アミノ酸の疲労軽減作用に関する研究
日本大学大学院生物資源科学研究科
津田 悠一
2021
目次
緒論 1
第1章 分岐鎖アミノ酸が運動時の生体応答に与える影響 5
第1節 緒言 5
第2節 材料および方法 6
第3節 結果 8
第4節 考察 10
図表 13
第2章 アミノ酸混合物が運動時のコルチゾール応答に与える影響 21
第1節 緒言 21
第2節 材料および方法 23
第3節 結果 26
第4節 考察 28
図表 32
第3章 アミノ酸混合物が運動時の疲労感に与える影響 39
第1節 緒言 39
第2節 材料および方法 41
第3節 結果 45
第4節 考察 47
図表 51
総括 62
謝辞 64
引用文献 65
原著論文リスト 79
1 緒論
現在、肥満・糖尿病・高血圧・脂質異常症などの生活習慣病が世界的に蔓延し、社会問 題となっている。世界の肥満者数は、1980年に8億8,500万人だったのが、2013年には2.5 倍の21億人にまで増加したと報告されている[1]。世界保健機関(WHO)やNCD Risk Factor
Collaborationなどの報告では、世界の成人の糖尿病有病者数は 4 億2,200 万人、高血圧有
病者数は11億3,000万人にのぼっており、今後さらに増加すると推測されている[2,3]。厚
生労働省の調査によると、わが国においても高血圧患者数は993万人、糖尿病患者数は329 万人、脂質異常症患者数は220万人にのぼっており[4]、今後の患者数の増加が懸念されて いる。これらの疾患は、日本人の三大死因である、がん、心疾患、脳血管疾患をはじめと したあらゆる疾患の発症リスクを高める[5-8]。
わが国が抱えるもう一つの問題が高齢化である。内閣府の報告では、わが国の総人口に 占める65歳以上人口の割合(高齢化率)は28.4%となっており、2065年には約2.6人に1人 が65歳以上、約3.9人に1人が75歳以上となると推測されている[9]。加齢は骨格筋量を 低下させることが知られており、それに伴って筋力や身体機能が大幅に低下する[10]。こ の症状はサルコペニアと呼ばれ、要介護リスクや転倒などによる骨折のリスクが高まるこ とが明らかとなっている[11,12]。また、加齢によって認知障害を引き起こすリスクが顕著 に増大することも数多く報告されている[13,14]。これらの予防・改善は、高齢化社会にお ける健康寿命延伸を目指すにあたっての重大な課題と言える。
生活習慣病の予防・改善には、運動が有効であることが示されている[15-17]。複数のシ ステマティックレビューの結果から、運動介入により体重、血圧、中性脂肪、空腹時血糖 などが有意に改善することが明らかとなっている[15,16]。また、運動は筋肉量および筋力 を維持・向上させ、サルコペニアの予防に大きく寄与することが報告されている[18,19]。 さらに、運動が認知障害の予防・改善に大きな効果を示すことも確認されている[20,21]。 こうしたエビデンスの蓄積に加え、近年では、健康維持・増進に対する国民の意識の向上 や、東京オリンピック・パラリンピックの開催(予定)などを背景に、アスリートのみならず
2
一般層においても運動・スポーツに対する関心が高まっている。スポーツ庁が実施した調 査によると、2019年における成人の週1日以上のスポーツ実施率は53.6%であり、1991年
の27.8%から大幅に増加している[22]。また、スポーツサプリメントの市場規模に目を向け
ると、2014年には295億円であったのが2018年には64%増の485億円にまで拡大してお り、今後も伸長していくことが予想されている[23]。
一方、運動を実施する際に克服しなくてはならない問題として疲労がある。日本疲労学 会では、疲労とは、過度の肉体的および精神的活動、または疾病によって生じた独特の不 快感と休養の願望を伴う身体の活動能力の減退状態と定義している。運動をすると身体が 疲労し、その疲労感がパフォーマンスの低下や運動に対するモチベーションの低下につな がることは、多くの人が経験している現象である。疲労感が発生する機序には、様々な因 子の関与が報告されている。例えば、長時間運動時では、グリコーゲンの枯渇により血糖 が低下し、疲労感が増加することが報告されている[24]。アンモニアは、長時間運動時のエ ネルギー枯渇により筋肉タンパク質の分解が促進することで産生され、神経毒性を介して 疲労感を誘発する[25,26]。Fernstromらは、運動により脳内においてセロトニン産生が促進 し、それが中枢性疲労の発生に寄与していると報告している[27]。疲労感の発生に関する 明確な機構については未だ不明な部分も多いが、このように多くの因子が疲労感の発生に 関与していることは明らかにされつつある。
パフォーマンスの向上を目指すアスリートや、趣味や健康のために運動を実施したい 人々にとって、運動による疲労感を軽減する機能性食品に対するニーズは高い。しかしな がら、運動・スポーツにおける疲労感を軽減する科学的エビデンスに基づいた機能性食品 はほとんど開発されていない。今般のスポーツサプリメント市場は、プロテイン商品群と アミノ酸商品群が双璧をなしている。プロテイン商品群は、ホエイやソイ由来のタンパク 質を用いた粉末食品や飲料が主要な商品であり、機能性としては、運動後の摂取による筋 合成促進作用や翌日以降の筋損傷回復促進作用が数多く報告されている[28-31]。しかしな がら、運動時の疲労感に対する作用に関するエビデンスはほとんどない。アミノ酸商品群 は、タンパク質原料から微生物発酵、抽出法、酵素法などを用いて製造したアミノ酸を活
3
用し、顆粒や飲料の形態で商品化されている。アミノ酸の運動に関連する生理作用につい ての科学的エビデンスとしては、ロイシンあるいはロイシン高含有アミノ酸混合物が、運 動後の筋合成促進作用および筋肉痛軽減作用を有することが報告されている[32-34]。運動 時の疲労感に関する報告としては、分岐鎖アミノ酸(BCAA)混合物の効果を検証した研究が いくつかなされている。例えば、運動後のBCAA混合物の摂取により回復期の疲労感が改 善したとの報告がある[35]。しかしながらこの報告では、運動時の疲労感に対する作用に ついては検証していない。テコンドー選手を対象に行った臨床試験では、BCAA・アルギ ニン・シトルリン混合物の摂取により、試合後に実施した反応テストのパフォーマンスが 改善したとの結果が得られているが、試合直後の自覚運動強度(RPE)には影響を及ぼして いなかった[36]。これらのBCAAに関する研究では、そのほとんどがBCAA混合物の効果 を検証している。すなわち、バリン、ロイシン、およびイソロイシンなど、各BCAA単独 の効果についてはほとんど知見がない。また、BCAA以外のアミノ酸では、オルニチンの 摂取が運動時の疲労感に及ぼす影響をビジュアルアナログスケール(VAS)により評価した 報告があるが、プラセボ群との有意差は認められていない[37]。上述のように、運動時にお けるアミノ酸の疲労感軽減作用について、ヒトを用いて明確に示した研究は少ない。一方、
疲労感を直接評価はしていないが、運動による疲労感に影響を与える可能性を示唆する報 告もある。例えば、BCAA混合物の摂取は、血液脳関門でのトリプトファン輸送の阻害を 介して運動時の脳内セロトニン産生を抑制することが動物実験により明らかにされており [38,39]、運動時の疲労感軽減につながる可能性がある。アルギニンは運動時の脂質代謝を 促進することが報告されており[40-42]、脂質代謝亢進による運動時の血糖あるいはグリコ ーゲンの節約により、疲労感を軽減する可能性がある。これらの背景から、運動時の疲労 軽減作用に着目してアミノ酸の生理機能を研究することは、新規性が高く有用な知見が得 られると考えられる。
本研究では、運動時に摂取することで疲労感を軽減することができる新規アミノ酸混合 物を提案することを目的とし、アミノ酸およびアミノ酸混合物の効果について検証を行っ た。まず、運動時における様々な生理作用が報告されているBCAAに着目し、各BCAA、
4
すなわち、バリン、ロイシン、およびイソロイシンが運動時の生体応答に与える影響につ いてラット水泳モデルを用いた試験により比較検討した。さらに、新規な比率で混合した アミノ酸混合物を作製し、そのアミノ酸混合物が運動時のホルモン応答や疲労感に与える 影響を、臨床試験を実施して評価した。
5
第1章 分岐鎖アミノ酸が運動時の生体応答に与える影響
第1節 緒言
いくつかのアミノ酸は運動時の生理機能に影響を与えることが知られている。例えば、
アルギニンは運動時の血中アンモニアおよび乳酸の上昇を抑制する[43]。β-アラニンの摂 取は筋中カルノシン合成を促進させ、運動パフォーマンスを向上させることが複数の臨床 研究により示されている[44,45]。BCAA は、運動時の生体応答に多様な影響を及ぼすアミ ノ酸として知られ、多数の研究が実施されている。例えば、BCAA混合物の摂取により運 動時の筋損傷の指標である血清クレアチンキナーゼ(CPK)活性の上昇が抑制されることが 報告されており、BCAA混合物が運動に起因する筋損傷を軽減することが示唆されている
[46]。De Paloらは、継続的なBCAA混合物摂取が運動後の血中乳酸濃度を低減することを
見出している[47]。また、BCAA混合物は運動時の乳酸閾値を上昇させるとの報告もあり、
運動能力向上への寄与も示唆されている[48]。しかしながら、各 BCAA、すなわち、バリ ン、ロイシン、およびイソロイシンが運動時の生体応答に与える影響についてはほとんど 知見がない。
そこで本章では、各BCAAが運動による疲労に及ぼす影響を明らかにすることを目的と し、バリン、ロイシン、およびイソロイシンの単回投与が運動時の各種代謝関連マーカー に与える影響について、ラット水泳モデルを用いた試験により比較検討した。さらに、バ リン、ロイシン、およびイソロイシンのうち、代謝関連マーカーに影響を及ぼしたアミノ 酸について、運動後のラットの自発行動量に与える影響を検討することで、疲労に対する 作用を行動学的に評価した。
6 第2節 材料および方法
実験動物
6 週齢の雄性 Sprague-Dawley ラットを日本クレア(東京)より購入した。ラットは、温度
22 ± 1℃、湿度55 ± 15%、照明1日12 時間に環境調節を施した飼育室において飼育し、購
入後1週間の馴化期間を設けた。全ての動物実験は、株式会社明治動物実験倫理委員会の 承認を受け(Approval No.2017_3871_0017, 2017_3871_0087)、株式会社明治動物実験倫理委 員会の定める動物実験における倫理規定に則って実施した。
水泳運動試験
ラットを約 18 時間絶食させ、バリン(n=10)、ロイシン(n=10)、イソロイシン(n=10)、ま たは対照として蒸留水(n=9)を経口投与した。バリン、ロイシン、およびイソロイシンは和 光純薬工業(大阪)より購入した。投与用量は1 g/kg体重、投与容量は10 mL/kg体重とした。
投与30分後から、ラットに水泳運動を60分間実施させた。水泳運動は、バケツに45 cm
の深さで35 ± 1℃の温水を張り、常に同時に6匹のラットが水泳運動を実施しているよう
に調節し、1匹あたりの遊泳可能面積が250 cm2となるようにした。水泳運動の条件は神田 らの方法を参考に設定した[49]。投与前および運動直後に尾静脈より採血を行い、血糖自 己測定器ブリーズ2(バイエル薬品、大阪)を用いて血糖値を測定した。また、ヘパリン入り ヘマトクリット管で血液を採取し、12,000 rpm、4℃で15 分間遠心分離して尾静脈血漿を 得た。その後、イソフルラン麻酔下にて腹部大静脈より採血を行い、肝臓を摘出した。得 られた血液を3,000 rpm、4℃で15分間遠心分離し、大静脈血漿を得た。
血液検査
尾静脈血漿乳酸濃度はLactate Colorimetric Assay Kit II K627-100 (BioVision, CA)により測 定した。大静脈血漿遊離脂肪酸濃度および総ケトン体濃度はNEFA C-test Code 279-75401お よびAutokit Total Ketone Bodies Code 415-73301, 411-73401 (和光純薬工業)を用いて測定し
7
た。大静脈血漿グリセロール濃度は Glycerol Colorimetric Assay Kit No.10010755 (Cayman
Chemical, MI)により測定した。大静脈血漿中性脂肪濃度および CPK活性はFuji Dri-Chem
Slide TG-PIIIおよびCPK-PIII (富士フイルム、東京)を用いてDri-Chem 7000 chemistry analyzer により測定した。大静脈血漿コルチコステロン濃度は Corticosterone Enzyme Immunoassay Kit K014-H5 (Arbor Assays, MI)を用いて測定した。
肝グリコーゲン量測定
採取した肝臓を均質化し、95℃で10分間加熱した。その後、18, 000 g、4℃で10分間遠 心分離し、得られた上清を肝臓抽出液とした。肝臓抽出液中のグリコーゲン量をGlycogen Colorimetric Assay Kit K646-100 (BioVision)により測定した。
自発行動量測定
ラットを約18時間絶食させ、バリン(n=17)、ロイシン(n=16)、または対照として蒸留水
(n=17)を経口投与した。投与用量は1 g/kg体重、投与容量は10 mL/kg体重とした。投与30
分後から、ラットに水泳運動を60分間実施させた。水泳運動の条件は前述と同様とした。
運動後から10分間、ラットの自発行動量をactivity monitoring system, ACTIMO-10 (シンフ ァクトリー、福岡)により測定した。
統計解析
全てのデータは、平均値±標準誤差で表した。One-way analysis of variance (ANOVA)を用 いて検定を行い、有意差が認められた場合にDunnett's testを行った。いずれの検定も有意 水準は両側5%とした。これらの検定には、エクセル統計(社会情報サービス、東京)を使用 した。
8 第3節 結果
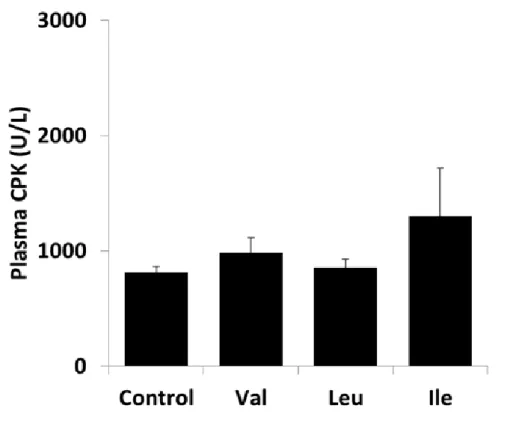
運動時の肝グリコーゲン、血糖、血漿中乳酸、CPK、コルチコステロンに与える影響 運動後の肝グリコーゲン量の結果をFig. 1-1 に示す。バリン投与により、対照群と比較 して運動後の肝グリコーゲン量が高値を示した(p<0.05)。一方、ロイシンまたはイソロイシ ン投与では対照群と差はなかった。運動前後の血糖値の結果をFig. 1-2に示す。60分間の 水泳運動により、血糖値は顕著に低下した。また、バリン投与により、対照群と比較して 運動後の血糖値が有意に高値を示した(p<0.05)。一方、ロイシン投与では、対照群と比較し て運動後の血糖値は有意に低値であり(p<0.01)、イソロイシン投与では、対照群と比較して 運動後の血糖値に有意な変化は見られなかった。運動前後の血漿乳酸濃度の結果をFig. 1- 3に示す。血漿乳酸濃度は60分間の水泳運動により顕著に上昇したが、群間で有意な差は なかった。運動後の血漿CPK活性の結果をFig. 1-4に示す。血漿CPK活性は群間で有意 な差はなかった。運動後の血漿コルチコステロン濃度の結果を Fig. 1-5 に示す。バリンま たはロイシン投与により、対照群と比較して運動後の血漿コルチコステロン濃度が有意に 低値を示した(p<0.05)。一方、イソロイシン投与では、対照群と比較して差はなかった。
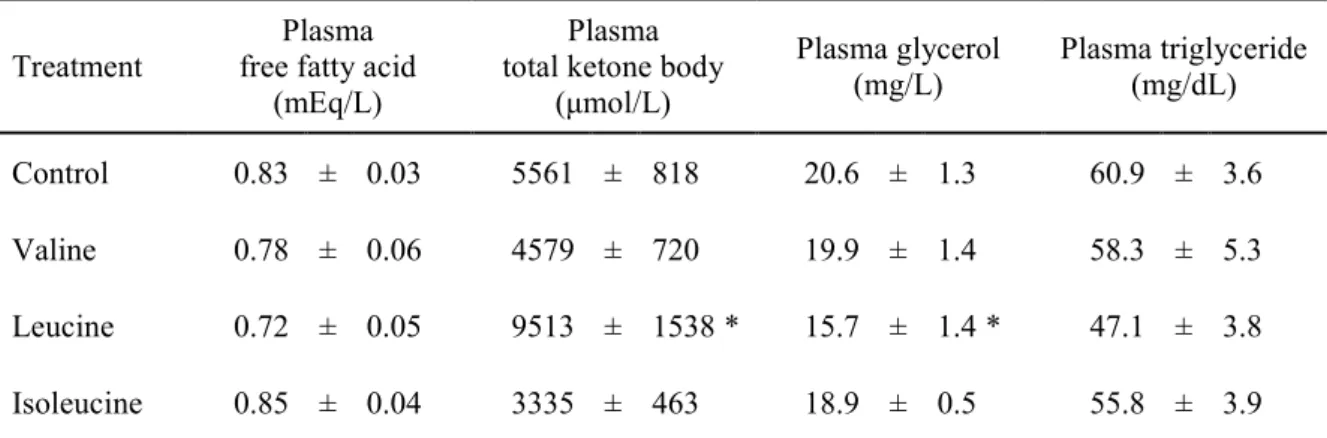
運動時の血漿中遊離脂肪酸、総ケトン体、グリセロール、および中性脂肪に与える影響 運動後の血漿中遊離脂肪酸、総ケトン体、グリセロール、および中性脂肪の結果をTable 1-1 に示す。ロイシン投与により、対照群と比較して血漿総ケトン体濃度が有意に高値を 示し(p<0.05)、血漿グリセロール濃度は有意に低値を示した(p<0.05)。バリンまたはイソロ イシン投与では、対照群と比較してこれらの血中指標に有意差はなかった。
運動後の自発行動量に与える影響
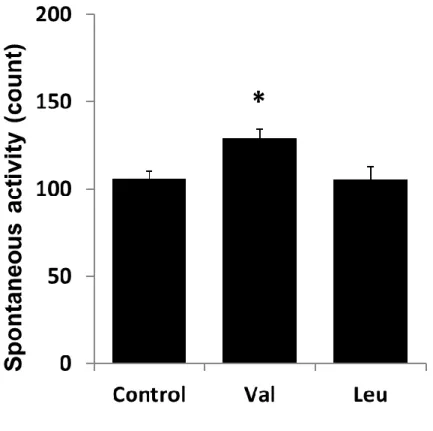
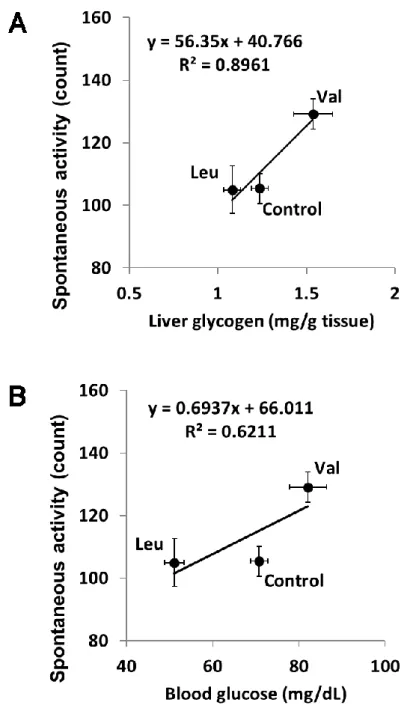
運動後の自発行動量の結果をFig. 1-6 に示す。バリン投与により、対照群と比較して運 動後の自発行動量が有意に高値を示した(p<0.05)。一方、ロイシン投与では対照群と有意な 差はなかった。また、運動後の自発行動量と運動後の肝グリコーゲン量または血糖値の間
9 には強い正の相関が見られた(Fig. 1-7)。
10 第4節 考察
運動開始時にはまず、エネルギー源として主に糖質が利用される。すなわち、筋肉や肝 臓に貯蔵されたグリコーゲンが分解されてエネルギー源として利用されるとともに、血糖 レベルの維持にも働く。長時間運動時にはグリコーゲンの枯渇により血糖が低下し、疲労 感が増加することが報告されている[24]。ヒトにおいては、筋グリコーゲンは肝グリコー ゲンよりも多く存在し、エネルギーへの寄与が大きいと考えられるが、ラットにおいては、
筋グリコーゲンよりも肝グリコーゲンが運動時のエネルギー源として重要であることが報 告されている[50]。これらのことから、ラットにおいて、運動時に血糖値あるいは肝グリコ ーゲン量を維持することは疲労軽減やパフォーマンス維持に重要であると考えられる。本 研究において、バリン投与によって運動後の血糖値および肝グリコーゲン量が対照群と比 較して有意に高く維持された。これらの結果から、運動前のバリン摂取は肝グリコーゲン および血糖の低下を抑制し、運動時の身体コンディションの維持あるいは疲労軽減に寄与 することが明らかとなった。げっ歯類における運動時のBCAA代謝は、ヒトとは対照的に 筋肉よりも肝臓において高い活性を有することが報告されている[51,52]。糖原性アミノ酸 の一種であるバリンの摂取により、運動時にラット肝臓において糖質源の代替としてバリ ンが代謝され、血糖値および肝グリコーゲンの維持に貢献した可能性がある。肝臓の代謝 関連酵素の活性などを評価することで、作用機序に関するより明確な知見が得られると考 えられる。一方、本研究において、同じく糖原性アミノ酸であるイソロイシンの投与では 血糖および肝グリコーゲン維持作用は認められなかった。イソロイシンはインスリン非依 存的に糖取り込みを促進する作用を有することが報告されていることから[53]、その作用 によって糖利用が亢進したことにより、バリンのような血糖および肝グリコーゲン維持作 用が見られなかったと考えられる。また、ロイシン投与では、運動後の血糖値は対照群と 比較して有意に低値となった。ロイシンはインスリン分泌を介して糖取り込みを顕著に促 進することが報告されていることから[54]、本結果は糖利用が亢進したことによるもので あることが示唆された。この結果から、運動前のロイシン摂取は、運動時の身体コンディ
11
ションや疲労感を悪化させる可能性があると考えられた。
低強度あるいは中強度の長時間運動時には、エネルギー源として主に脂質が利用される
[55]。本研究において実施した水泳運動は、60 分間に及ぶ長時間運動であるため、脂質代
謝が亢進する条件であると考えられる。本研究では、ロイシン投与により、対照群と比較 して運動後の血漿中総ケトン体量が有意に上昇し、血漿グリセロール濃度が有意に低下し た。ロイシン投与により運動後の血糖値が顕著に低下していたことから、ロイシン投与群 では対照群よりも運動時の血糖が速やかに利用されることが推察された。また、脂質代謝 は体内の糖質源が枯渇した際に特に亢進する。したがって、ロイシン投与群では、運動時 の速やかな血糖利用により糖質源が枯渇し、脂質代謝が亢進した結果、ケトン体レベルが 上昇したと考えられる。一方、グリセロールは、エネルギー必要時にはグリセロールキナ ーゼおよびグリセロール-3-リン酸デヒドロゲナーゼにより異化され、解糖系にて代謝され る。したがって、ロイシン投与による糖質源の枯渇により、グリセロール異化反応が亢進 したと考えられる。
げっ歯類におけるコルチコステロン(ヒトにおけるコルチゾール)は副腎皮質より分泌さ れる糖質コルチコイドである。コルチコステロンあるいはコルチゾールは、血圧上昇作用、
血糖上昇作用、抗炎症作用などを示す。臨床研究においては、運動時の血糖低下に応じて 血中コルチゾールが顕著に分泌されることが報告されている[56,57]。また、血中コルチゾ ールは運動後の疲労感と強い相関を有することが確認されている[58,59]。本研究において、
バリン投与により、血漿コルチコステロン濃度が有意に低下した。バリンは血糖および肝 グリコーゲンを維持することが示されているため、それらを介してコルチコステロンの上 昇抑制に寄与していると考えられる。これらのことから、運動前のバリン摂取は、血糖お よび肝グリコーゲンの維持を介して、運動時の疲労軽減あるいは身体コンディションの向 上に寄与することが示唆された。また、本研究では、ロイシン投与においても血漿コルチ コステロン濃度の有意な低下が認められたが、ロイシン投与では運動後の血糖値はむしろ 低下していたため、ロイシン投与によるコルチコステロン低減作用には、糖代謝以外の因 子が関与していると考えられる。具体的には、血糖低下によりコルチコステロンが一時的
12
に急激な上昇を示し、フィードバック制御によって上流の因子の分泌を抑制した結果、本 研究における測定時点でのコルチコステロンが低値を示した可能性が考えられる。
自発行動量は、動物実験における行動学的評価として一般的に用いられる[60]。自発行 動量の低下は疲労感の上昇による行動のモチベーションの低下に起因することが報告され ているように[61]、自発行動量は疲労感の指標として用いることができる。本研究では、代 謝関連マーカーに影響を及ぼしたバリンおよびロイシンの投与が運動後の自発行動量に与 える影響を評価した。その結果、バリン投与により、対照群と比較して運動後の自発行動 量が有意に高値を示した。一方、ロイシン投与では対照群と比較して顕著な変化は見られ なかった。これらの結果から、運動前のバリン摂取は運動による疲労感を軽減する作用を 有することが示された。また、運動後の自発行動量と肝グリコーゲン量または血糖値には、
強い正の相関が確認された。したがって、バリン投与による自発行動量の増加は血糖およ び肝グリコーゲン維持作用が寄与していることが示唆された。
BCAA 混合物の継続摂取により、運動後の乳酸および CPK の上昇が抑制されることが 報告されているが[46,47]、本研究においては、バリン、ロイシン、またはイソロイシン投 与は、運動後の血中乳酸および CPK には影響を及ぼさなかった。これらの結果について は、本研究が単回投与実験であったことが考えられる。今後、バリン、ロイシン、または イソロイシンの継続投与による影響を評価することで、血中乳酸および CPK に及ぼす影 響を明確にすることが可能である。また、血中乳酸およびCPKは運動強度や測定タイミン グが結果に大きく影響する指標であるため、より高強度の運動条件や異なるタイミングで の測定を実施することで、本研究とは異なる結果が得られる可能性がある。
本研究の結果から、BCAAのうち特にバリンが運動時の血糖および肝グリコーゲンの低 下を抑制すること、コルチコステロン分泌を抑制すること、運動後の自発行動量を増加さ せることを見出し、バリンが運動時の疲労軽減作用を有する可能性を明らかにした。一方、
ロイシン投与は運動時の血糖を低下させ、身体コンディションを悪化させる可能性が示唆 された。また、イソロイシン投与では、運動に関連する代謝マーカーに顕著な変化は見ら れないことを確認した。
13
Fig. 1-1. Effects of valine, leucine, and isoleucine on liver glycogen content during exercise.
Rats were fasted for 18 h, and then valine, leucine, or isoleucine (1 g/kg body weight) was orally administered. At 30 min after the administration, the animals swam for 60 min. Liver samples were collected after the exercise, and then liver glycogen contents were measured. Values are mean ± SEM (n=9–10). * p<0.05 compared to the control group.
14
Fig. 1-2. Effects of valine, leucine, and isoleucine on blood glucose level during exercise.
Rats were fasted for 18 h, and then valine, leucine, or isoleucine (1 g/kg body weight) was orally administered. At 30 min after the administration, the animals swam for 60 min. Blood glucose concentrations were measured using whole blood from a tail vein before the administration and immediately after the exercise. Values are mean ± SEM (n=9–10). * p<0.05, ** p<0.01 compared to the control group.
15
Fig. 1-3. Effects of valine, leucine, and isoleucine on plasma lactate level during exercise.
Rats were fasted for 18 h, and then valine, leucine, or isoleucine (1 g/kg body weight) was orally administered. At 30 min after the administration, the animals swam for 60 min. Blood samples were collected from a tail vein before the administration and immediately after the exercise, and then plasma lactate concentrations were measured. Values are mean ± SEM (n=9–10).
16
Fig. 1-4. Effects of valine, leucine, and isoleucine on plasma CPK level during exercise.
Rats were fasted for 18 h, and then valine, leucine, or isoleucine (1 g/kg body weight) was orally administered. At 30 min after the administration, the animals swam for 60 min. Blood samples were collected from inferior vena cava after the exercise, and then plasma CPK concentrations were measured. Values are mean ± SEM (n=9–10). CPK: creatine phosphokinase.
17
Fig. 1-5. Effects of valine, leucine, and isoleucine on plasma corticosterone level during exercise.
Rats were fasted for 18 h, and then valine, leucine, or isoleucine (1 g/kg body weight) was orally administered. At 30 min after the administration, the animals swam for 60 min. Blood samples were collected from inferior vena cava after the exercise, and then plasma corticosterone concentrations were measured. Values are mean ± SEM (n=9–10). * p<0.05 compared to the control group.
18
Table 1-1. Effects of valine, leucine or isoleucine on plasma free fatty acid, total ketone body, glycerol and triglyceride concentrations during exercise.
Treatment Plasma
free fatty acid (mEq/L)
Plasma total ketone body
(μmol/L)
Plasma glycerol
(mg/L) Plasma triglyceride (mg/dL) Control 0.83 ± 0.03 5561 ± 818 20.6 ± 1.3 60.9 ± 3.6
Valine 0.78 ± 0.06 4579 ± 720 19.9 ± 1.4 58.3 ± 5.3
Leucine 0.72 ± 0.05 9513 ± 1538 * 15.7 ± 1.4 * 47.1 ± 3.8 Isoleucine 0.85 ± 0.04 3335 ± 463 18.9 ± 0.5 55.8 ± 3.9 Rats were fasted for 18 h, and then valine, leucine or isoleucine (1 g/kg body weight) was orally administered. At 30 min after the administration, the animals swam for 60 min. Blood samples were collected from inferior vena cava after the exercise and then plasma free fatty acid, total ketone body, glycerol and triglyceride concentrations were measured. Values are mean ± SEM (n=9–10). * p<0.05 compared to the control group.
19
Fig. 1-6. Effects of valine or leucine on spontaneous activity after exercise.
Rats were fasted for 18 h, and then valine or leucine (1 g/kg body weight) was orally administered.
At 30 min after the administration, the animals swam for 60 min. After the exercise, spontaneous activities were estimated for 10 min. Values of the spontaneous activity are mean ± SEM (n=16–17).
* p<0.05 compared to the control group.
20
Fig. 1-7. Correlations between the spontaneous activity after exercise and the liver glycogen level (A) or blood glucose level (B) after exercise.
Values of the spontaneous activity are mean ± SEM (n=16–17). Liver glycogen and blood glucose values are mean ± SEM (n=9–10).
21
第2章 アミノ酸混合物が運動時のコルチゾール応答に与える影響
第1節 緒言
ストレスに対して生体は hypothalamic-pituitary-adrenal (HPA)軸を活性化することにより
対抗する[62,63]。この反応は、副腎皮質刺激ホルモン放出ホルモン(CRH)が視床下部から分
泌されることから開始される。続いて下垂体から副腎皮質刺激ホルモン(ACTH)分泌が刺激 され、副腎皮質からの糖質コルチコイド(ヒトにおけるコルチゾール)の分泌を刺激するこ とによりストレスに対抗する。運動によるストレスはHPA軸を刺激し、コルチゾール分泌 を亢進することがこれまでに数多く報告されている[64-67]。また、長時間の運動における 血糖値の低下が下垂体、副腎皮質からのACTH、コルチゾール分泌亢進を引き起こすこと も明らかとなっている[56,57]。さらに、ストレスフルな運動時におけるコルチゾール応答 と疲労感には顕著な相関が認められる[58,59]。これらの報告から、コルチゾールはグリコ ーゲン枯渇や血糖低下に応答し、血糖を維持して疲労を遅らせるために分泌されると考え
られる。Lugerらは、身体コンディションは運動によるHPA軸亢進の抑制と関与している
こと、実際、運動鍛錬者はACTH、コルチゾール応答が低減されていることを報告してい る[68]。これは、トレーニングによりミトコンドリア機能が向上し、脂質代謝が亢進したこ とで、グリコーゲンおよび血糖の低下が抑えられ、結果としてコルチゾール応答の抑制に 結びつくものと考えられる。また、コルチゾール分泌上昇は筋分解を誘発し[69-72]、糖質 コルチコイドに起因する筋委縮を引き起こす可能性を示した報告も存在する[73,74]。した がって、運動時のコルチゾールレベルを抑制することは、運動時に誘発されるストレス・
疲労の軽減に加え、グリコーゲンや血糖維持といったより良い身体コンディションを維持 し、また、運動後の筋分解を軽減することにもつながると考えられる。
運動時のコルチゾール分泌を抑制するための栄養学的戦略について、いくつかの研究が 報告されている。例えば、運動時の糖質摂取は運動後の血中コルチゾール濃度を抑制する ことが報告されている[75-77]。アミノ酸摂取が運動時のコルチゾール応答に与える影響を
22
評価した報告もあるが、アミノ酸摂取が運動時のコルチゾール応答を抑制したというヒト での報告はほとんどない。ある報告では、運動前のBCAA摂取は、運動時の成長ホルモン 分泌を抑制し、運動後のテストステロンレベルを上昇させたが、運動時のコルチゾール応 答には影響を与えなかった[78]。また、BCAA 5.2 g、 必須アミノ酸4.3 g、タウリン1.5 g を含むサプリメントを摂取しても、運動時のコルチゾール応答には影響を及ぼさなかった [79]。しかしながら、アミノ酸の中には運動によるコルチゾール応答に影響を与える可能 性を有するものが存在する。例えば、本学位論文の第1章において、BCAAの中でもバリ ンが運動時の肝グリコーゲンおよび血糖維持を介して顕著に糖質コルチコイド分泌を抑制 することを動物実験により明らかにしている。また、アルギニンは脂質代謝を亢進するこ とが動物およびヒトにおいて示されており[40-42]、脂質代謝亢進による運動時の血糖ある いはグリコーゲンの節約により、コルチゾール応答が抑制される可能性がある。セリンを 基質として合成されるホスファチジルセリンは、運動時のコルチゾール分泌を抑制したと 報告されている[80]。そこで本章では、これら 3 種のアミノ酸の効果が相乗的に働くこと によって、運動時のコルチゾール応答を抑制することができるのではないかと考え、これ らを利用した新規アミノ酸混合物を作製し、このアミノ酸混合物の摂取が運動時のコルチ ゾール応答に与える影響についてランダム化二重盲検プラセボ対照クロスオーバー比較試 験を行った。
23 第2節 材料および方法
被験者
習慣的に運動を実施している 20~39 歳の健常男性48 名が試験に参加した。全ての参加 者は、本試験の内容を理解し、文書同意が得られた者である。食物アレルギーを有する者、
喫煙習慣のある者、試験開始前1ヶ月以内に200 ml、または3か月以内に400 ml以上の採 血を実施した者、1 ヶ月以内に他の臨床試験に参加していた者、その他の理由で試験責任 医師により不適と判断された者は、試験から除外した。また、参加者には本試験で実施す る運動と同様の運動を事前に実施させ、その運動を完遂できない者、またはその運動によ り血漿コルチゾール濃度が全く上昇しない者は、試験から除外した。被験者募集および選 抜は、2016年8月から9月に実施した。合計20名の被験者を選抜し、乱数を用いてラン ダムに2群に群分けした。割付情報は試験終了まで被験者および研究実施者には明かさず、
盲検性を維持した。被験者のサンプルサイズは、5%水準で80%の検出力が達成されるよう に設定した。また、サンプルサイズの設定には、栄養介入が運動時の血中パラメーターに 与える影響を評価した過去の文献も参考にした[78,79]。本研究では、全ての被験者が全プ ロトコルを完遂した。統計解析実施前に、介入とは関係しない血液検査の異常値により 3 名、プロトコルの逸脱により 2 名が解析対象から除外されたため、最終的に 15 名の被験 者を解析対象とした(Fig. 2-1)。被験者情報をTable 2-1に示した。試験中、被験者は普段の 運動量および食事量を変えないよう指導され、運動試験実施当日にはサプリメントを摂取 しないよう指導された。また、運動試験実施前日および当日には、アルコールの摂取を禁 じ、カフェイン摂取量を変えないよう指示した。
研究デザイン
ランダム化二重盲検プラセボ対照クロスオーバー比較試験を CPCC 株式会社(東京)で実 施した。まず、各被験者の最大酸素摂取量(VO2max)を自転車エルゴメーター(Aerobike
75XLIII、コナミスポーツ、東京)を用いた漸増負荷試験により測定した。1週間以上経過後、
24
被験者は本試験と同様の運動試験を実施し、本試験の運動を完遂できるか、また、この運 動により血漿コルチゾール濃度が上昇するかを確認した。被験者選抜および割付後、被験 者は本試験を実施した。本試験実施当日は、被験者には朝食として規定食を摂取させた。
来院後、被験者はアルギニン1.8 g、バリン1.1 g、セリン0.1 gを含むセルロースカプセル
(松谷化学、兵庫)、または空カプセルを運動開始30分前に摂取した。摂取する順番はラン
ダム化され、試験期間中の二重盲検性は維持された。被験者にはカップに入ったカプセル を渡し、それを直接掴むことなく素早く摂取するよう指示することで、被験者がどちらの 試験食品を摂取しているのか分からないようにした。アミノ酸の配合比および用量は、各 アミノ酸が期待した効果を発揮するための必要量を勘案するとともに、物性などの面にお いて実用化に適した配合となるように設定した。具体的には、アルギニンの脂質代謝亢進 作用やバリンの血糖維持・コルチゾール低減作用には一定の用量が必要であること、ホス ファチジルセリンは低用量で効果が認められているため、セリンはアルギニンおよびバリ ンよりも低用量で効果が期待できること、アルギニンは吸湿性が高いため顆粒状の食品を 製造する際には配合量に限界があること、バリン摂取によって疲労感を誘発するアンモニ アを過剰に産生させないようにすること、などの条件をもとに配合比および用量を設定し た。その後、被験者は自転車エルゴメーター(Aerobike 75XLIII)を用いて50%VO2maxで80 分間の運動を実施した。中間には3分間の休憩を挟み、被験者はその休憩中に十分な水分 補給を行った。運動条件は、非鍛錬者の血中コルチゾール濃度が上昇していた過去文献を 参考にして設定した[81]。運動試験は、恒温恒湿室にて実施した。1週間以上のウォッシュ アウト期間を挟み、被験者はもう一方の試験食品で同様の運動試験を実施した。これらの 運動試験は、2016年9月から10月に実施した。本研究は、チヨダパラメディカルケアク リニック臨床試験審査委員会(Approval No. 20160720)および明治臨床試験審査委員会
(Approval No. 91)の承認を受け、ヘルシンキ宣言に基づく倫理原則、人を対象とする医学系
研究に関する倫理指針(文部科学省、厚生労働省告示)を遵守し、実施した。また、本試験は UMIN Clinical Trials Registryに2016年8月19日に登録した(UMIN000023587)。
25 血液検査
運動開始前および運動終了直後に上腕静脈より採血した。EDTA-2Na 入り採血管で採取 した血液を1,700 g、4℃で10分間遠心分離し、血漿を得た。得られた血漿は、血漿コルチ ゾール濃度およびACTH濃度の測定に用いた。プレーン採血管で採取した血液を1,700 g、
4℃で10分間遠心分離し、血清を得た。得られた血清は、血清総ケトン体濃度、遊離脂肪 酸濃度、およびCPK活性の測定に用いた。1N過塩素酸入り採血管で採取した血液を15~60 分間氷冷してタンパク除去した後、1,700 g、4℃で10分間遠心分離し、除タンパク血漿を 得た。得られたサンプルは、血漿乳酸濃度およびアンモニア濃度の測定に用いた。NaF・
EDTA-2Na 入り採血管で採取した血液は、血糖の測定に用いた。全ての血中パラメーター
の測定はLSIメディエンス(東京)で実施した。
統計解析
本研究では、血漿コルチゾール濃度を主要評価項目、血漿ACTH濃度、血糖、血漿乳酸 濃度、血漿アンモニア濃度、血清中CPK活性、血清総ケトン体濃度、および血清遊離脂肪 酸濃度を副次評価項目とした。全てのデータは、平均値±標準誤差または変化量±95%信 頼区間で表した。Two-way repeated measures ANOVAを用いて検定を行い、主効果または交 互作用がみられた場合にBonferroni-corrected post hoc t-testを行った。運動前後の変化量に ついては、対応のあるt-testで検定を行った。コルチゾール変化量とACTH 変化量の相関 については、Spearman’s rank correlation analysisで検定を行った。いずれの検定も有意水準 は両側 5%とした。これらの検定には、SPSS v. 22 (IBM Japan, Ltd.、東京)を使用した。
26 第3節 結果
本研究で用いた試験食品の摂取に関連した副作用の報告はなかった。また、1 回目にア ミノ酸混合物を摂取し、2 回目にプラセボを摂取する場合、あるいはその逆順で摂取する 場合の試験の順番による影響の有無を確認するため、血漿コルチゾール濃度について順序 効果を評価した結果、本研究において順序効果は認められなかった。
コルチゾールおよびACTH
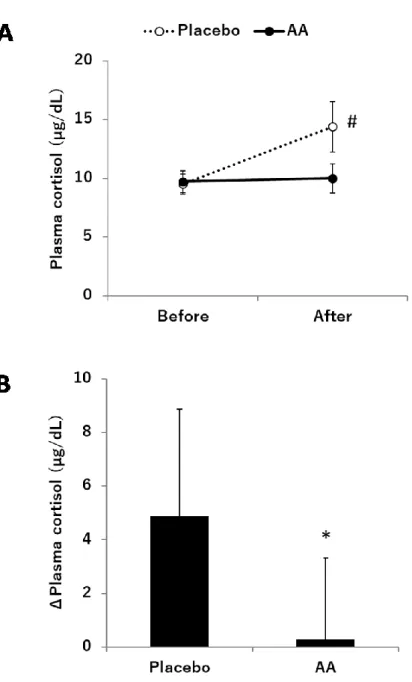
運動前後における血漿コルチゾール濃度を Fig. 2-2に示す。血漿コルチゾール濃度にお いて、食品と時間の交互作用に有意差が認められた(p<0.05)。Post hoc t-testの結果、プラセ ボ摂取条件において、運動後の血漿コルチゾール濃度は運動前と比較して有意に高値を示 した(p<0.05)。一方、アミノ酸混合物摂取条件においては、血漿コルチゾール濃度は運動前 後で有意差は認められなかった(p=0.846)。さらに、アミノ酸混合物摂取条件では、プラセ ボ摂取条件と比較して運動前後の血漿コルチゾール濃度の上昇が有意に低値であった
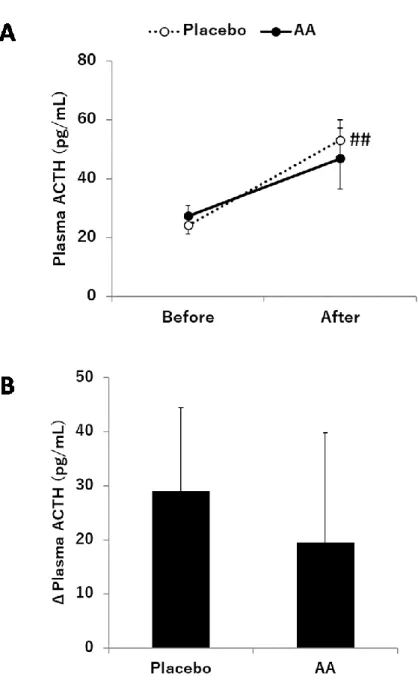
(p<0.05)。運動前後における血漿ACTH濃度をFig. 2-3に示す。血漿ACTH濃度において
は、時間の主効果に有意差が認められた(p<0.01)。Post hoc t-testの結果、プラセボ摂取条件 においてのみ運動前後の血漿 ACTH 濃度で有意な上昇が認められ(p<0.01)、アミノ酸混合 物摂取条件では運動前後での有意な上昇は見られなかった(p=0.057)。運動前後の血漿 ACTH濃度の変化量は、条件間で有意差は認められなかった(p=0.454)。運動前後における コルチゾール/ACTH比をFig. 2-4に示す。コルチゾール/ACTH比においては、時間の主効 果に有意差が認められ(p<0.01)、post hoc t-testの結果、両条件ともに運動前後で有意差が認 められた(p<0.01)。
運動前後の血漿コルチゾール変化量と血漿ACTH変化量の相関をFig. 2-5に示す。両条 件ともに血漿コルチゾール変化量と血漿 ACTH 変化量の間に強い相関が認められ(プラセ ボ摂取条件; r=0.58, p<0.01、アミノ酸混合物摂取条件; r=0.95, p<0.01)、両条件における近似 曲線はほぼ同等であった。
27 その他の血中パラメーター
運動前後におけるその他の血中パラメーターをTable 2-2に示す。血糖、血漿乳酸濃度、
血漿アンモニア濃度、血清CPK活性、血清総ケトン体濃度、および血清遊離脂肪酸濃度に おいて、時間の主効果が認められた(p<0.01)。Post hoc t-testの結果、両条件ともに、運動前 後で血糖が有意に低下し(p<0.01)、血漿乳酸濃度、血漿アンモニア濃度、血清CPK活性、
血清総ケトン体濃度、および血清遊離脂肪酸濃度が有意に上昇した(p<0.01)。
28 第4節 考察
運動によりグリコーゲンの枯渇や血糖値の低下などが生じると、血糖を維持して疲労を 遅らせるためにコルチゾールが分泌される[64-67]。また、その過剰分泌は筋分解や筋委縮 を引き起こす[69-72]。本研究では、プラセボ摂取条件において、運動後の血漿コルチゾー ル濃度が顕著に上昇していた。したがって、本研究で実施した運動条件は、本研究に参加 した被験者にとってコルチゾール分泌を誘導するのに十分な条件であったと考えられる。
一方、アミノ酸混合物摂取条件においては、運動前後で血漿コルチゾール濃度は変動せず、
運動前後の血漿コルチゾール濃度の上昇はプラセボ摂取条件と比べて有意に低値であった。
これらの結果は、アミノ酸混合物の単回摂取が長時間運動時のコルチゾール上昇を有意に 抑制することを示しており、本アミノ酸混合物が運動時のストレス軽減あるいは身体コン ディションの向上、さらには運動後の筋分解抑制に寄与することを示唆している。
ACTH は HPA 軸のストレス応答に関与する因子の一つであり、下垂体から分泌された ACTH は副腎皮質からのコルチゾール分泌を促進する[62,63]。本研究では、プラセボ摂取 条件において、血漿ACTH濃度が運動によって顕著に上昇していた。この結果から、プラ セボ摂取条件におけるコルチゾールの上昇は、主としてACTHの分泌亢進が寄与している と考えられた。アミノ酸混合物摂取条件においては、運動前後の血漿ACTH濃度に有意な 差はなかったものの、上昇傾向が認められた。さらに、条件間においてもACTHレベルに 差は認められなかった。これらの結果から、本研究において、アミノ酸混合物摂取は直接 的にACTH分泌に影響を及ぼしてはいないと考えられた。
コルチゾール/ACTH比は副腎応答の評価に用いられる指標である[82-84]。本研究におい て、条件間でコルチゾール/ACTH比に有意差は認められなかったことから、アミノ酸混合 物摂取は副腎応答には影響を及ぼしていないことが示唆された。また本研究では、運動前 後の血漿コルチゾール変化量と血漿ACTH変化量の相関を評価した。アミノ酸混合物摂取 によるコルチゾール分泌あるいはACTH分泌の抑制の程度が異なっていた場合、コルチゾ ールと ACTH の相関における近似曲線の傾きはプラセボ摂取条件とは異なることが予想
29
される。しかしながら、本研究では両条件の近似曲線の傾きはほぼ同等であった。この結 果から、下垂体からのACTH分泌と副腎皮質からのコルチゾール分泌はアミノ酸混合物摂 取によって同程度抑制されることが示唆された。ただし、被験者1名分のデータが他デー タと大きく異なっており、近似曲線の算出に大きな影響を及ぼしていた。より正確な知見 を得るためには、さらにサンプルサイズを増やした検討が必要である。また、アミノ酸混 合物摂取がHPA軸の最上流に位置する視床下部からのCRH分泌に与える影響についても 検討することで、アミノ酸混合物の作用機序の解明につながることが期待される。
血糖レベルはコルチゾール分泌に強く関係している。Tabata らは、長時間運動時の血糖 低下がACTHおよびコルチゾールの分泌を促進することをヒトで明らかにしている[56,57]。
本研究では、両条件ともに運動によって血糖が低下したが、条件間での差は認められなか った。このことから、アミノ酸混合物摂取によるコルチゾール応答抑制には、血糖は関与 していないことが示唆された。血糖が低下すると、コルチゾールは筋または肝グリコーゲ ンを分解して血糖レベルを維持する[85,86]。著者らは、ラットを用いた検討でバリンの経 口摂取が肝グリコーゲンを維持し、コルチコステロン分泌を抑制することを明らかにして いる。これらの知見から、本研究においてもアミノ酸混合物摂取によって筋または肝グリ コーゲンレベルが維持された可能性が考えられた。
アルギニンは脂質代謝を亢進することが報告されている[40-42]。Lucoti らは、アルギニ ン(8.3 g/day)の長期摂取が体脂肪量および腹囲を減少させ、さらには糖代謝およびインスリ ン応答を改善することを見出している[87]。Hurtらは、3 gのアルギニンを1日3回12週 摂取することで、肥満者の内臓脂肪が低減することを報告している[88]。本研究では、条件 間で血清総ケトン体濃度および血清遊離脂肪酸濃度に差は認められなかった。これらの結 果から、アルギニン1.8 gを含むアミノ酸混合物の単回摂取は、脂質代謝に対して顕著な影 響を及ぼさないこと、アミノ酸混合物のコルチゾール上昇抑制効果に脂質代謝を変動させ るメカニズムは関与していないことが示唆された。
CPKは筋肉に傷害が生じた際に血中に逸脱する酵素で、細胞傷害のマーカーとして利用
される[89,90]。BCAAやホスファチジルセリンの摂取により、運動後のCPKレベルの上昇
30
が抑制されることが報告されている[91,92]。アンモニアは、運動時にエネルギーが枯渇し た際の筋タンパク分解によって生じ、神経毒性を有することが知られている[25,26]。また、
アルギニンにより運動時のアンモニアレベルの上昇が抑制されることが報告されている
[43]。本研究では、条件間で血清CPK活性および血漿アンモニア濃度に差は認められなか
った。これらの結果から、アミノ酸混合物摂取による運動時のコルチゾール応答抑制にこ れらの因子は関与していないことが示唆された。
本研究では、アミノ酸混合物摂取によるコルチゾール応答抑制作用のメカニズムを明確 にするには至らなかったが、メカニズムに関していくつかの仮説を述べる。まず、アルギ ニンは一酸化窒素(NO)産生の基質として知られ[93]、NOはHPA軸の応答を制御すること が報告されている[94,95]。これらのことから、アミノ酸混合物摂取がNO産生を介してコ ルチゾール分泌を抑制した可能性が考えられる。次に、本研究で用いたアミノ酸混合物の 摂取は運動時の血中トリプトファン/BCAA比を低下させ、脳内セロトニン産生を抑制する ことが示唆されており(第3章参照)、セロトニンはセロトニン受容体刺激を介してHPA軸 を活性化させることが報告されている[96]。これらのことから、アミノ酸混合物摂取がセ ロトニン産生抑制を介してコルチゾール応答を抑制した可能性が考えられる。さらに、ホ スファチジルセリンは運動時の ACTH およびコルチゾールの分泌を抑制することが報告 されており[80]、セリンの経口摂取で脳内のセリン濃度が上昇し、ホスファチジルセリン が合成されることも報告されている[97,98]。これらのことから、アミノ酸混合物摂取がホ スファチジルセリン産生を介してコルチゾール分泌を抑制した可能性が考えられる。
本研究の結果から、アルギニン、バリン、セリンを含むアミノ酸混合物の単回摂取は、
運動時のコルチゾール応答を顕著に抑制することが明らかとなり、運動時におけるストレ ス・疲労の軽減、あるいは身体コンディションの改善に貢献する可能性が示された。また、
その作用機序には、少なくとも糖代謝や脂質代謝に関連する血中マーカーは関与していな いことが示唆された。本研究で示されたアミノ酸混合物の効果は、NO やホスファチジル セリン産生、あるいは脳内セロトニン産生の抑制を介したものである可能性が考えられ、
アミノ酸混合物摂取がこれらの因子の産生に及ぼす影響を評価することで、本研究結果の
31 作用機序解明につながると期待される。
32 Fig. 2-1. Flowchart of the study participants.
AA: amino acid.
33 Table 2-1. Basic characteristics of the study subjects.
n=15
Age (years) 32.3 ± 1.2
Height (cm) 169.9 ± 1.5
Body weight (kg) 64.4 ± 1.5
BMI (kg/m2) 22.3 ± 0.4
VO2max (mL/min/kg) 39.8 ± 1.7
Values are the mean ± SEM. BMI: body mass index.