導がみられることから CAR の活性化が示唆される抗真 菌剤も多く存在する。しかし,実際に抗真菌剤による肝発がんにCAR が関与しているか調べ た報告はない。そこで本研究では,上記 2 つの問題点を踏まえて,フェノバルビタール以外 の化学物質として抗真菌剤を選択し,抗真菌剤による肝肥大及び肝発がんへのCAR の関与を 調べること,また肝肥大が肝発がんのkey event であるかどうか調べることを目的とした。
CAR の関与は,CAR ノックアウトマウスを用いて調べた。CAR ノックアウトマウスは, フェノバルビタールによる肝肥大及び肝発がんへの CAR の関与を調べた際にも使用された 有用な試験系である。また,肝発がんは,肝発がんのイニシエーターであるジエチルニトロ ソアミン (DEN) を用いたイニシエーションプロモーションモデルによって検討した。イニシ エーションプロモーションモデルは,げっ歯類の肝発がんを調べる方法として,成書でも紹 介されるような古くからよく使用されている方法である (7, 25, 43)。 第1 章では,トリアゾール系抗真菌剤であるシプロコナゾール,テブコナゾール,及びフ ルコナゾールによる肝肥大及び肝発がんへのCAR の関与について調べた。トリアゾール系抗 真菌剤は,多くの剤でげっ歯類に肝肥大及び肝発がんを誘発するが,肝発がんへのCAR の関 与が明らかにされた報告はなく,肝肥大についてもわずかな報告しかない。また,各トリア ゾール系抗真菌剤による肝肥大及び肝発がんへのCAR の関与の有無を調べることで,肝発が んと肝肥大の関係性について検討した。 第1 章の結果から,使用した 3 種のトリアゾール系抗真菌剤による肝肥大には CAR 以外の 核内受容体の関与が示唆された。核内受容体は多岐にわたり,化学物質によっては複数の核 内受容体をそれぞれ違う割合で活性化させることが考えられる。実際に,投与用量によって 活性化される核内受容体が異なる可能性が示唆されている (10)。第 1 章では,肝発がんの解 析に注力しため,腫瘍誘発用量で投与した。しかし,当該用量では,テブコナゾールによる
4 肝肥大は程度が強く,フルコナゾールによる肝肥大は程度が弱かったため,CAR を含めた核 内受容体の肝肥大への関与を詳細に調べるためには課題があった。そこで,第 2 章では,第 1 章で用いた 3 種のトリアゾール系抗真菌をそれぞれ複数用量投与し,肝肥大への CAR を含 めた核内受容体の関与を詳細に検討することとした。 第3 章では,第 1 章及び第 2 章で確立した試験系を用いて,げっ歯類に肝肥大及び肝発が んを誘発することが知られているイマザリルを選択し,同様に肝肥大及び肝発がんへのCAR の関与の有無及び複数用量投与による肝肥大に関する核内受容体の詳細解析を検討した。イ マザリルは,イミダゾール系抗真菌剤の 1 つであり,げっ歯類の肝肥大及び肝発がんのいず れについてもCAR の関与に関する報告はない。 以上の実験により,トリアゾール系3 剤,イミダゾール系 1 剤の抗真菌剤による肝肥大及 び肝発がんへのCAR の関与を調べた。また,それぞれの剤による肝肥大及び肝発がんに関わ るCAR あるいはその他の核内受容体を調べることで,肝肥大と肝発がんの関係性についても 検討した。本研究では,肝発がんの外挿性についての新たなエビデンスを取得し,化学物質 のリスク評価に貢献することを目的とした。
5 第1 章
トリアゾール系抗真菌剤シプロコナゾール,テブコナゾール,フルコナゾールによる 肝肥大及び肝発がんへのCAR の関与
6 緒言 トリアゾール系抗真菌剤は,農薬や医薬品として広く用いられている。トリアゾールは, 14-α demethylase 活性を阻害し,真菌のステロール合成を阻害することで抗真菌作用を発揮す る (9)。毒性及び発がん性評価において,多くのトリアゾール系抗真菌剤がげっ歯類に肝肥大 及び肝発がんを誘発することが報告されている (15, 16, 17, 18, 19, 20, 21, 45)。しかし,トリア ゾール系抗真菌剤の肝肥大あるいは肝発がん性の機序研究は少なく,CAR の関与についての 報告はわずかである。すなわち,後述するシプロコナゾールを除くと,トリアゾール系抗真 菌剤とCAR の関係性については,プロピコナゾール,トリアジメフォン,及びボリコナゾー ルについて限られた報告があるだけである。プロピコナゾール及びトリアジメフォンが,フ ェノバルビタールと同様にマウスに肝肥大,CYP2B 発現の増加,及び肝細胞増殖の活性化を 誘発し,アポトーシスを抑制したため,CAR の関与が示唆されていた (1, 15, 16)。しかし, マイクロアレイ解析の結果では,これら 2 つのトリアゾール系抗真菌剤を投与したマウスの 肝臓における遺伝子発現はフェノバルビタールによるものと明らかに異なっており,肝肥大 や肝発がんへのCAR の関与は不明なままである (33)。プロピコナゾールについては,トキシ コゲノミクス解析によりフェノバルビタール様のMOA であると報告されているが,CAR ノ ックアウトマウスを用いた実験ではないことから直接的なCAR の関与は示されていない (5)。 ボリコナゾールはマウスにおける代謝にCAR 及び pregnane X receptor (PXR) の関与が示唆さ れたが,肝肥大へのCAR の関与については調べられていない (34)。また,トリアゾール系抗 真菌剤による肝発がんへのCAR の関与を明らかにした報告は認められなかった。 シプロコナゾールとテブコナゾールは農薬の殺菌剤として,フルコナゾールは医薬品の抗 真菌剤として用いられている。いずれの 3 剤も肝肥大あるいは肝発がんを誘発することが知 られているトリアゾール系抗真菌剤である (18, 45, 47)。シプロコナゾールについては,CAR ノックアウトマウスを用いた短期間の実験によって,シプロコナゾールによる肝肥大及び肝 細胞増殖活性にCAR が関与していることが報告されている (38)。テブコナゾールについての
7
CAR に関連する報告はない。フルコナゾールは,CAR の下流の CYPs 遺伝子発現や酵素活性 を増加させることからCAR の関与が示唆されている (10)。 そこで本章では,CAR ノックアウトマウスを用いて,これら 3 種のトリアゾール系抗真菌 剤による肝肥大及び肝発がんにCAR が関与しているか調べた。肝発がんは,イニシエーショ ンプロモーションモデルを用いて評価した。本実験で検索した物質の遺伝毒性については, 農薬及び医薬品ガイドラインを満たす十分な数のin vitro,in vivo 遺伝毒性試験が実施されて おり,いずれの化学物質についても遺伝毒性はないことが知られている (6, 18, 45)。
8
実験材料及び実験方法
化合物
本実験で用いたトリアゾール系抗真菌剤の化学構造式を図 1-1 に示す 。シプロコナゾール (CAS No. 94631-06-5,純度 96.7%),テブコナゾール (CAS No. 107534-96-3,純度 97.3%),及 びフルコナゾール (CAS No. 86386-73-4,純度 99.5%以上) は,LKT ラボラトリー社 (St. Paul, MN, USA) より入手した。DEN (CAS No. 55-18-5,純度 99.9%) は,東京化成工業株式会社 (Tokyo, Japan) より入手した。 動物 C3H/HeNCrl マウスを背景とした雄性 CAR ノックアウトマウス (26, 55) は根岸博士よりご 厚意でご提供いただいた。同ノックアウトマウスには C57BL 系統由来と C3H 系統由来マウ スが存在するが,今回の実験の目的が肝発がん性であることから,同博士より肝腫瘍好発系 であるC3H 系統を供与頂いた。また,雌よりも雄で肝臓腫瘍が好発することから雄を選択し た。供与後,CAR ノックアウトマウスは,国立医薬品食品衛生研究所内で継代繁殖させ,試 験実施可能な個体数まで繁殖用動物数を確保したのち,本試験に用いた。野生型動物として 5 週齢の雄性 C3H/HeNCrlCrlj を日本チャールス・リバー株式会社 (Kanagawa, Japan) より購 入した。なお,全ての動物実験は,国立医薬品食品衛生研究所の動物実験委員会に承認を受 け,国立医薬品食品衛生研究所の動物実験の適切な実施に関する規定に準拠して行った。 化合物の投与 肝発がんを調べる方法として,肝発がんのイニシエーターであるDEN を用いたイニシエー ションプロモーションモデルを用いた (7, 43)。肝発がんのイニシエーション処置として 13 及び27 週投与群には,6 週齢時に DEN を 90 mg/kg の用量で単回腹腔内投与した。4 週間投 与群は主に肝肥大について解析するため DEN 処置は実施しなかった。6 週齢の雄性 CAR ノ
9 ックアウトマウス及び野生型マウスを,体重を指標として無作為に4,13,及び 27 週投与群 にそれぞれ群分けした。投与期間は,それぞれ4,13,あるいは 27 週間とした。各群の動物 数は,4 及び 13 週投与群は 5 あるいは 6 匹とし,27 週投与群は 22 から 25 匹とした。7 週齢 からシプロコナゾール,テブコナゾール,あるいはフルコナゾールをそれぞれ 200,1500, 200 ppm の濃度で混餌投与した。また,対照として,粉末基本飼料 (CRF-1,オリエンタル酵 母工業株式会社) を自由摂餌させる群を設定した。シプロコナゾール及びテブコナゾールの 投与濃度は,肝肥大及び肝発がんを誘発することが知られている投与濃度を設定した (18, 38, 45)。フルコナゾールは,diflucan (フルコナゾールの医薬品名,ファイザー株式会社) の経口 投与によるマウスのがん原性試験において,10 mg/kg/day まで肝発がんを誘発しなかった (6)。 そのため,本実験では,マウスにおいて Cyp2b10 遺伝子発現の増加及び肝肥大を誘発するこ とが報告 (47) されている 25 mg/kg/day の用量に相当するように 200 ppm の投与濃度を設定し た。混餌飼料は動物に与えるまで冷蔵保存した。給餌は毎週新しい飼料と交換し,餌缶内に 残った飼料は廃棄した。 動物の観察及び肝毒性指標の測定 一般状態観察を1 日 1 回以上実施した。体重及び摂餌量は,投与 13 週までは 1 週間に 1 回 測定し,その後は2 週間に 1 回測定した。投与 4,13,あるいは 27 週間後に,吸入麻酔によ る全身深麻酔下で心臓から採血し,マウスを放血により安楽死させた。血清試料は,使用す るまで-30℃で保存した。肝毒性を調べることを目的として,血清中のアラニンアミノトラン スフェラーゼ (ALT) を測定した。ALT の測定は,株式会社エスアールエル (Tokyo, Japan) で 実施した。
肝臓の病理学的及び免疫組織化学的評価
4 及び 13 週投与群では肉眼的に観察し,肝臓重量を測定した後に,10%中性緩衝ホルマリ ン溶液によって肝臓の中間葉を固定した。パラフィン包埋した肝臓を,薄切し,常法に従っ
10
てヘマトキシリンエオジン (HE) 染色した。肝細胞肥大の程度は,軽度 (mild),中等度 (moderate),重度 (marked) の 3 段階に分類した。肝細胞肥大が小葉中心性のみに限局してい た場合は,軽度あるいは中等度とし,小葉中心から中間帯まで認められた場合に重度とした。
4 週投与群の肝臓を用いて免疫組織化学的に評価した。HE 染色標本と連続した切片に一次 抗体として抗 PCNA 抗体 (Dako, Japan) あるいは抗 CYP2B 抗体 (Enzo Life Sciences Inc., Farmingdale, NY, USA) を添加し,4℃条件下で一晩免疫反応した。その後,ペルオキシダーゼ で標識したデキストランポリマーが結合した二次抗体 (Histofine Simple Stain mouse MAX PO, Nichirei, Tokyo, Japan) を反応させ,3-3’-diaminobenzidine (Sigma Chemical Co., St. Louis, MO, USA) により視覚化した。また,核を染色するため,ヘマトキシリン染色した。 肝細胞増殖活性は,PCNA 染色した切片を用いて,視野中にある PCNA 陽性細胞数を指標 に評価した。すなわち,対物レンズ10 倍の光学顕微鏡下で,各動物の肝臓を 5 視野 (1 視野 に肝小葉全体が入るようにする) ずつ観察し,各視野における陽性細胞数を数えた。次に, 各視野の陽性細胞数を合計し,合計した数に応じて 4 つのグレードに分類した。各グレード の合計陽性細胞数は,グレード0:10 未満,グレード 1:10~25,グレード 2:26~50,グレ ード3:51 以上とした。 27 週間投与群では,変異肝細胞巣及び肝腫瘍の数を数えるため,肝臓の中間葉及び左葉を それぞれ 6 切片ずつ作製した。肝臓の増殖性病変の分類は,the criteria of the International Harmonization of Nomenclature and Diagnostic Criteria of Lesions in Rats and Mice (49) に従った。 フェノバルビタールは主として好酸性の変異肝細胞巣及び肝腺腫を誘発することが知られて いる (55)。そのため,本実験において,変異肝細胞巣及び肝腫瘍は,それぞれの形態的特徴 及び染色性に基づいて,好酸性,好塩基性,その他 (明細胞性及び混合型) のいずれかに分類 した。増殖性病変の発生頻度 (病変が認められた動物数) 及び multiplicity (1 個体あたりの病 変数) について計算した。
11 RT-real time PCR
肝薬物代謝酵素で肝肥大に関連すると報告されている分子種を中心に解析を行った。4 週 投与群では,肝臓左葉の一部を安楽死後即座に-80℃で凍結した。凍結した肝臓から,ISOGEN (Nippon Gene Co. Ltd., Tokyo, Japan) を用いて RNA を抽出した。抽出した 1 μg の RNA を High Capacity Reverse Transcription kit (Applied Biosystems, Foster City, CA, USA) を用いて逆転写し, TaqMan Fast Universal PCR Master Mix (Applied Biosystems) 及び TaqMan Gene Expression Assays (Applied Biosystems) を用いてリアルタイム PCR (7900HT Fast Real-time PCR System, Applied Biosystems) を 実 施 し た 。 本 実 験 で は , Cyp1a2 (Mm00487224_m1) , Cyp2b10 (Mm01972453_s1),Cyp3a11 (Mm00731567_m1),Cyp4a10 (Mm01188913_g1),及び P450 reductase (Mm00435876_m1) のプライマープローブセットを用いた。それぞれの遺伝子発現レベルは,
内因性のGAPDH (TaqMan Rodent GAPDH Control Reagent, Applied Biosystems) で標準化し,相
対標準曲線法により計算した。また,Gadd45β は抗アポトーシス及び CAR に関連することが 知られている (36, 37, 56) ことから,過去に小澤らが報告 (35) した方法で Gadd45β について も遺伝子発現レベルを調べた。。 統計解析 統計解析は,対照群と化合物投与群 (シプロコナゾール投与群,テブコナゾール投与群, 及びフルコナゾール投与群) の比較を CAR ノックアウトマウス及び野生型マウスそれぞれに ついて実施した。また,CAR ノックアウトマウスと野生型マウスの比較を同じ投与群同士で 比較した。体重,肝臓絶対重量,肝臓相対重量,mRNA 発現量,及び肝臓増殖性病変の multiplicity のデータについて,F 検定により等分散性の検定を実施した.等分散性が認められ た場合はStudent の t 検定を,等分散性が認められなかった場合は Welch の t 検定を用いて, それぞれ媒体対照群と化合物投与群との間で比較した。肝臓増殖性病変の発生頻度のデータ は,Fisher の直接確率検定によって統計解析した。有意水準は 5%及び 1%とした。
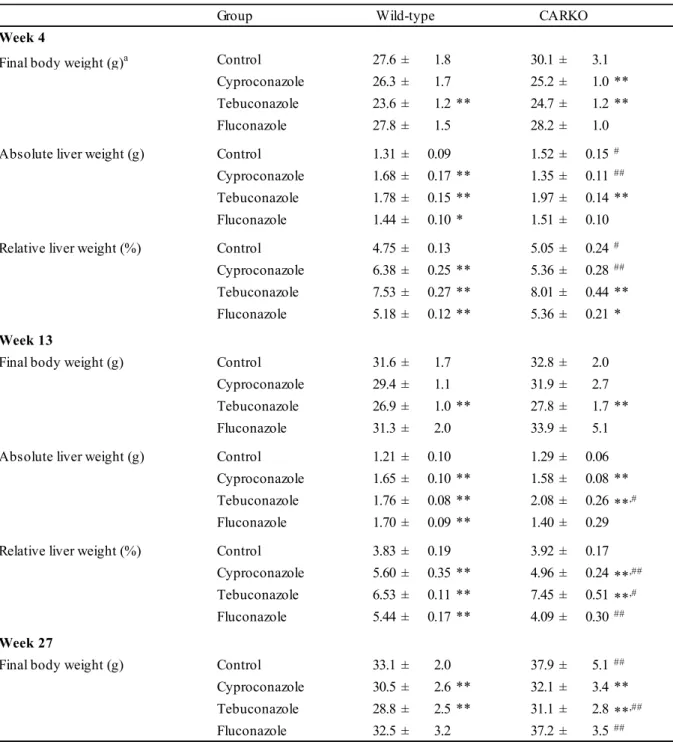
12 実験結果 一般状態,体重,摂餌量,及び肝臓重量 投与4,13,及び27週後の最終体重及び肝重量の結果を表1-1に示す。投与期間中に化合物 に起因した死亡及び一般状態の異常は認められなかった。シプロコナゾール投与群及びテブ コナゾール投与群では,両遺伝子型のマウスにおいて持続的に体重が減少した。フルコナゾ ールの投与は体重に影響しなかった。テブコナゾール投与群の野生型マウスでは,摂餌量が 対照群と比較して減少していたが,その他の群では対照群と同等以上であった。投与27週間 のシプロコナゾール,テブコナゾール,及びフルコナゾールの摂取量は,それぞれ34~39, 226~336,及び33~34 mg/kg/dayであった。 野生型マウスでは,全ての化合物投与群において,肝臓絶対及び相対重量が有意に増加し た。肝臓重量増加の程度は,テブコナゾール,シプロコナゾール,フルコナゾールの順に強 かった。シプロコナゾールを投与したCARノックアウトマウスでは,投与13週後に肝臓重量 が増加したが,その増加量は野生型マウスと比較して少なかった。テブコナゾール投与群の CARノックアウトマウスでは,投与4及び13週後のいずれにおいても,肝重量が野生型マウス と同程度に増加した。フルコナゾール投与群のCARノックアウトマウスでは,いずれの剖検 時期でも対照群と同程度の肝臓重量であった。フルコナゾール投与群において,投与4週後で は野生型マウスとCARノックアウトマウスの肝臓重量に差は認められなかったが,投与13週 後では有意な差が認められた。 肝臓の形態学的変化 (肝細胞の増殖性病変以外) 及び ALT の変動 投与4,13,及び27週後の肝臓の病理組織学的変化及びALTの結果を表1-2,表1-3,及び図 1-2に示す。野生型マウスでは,化合物投与群の全てのマウスで小葉中心性肝細胞肥大が認め られた。しかし,その重篤度は,化合物によって異なっていた。テブコナゾール投与群では, 重篤な肝細胞肥大が認められた。シプロコナゾール投与群では,ほとんどの動物で中等度の
13 小葉中心性肝細胞肥大が認められた。フルコナゾール投与群では,他の2つのトリアゾールと 比較して軽微な肝細胞肥大が認められた。CARノックアウトマウスにおいては,シプロコナ ゾール投与群では,投与13週後に軽度な肝細胞肥大が認められた。しかし,その肥大の程度 は,野生型マウスよりも軽度であった。一方,テブコナゾール投与群では,CARノックアウ トマウスにおいても野生型マウスと同程度に重度な肝細胞肥大が認められた。フルコナゾー ル投与群のCARノックアウトマウスでは,投与4及び13週後には肝細胞肥大は認められなかっ た。投与27週後では,わずかに肝細胞肥大がみられた。 化合物に起因したその他の病理所見として,シプロコナゾール投与群では,投与27週後に 褐色色素沈着が両遺伝子型マウスで認められた。テブコナゾール投与群では,投与4週後に肝 細胞の空胞化及び炎症性細胞浸潤,投与27週後にoval cellの過形成 (増生) があり肝傷害像と それに伴う炎症性変化が認められた。全ての投与期間において,テブコナゾール投与群の両 遺伝子型のマウス及びシプロコナゾール投与群の野生型マウスにALTの増加が認められた。 フルコナゾール投与群では,いずれのマウスにおいても肝毒性を示唆する変化は認められな かった。 肝細胞増殖活性 肝細胞増殖活性の結果を図1-3に示す。肝細胞増殖活性は,PCNA陽性細胞数として評価し た。テブコナゾール投与群では,投与4及び13週後に,両遺伝子型マウスで,肝細胞増殖活性 が対照群と比較して2.3~7.2倍増加していた。シプロコナゾール投与群及びフルコナゾール投 与群では,投与に起因した肝細胞増殖活性の変化はいずれの遺伝子型マウスでも認められな かった。 肝臓におけるCYP 発現 CYPs及びGADD45BETAをコードしたmRNA発現量を図1-4に示す。野生型マウスでは, Cyp2b10遺伝子の発現量がいずれの化合物投与群においても増加しており,その増加量は対照
14 群と比較して69~337倍であった。シプロコナゾール投与群及びテブコナゾール投与群では, CARノックアウトマウスにおいてもCyp2b10発現量が有意に増加した。しかし,シプロコナゾ ール投与群では,CARノックアウトマウスにおけるCyp2b10発現量は,野生型マウスと比較し て低かった。Cyp3a11発現量は,いずれの化合物投与群においても両遺伝子型マウスで有意に 増加した。シプロコナゾール投与群及びフルコナゾール投与群では,CARノックアウトマウ スにおいてCyp3a11発現量が野生型マウスよりも低かったが,その減少幅はわずかであった。 テブコナゾール投与群では,両遺伝子型マウスにおいてCyp4a10発現量が増加した。テブコナ ゾール投与群の両遺伝子型マウス及びシプロコナゾール投与群の野生型マウスにおいて,わ ずかではあるもののCyp1a2発現量が有意に増加した。CARノックアウトマウスにおける Cyp1a2発現量は,全ての群において野生型マウスよりも低かった。P450 reductase発現量は, いずれの化合物投与群においても対照群よりも高く,その増加量はCARノックアウトマウス よりも野生型マウスの方が高かった。シプロコナゾール投与群及びテブコナゾール投与群で は,Gadd45β発現量が両遺伝子型マウスで増加した。 抗CYP2B抗体による免疫組織学的検査の結果を図1-5に示す。投与4週後の肝臓を用いた CYP2Bの免疫組織化学的評価では,全ての化合物投与群の野生型マウスにおいて,肥大した 肝細胞に強い陽性反応を示した。シプロコナゾール投与群のCARノックアウトマウスでは, 小葉中心部に弱い陽性反応が認められたが,その反応は明らかに野生型マウスよりも減弱し ていた。一方,テブコナゾール投与群では,CARノックアウトマウスでも野生型マウスと同 等の強い陽性反応が認められた。フルコナゾール投与群のCARノックアウトマウスでは,陽 性反応はなかった。 肝発がん性の解析 変異肝細胞巣あるいは肝細胞腺腫の発生頻度を表1-4に示す。投与13週後では,野生型マウ スにわずかな変異肝細胞巣が認められたものの,化合物投与による発生頻度の差はなかった。 また,投与13週後のCARノックアウトマウスでは,増殖性病変は認められなかった。投与27
15 週後では,シプロコナゾール投与群の野生型マウスに好酸性肝細胞腺腫の発生頻度が増加し た。一方,CARノックアウトマウスのシプロコナゾール投与群では,好酸性肝細胞腺腫の発 生頻度が野生型マウスよりも低く,CARノックアウトマウスの対照群と同程度であった。テ ブコナゾール投与群及び対照群では,好酸性変異肝細胞巣の発生頻度が野生型マウスよりも CARノックアウトマウスで低かった。フルコナゾール投与群では,好酸性肝細胞腺腫の発生 頻度が野生型マウスよりもCARノックアウトマウスで低かった。テブコナゾール投与群では, 好塩基性変異肝細胞巣あるいは肝細胞腺腫の発生頻度が対照群と比較して低かった。また, テブコナゾール投与群のCARノックアウトマウスでは,野生型マウスと比較して,好塩基性 変異肝細胞巣の発生頻度が低かった。 変異肝細胞巣及び肝細胞腺腫のmultiplicityを図1-6に示した。野生型マウスにおいて,シプ ロコナゾールは,好酸性変異肝細胞巣及び肝細胞腺腫のmultiplicityを増加させた。テブコナゾ ールは好酸性変異肝細胞巣について増加させた。フルコナゾール投与群では,好酸性増殖性 病変の増加に統計学的有意差がなかった。シプロコナゾール投与群では,CARノックアウト マウスの好酸性変異肝細胞巣及び肝細胞腺腫のmultiplicityが,野生型マウスと比較して有意に 減少していた。シプロコナゾールは,CARノックアウトマウスにおいても好酸性変異肝細胞 巣を増加させたが,好酸性肝細胞腺腫は増加させなかった。テブコナゾール投与群では,シ プロコナゾールと同様に,CARノックアウトマウスの好酸性変異肝細胞巣及び肝細胞腺腫の multiplicityが,野生型マウスと比較して減少していた。フルコナゾール投与群では,CARノッ クアウトマウスの好酸性肝細胞腺腫のmultiplicityが野生型マウスよりも減少していた。対照群 において,好酸性変異肝細胞巣のmultiplicityが,野生型マウスよりもCARノックアウトマウス で低かった。野生型マウスにおいて,シプロコナゾールは,好塩基性変異肝細胞巣のmultiplicity を増加させたが,好塩基性肝細胞腺腫は増加しなかった。テブコナゾール投与群では,野生 型及びCARノックアウトマウスで,好塩基性変異肝細胞巣及び肝細胞腺腫のmultiplicityが減少 した。シプロコナゾール及びテブコナゾール投与群のCARノックアウトマウスにおける好塩
16
基性変異肝細胞巣及び肝細胞腺腫のmultiplicityは,野生型マウスよりも低かった。その他 (明 細胞性及び混合型) の増殖性病変については,いずれの化合物あるいは遺伝子型でも有意な 差は認められなかった。
17 考察 本実験で用いた 3 種のトリアゾール系抗真菌剤による肝肥大及び肝発がんへの関与が示唆 される経路を図 1-7 に示す。本実験で明らかとなった 1 点目として,同じトリアゾール系抗 真菌剤に属していても化合物によって肝肥大における CAR の関与が異なることが明らかに なった。すなわち,シプロコナゾール及びフルコナゾールによる肝肥大にはCAR が関与して いたが,テブコナゾールによる肝肥大にはCAR が関与していなかった。特にフルコナゾール では,本実験に用いた投与濃度において,CAR ノックアウトマウスで肝肥大は観察されず, CAR に依存した肝肥大であると考えられた。加えて,シプロコナゾール及びフルコナゾール 投与群では,投与4 週後に,Cyp2b10 遺伝子及び CYP2B 蛋白の発現量が CAR ノックアウト マウスでは抑制されたことから,CAR が関与した CYP2B 発現と考えられた。シプロコナゾ ール及びフルコナゾール投与群では,CAR ノックアウトマウスにおいても Cyp3a11 発現量が 増加していたことから,PXR も肝肥大に関与している可能性が考えられた。シプロコナゾー ル投与群のCAR ノックアウトマウスに肝肥大が認められたことも PXR が肝肥大に関与して いることを支持している。これら 2 つのトリアゾールに対してフェノバルビタールは,野生 型マウスにおいて CAR しか活性化させず,CAR ノックアウトマウスでは肝肥大や肝発がん を誘発しないことが本実験と同様の方法で実施した実験で明らかとなっている (43)。一方, テブコナゾール投与群では,CAR ノックアウトマウスにおいても野生型マウスと同等の肝肥 大及び CYP2B 発現が認められていることから,テブコナゾールによる肝肥大には主として CAR 以外の経路が関与していることが考えられた。テブコナゾールは,Cyp3a11 及び Cyp4a10 発現量を両遺伝子型マウスで増加させた。従って,テブコナゾールによる肝肥大には,PXR あるいはperoxisome proliferator activated receptor alpha (PPARα) が関与している可能性が考え られた。
本実験で明らかになった 2 点目は,げっ歯類のイニシエーションプロモーションモデル を用いた肝発がん性の結果より,テブコナゾールを含む 3 種のトリアゾール系抗真菌剤によ
18
る肝発がんにCAR が重要な役割を果たしていることが明らかになった点である。これまで肝 肥大は,げっ歯類のCAR が関与する肝発がんの MOA の中で key event として認識されてき た(12)。しかし,テブコナゾールは,CAR 以外の経路で肝肥大を誘発し,CAR を介して肝発 がんを誘発した。また,CYP2B 誘導剤である Piperonyl Butoxide (PBO) は,テブコナゾール と同様に,肝肥大に CAR が関与していないにも関わらず,CAR を介した肝発がんを誘発し た(43)。このように,肝発がんと肝肥大への CAR の関与が異なっていたことから,肝肥大は 肝発がんのkey event ではないことが示唆された。肝発がんの key event の候補の 1 つとして細 胞増殖活性の増加が考えられている。本実験において,テブコナゾール投与群では,投与 4 及び13 週に,両遺伝子型マウスで肝細胞増殖活性が増加傾向であった。テブコナゾール投与 群では,両遺伝子型マウスに炎症性細胞浸潤,oval cell 過形成,及び ALT 増加といった肝毒 性を示唆する所見が投与 4 週後から認められた。これらの結果は,テブコナゾールが細胞毒 性による持続的な細胞増殖を誘発していたことを示唆していた。しかし,CAR ノックアウト マウスに持続的な肝毒性があったにも関わらず,肝発がんが誘発されなかったことから,肝 毒性が肝発がんに影響していなかったと考えられた。そのため,テブコナゾールによる肝発 がんには肝毒性による細胞増殖活性の増加ではなく,CAR が関与していると考えられた。
Gadd45beta は,NF-κB が関与した c-Jun N-terminal kinase の抑制による抗アポトーシス要因
の1 つである (36, 37)。また,Gadd45beta の発現は CAR によって調整されていることが知ら れている (56)。しかしながら,本実験では,シプロコナゾール及びテブコナゾール投与群に おけるGadd45β 発現量の増加は,野生型マウスに加え,CAR ノックアウトマウスにおいても 認められた。この結果から,Gadd45β が関与した抗アポトーシスシグナルは,CAR を介した 肝発がんには関与していない可能性が考えられた。 シプロコナゾールを用いた過去の報告では,肝肥大及び肝細胞増殖活性へのCAR の関与と 投与用量の関係が示されている (38)。本実験では,トリアゾール系抗真菌剤による肝発がん へのCAR の関与を明らかにすることを第一の目的に実施したため,腫瘍誘発用量で投与した。
19
そのため,テブコナゾールでは肝肥大の程度が強く,フルコナゾールでは肝肥大の程度が弱 かった。本実験では,肝肥大におけるCAR の関与を詳細に調べることができなかったことが 課題として残された。
20 小括 本実験では,シプロコナゾール,テブコナゾール,及びフルコナゾールの 3 種のトリアゾ ール系抗真菌剤を野生型及びCAR ノックアウトマウスに投与し,これらのトリアゾール系抗 真菌剤による肝肥大及び肝発がんにおけるCAR の関与を調べた。その結果,シプロコナゾー ル及びフルコナゾールによる肝肥大ではCAR が主として関与していたが,テブコナゾールに よる肝肥大にはCAR は関与していなかった。一方で,3 種のトリアゾール系抗真菌剤による 肝発がんには,いずれもCAR が重要な役割を果たしていた。これらの結果から,げっ歯類の CAR を介した肝発がんでは,肝肥大は key event ではないことが示唆された。
21 第2 章
トリアゾール系抗真菌剤シプロコナゾール,テブコナゾール,フルコナゾールによる 肝肥大におけるCAR の関与と投与用量の関係
22 緒言 第1 章の結果から,使用した 3 種のトリアゾール系抗真菌剤による肝肥大には CAR 以外の 核内受容体の関与が示唆された。核内受容体は多岐にわたり,化学物質によっては複数の核 内受容体をそれぞれ違う割合で活性化させることが考えられる。実際に,投与用量によって 活性化される核内受容体が異なる可能性が示唆されている (10)。第 1 章では,腫瘍誘発用量 における肝肥大あるいは関連遺伝子の発現を調べたが,CAR やそれ以外の核内受容体の関与 は明確に示すことができなかった。また,第 1 章で用いたテブコナゾールの腫瘍誘発用量は maximum tolerated dose (MTD) を超えており,肝肥大の程度も重度であった。重度の肝肥大で は,核内受容体の関与があったとしても,弱い活性化ではマスクされてしまい,その関与を 確認することができない可能性がある。一方,フルコナゾール投与群では,肝肥大の程度は 弱かったため,核内受容体の関与があったとしても検出できなかった可能性が考えられる。 そのため,フルコナゾールでは,第 1 章で用いた用量より高い用量における肝臓への影響を 調べることは有意義であると考えられた。 既存の文献によるとシプロコナゾールによる肝肥大及び細胞増殖活性では,CAR ノックア ウトマウスを用いた短期間の試験において,CAR が重要な役割を果たしていることが報告さ れている (38)。また,同報告では,低用量では CAR ノックアウトマウスにおいて細胞増殖活 性が完全に抑制されているのに対し,高用量ではCAR ノックアウトマウスにおいてもわずか に細胞増殖活性の増加がみられた。このように,投与用量によって反応が異なり,関与して いる経路も異なる可能性が示唆されている。また,テブコナゾール及びフルコナゾールにつ いては,CAR の関与と投与用量の関係について調べた報告はない。 加えて,投与初期における細胞増殖活性の増加が肝発がんに関連している可能性が考えら れており (42),短い投与期間における細胞増殖活性について調べる意義は大きい。また,テ ブコナゾール及びフルコナゾールでは,投与初期の細胞増殖活性へのCAR の関与の報告はな い。
23 そこで,本章では,第 1 章で用いた 3 種のトリアゾール系抗真菌剤 (シプロコナゾール, テブコナゾール,及びフルコナゾール) の投与 1 週間後における肝肥大,CYP2B を主体とし た関連遺伝子の発現量,及び肝細胞の細胞増殖活性へのCAR の関与と投与用量の関係性につ いて検討した。本章では,フェノバルビタールとの違いをより明確にするために,比較対象 としてフェノバルビタールについても評価を行った。
24
実験材料及び実験方法
化合物
シプロコナゾール,テブコナゾール,及びフルコナゾールは,第 1 章で用いたものと同じ ものを使用した。フェノバルビタール (Phenobarbital sodium, CAS No. 57-30-7, purity 98.5%) は, 和光純薬工業株式会社 (Osaka, Japan) より購入した。 動物 第1 章と同様に,国立医薬品食品衛生研究所で繁殖している雄性 CAR ノックアウトマウス 及び日本チャールス・リバー株式会社より購入した野生型マウスを用いた。なお,全ての動 物実験は国立医薬品食品衛生研究所の動物実験委員会に承認を受け,国立医薬品食品衛生研 究所の動物実験の適切な実施に関する規定に準拠して行った。 化合物の投与 7 週齢の CAR ノックアウトマウス及び野生型マウスを,体重を基にして無作為に 13 群に 群分けした。投与群は,シプロコナゾール,テブコナゾール,フルコナゾール,及びフェノ バルビタールの4 種の化合物投与群それぞれに低,中,及び高用量群の 3 群の計 12 群を設定 し,それに加えて粉末基本飼料 (CRF-1, Oriental Yeast Co., Ltd., Tokyo, Japan) を自由摂餌させ る対照群を設定した。各群の動物数は 5 匹とした。投与濃度は,シプロコナゾールを 100, 200,及び 400 ppm,テブコナゾールを 375,750,及び 1500 ppm,フルコナゾールを 200,400, 及び800 ppm,そしてフェノバルビタールを 250,500,及び 1000 ppm の濃度で 7 日間投与し た。投与濃度及び投与期間は,過去の報告 (18, 38, 45, 47) や第 1 章の結果を根拠に設定した。 動物の観察及び肝毒性指標の測定 動物の一般状態は,1 日 1 回以上観察した。体重は投与期間中に 3 回,摂餌量は投与期間 中に 2 回測定した。解剖は,第 1 章と同様に実施した。すなわち全身深麻酔下で採血し,全
25
身の肉眼的観察,肝臓重量測定,肝臓サンプル採取を実施した。また,血清を用いたALT の 測定は,株式会社エスアールエル (Tokyo, Japan) で実施した。
肝臓の病理学的及び免疫組織化学的評価
肝臓の中間葉を10%中性緩衝ホルマリン溶液に浸漬させ,第 1 章と同様に,HE 染色標本 を作製した。また,抗CYP2B 抗体 (Enzo Life Sciences Inc., Farmingdale, NY, USA) 及び抗 PCNA 抗体 (PCNA, Clone PC10, DAKO, Glostrup, DNK) を用いて,第 1 章と同様に免疫組織化学的 検査を実施した。肝細胞増殖活性は,第 1 章と同様に,1 視野あたりの PCNA 陽性細胞数に 基づいて,グレード0~3 で評価した。
RT-real time PCR
肝臓の左葉を用いて,第1 章と同様の手法により RT-real time PCR を用いて遺伝子発現量を 評価した。Cyp1a2 (Mm00487224 m1), Cyp2b10 (Mm01972453 s1), Cyp3a11 (Mm00731567 m1), Cyp4a10 (Mm01188913 g1),及び P450 reductase (Mm00435876 m1) の発現量を GAPDH で標準
化して比較した。
肝肥大の解析
肝肥大については,肝臓重量及び病理組織学的に診断した肝細胞肥大を以下の基準よりス コア化し,その合計を肝肥大スコアとして評価した。肝臓重量及び肝細胞肥大のスコアは, 野生型マウスのフェノバルビタール低用量群を 100 とした相対値で表した。肝細胞肥大は, 所見の重篤度をスコア化した。異常なし,極軽微 (very slight),軽微 (slight),軽度 (mild), 及び中等度 (moderate) の重篤度は,それぞれスコア 0,1,2,3,及び 4 として計算した。各 遺伝子型マウスの肝臓重量と肝細胞肥大の合計スコアから肝肥大への CAR の関与を評価し た。
26 統計解析 統計解析は,対照群と各用量の化合物投与群の比較を各遺伝子型マウスで,それぞれ実施 した。また,CAR ノックアウトマウスと野生型マウスの比較をそれぞれの同じ投与群 (同化 合物同用量) 同士で比較した。対照群と化合物投与群の比較は Dunnett 検定 (等分散性あり) あるいはSteel 検定 (等分散性なし) を用い,野生型と CAR ノックアウトマウスとの比較では, 第1 章と同様に,Student の t 検定 (等分散性あり) あるいは Welch の t 検定 (等分散性なし) を 用いた。統計解析は,体重,肝臓絶対重量,肝臓相対重量,ALT,及び mRNA 発現量のデー タを対象とした。有意水準は5%及び 1%とした。
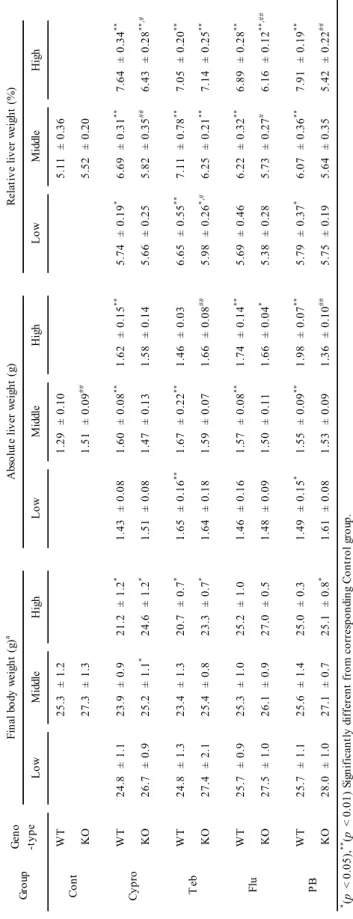
27 実験結果 一般状態,体重,摂餌量,及び肝臓重量 最終体重及び肝臓重量の結果を表 2-1 に示す。投与期間中に化合物に起因した一般状態の 異常は認められなかった。シプロコナゾール中用量群のCAR ノックアウトマウス,シプロコ ナゾール及びテブコナゾール高用量群の両遺伝子型マウス,及びフェノバルビタール高用量 群のCAR ノックアウトマウスでは,対照群と比較して有意に体重が減少した。全ての化合物 は,野生型マウスの肝臓重量を増加させた。野生型マウスでは,フルコナゾール低用量群を 除き,全ての化合物で低用量から用量依存的に有意な肝臓重量増加がみられた。野生型マウ スにおいて,シプロコナゾール高用量群が最も肝臓重量増加が顕著であった。CAR ノックア ウトマウスでは,シプロコナゾール及びフルコナゾール投与群は高用量のみで肝臓重量が増 加した。一方,テブコナゾール投与群では,CAR ノックアウトマウスにおいても,全ての投 与用量で肝臓重量が増加した。シプロコナゾール及びフルコナゾールの中・高用量群及びテ ブコナゾール低用量群では,野生型マウスに比較して,CAR ノックアウトマウスの肝重量が 減少していた。フェノバルビタールは,全ての投与用量においてCAR ノックアウトマウスに 肝重量を増加させなかった。摂餌量は,体重減少が認められた群で減少していた。化合物摂 取量を表2-2 に示す。 肝臓の病理組織学的変化及びALT 肝臓の病理組織学的所見及びALT の結果を表 2-3 に示す。野生型マウスでは,全化合物で, 投与用量に依存した小葉中心性肝細胞肥大が認められた (図 2-1,表 2-4)。シプロコナゾール 及びフルコナゾールは,中用量以上でCAR ノックアウトマウスにおいても肝細胞肥大を誘導 したが,その肝細胞肥大の程度は同群の野生型マウスと比較して弱かった。テブコナゾール を投与したCAR ノックアウトマウスでは,中用量以上では野生型マウスと同等の重篤な肝細 胞肥大が認められたが,低用量では野生型マウスと比較して肝肥大の程度が弱かった。CAR
28 ノックアウトマウスのフェノバルビタール投与群では,いずれの用量においても肝細胞肥大 は認められなかった。その他の病理組織学的所見として,シプロコナゾール投与群の野生型 マウス及びテブコナゾール投与群の両遺伝子型マウスでは,肝細胞の空胞化,単細胞壊死, 及び炎症性細胞浸潤が認められた。ALT は,野生型マウスでは中用量以上のテブコナゾール 投与群,シプロコナゾール及びフルコナゾールの高用量群,CAR ノックアウトマウスではテ ブコナゾールの高用量群で有意に増加した (表 2-3)。 肝臓におけるCYP 発現 抗CYP2B 抗体による免疫組織学的検査の結果を図 2-2 に示す。抗 CYP2B 抗体による免疫 組織学的検査では,野生型マウスにおいて,全化合物で低用量から用量依存的に,小葉中心 の肝細胞が強陽性を示した。CAR ノックアウトマウスでは,フェノバルビタールの全投与用 量で陽性反応が認められなかった。シプロコナゾール及びフルコナゾール投与群では,CAR ノックアウトマウスでも弱い陽性像が観察されたが,野生型マウスと比較して弱い反応であ った。テブコナゾール投与群では,CYP2B 陽性反応は,両遺伝子型で同等であった。 CYP 関連遺伝子の mRNA 発現量を図 2-3 に示す。野生型マウスでは,すべての化合物投与 群でCyp2b10 発現量の用量依存的な著しい増加が認められた。CAR ノックアウトマウスでは, いずれのトリアゾール系抗真菌剤も,高用量のみで Cyp2b10 発現量が増加した。一方,フェ ノバルビタール投与群では,CYP2B 蛋白発現と同様に,CAR ノックアウトマウスの全投与用 量で Cyp2b10 発現量が増加しなかった。フェノバルビタールを含む全ての化合物投与群の両 遺伝子型マウスで,Cyp3a11 発現量が用量依存的に増加した。Cyp3a11 発現量の増加は,いず れの化合物においても対照群と比較して5 倍未満であった。Cyp1a2 発現量は,野生型マウス の全ての化合物投与群において,用量依存的な軽微な増加が認められた。一方,CAR ノック アウトマウスでは,シプロコナゾール,フルコナゾール,及びフェノバルビタールの全投与 用量で Cyp1a2 発現量が増加しなかった。高用量のテブコナゾールだけが,CAR ノックアウ トマウスに有意にCyp1a2 を増加させた。テブコナゾール投与群では,統計学的有意差がない
29 群があるものの,Cyp4a10 発現量が増加傾向であった。一方,フルコナゾール及びフェノバ ルビタールは,Cyp4a10 発現量を減少させた。P450 reductase 発現量は,全化合物で用量依存 的に増加した。また,野生型マウスのP450 reductase 発現量は,いずれの群においても CAR ノックアウトマウスよりも高かった。 肝細胞増殖活性 肝細胞増殖活性の結果を図 2-4 に示す。肝細胞の増殖活性は,PCNA 陽性細胞の数として 評価した。野生型マウスでは,シプロコナゾール,テブコナゾール,及びフェノバルビター ルの中用量以上の群及びフルコナゾール高用量群で肝細胞増殖活性が増加した。一方,CAR ノックアウトマウスでは,全ての化合物投与群で対照群と同程度の肝細胞増殖活性であった。
30 考察 本実験では,トリアゾール系抗真菌剤であるシプロコナゾール,テブコナゾール,及びフ ルコナゾールをそれぞれ低,中,及び高用量の 3 用量群で混餌投与し,これらによる肝肥大 に関わる経路と投与用量の関係について検索した。 本実験において,CAR ノックアウトマウスで肝肥大が減弱したことから,いずれのトリア ゾール系抗真菌剤による肝肥大にも程度の差はあれ CAR が関与していることが明らかとな った。また,CAR ノックアウトマウスにおいても肝肥大を誘発したことから,いずれの化合 物による肝肥大にもCAR 以外の経路が存在した。一方,フェノバルビタールによる肝肥大で は,いずれの用量であってもCAR 以外の関与は認められなかったことから,本試験で用いた トリアゾール系抗真菌剤による肝肥大はフェノバルビタールとは異なっていた。本実験から 得られた結果に基づいた肝肥大におけるCAR あるいは CAR 以外の関与と投与用量の関係を 図2-5 に示す。肝肥大への CAR の関与は,その程度により,「完全なCAR 依存」,「主にCAR 依存」,「わずかなCAR の関与」の 3 段階に分けることができた。「完全なCAR 依存」は,フ ェノバルビタールによる肝肥大に認められた。シプロコナゾール及びフルコナゾールは高用 量のみでCAR 以外の影響がみられたことから,「主にCAR 依存」型に分類された。テブコナ ゾールによる肝肥大は,低用量でのみCAR の影響がみられたことから,「わずかなCAR の関 与」型であった。本章では,第 1 章では検出することができなかったテブコナゾールによる 肝肥大へのわずかなCAR の関与を明らかにすることができたことから,用量依存性の詳細な 解析が肝肥大に関与する核内受容体を明らかにするために有用であることを示した。同様に 用量依存性の解析の重要性については,いくつかの報告がなされている (2, 3, 11)。 野生型マウスにおいて,全てのトリアゾール系抗真菌剤投与群では,CAR の活性化を示唆 するCYP2B 蛋白及び Cyp2b10 発現量の増加が用量依存的に認められた。一方,両遺伝子型マ ウスにおいて Cyp3a11 発現量が増加したことから,全てのトリアゾール系抗真菌剤高用量群 では,PXR が関与していた可能性が考えられた。PXR と CAR のリガンドやアゴニスト・ア
31
ンタゴニストは,オーバーラップしていることが知られている (27, 52)。トリアゾール系抗真 菌剤においても,フルコナゾールを含む 4 つのトリアゾール系抗真菌剤がマウスの肝臓にお いてCAR 及び PXR の両経路を活性化することが報告されている (10)。加えて,Cyp2b10 よ りは増加の程度が弱いものの,Cyp1a2 も全てのトリアゾール投与群の野生型マウスで増加し
ていた。Cyp1a2 発現は主として AhR に関与している。しかし,これまでの報告で,CAR ノ
ックアウトマウスでは,Cyp1a2 が減少していたことから,CAR にも依存していることが示唆 されている (54, 57)。テブコナゾールは統計学的に有意に Cyp1a2 を増加させたが,その程度 が軽微 (1.88 倍) であることから,肝肥大への関与の可能性は低いと考えられた。一方, Cyp4a10 はテブコナゾールの投与により数~10 倍程度増加したことから,テブコナゾールに よる肝肥大には主としてPPARα が関与している可能性が示唆された。フルコナゾール及びフ ェノバルビタール投与群の野生型マウスにおいて,Cyp4a10 発現量が減少した結果は,CAR の活性化により Cyp4a 発現量が減少した過去の報告と一致していた (50)。CAR/PXR ダブル ノックアウトマウスから採取した初代培養肝細胞ではPPARα 及びその下流の Cyp4a の遺伝子 発現量が増加していた。CAR/PXR の非存在下ではフェノバルビタールは HNF4 を活性化する ことが知られており,PPARα 及び Cyp4a は HNF4 の標的遺伝子である (48)。これらの報告は, 本実験でCAR ノックアウトマウスのフェノバルビタール高用量群に Cyp4a10 発現量の増加が 認められたことに関連している可能性がある。 フェノバルビタールあるいは CAR アゴニストである 1,4-bis[2-(3,5)-dichoropyridyloxy] benzene (TCPOBOP) は,5-bromodeoxyuridine (BrdU) の肝臓における取り込みを CAR 依存的 に増加させる (51)。本試験においてもフェノバルビタールによる肝細胞増殖活性の増加は CAR ノックアウトマウスでは認められなかった。興味深いことに,本試験において,3 種の トリアゾール系抗真菌剤による肝肥大にはCAR 以外の関与が認められた一方で,肝細胞増殖 活性化はトリアゾール抗真菌剤も CAR 依存的であった。本試験で用いた投与濃度では CAR 以外の経路による肝細胞増殖活性は認められなかったが,フェノバルビタールは肝細胞増殖
32
活性よりも低い用量で肝肥大を誘発する (23)ことから,本試験で用いたトリアゾール系抗真 菌剤の用量よりも高用量では,CAR ノックアウトマウスにおいても肝細胞増殖の活性化がみ られるかもしれない。
33 小括 本実験では,シプロコナゾール,テブコナゾール,及びフルコナゾールによる肝肥大には CAR が関与していることが明らかとなった。シプロコナゾール及びフルコナゾールによる肝 肥大には主にCAR が関与していたが,テブコナゾールによる肝肥大には CAR はわずかしか 関与していなかった。また,トリアゾール系抗真菌剤による肝肥大では,PXR を含む CAR 以外の経路の関与が認められた。一方,フェノバルビタールによる肝肥大は完全にCAR に依 存していた。これらの結果から,3 種のトリアゾール系抗真菌剤による肝肥大は,フェノバ ルビタールとは異なる経路であることが示唆された。本章では,肝肥大への核内受容体の関 与をより詳細に解析するため,3 用量で肝肥大への影響を調べた。その結果,用量を増減さ せることで,テブコナゾールによる肝肥大にCAR の関与があること,フルコナゾールによる 肝肥大にCAR 以外の関与があることを新たに見出した。以上から,肝肥大における核内受容 体の関与を調べるためには,CAR ノックアウトマウスに化合物を複数用量投与して評価する 方法が有用であることが示された。
34 第3 章
35 緒言 イマザリルは,1970 年代より果実の収穫後農薬いわゆるポストハーベストに使用されてい る広域スペクトラムのイミダゾール系抗真菌剤である。雄性 Swiss マウスを用いたイマザリ ルの発がん性試験では,200 ppm 以上の投与濃度から肝細胞腺腫の発生頻度が増加した (20)。 また,イマザリルの投与によってマウス及びラットの肝重量が増加した (20)。さらに,イマ ザリルは,マウスの肝臓及び小腸に対して,CYP に依存した pentoxyresorufin O-dealkylase (PROD) 活性を増加させた (31)。マウス及びラットにおける PROD 活性の増加は CYP2B 誘 導と一致することが知られている (30, 32)。そのため,イマザリルはフェノバルビタールと同 様にCYP2B 誘導剤であると考えられている (4)。これらの実験結果から,イマザリルによる 肝肥大及び肝発がんにCAR が関与している可能性が示唆されるが,イマザリルによる肝肥大 及び肝発がんへのCAR の関与についての報告はない。 そこで本章では,第1 章と同様に,CAR ノックアウトマウスを用いた肝臓のイニシエーシ ョンプロモーションモデルによって,イマザリルによる肝発がんにCAR が関与しているか調 べた。また,第 2 章で確立させた複数用量投与による肝肥大への核内受容体の関与を調べる 評価系により,イマザリルによる肝肥大にCAR あるいは他の核内受容体が関与しているか調 べた。イマザリルは,農薬ガイドラインを満たした遺伝毒性試験により遺伝毒性がないこと が知られている (20)。
36
実験材料及び実験方法
化合物
本実験で用いたイマザリルの化学構造式を図3-1 に示す 。イマザリル (CAS No. 35554-44-0, 純度95.2%) 及び DEN (CAS No. 55-18-5,純度 99.9%) は,それぞれ LKT ラボラトリー社 (St. Paul, MN, USA) 及び東京化成工業株式会社 (Tokyo, Japan) より入手した。
動物 第1 章及び第 2 章と同様に,国立医薬品食品衛生研究所で繁殖している雄性 CAR ノックア ウトマウス及び日本チャールス・リバー株式会社より購入した野生型マウスを用いた。なお, 全ての動物実験は国立医薬品食品衛生研究所の動物実験委員会に承認を受け,国立医薬品食 品衛生研究所の動物実験の適切な実施に関する規定に準拠して行った。 実験方法 本実験は,実験1 と実験 2 に分けて実施した。実験 1 では,第 1 章と同様にイニシエーシ ョンプロモーションモデルを用いてイマザリルによる肝発がんへの CAR の関与を検討した。 また,同時にイマザリルによる肝肥大へのCAR の関与も併せて解析した。肝肥大及び肝発が ん性への継時的な変化を検索するため,6 週齢の雄性 CAR ノックアウトマウス及び野生型マ ウスを体重を基に無作為に群分けし4,13,及び 27 週投与群に割り付けた。各群の動物数は, 4 及び 13 週投与群は 5 匹とし,27 週投与群は 23 あるいは 24 匹とした。13 及び 27 週投与群 には,イニシエーション処理として6 週齢時に DEN を 90 mg/kg の用量で単回腹腔内投与し た。イマザリル投与群 (4,13,及び 27 週投与群) には,7 週齢より粉末基本飼料 (CRF-1, Oriental Yeast, Co. Ltd, Tokyo, Japan) にイマザリルを 500 ppm の濃度で混合した飼料で 4,13, あるいは27 週間混餌投与した。イマザリルの投与濃度は,過去の報告において肝肥大及び肝 発がんを誘発している投与濃度に基づいて設定した (20)。野生型及び CAR ノックアウトマウ
37 スの対照群には,粉末基本飼料を4,13,あるいは 27 週間自由摂餌させた。 実験2 では,イマザリルの肝肥大と用量依存性に対する CAR 等の核内受容体の関与を調べ ることを目的として実施した。7 週齢の野生型あるいは CAR ノックアウトマウスを各群 5 匹 で4 つのグループ (対照群,低・中・高用量群) に分けた。イマザリルは 500,1000,あるい は2000 ppm の投与濃度で 1 週間混餌投与した。投与濃度は,過去の報告 (31) を基に,投与 用量としてCYP2B を十分に活性化させる投与濃度を設定した。すなわち,10 mg/kg/day の 3 日間の投与でマウスの肝臓における PROD 活性が増加した (31) ことから,当該投与用量を 上回ることが期待される投与濃度として 500,1000,及び 2000 ppm の投与濃度を設定した。 対照群には粉末基本飼料を自由摂餌させた。 実験1 及び 2 に共通して,以下のように実施した。一般状態観察は 1 日 1 回以上,体重及 び摂餌量測定は1 週間に 1 回以上実施した。実験 1 の 13 週以降では,体重及び摂餌量は 2 週 間に 1 回測定した。計画剖検時において,全身深麻酔下で採血し,マウスを安楽死させた。 血液サンプルは,血清に分離し,血清は-30℃で保存した。ALT は,株式会社エスアールエル (Tokyo, Japan) で測定した。肝臓は,肉眼的観察後,重量を測定 (実験 1 の 27 週投与群を除 く) し,10%中性ホルマリン緩衝液で固定した。 肝臓の病理学的及び免疫組織化学的評価 第1 章及び第 2 章と同様に,肝臓の中間葉 (実験 1 の 27 週投与群を除く) を用いて HE 染 色,免疫組織化学的染色を実施した。免疫組織化学的染色には,抗 CYP2B 抗体 (Enzo Life Sciences Inc., Farmingdale, NY, USA) 及び抗 PCNA 抗体 (PCNA, Clone PC10, DAKO, Glostrup, DNK) を用いた。
実験1 の 27 週投与群では,第 1 章と同様の方法で肝発がん性について解析した。すなわち, 変異肝細胞巣及び肝腫瘍を数えるため,肝臓の中間葉及び左葉をそれぞれ 6 切片ずつ作製し た。肝臓の増殖性病変は,好酸性,好塩基性,その他 (明細胞性及び混合型) のいずれかに分 類し,発生頻度及びmultiplicity について計算した。
38 RT-real time PCR
実験2 では,肝臓の左葉を用いて,第 1 章及び第 2 章と同様の手法により RT-real time PCR を 用 い て 遺 伝 子 発 現 量を 評 価 し た 。 測 定 対 象は ,Cyp1a2 (Mm00487224 m1) ,Cyp2b10
(Mm01972453 s1),Cyp3a11 (Mm00731567 m1),Cyp4a10 (Mm01188913 g1),及び P450 reductase (Mm00435876 m1) として,GAPDH で標準化した発現量によって比較した。 肝細胞増殖活性の解析 投与1 週 (実験 2),4 週,及び 13 週 (実験 1) において,第 1 章及び第 2 章と同様に,PCNA 陽性肝細胞のグレーディングを実施した。細胞増殖活性は,1 視野あたりの PCNA 陽性細胞 数に基づいて,グレード0~3 で評価した。 統計解析 実験1 では,対照群とイマザリル投与群の比較を CAR ノックアウトマウス及び野生型マウ スについてそれぞれ実施した。また,CAR ノックアウトマウスと野生型マウスの比較をそれ ぞれの同じ投与群同士で比較した。体重,肝臓絶対重量,肝臓相対重量,及び肝臓増殖性病 変の multiplicity のデータについて,F 検定により等分散性の検定を実施した.等分散性が認 められた場合はStudent の t 検定を,等分散性が認められなかった場合は Welch の t 検定を用 いて,それぞれ媒体対照群と化合物投与群との間で比較した。肝臓増殖性病変の発生頻度の データは,Fisher の直接確率検定によって統計解析した。有意水準は 5%及び 1%とした。 実験 2 では,対照群と各用量のイマザリル投与群の比較を各遺伝子型のマウスの間で,そ れぞれ実施した。また,CAR ノックアウトマウスと野生型マウスの比較をそれぞれの同じ投 与用量同士で比較した。対照群とイマザリル投与群の比較はDunnett 検定 (等分散性あり) あ るいはSteel 検定 (等分散性なし) を用い,野生型と CAR ノックアウトマウスとの比較では, Student の t 検定 (等分散性あり) あるいは Welch の t 検定 (等分散性なし) を用いた。統計解
39
析は,体重,肝臓絶対重量,肝臓相対重量,ALT,及び mRNA 発現量のデータを対象とした。 有意水準は5%及び 1%とした。