膜の構造と機能
慶應義塾大学環境情報学部・基礎分子生物学
3
平成22
年11
月24
日本テキストを利用するに当たっての注意
本テキストは慶應義塾大学湘南藤沢キャンパス(SFC)で行われている基礎分子生物学3の履修者 有志によって作成されたものです。生物学の専門家によって作成されたものではありませんので、
重要なポイントについては元となった書籍「Essential細胞生物学(原著第2版)」[1]でしっかり確 認して下さい。
ただ学生が作ったものとはいえ、独自に作成した豊富な図を用いて初学者の視点から分子生物学 を分かりやすく解説することを試みています。
また本テキストはウィキ(Wiki)方式で毎年履修者によって改良され、どんどん良い物になってゆ くでしょう。本テキストを受動的に利用するのではなく、ぜひ執筆活動に積極的に参加する姿勢で 利用して頂きたいと思います。
Saito R.
目 次
1 膜の役割と基本構造 4
2 脂質二重層の流動性と非対称性 4
3 膜タンパクの構造と膜への結合 8
4 膜と細胞表面 8
5 膜輸送の原理 10
6 イオンチャネルのイオン選択性 16
7 神経細胞のイオンチャネルとシグナル伝達 16
8 細胞の区画とタンパク質の輸送 20
9 糖と脂肪の分解 27
10 ミトコンドリアの4つの区画とその役割 29
11 クエン酸回路と電子伝達系 30
12 食物の備蓄と利用 32
13 葉緑体の構造と機能 34
14 光合成の仕組みとその進化 36
1
膜の役割と基本構造膜は細胞の内容物を入れる容器の役割を果たすもので、細胞が生きてゆく上で欠かせない物質で す。化学的には膜の正体は脂質二重層であり、厚さは約5nmです1。細菌では細胞膜が唯一の膜で すが、真核生物には細胞内区間を仕切る細胞内膜も存在します。細胞膜には多数の膜タンパク質が 存在しており、
1. 無秩序な細胞外部液の流入、細胞内部液の流出の防止 2. 栄養物の取り込み
3. 老廃物の排出
4. 外界の刺激に対する応答 5. 細胞の運動
を行うことが可能です。
親水性の頭部
疎水性の尾部
図 1: 膜を作る脂質分子 図1に示すように、脂質二重層は親水性の頭部と、疎水性の
尾部でできています(両親媒性)。尾部の炭化水素間に二重結合 がある部分では、尾部が折れ曲がります。親水性の頭部は水に 引き寄せられ、疎水性の尾部は水を避けて他の疎水性分子と会 合しようとする性質があります。この2つの性質をちょうど満 たす構造が脂質二重層で、頭部が外側に出て水分子と接してお り、尾部が内側に入っています(図2)。脂質二重層はそれぞれ が引きつけ合って形成されるというよりむしろ、疎水性の尾部 が周りの水分子と結合を持てないことによって結果的に形成さ れています。脂質二重層に末端があると、尾部が水分子と接し てしまうことになるので、それを避ける構造、つまり丸まった 構造が安定したものになり、これがまさに細胞膜が丸まる原理
です。生体内で最も多く存在する膜脂質はリン脂質であり、親水性の頭部とそれ以外の部分がリン 酸基を介して結合しています。
2
脂質二重層の流動性と非対称性細胞膜中の個々の分子の並び順はほぼ一定というイメージがあるかも知れませんが、実際には細 胞膜を構成する脂質分子は自由に動きまわることができます2。脂質分子がどの程度の速さで動き回 れるかは温度などに依存しますが、場合によっては、大型の細胞膜全体(2µm)にほぼ1秒で拡散す ることが可能なのです。また脂質分子は細胞質→細胞外の方向を軸として回転することが可能で、
速いものは毎秒30,000回転にも達します。しかしながら、細胞質側の脂質分子が細胞外側に移った
15×10−9m、原子ほぼ50個分に相当する。
2単粒子追跡(SPT顕微鏡観察法)を用いると、個々の分子あるいは小集団の分子を標識して追跡することができる。
水
図2: 脂質二重層
細胞小器官
小胞
細胞質
細胞外液
図3: 膜の合成
り、また逆に細胞外側の脂質分子が細胞質側に移ったりする現象(フリップ・フロップ)は自発的に はほとんど起こりません。リン脂質分子の流動性を決める要因はいくつかあります。まずは炭化水 素鎖の長さで、 短いとそれ同士の相互作用が少なくなり、流動性が増します。次に、炭化水素尾部 の二重結合の数(不飽和度)で、これが多いと、尾部をきっちり並べるのは難しくなり、流動性が増 します。そして、コレステロールは不飽和炭化水素鎖の尾部の折れ曲がりによって生じたリン脂質 分子間の隙間を埋め、流動性を低下させます。
新しい膜の合成はほとんど小胞体(ER, 8節も参照のこと)で行われ、細胞小器官から出芽した小 胞が細胞質を移動し、細胞膜に取り込まれます(図3)。このとき、細胞質に向いている側の脂質層は 常に一定です。膜の合成過程では、小胞体の内腔にて、脂質はまず膜の細胞質側に取り込まれ、フ リッパーゼによって一部は細胞外部側へ運ばれます。フリッパーゼにはリン脂質の選択性があるた め、脂質二重層の非対称性が生まれます(図4a)。ゴルジ体内の酵素群は細胞質と接していない側の 膜だけに糖残基を付加するため、これも非対称性を生む要因となります(図4b)。
細胞外部
細胞質
(a) (b)
図4: 脂質分布の非対称性
(a)フリッパーゼのリン脂質選択性によって生まれる非対称性、(b)ゴルジ体内の酵 素群が細胞質と接していない側の膜だけに糖残基を付加するために生まれる非対称性。
6角形が糖残基を表す。
X Y 細胞外部
細胞質
(a) (b)
(c) (d)
図 5: 膜タンパクの様々な機能
膜タンパク質が持つ機能のうち、主なものを4つ図示した。(a)輸送体、(b)連結体、(c)受 容体、(d)酵素。
N末端
C末端 膜貫通型
単層結合型
脂質連結型
タンパク質付着型
図6: 膜タンパク質の膜への結合様式
3
膜タンパクの構造と膜への結合膜タンパク質は膜に結合したタンパク質、もしくは膜と連動しているタンパク質を指し、膜の機 能のほとんどは膜タンパク質が担っています。膜タンパク質の機能は図5に示すように主に、細胞 内外への分子の輸送(細胞外へNa+を運び出し、細胞内へK+を運び入れるNa+ポンプなど)、他 のタンパク質の膜付近への固定(アクチンフィラメントとマトリックスをつなぐインテグリンなど)、
細胞外からのシグナルの受容(PDGFを結合し、シグナル伝達によって細胞の成長、分裂を起こす 血小板由来増殖因子受容体など)、そして代謝(外のシグナルに応じて環状AMPの生産を触媒する アデニル酸環化酵素など)です。
図6に示すように、膜タンパク質の膜への結合様式は様々です3。タンパク質の膜貫通領域では、
疎水性の側鎖が同じく疎水性の脂質の尾部と接触し、親水性の主鎖が互いに水素結合を作ると安定 した立体構造になります。親水性の主鎖の結合数はタンパク質がαへリックスという構造をとると 最大になります。このため、膜貫通型のタンパク質にはαへリックスが多いことが知られています。
4
膜と細胞表面細胞膜自体は極めて薄く破れやすい構造になっているので、ほとんどの細胞膜は膜貫通タンパク を介して膜に付着しているタンパク質で出来た枠組みで支えられています。細胞膜の力学的な性質
3ここに示してある例は内在性膜タンパク質。界面活性剤を用いて脂質二重層を壊さなければ膜から離れない。一方、表 在性膜タンパクは脂質二重層を壊さなくてもタンパク質間の相互作用だけを変える抽出処理によって膜から分離できる。
脂質分子と界面活性剤分子
細胞膜に最も多く使われているホスファチジルコリンというリン脂質分子は、親水性の頭部と 疎水性の尾部を持った両親媒性の分子です。界面活性剤分子も基本的な構造は同じですが、こ の2つは疎水性の尾部の構造が異なっています。リン脂質分子は疎水性尾部が2本の鎖状アル キル基からなっているのに対し、界面活性剤分子は1本しかありません。図形的に考えるとリ ン脂質分子は頭部と尾部の横幅がほとんど同じ円柱状の分子です。一方界面活性剤分子は親水 性頭部に比べて疎水性尾部の横幅が短いので、頭部を底面とする円錐状の分子です。水中では 円柱状のリン脂質分子の疎水性尾部は水を避ける傾向があり、その結果として二重層を形成し て安定した構造を保ちますa。細胞質ゾルには水が含まれるので細胞内での振る舞いもほぼ同 じです。一方、界面活性剤分子は同じように疎水性尾部を水から隠すように結合すると、ミセ ルと呼ばれる球体を形成します。リン脂質分子の疎水性尾部の片方を切断し一本にするか親水 性の頭部を大きくすると、円錐型の立体構造に変わり、界面活性剤として働くようになります。
親水性
疎水性
単純な図形的イメージ
膜の脂質分子 界面活性剤分子
リン脂質分子 界面活性剤分子
構造 疎水性尾部が2本 疎水性尾部が1本 水中状態 疎水性尾部を内側に集めた二重層を
作る
疎水性尾部を中心に集めたミセルを 作る
性質 疎水性尾部を完全に水から隔離する 膜 を作る
疎水性尾部を他のタンパクや油脂に 結合させ、 洗剤 としてはたらく
a親水性頭部には極性があるので水分子と水素結合を形成するが、疎水性尾部は無極性なので、ファンデルワール ス力のみで結合している。
スペクトリン アクチン 膜貫通タンパク
~100 nm
付着タンパク
図7: ヒト赤血球の細胞皮層
は、繊維状のタンパク質が作る細胞皮層と呼ばれる網目構造で決まります(図7)。皮層の主成分は、
スペクトリンという細長くて柔軟な、長さ約100 nmの棒状タンパクです。スペクトリンの作る網 目構造が細胞膜を支え、細胞を平たい形に保っているのです。細胞膜にスペクトリンをつないでい るのは細胞内付着タンパクで、これがスペクトリンを特定の膜貫通タンパクに結びつけています。
赤血球のスペクトリンに遺伝的な異常が見られるマウスやヒトは貧血症状を呈し、赤血球細胞の数 が正常より少なく、形も扁平ではなく球状で極めて壊れやすくなっています。ヒトのほとんどの細 胞の細胞皮層には、スペクトリンとその付着タンパクに似たタンパク質がありますが、それは赤血 球細胞のものより遥かに複雑です。これは赤血球は血管内を押し流される際の圧力に耐える力学的 な強度があれば十分なのに対し、他の細胞は活発に形を変えたり移動したりしなければならないた めです。
真核細胞の細胞表面の脂質や膜タンパクには多くの場合、オリゴ糖が結合しており、これらは糖 衣という被覆を作っています(図8)。オリゴ糖は水を吸収するので、細胞表面はぬめぬめしていて、
潤滑剤の役割を果たしています。これによって細胞表面が機械的、化学的な損傷を受けるのを防い でいるのです。また糖衣は細胞同士の識別や接着にも重要な役割を果たしています。レクチンと呼 ばれる一群のタンパク質は、特定のオリゴ糖側鎖を識別してそれに結合します。
5
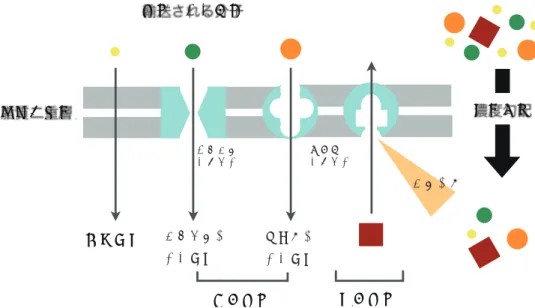
膜輸送の原理細胞の持つ細胞膜は脂質二重層なので、極性のある大きな分子やイオンを拡散によって細胞内に 取り込むことはできません。実験的に脂質二重膜のみを作った場合、水溶性の物質は膜をほとんど 通過できないでしょう。そこで、細胞が生きていくうえで必要な物質は細胞膜上に存在する膜輸送 タンパクを利用して細胞内に輸送されます(図9)。
膜輸送タンパクにはチャネルタンパクと運搬体タンパクの2種類があります。チャネルタンパク (図10)はランダムに開いたり閉じたりを繰り返すタンパク質で、分子の持つ電荷とその大きさで通 過できる分子を選択します。
細胞質
脂質二重層 糖衣
細胞外部 糖残基
膜貫通タンパク質 糖脂質
膜貫通プロテオグリカン 表面に吸着された
糖タンパク
図 8: 細胞表面に存在する糖衣
人工脂質二重層 疎水性の小分子
電荷を持たない 極性小分子
大型で電荷を持 たない極性分子
イオン
O2 CO2 N2 ベンゼン H2O グリセリン エタノール アミノ酸グルコース ヌクレオチド H+, Na+, HCO3-,K+ Ca2+,Cl- Mg2+
(a)
(b) (c)
図9: 人工脂質二重層と細胞膜の違い
(a) 人工脂質二重層を通って分子が拡散する速度は分子の大きさと溶解度に依存す る。(b)タンパク質を含まない人工脂質二重層は水溶性分子をほとんど通過させない。
(c)細胞膜にある輸送タンパクはそれぞれ特定の分子だけを運搬する。このため、膜内 の区画には選ばれた溶質だけが蓄積する。
開 閉
イオンなど
図 10: チャネルタンパク
図 11: 運搬体タンパク
⢽⾰ੑ㊀ጀ Ớᐲ൨㈩
න⚐ᢔ
ャㅍߐࠇࠆಽሶ
࠴ࡖࡀ࡞ߦ ࠃࠆᢔ
࠴ࡖࡀ࡞
࠲ࡦࡄࠢ
ㆇ៝
࠲ࡦࡄࠢ
ㆇ៝ߦ ࠃࠆᢔ
ࠛࡀ࡞ࠡ
ฃേャㅍ ⢻േャㅍ
図12: 受動輸送と能動輸送
運搬体タンパク(図11)は、タンパク質の一部に分子が結合するとタンパクが変形し、それによっ て内外への輸送を行います。その機構の性質上、輸送できる分子に選択性があります。
輸送には、エネルギーを必要としない受動輸送と、その逆の能動輸送があります(図12)。溶質は その濃度勾配と膜をはさんだ電位差に従って自発的に移動します。この2つの駆動力を電気化学的 勾配と呼びます。受動輸送では電気化学的勾配を利用して輸送を行っており、エネルギー消費は起 こりません。例えるならば、坂道の頂上に置いたボールが坂のふもとへ自然に転がっていくように 溶質が輸送されるのです。一方、運搬体タンパクによって何らかのエネルギーを輸送に用いなけれ ばならないのが能動輸送です。これは先ほどの例に例えるなら、坂道のふもとに置いたボールを労 力を使って坂の頂上にもっていくように溶質を輸送するのです。つまり能動輸送は電気化学的勾配 に逆らって溶質を輸送します。これには以下に挙げる方法があります。
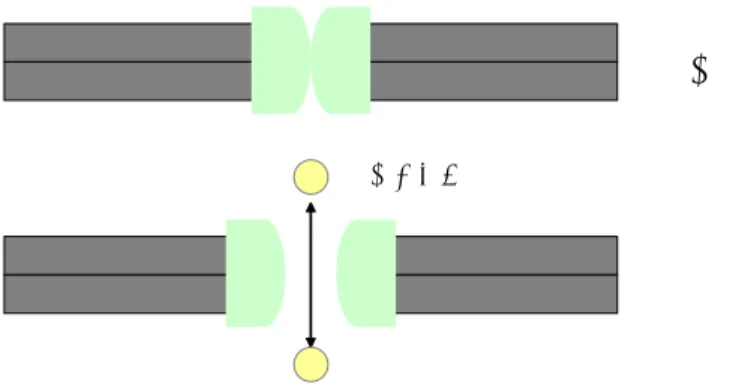
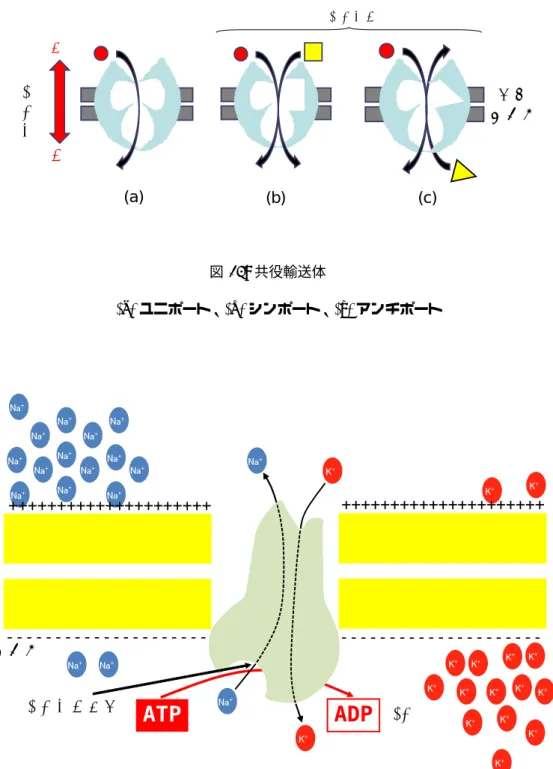
共役輸送 ある溶質の勾配に逆らう方向の輸送と別の溶質の勾配に従う方向の輸送とを組み合わせ る方法です。2種類の溶質が同じ方向に輸送されるものをシンポートといい、その逆をアンチ ポートといいます(図13)。また、共役輸送ではありませんが、1つの溶質のみで行われるも のをユニポートといいます。
ATP駆動ポンプによる輸送 ATP→ADP + Piの過程で得られるエネルギーを利用する方法です (ATPについては9節で説明します)。人体のNa+ポンプはこれにあたります。
光駆動ポンプによる輸送 細菌細胞等が使う方法で光エネルギーを利用して勾配に逆らう輸送を行う 場合をいいます。
図14にATP駆動ポンプの一種であるNa+-K+ポンプが機能する様子を示します。このポンプは 以下のステップでNa+を細胞外にK+を細胞内に運びます。
濃 度 勾 配
高 低
脂質 二重層
(a) (b) (c)
共役輸送体
図13: 共役輸送体
(a) ユニポート、(b)シンポート、(c)アンチポート
Na+ Na+ Na+
Na+ Na+
Na+ Na+
Na+ Na+
Na+ Na+
Na+ Na+
Na+ Na+ Na+
K+ K+
K+ K+
K+ K+ K+ K+
K+
K+ K+
K+ K+ K+ K+
Na+の結合部位
+++++++++++++++++++++ +++++++++++++++++++++
--- ---
Na+
K+
ATP ADP
+Pi細胞質側
Na+
K+
図 14: Na+-K+ポンプの機能
1. Na+がチャネルに結合
2. ATPの加水分解(ATP+H2O→ADP+Pi)
3. リン酸基(Pi)がチャネルに結合してリン酸化され、チャネルの構造が変化する →Na+が細 胞外に放出される
4. 細胞外のK+がチャネルに結合して脱リン酸され、K+は細胞内へ流入する
運搬体タンパクは選択制が極めて高いため、多くの場合、1種類の分子しか運びません。そのた め、膜で包まれた各々の細胞小器官には、必要な分子を取り込むための運搬体タンパクが独自の組 み合わせで備わっているのです。
6
イオンチャネルのイオン選択性イオンを透過させる働きのあるチャネルをイオンチャネルと呼びます。イオンチャネルには選択 性があります。例えばNa+チャネルはNa+イオンしか通さず、K+チャネルであれば、K+イオン しか通しません。このように特定の無機イオンしか通さない細胞膜表面の小さな穴が、イオンチャ ネルの実体です。イオンは膜の外と内のイオン勾配差によって起きる受動輸送によって運ばれます。
イオンチャネルがイオンを特異的に通す仕組みとしてまず挙げられるのが、イオンの直径や形で す。穴に合わない形のイオンや穴より大きいイオンは通りません。次にチャネル内の電荷の分布で す。例えばチャネル内が+に電荷している場合、+の電荷を帯びたイオンは反発しあうため、通る 事ができません。
また図15に示すように、チャネルの一番狭いところでは、選択フィルターと呼ばれる、関所が存 在し、イオン選択性を助けています。例えば、K+イオンチャネルでは、チャネルの内側に負の電 荷を帯びたカルボニル基が並び、K+イオンが一時的に結合する部分を作ります。またイオンはた いてい水分子をまとっていますが、水分子をつけたままこの狭いフィルターを通る事は不可能なた め、水分子をほとんど外してこのフィルターを通ります。イオン輸送のこの段階がチャネル通過の 最大速度を制限しています。したがってイオン勾配の差が増大するとチャネルを通るイオンの流れ は勾配に比例して増加しますが、このフィルターの働きによって速度には上限ができるのです。
7
神経細胞のイオンチャネルとシグナル伝達ニューロン(神経細胞)はシグナルを受け取って次の目的地に運ぶ重要な細胞です。多くの動物は 神経系を持っており、どの神経系も、数多くのニューロンからなるネットワークなのです。図16に 示すように、ニューロンには核を含む細胞体と、細胞体から遠く離れた細胞までシグナルを伝達する 長い軸索、他のニューロンからのシグナルを受け取る表面積を増やすための樹状突起、他のニュー ロン、筋肉などたくさんの細胞にシグナルを伝達するためたくさんの枝に分かれた神経末端から構 成されています。
細胞外を基準とした細胞内の電位を膜電位と呼びます。ちなみにネルンストの式を使うと、細胞 内外のイオン濃度の比から理論上の静止膜電位を計算することが可能です(コラム参照)。静止状態
K+
o o o
o o o カルボニル
カルボニルカルボニル カルボニル基基基基
K+
K+
K+
K+
K+
K+ K+
K+
K+
水分子 水分子 水分子 水分子
図15: K+イオンの選択フィルター
チャネルの一番狭いところには選別の関所(選択フィルター)が存在する。
樹状突起
軸索
細胞体
神経末端
図 16: 神経細胞(ニューロン)
ネルンストの式
以下のネルンスト(Nernst)の式は平衡状態を定量的に表すもので、これを用いると、細胞内外 のイオン濃度の比から理論上の静止膜電位の値を計算できます(イオンが1個の正電荷をもち、
温度が37℃のとき)。
V = 62 log10(Co/Ci)
ここで、V はミリボルト(mV)で表した膜電位で、Co/Ciは細胞外/内のイオン濃度を表しま す。この式の導出過程を下図の膜を間に挟んだ2種類の溶液を使いながら、説明します。
C1 C2 膜
濃度C1の溶液中から1 molの物質を濃度C2の溶液中に輸送するときの自由エネルギー変化
∆Gは、
∆G=RTln(C2/C1)
です(自由エネルギーに関してはp.33のコラム参照)。ここで、Rは気体定数(8.314J/K·mol) でTは絶対温度です。輸送はC1=C2、すなわち∆G= 0となるまで続きます。
そして、細胞膜の両側で生じている電位差、すなわち膜電位を式に組み込むと、
∆G=RTln(C2/C1) +ZF∆Ψ
となります。ここで、Zはイオンの電荷、Fはファラデー定数(96.5 kJ V−1mol−1=23.06 kcal V−1mol−1)、∆Ψはボルト(V)で表した膜電位です(方向は外→内とする)。ZF∆Ψは1 mol のイオンが電位差を超えて移動する時のエネルギー変化を意味します。静止膜電位は細胞内外 で電気化学的勾配が0になっている状態における膜電位なので、∆G= 0のときの∆Ψを求め ればいいことになります。これは∆G=RTln(Ci/Co) +ZF∆Ψより
∆Ψ = ln(Co/Ci)·RT /ZF
となります。従って、37℃(T = 310)で1価のイオン(Z =±1)という条件では、
∆Ψ≃ ±27 ln(Co/Ci)≃ ±62 log10(Co/Ci)
となります。ネルンストの式は数種類のイオンを同時に扱うことはできません。そこで異なっ たイオンの膜に対する相対的な透過性を考慮したゴールドマンの式があります。
0 1 2 0
-50 -50 膜電位(mV) (a)
(b)
時間(ms) 刺激電流
(mV)
閉 開 不活性 閉
(c)
図17: 電位依存Na+チャネルと膜電位
(a)短いパルス電流。これが活動電位を発生させる。(b)刺激電流が流れた後の膜電位の変化。
(c) Na+チャネルの状態。
のニューロンでは、軸索の外側は内側に比べ正に荷電しており、これを分極しているといいます。静 止状態で分極が起こっているのは、K+漏洩チャネルによって細胞内に多く存在するK+が細胞外 に流出し、結果として負の電荷を持つイオンの割合が増えているためです。実際に電位差の測定を 行うと、細胞膜の内側が外側に対して-20mVから-200mV程度の電位を持っていることが分かりま す。このように観測される電位差を静止膜電位といいます。ニューロンによる情報伝達は、この膜 電位の変化の電気信号によって行われます。電気信号は減衰して消えてしまわないように所々で増 幅されながら伝えられますが、この電気的興奮が伝わる波を活動電位といいます。以下では活動電 位が生じて機能する仕組みを説明します。
図17に示すように、電位依存Na+チャネルは閉、不活性、開の3種類の形状をとります。ニュー ロンが刺激を受けると電位依存Na+チャネルが開き、細胞外のNa+が細胞内に流入し、膜電位の 負の値がわずかに正に近づきます。このような膜電位の変化を脱分極といいます。これによって細 胞内と細胞外の電荷が逆転する活動電位を生じます。そしてチャネルはすぐさま不活性となります。
これにより刺激の直後、再び刺激を受けてもNa+チャネルが不活性状態から閉状態に戻るまでの 間、次の活動電位を発生する事はできません。
活動電位はすぐさま軸索の膜上のNa+チャネルに沿って隣へ隣へと伝えられます。刺激がある一
Ca2+イオン
シナプス前細胞 シナプス後細胞
神経伝達物質 電位依存
Ca2+チャネル
イオン
神経伝達物質受容体 ++++++---+
---+++-
---+++- ++++++---+
--+++++++
++---
++--- --+++++++
図 18: ニューロンが他のニューロンへ情報を伝える仕組み
定の値(閾値)を超えると活動電位を生じますが、その値を超えなければ、静止膜電位に戻ります。
あるニューロンが他のニューロンへ情報を伝える仕組みを図18に示します。シグナルを伝達する 細胞(シナプス前細胞)と受け取る細胞(シナプス後細胞)の間にはシナプスと呼ばれる特別な接 合部があります。ニューロンがニューロンへ情報を伝えるためには神経伝達物質がシナプスを通る 必要があります。神経伝達物質は神経末端でシナプス小胞に蓄えられています。神経末端にシグナ ルが電気信号として到着すると、電位依存Ca2+チャネルが開き、Ca2+イオンが細胞内に流入しま す。これをきっかけにシナプス小胞の中に含まれる神経伝達物質が化学信号として細胞外に分泌さ れます。分泌された神経伝達物質は神経伝達物質受容体によって受容されます。受容体が働くとイ オンが細胞内に流入し、シグナルが伝達されます。この際、化学信号は再び電気信号に変換される ことになります。神経伝達物質には興奮を伝えるアセチルコリン、グルタミン酸などの興奮性伝達 物質と、興奮を抑えるGABA、グリシンなどの抑制伝達物質が存在します。興奮伝達物質はNa+ やCa2+ などを受容するイオンチャネルに結合し(図19)、抑制伝達物質はCl−などを受容するイオ ンチャネルに結合します。
シナプスでは、複数の化学信号が加算されることや、シナプスでの化学的活動がそのシナプスの 強度を変えることがあります(シナプス可塑性)。シナプスでの化学的な信号伝達は、一見神経系の 活動を非効率にするようにも見えますが、このようなニューロンによる演算や記憶を容易にすると いう重要な側面を持っています。
サリンは副交感神経毒です。受容体によって受容されないで余ったアセチルコリンを分解するコ リンエステラーゼの合成を阻害し、細胞内にアセチルコリンがたまったままにすることで、副交感 神経を興奮させたままにする働きがあるのです。
8
細胞の区画とタンパク質の輸送バクテリアの細胞には細胞質(cytosol)という細胞膜で囲まれた区画が1つあるだけですが、真核 生物の細胞の内部は図20に示すように、細胞内膜によって複雑に分割され、特定の機能を果たすよ
䜰䝉䝏䝹䝁䝸䞁 - -
- - - - - -
- - - - - - - -
Na+
䜰䝉䝏䝹䝁䝸䞁
⤖ྜ㒊
(a) (b)
(c)
-- --
--- -
⣽⬊㉁
⬡㉁㔜ᒙ
- -
Na+
図 19: アセチルコリン受容体の構造
筋細胞膜にあるものを例として示した。アセチルコリンがチャネルに結合すると開く。小孔の 両側には負電荷を持つアミノ酸側鎖があり、正電荷を持つイオン(主にNa+やK+)が通過でき る。(a)全体の構造、(b)閉状態、(c)開状態
区画 主要機能
細胞質 多数の代謝経路、タンパク質合成系 核 ゲノムがある、DNAとRNAを合成する
小胞体 脂質のほとんどを合成、多数の細胞小器官や細胞膜に送るタンパク質の 合成
ゴルジ体 タンパク質と脂質を修飾・選別して、分泌や別の細胞小器官に送る小胞 に詰め込む
リソソーム 細胞内消化
エンドソーム エンドサイトーシスにより取り込まれた物質の選別 ミトコンドリア 酸化的リン酸化によるATPの合成
葉緑体(植物細胞) 光合成によるATPの合成と炭素固定
ペルオキシソーム 有毒分子の酸化
表 1: 真核細胞に存在する膜で包まれた小区画の主要機能
ヤリイカを用いて膜の興奮の秘密を探る
ヤリイカの持つ軸索は太くて長いため、神経の軸索を伝わる電気シグナルの研究をするのにヤ リイカは都合のよい生物です。神経の興奮状態を知る上で実際にアメリカケンサキイカが研究 に使われていました。その巨大な軸索に毛細管電極を挿し、刺激を与えることで活動電位を直 接測ることができます。そのため、軸索の中を空にした後、そこに軸索の内外に存在する主な イオンの純粋溶液で満たす灌流を行うことで、活動電位に関係しているイオン(Na+とK+)を 同定することができたのです。またそれぞれのイオンがどのような働きをしているかも、イカ の巨大軸索によって明らかになりました。また膜外のK+とNa+の濃度を変化させ、それが膜 電位にどのような影響を与えたのかを調べた結果、Na+は活動電位のピークに、K+は静止状 態の軸索の膜電位に関係していることが分かりました。さらに、活動電位によってNa+チャネ ルが開くこと、Na+チャネルが閉じた後は余分のK+チャネルが開くことによって膜電位が静 止状態に戻るのを促進していることも判明したのです。
軸索の細胞質
灌流用カニューレ
灌流液 灌流液 ゴムのローラー 巨大軸索
ヤリイカの軸索内の細胞質をゴムのローラによって取り除き、カニューレを用いて灌流 液に置き換える操作。
ミトコンドリア
核 細胞質
小胞体 ゴルジ体 細胞膜
ペルオキシソーム
リソソーム
エンドソーム
図20: 小腸上皮細胞の模式図
真核生物の場合、このように細胞内に多様な区画が存在する。小さな黒い粒はリボソームを表 す。リボソームには膜に結合しているものもあれば、遊離しているものもある。
核膜孔を通る輸送
膜を通る 輸送
小胞による 輸送 細胞質で合成されたタンパク質
葉緑体
ミトコンドリア
ペルオキシソーム
図21: 細胞小器官のタンパク質の取り込み
うに専門化された構造、すなわち細胞小器官(organelle)が見えます。その主な機能を表1にまと めます。
タンパク質の合成はほとんど細胞質で行われます4。その中で細胞質に留まるタンパク質もあれ ば、他の区画に輸送されるタンパク質もあります。後者のタンパク質の場合、図21に示すような3 種類の方法で輸送が行われます。どの輸送機構でもエネルギーを必要としますが、膜を通る輸送で はさらにタンパク質の立体構造をほどく必要があります。
水溶性のタンパク質が小胞によって輸送される場合、そのタンパク質は翻訳終結前に小胞体の中 に入りこみます。そして図22に示すように、合成されたタンパク質は輸送小胞の出芽・融合の繰り 返しによって他の細胞小器官や細胞膜へと輸送されます。小胞による輸送経路は、小胞体からゴル ジ体を介して細胞膜を抜けて外へ向かうものと、細胞膜からエンドソームを介しリソソームという ように内に向かうもの(エンドサイトーシス経路)があります。
膜から出芽する小胞は細胞質側の表面に独自の被覆を持ちます。最も研究が進んでいるのはクラ スリンというタンパク質の被覆です。それぞれの区画から出芽する輸送小胞が正しく働くためには、
まず輸送の対象となる小胞体中のタンパク質が小胞に詰め込まれなければなりませんが、アダプチ ンというタンパク質が輸送対象のタンパク質の選別を行い、クラスリンとともに小胞を覆う被覆を 形成するのです。小胞の出芽後は被覆は離脱します。
その後、小胞は標的となる膜とだけ正確に融合しなければなりません。小胞の特異的融合はSNARE とよばれる一群の膜貫通タンパクの働きによるものです。図23に示すように、小胞に存在するv- SNAREを、標的膜にある相補的なt-SNAREが識別するのです。
核核 核核
ゴルジ ゴルジ ゴルジ ゴルジ体体体体
細胞外部 細胞外部細胞外部 細胞外部
小胞体 小胞体小胞体 小胞体
リソソーム リソソームリソソーム
リソソーム 後期後期エンドソーム後期後期エンドソームエンドソームエンドソーム 初期 初期 初期
初期エンドソームエンドソームエンドソームエンドソーム
輸送小胞 輸送小胞 輸送小胞 輸送小胞 核膜
核膜 核膜 核膜
細胞質 細胞質細胞質 細胞質
図 22: 小胞による輸送
細胞外に向かう経路を赤い矢印で、エンドサイトーシス経路を緑色の矢印で示す。
v-SNARE
v-SNARE
t-SNARE
t-SNARE
融合 接着
積み荷b 積み荷a
積み荷受容体
被覆
他の細胞区画
小胞
図 23: SNAREによる標的膜の認識
O P O P
O O
O-
O- O
OH OH H2N
N
N N N
HO
O P O P
O O
O-
O O
OH OH H2N
N
N N N
O P O O
O- -
-
ATP
ADP
アデニン
リン酸 リボース
細胞活動や化学合成に 利用できるエネルギー 太陽光や食物からの
エネルギー
図 24: ATPとADPの化学構造
食物 糖類
タンパク質 脂質
アミノ酸
グルコース
グリセリン 脂肪酸
ピルビン酸
アセチルCoA
TCA回路 CO2
NADHの形での
還元力 電子伝達系
O2
ATP
H2O 解糖系
細胞質
ミトコンドリア
図25: 食物からエネルギーを得る仕組み
ATP ATP
グルコース
フルクトース
1,6-ビスリン酸
ジヒドロキシ アセトンリン酸 ジヒドロキシ アセトンリン酸 ピルビン酸
ピルビン酸
ATP ATP
ATP ATP
NADH NADH
HO HO
OH OH OH
O
図 26: 解糖系
9
糖と脂肪の分解我々が生きてゆくためにはもちろん食物を食べる必要があります。その最も大きな理由は、食物 から生きてゆくために必要なエネルギーを得るためです。化学的にはこのエネルギーはATP(アデ ノシン三リン酸)と呼ばれる分子に蓄えられています(図24)。ATPがADP(アデノシン二リン酸) という分子に変わるとき蓄えられたエネルギーが放出され、様々な活動に使われます。食物からエ ネルギーを得る過程は、食物からATPを合成する過程であると考えていいでしょう。ではどのよう な過程で食物からATPが合成されるのでしょうか。その全体像を図25に示しました。
食物は消化されたあと、糖類は解糖系と呼ばれる一連の化学反応の経路へ進みます(図26)。解糖 系ではATP生成の中心的経路であり、そこでは我々にとって最も重要な糖であるグルコース1分子 がピルビン酸2分子になり、その過程で2分子のATPとNADH(ニコチンアミドアデニンジヌクレ オチド)ができます。これをもう少し詳しく説明すると図26に示すように、グルコース1分子につ きATP2分子が消費されますが、その後の反応で4分子のATPが得られるため、差し引き2分子 のATPが得られるのです。このときのATPはADPから生成されます。ADPはATPからエネル ギーが放出されたときにできる低エネルギーの分子なので、そこからまたATPができるのは不思 議に思えるかも知れませんが、図27に示すように、この反応ではエネルギー的に起こりにくい反応 をエネルギー的に起こりやすい反応と共役させる酵素が用いらているのです。このとき、グリセル アルデヒド3-リン酸がグリセルアルデヒド3-リン酸脱水酵素によって3-ホスホグリセリン酸に変換 され、これによって生じた高エネルギーのリン酸基がADPに転移し、ATPができます。
嫌気状態ではこれ以上酸化が進まず、解糖系でATPを作るために必要なNAD+ を再生する発 酵を行います(図28)。好気状態の場合、ピルビン酸・脂肪酸はミトコンドリアでアセチルCoAに 変換されてクエン酸回路に入っていき、さらにその先の電子伝達系で多量のATPが生産されます。
4ミトコンドリアと葉緑体は独自のゲノムを持っており、そこで合成されるタンパク質もある。
グリセルアルデ ヒド3-リン酸のC- H結合の酸化エ ネルギー
自由エネルギー NADH
NAD
高エネルギーの 結合を持つ1,3ビ スホスホグリセリ ン酸の合成
1,3ビスホスホグ リセリン酸の高 エネルギー結合 の切断
ADP ATP
図 27: 共役反応によりNADHとATPが生成される仕組み
グルコース
ピルビン酸2分子 ATP
ADP
乳酸2分子 NADH
2 2
解糖 2
NAD+ 2
2 NAD+
NAD+の再生
+2H+
グルコース
ピルビン酸 2分子 ATP
ADP
エタノール 2分子 NADH
2 2
解糖
2 NAD+ 2
2 NAD+
NAD+の再生
+2H+ アセトアルデヒド
2分子
H+ CO2 2分子 (a)
(b)
図 28: 発酵
(a)乳酸が作られる発酵(b)アルコールとCO2が作られる発酵
ࡑ࠻࠶ࠢࠬ
ౝ⤑
ᄖ⤑
⤑㑆ㇱಽ
図29: ミトコンドリアの構造
10
ミトコンドリアの4つの区画とその役割ミトコンドリアはATPを生産する細胞小器官で、真核生物の細胞に広く存在します。ミトコン ドリアには、外膜、内膜、マトリックス、外膜と内膜の間の部分である膜間部分の4つの区画が存 在し、エネルギーの合成においてそれぞれが重要な役割を担っています(図29)。
外膜 ポリンと呼ばれる輸送タンパクをもち、それが大きな親水性チャネルを構成しています。その ため膜間部分の分子構成は、5000ドルトン以下の分子については細胞質とほぼ同じになって います。
内膜 膜輸送タンパクによってマトリックスに入る分子の種類の選択をするという機能と、マトリッ クスから送られてきた高エネルギー電子を使って電子伝達系でプロトンを膜間部分に汲み出 すという機能を持っています。
マトリックス 膜を通ってきたピルビン酸と脂肪酸を酸化してアセチルCoAを作り、クエン酸回路 を回します。そしてできた高エネルギー電子を内膜の電子伝達系に送ります。また、電子伝達 系で形成されたプロトン勾配を使ってマトリックス内部にあるADPからATPを合成します
(これについては11節で詳しく説明します)。
膜間部分 マトリックスから送られてきたATPを用いて、ほかのヌクレオチドをリン酸化します。
ミトコンドリアは独自のゲノムおよび転写・翻訳系を持っており、ミトコンドリア内に存在する タンパク質の一部を合成しています。ミトコンドリアは真核細胞の中に存在する微生物のようにも 見えますが、実際にミトコンドリアの祖先は他の微生物で、それが原始的な真核細胞の中に取り込 まれたと考えられています(図30)。
原始的な真核生物
細菌(ミトコンドリアの祖先)
図30: ミトコンドリアの進化
11
クエン酸回路と電子伝達系クエン酸回路5と電子伝達系は、共に酸素呼吸が可能な細胞すなわち、ミトコンドリアを有する細 胞のみに存在するエネルギー代謝経路です。クエン酸回路はミトコンドリア内膜内(マトリックス) に存在し、電子伝達系はミトコンドリア内膜上に存在しています。
ミトコンドリアを有する細胞では、解糖系によって生じたピルビン酸を二酸化炭素とアセチル基 に分解し、さらにこのアセチル基をCoA(補酵素A)に結合させ、アセチルCoA6 を生産します。
アセチルCoAはオキサロ酢酸と結合し、クエン酸となってクエン酸回路というループ状の代謝経 路に入ります(図31)。アセチルCoAは2つの炭素を有し、オキサロ酢酸は4つの炭素を有する分 子です。クエン酸はその合計、6つの炭素を有しています。クエン酸回路は回転しながら徐々に炭素 を減らしつつ電子をNAD+に渡してゆきます。この「電子を渡す」ことこそがクエン酸回路の大き な意義なのです。NAD+は電子を受け取って大きな還元力を持つNADHになります。
これに対して、電子伝達系の意義は「ATPを生産する」ことにあると言えるでしょう。クエン酸 回路で合成されたNADHから得た電子が電子伝達系を流れるときのエネルギーを用いてH+がマ トリックスから膜間部分に汲み上げられます。結果的にミトコンドリア内膜を境に、膜間部分では H+が増え、マトリックス側ではe−が増えます(図32)。なおこのような電子の動きを予測するに は酸化還元電位の計算が有効です。
このようにして生じたプロトン勾配を利用してATP合成酵素がリン酸を使ってADPをリン酸化 し、ATPを合成します。NADHが酸化されて放出された電子は最終的に酸素に移って水分子が出 来上がるため、ここで酸素が消費されます。このようにATPが合成される工程は酸化的リン酸化と 呼ばれます。
5他にTCA回路、TCAサイクル(tricarboxylic acid cycle)、トリカルボン酸回路、クレブス回路(Krebs cycle)など の呼び方がある
6「アセチルコーエー」と読む。
アセチルCoA 2C
6C
6C クエン酸
NADH + H+ CO2 5C
NADH + H+
CO2 4C
GTP 4C
FADH2 4C 4C
4C オキサロ酢酸
クエン酸回路
H3C C S CoA O
NADH + H+
図31: クエン酸回路
クエン酸回路 電子伝達系
電子伝達系 電子伝達系 電子伝達系
H+ H+ H+
アセチルCoA
ピルビン酸 脂肪酸
ADP + Pi ADP + Pi
H+
H+ ATP
ピルビン酸 脂肪酸
O2 O2
2H2O
CO2 CO2
ATP
内膜
ATP合成酵素 外膜 膜間部分
マトリックス
NAD+ NADH
e-
NADH脱水素酵素複合体 シトクロムb-c1複合体
シトクロム酸化酵素複合体
NADH
図32: 電子伝達系
酸素のない発酵で1分子のグルコースから獲得できるATPの数は2個です。これに対し、酸素 を使う場合は解糖系から2個、TCA回路から2個、電子伝達系から約30個のATPを獲得できる のです。酸素を用いる呼吸代謝がATPを獲得する上でいかに効率が良いか、よく分かります。
12
食物の備蓄と利用これまで述べてきたように、生物が活動するためには常に細胞内でATPが必要となります。こ れは生体内で起こる多くの化学反応のエネルギー源となるからです。しかし現実には生物は常に食 物にありつけるわけではありません。そこで生物には食物分子を備蓄する機構が備えられています。
動物の場合、肝臓や筋肉など多くの細胞の細胞質にグリコゲンという多糖が小さな顆粒として存 在し、急な必要性に素早く対応できるようになっています。一方、脂肪は脂肪組織に蓄えられ、必 要なときは血中に放出されます。これはグリコゲンに比べ備蓄性に優れるという長所があり、また 1g当たりのエネルギーもグリコゲンの約2倍です。
植物の場合、食物をデンプンと脂肪という形で蓄えています。どちらも葉緑体に蓄えられ、日光 からATPを生産できない時に使われます。種子には多量のデンプンや脂肪が蓄えられており、成長 するためのエネルギーや生体分子として使われています。発芽中の種子は、必要に応じて、脂肪を グルコースに変えられます7。
7動物細胞では糖は脂肪に変えられるが、脂肪酸は糖に変えられない。