総 説
月
)
免 疫(
1
)
自然免疫と獲得免疫
東京女子医科大学医学部微生物学免疫学教室 ヤ ギ ジ‘ュンジ 八 木 淳 二 ( 受 理 平 成27年 1月19日) Immunity(1) Innate and Acquired Immunity
Junji YAGI
Department of Microbiology and Immunology, Tokyo Women's Medical University School of Medicine
The immune system consists of innate and acquired immunity. The molecular basis of pathogen recognition by pattern recognition receptors (PRRs) of innate immunity has been extensively investigated. The subsequent immunity, known as acquired immunity, is characterized by specific antigen recognition by antigen receptors on T and B cell clones. However, the discrimination between self and non-self is not strict in acquired immunity, thus relying on regulatory T (Treg) cells to prevent autoimmune responses. The immune system apparently develops while interacting with microbes, enabling a co-existence. Treg cells are also key regulatory cells that enable our co-existence with microbes. Here, innate and acquired immunity will be described as concisely as possible to pro -vide the readers with an understanding of how our immune system allows or inhibits co-existence with“non -self."
Key W ords: innate immunity, acquired immunity, pattern recognition receptors, regulatory T cells
はじめに 人類の直接的な祖先である原人は,約
3
0
0
万年前 に誕生した.地球の生物進化のカレンダーでは,大 みそかの午後7時にあたる. もちろんそれよりず、っ と以前から微生物は存在していたのであって,人類 誕生からただちに微生物とのせめぎ合いが始まっ た.われわれの免疫システムは,われわれの生体内 に侵入した自己 selfではない非自己 non-selfである 微生物や物質(抗原)を排除して恒常性を維持する システムである.せめぎ合いの過程で,微生物は, 免疫システムをすり抜ける機序を獲得してきたし, 免疫システムは,それに対抗する機序を獲得してき たというように,共に相互作用を繰り返しつつ共進 化してきたと考えられる.その過程で免疫システム は異物である微生物と共存する機構も得てきた.免 疫システムは,互いに連動しあう 2つのシステム, 自然免疫と獲得免疫からなる.本編では,それらに ついて免疫システムの特性を考えながら簡潔に記し たいと思う. 免疫システムの特性 免疫システムは,多くの機能を異にする免疫担当 細胞がお互いに相互作用をしながら,平衡系を形 イ乍っている.そのなかには,免疫反応を正に制御(異 物を排除)する細胞と負に制御(反応を抑制)する 細胞が含まれ,巧妙な制御下で反応が起こり,収束 する (Fig.1).異物が侵入すると,可塑性という言葉 が適切ではないかもしれないが,平衡系は,いった ん偏向するが異物の排除に伴って元の平衡状態に戻 図:八木淳二 干162-8666東京都新宿区河田町8-1 東京女子医科大学医学部微生物学免疫学教室 E-mail: [email protected]=〈川州
APC MHC
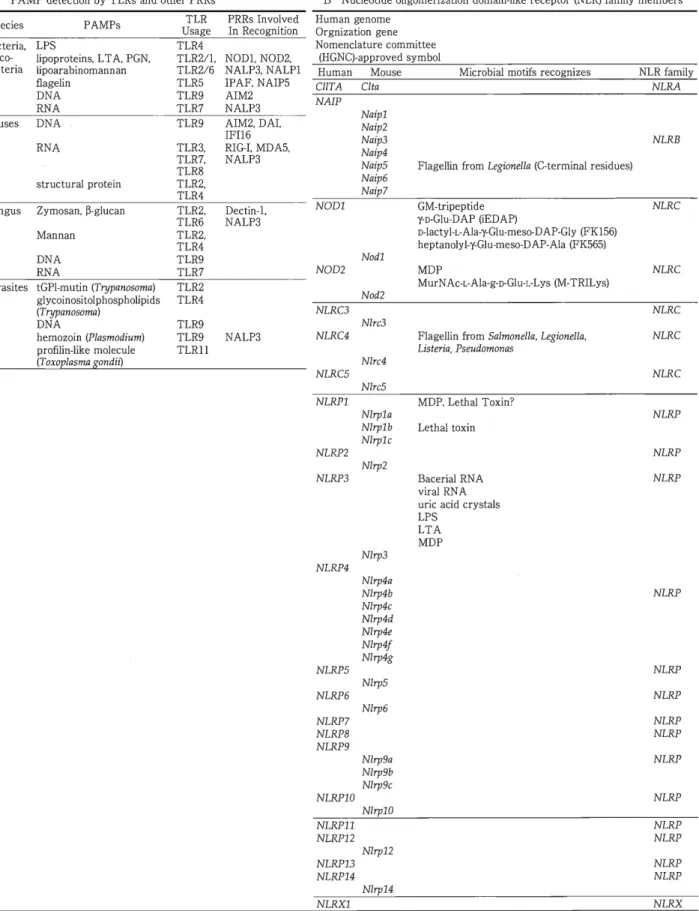
Activation' proliferation CytotoxicT cell Removal ofvirusand tumor cells KillerT cell Fig.1 Interactionamong immunocompetent cellsandT cell subsets Naive CD4+ and CD8+ T cellsrecognize complexesof antigenpeptides presented by MHC class IIand MHC class1 molecules, respectively, on APCs andareactivated. Under the in -fluenceof cytokines present in themicroenvironment, T cellsubsetsarematured and dif -ferentiated into effectorcells and exhibitimmune responses. Note thatthedifferentiation ofone T cell subsetregulatesthat of the other subset. る.ただし免疫記憶が残る.免疫担当細胞は,胸 腺や骨髄という一次リンパ組織のなかで生成され, まだ抗原とは出会っていない非感作の細胞が,末梢 の組織において抗原と出会い,分化・成熟してはじ めて機能を発揮しえる細胞(エフェクター細胞)と なる(Fig.l).一次リンパ組織で,無数の抗原と反応 しえるレパートリーが獲得されるとともに,一次リ ンパ組織と末梢の組織において 自己とは反応しな いレパートリーの形成,すなわち自己寛容と一部の 微生物と共存しうる制御機構が獲得される.免疫シ ステムの抗原に対するレパートリー形成は,以下で 簡単に触れるように,巧妙ではあるものの,無駄な かっ危険性を字む機構によっている.免疫異常に基 づく疾患(自己免疫疾患,アレルギーなど)を発症 する危険性は常に存在している. 自然免疫と獲得免疫 自 然 免 疫 は , 細 胞 因 子 と し て 好 中 球 や マ ク ロ ファージなどの貧食細胞や N K(naturalkiller)細胞 が,液性因子としては,補体などが働き異物排除の 最前線に位置している.貧食細胞がどのように異物 をnon-selfとして認識し,貧食するかについては,長 く全く不明であった.ところが,周知のように, 1997 年のJanewayのグループによる晴乳類のToll様受 容体(Toll-likereceptor : TLR) 4の発見)1を端緒とし -2-て,爆発的に解明が進んで、いる.貧食細胞は,微生 物 が 有 す る 病 原 体 関 連 分 子 パ タ ー ン (pathogen-associated molecular patterns : P AMPs) と呼ばれ る微生物独自の分子パターンを認識するパターン認 識受容体 (patternrecognition receptors : PRRs)を 介して活性化し免疫応答が誘導される.反応は,単 に異物を貧食し消滅させるだけではなく,獲得免疫 系の引き金を引くとともに,その方向性にも多大な 影響を与える.PRRsは多種類存在するが,膜貫通 型と細胞質型に分けることができる.Table 12 )3)に PRRsを網羅的に示す.解明が急速に進み,多くの PRRsやそれらに関連した複雑な反応が明らかにさ れつつあるが,一方で総合的に理解することが難し い現状を感じる.いずれにしても,病原微生物が侵 入すると,シグナル伝達型のPRRsがそれを察知 (sensing) し,獲得免疫系からのインターフエロン-y Gnterferon-y: IFN-y)などの刺激とあいまって, 貧食した微生物の殺滅や炎症性サイトカイン[イン ターロイキン・1sGnterleukin-1s: IL-1s)や腫蕩壊死 因子 (tumornecrosisfactor: TNF) ]の産生による 炎症の増強により,異物の排除にあたる.同時に, マクロファージや樹状細胞は, 自身の MHC分子や 補助刺激因子 [CD80(B7・1),CD86 (B7-2)]の発現 を増強し取り込んだ、抗原の獲得免疫系への提示を
Table 1 PRRs A PAMP detection by TLRs and other PRRs Species Bacteria, myco-bacteria vlruses Fungus PAMPs LPS lipoproteins, L T A.PGN, lipoarabinomannan flagelin DNA RNA DNA RNA structural protein Zymosan, s-glucan Mannan DNA RNA Parasites tGPl-mutinσrypanωoma) glycoinositolphospholipids (Trypanosoma) DNA hemozoin(Plasmodium) profilin-like molecule (Toxoplasma~ondii) TLR Usage TLR4 PRRs Involved In Recognition TLR2/1.NODl.NOD2, TLR2/6 NALP3, NALPl TLR5 IPAF, NAIP5 TLR9 AIM2 TLR7 NALP3 TLR9 AIM2, DA,I TLR3, TLR7, TLR8 TLR2, TLR4 TLR2, TLR6 TLR2, TLR4 TLR9 TLR7 TLR2 TLR4 TLR9 TLR9 TLRll IFIl6 RIG-,IMDA5, NALP3 Dectin-,l NALP3 NALP3 B Nucleotide oligomerization domain-like receptor(NLR) family members Human genome Orgnizationgene Nomenclature committee (HGNC)-approved symbol Human Mousε CllTA CIta NAIP NOD1 NOD2 NLRC3 NLRC4 NLRC5 NLRPl NLRP2 NLRP3 NLRP4 NLRP5 NLRP6 NLRP7 NLRP8 NLRP9 NLRP10 Na伊1 Na伊2 Na伊3 Na伊4 Na伊5 Na伊6 Naip7 Nodl Nod2 Nlrc3 Nlrc4 Nlrc5 Nlrpla Nl中lb Nlrplc Nlrp2 Nlrp3 Nlrp4a Nlrp4b Nlrp4c Nlrp4d Nlrp4e Nlrp4J Nlrp4g Nlrp5 Nl巾6 Nlrp9a Nlrp9b Nlrp9c Nlrpl0 NLRPll NLRP12 Nl中12 NLRP13 NLRP14 Nlrp14 Microbial motifs recognizes Flagellin from Legionella(C-terminal residues) GM・tripeptide チD-Glu-DAP(iEDAP) D-lactyl・レAla~千Glu-meso-DAP-Gly(FK156) heptanolyl-'子Glu-meso-DAP-Ala(FK565) MDP MurNAc-L-Ala-g-D-Glu-L-Lys (M-TRILys) Flagellin fromSalmonella, Legionella, Listeria, Pseudomonas MDP, Lethal Toxin? Lethal toxin Bacerial RN A viral RNA uric acid crystals LPS LTA MDP NLR family NLRA NLRB トlLRC NLRC トlLRC NLRC NLRC NLRP NLRP NLRP ふlLRP NLRP NLRP NLRP NLRP NLRP NLRP かlLRP NLRP NLRP NLRP NLRXl NLRX
PRRs recognize pathogen molecular patterns. Two representative PRRs, Toll-like receptors (TLRs) (A) and Nucleotide oligomeriza -tion domain (NOD)-like receptors (NLRs) (B) are shown.
PRRs recognize a wide range of pathogens, triggering the innate immunity response. Other PRRs include C-type lectin receptors and RIG-1-1ike receptors. Complicated responses elicited by innate immunity have been elucidated. (Adapted from ref. 2 Kawai T et al: Immunity 34: 637-650, 2011; ref. 3 Franchi L et al: Immunol Rev 227: 106-128,2009)
増強するとともに,
I
L
-
1
2
の産生によるl
型ヘルパーT
細胞の誘導など獲得免疫系の方向性にも関与す る.最近, PRRs は,微生物など外来異物の侵入のみ ならず,生体内部環境の危険な状態,すなわち,が んやストレスなどによる組織の障害において主に死 細胞から放出される成分 (dangersignal)をsensing し,生体反応を誘導することで,生体の恒常性の維 持にも寄与していることが明らかになってきてい る.この反応は,自己に対する反応であることから, 過剰や異常な反応が自己免疫疾患や炎症性疾患に関 連することも明らかにされつつある.外因性の病原 体物質に特異的と考えられてきた自然免疫センサー が病原体成分に類似する自己にも応答するために自 己への寛容の破綻をきたし自己免疫疾患をうむと考 えられる.移植免疫は多種多様なT
細胞抗原受容体 およびB細胞抗原受容体を介した獲得免疫が中心 と考えられてきた一方で、 移植後の臓器再潅流傷害 で生じる臓器障害とそれに対する初期免疫応答との 関連性が報告されている.ちなみに,筆者は, 1986 年から 1990年まで, Janewayの研究室に留学する 機会を得たが,彼がそのころ,動物に抗体を産生さ せるために,病原体由来の免疫賦活剤であるアジ、ユ バントを抗原と同時投与しなければならないことに ついて, 免疫研究者の"dirtysecret"と言いながら, アジュバントの必要性の解明に熱く取り組んでいた ことを思い出す. 獲得免疫は,B細胞,T細胞によるリンパ球の免疫 応答である.自然免疫系の細胞と決定的に異なる点 は,それらが,クローン性を有することである.抗 原を認識するそれぞれの受容体である細胞表面の免 疫グロプリンとT
細胞抗原受容体は,抗原認識にか かわる可変部のアミノ酸がクローンごとに異なり, 異なる抗原を認識する.一つのクローンは,多くと も数十個の細胞からなり,末梢リンパ組織には,無 数ともいえる抗原と反応しえるおびただしい種類の クローンが抗原と出会う以前からあらかじめ用意さ れている.ひとたび生体にある抗原が侵入すると, 対応するクローンが活性化し“クローン拡大"する とともに,非感作の細胞は,成熟,分化してエフェ クター細胞となる.ついで,エフェクター細胞は, 全身を巡り対応する異物の排除にあたる.かつて Barnettが提唱しノーベル賞に輝いた“クローン選 択説"は,現在も色あせることなく受け入れられて いる.獲得免疫系の細胞と自然免疫系の細胞の特性 について, Table 24 )に簡単に比較した. -4-抗原認識の多様性獲得の機構と自己寛容 リンパ球のクローン性を生み出す一次リンパ組織 における抗原レセプター遺伝子再構成の分子機序に ついては,一般の教科書に譲る.ここでは,可変部 遺伝子断片の結合部の自由度が抗原特異性の多様性 に大きな貢献をする際に,ランダムなヌクレオシド の欠失ないし挿入に伴ってoutofframeとなって, 蛋白をコードできなくなる危険性が非常に高い点を 強調したい.多様性を生むために多大の犠牲を伴っ ている無駄の多い機構と言える.実際,胸腺におい て分化する未熟T
細胞の大部分は,分化過程で死ん でしまう.また, 自己反応性のリンパ球,たとえばT
細胞を例にあげるなら,自己反応性T
細胞の抗原 受容体は,未熟な段階で胸腺上皮細胞や胸腺中のマ クロファージや樹状細胞の提示する特定の自己抗原 ペプチドとMHC
の複合体と強く結合する(高い結 合親和性で結合する)ことにより,アポトーシスの シグナルが誘導され死滅する.胸腺が提示する自己 抗原のすべてに対する自己反応性T細胞が末梢リ ンパ組織へ漏出することをあらかじめ阻止すること で, 自己寛容を成立させる機構である(中枢性自己 寛容).ただしこれは,不完全であることがわかっ ている.胸腺の自己抗原ペプチドのレパートリーは, 末梢組織のそれと完全には一致せず,一部末梢組織 に特異的な抗原ペプチドが存在するからである. し たがって中枢性自己寛容の機構では末梢にのみ存在 するペプチドに対して反応するT
細胞は,胸腺で異 物反応性T細胞として成熟し,末梢リンパ組織へ定 着してしまう.事実,末梢リンパ組織中の自己反応 性T
細胞クローンの存在をi
nv
i
t
r
o
で容易に確認す ることができる. 免疫システムは,このような不都合に対して,必 然的に末梢に存在する自己反応性T
細胞を不応答 性(anergy)に陥らせる機構によって対処している. この機構はいわゆるtwosignal theoryに基づいて いる.すなわち, T細胞が十分に活性化・増殖する ためには, T細胞受容体が抗原を提示する細胞(抗 原提示細胞=antigen presenting cell : APC)が提示 する抗原ペプチドとMHC
の複合体を認識して誘導 されるシグナル(第1のシグナル,自rstsignal)だけ では不十分で、あり,第2のシグナル(secondsignal, 補助(共)刺激シグナル=co-stimulatory signal)が 供給される必要がある.もし 第2のシグナルが供 給されず第1
のシグナルのみが供給された場合は, T細胞は反応しないだ、けではなく,冬眠状態に陥っTable 2 Differences in antigen recognition by antigen receptors of innate and acquired immunity and their clonality Innate i mmunity Acquired immunity Specificity Speci行cityagainst structure (molecular pattern) I Recognition offine structure (antigen) inmicrobial shared by microbial species I molecule: recognize non-microbial antigen
写
n
w
a叡
e
n
J
設
e
m
w
e2
5
叶 式 決 バ
[
血血血
Receptor Coded by germinal cells; poorindiversity Coded by a gene generated byrecombination of genetic segments; high in diversity LPS scavenger receptor Receptor distribution
I
No c1onality: receptors on all cells belonging to the same cell lineage are identical. c1onality: c10nes of Iymphocytes with different specificities express different receptors. yes; based on the selection of self-reactive Iymphocytes. Discrimination between selfand non-self isnot complete (elicits autoimmune diseases). Discrimination I yes; norecognition of host cells or host cells between self andnonI express molecules that suppress innate self I immunity responses.Since thereisno complete discriminationbetween self and non-self by acquiredimmunity, we arealways at a risk ofdevel -oping autoimmune responses.(Adapted from re.f4 Abbas AK et al:分子細胞免疫学,2008)
⑨
戸
長
lム
洗 /
Z
s
-
-
(
晃
ぎ
に
Tヂ竺
Absence of antigen Antigen alonel
l
Non-responsiveness ofT cell (anergy) lNo T叫 response Antigen and B7-1/2l
T cell proliferation and differentiation Cytokine production, promotion of cell cycie and cell survivalFig. 2 Two signal theory in T cell activation
In order for the naive T cells to be activated and to become effector cells, not only a first signa,lbut also a second signal must be received. The first signal alone renders naive T cells anergic. (Adapted from ref. 5 Yagi J:Annual Review Kokyuki2007)
T cell response homology 100 33 29 15 12 (CTLA固め PD-1 BTLA T CD28 CD152 1C05 ?
+X+ +
+
、
〆
+
APC CD80 PDL-1 PDL・2 (B7-1) homology 100 26 27 25 25 13 Fig. 3 CD28/B7 family members and their interactions B7 family members expressed on APC that supply co-stimulation and their receptors on T cells, CD28 family members, are shown. Among CD28 family members, some receptors promote and suppress responses, and regulate immune responses. V, C, IgV-like, and IgC -like domain sequence homology is shown in humans. (Adapted from ref. 5 YagiJ:Annual Review Kokyuki 2007) てしまう.次に第1のシグナルと第2のシグナルを 受けても不応答性anergyが維持され,全く反応で きない(Fig.2)5).第2のシグナルは単一の分子から ではなく,複数のファミリーを形成する分子群が供 給することが分かつている (Fig.3).代表的な補助 (共)刺激受容体は CD28である. T細胞上のCD28 は.APC上の補助刺激分子B7-1/2(CD80/CD86)分 子と結合することでT
細胞に第2
のシグナルを供 給する.APCとして機能するマクロファージや未成 熟の樹状細胞は,外来異物を捕捉すると,おのおの 活性化ないし成熟して 自己抗原ペプチドを抗原ペ プチドと入れ替えて提示すると共にB7・1/2の表現 が誘導される.抗原特異的なT細胞は,第1と第2 のシグナルを受けて活性化して異物の排除にあた る.休止期のマクロファージや未成熟の樹状細胞は, 自己抗原ペプチドを提示しているがB7-1/2の発現 はないか極めて少なく, したがって第2
のシグナル を送ることができず,自己反応性T細胞自己反応性 は活性化することはなく.anergyが誘導される.一 方,中心性自己寛容を補う末梢性自己寛容も危険性 を内に秘めている.微生物による感染は, ときに自 己免疫疾患を誘導するが, 自己抗原ペプチドを提示 する APCに感染により B7-1/2が誘導される場合 を想定すれば自己免疫の引き金がヲl
かれる可能性を 一-6-容易に理解できょう.このように免疫システムは, 容易に生体に不都合な反応を起こし得るシステムと 言える.不都合な反応に対してはそれを制御する機 構を備えることで危険性を押さえている. 自己反応 性T細胞の活性化は制御性T細胞 (regulator yT-cells : Treg cells)が抑制することが明らかになって いる6) 微生物との共存においても次に記すように Treg cellsが重要な働きをすることが明らかになり つつある.粘膜免疫を例にあげる. 粘膜免疫 生体の粘膜は直接外界に接しながらおびただしい 数の抗原や微生物と接触している.生体防御の第一 線として病原微生物の侵入を阻止している一方,多 種多様な常在細菌が存在し,お互いに影響しながら 共存している.近年,病原微生物に対しては排除す るものの,本来non-selfであり免疫システムの基本 からいえば排除の対照である常在細菌に対しては排 除せず共存する機構が明らかにされつつある. 粘膜免疫の代表的な場としては,鼻咽頭関連リン パ組織 (nasopharynx-associatedlymphoid tissues : NALT)や腸管関連リンパ組織 (gut-associate dlym-phoid tissues : GALT)があげられる.GALTは小腸 に散在する.腸内フローラの優性菌の一つである Clostridiumに関する最近の知見7)によると.Clos-Clostridium clusters IV and XIVa Othertissues nTreg¥
l
・
ノ
Colon Thymus-derived nTreg cells ThymusFig. 4 Induction of IL-IO-producing Treg cells by interactionbetween Clostridium and epithelial cells
After weaning, Clostridium clusters N and沼Vabecome prominent in the large intes
-tine and promote the production of matrix metallo proteases (MMPs) from epithelial cells. MMPs then promote the production of the active form of the tumor growth factor (TGF)-s. In cooperation with indoleamine2, 3-dioxygenase (IDO) that is produced by epithelial
cells, the active form of TGF-s makes non Treg cells differentiate intoIL-lO-produingand
CTLA-4-expressing iTreg cells. Locally differentiated iTreg cells suppress inflammatory and allergic responses in the intestine and presumably other tissues. In contrast, thymus -derived nTreg cells do not require any bacteria-mediated (intestinal flora) stimuli.(Adapted from re 7 f. Kunisawa J et al: Cell Host Microbe 9:83-84, 2011) tridiumは腸管上皮からのメタロプロテアーゼ誘導 を 介 し て , サ イ ト カ イ ン 腫 蕩 増 殖 因 子 (tumor growth factor-s: TGF-s)の産生を促進する.
TGF-p
は免疫応答特に自己免疫を抑制することで注目さ れている Tregcellsを誘導するサイトカインであ り,腸管内において Tregcellsが誘導される Treg cellsは,異物である Clostridiumに対する排除を抑 制することでClostridiumは腸内フローラとして定 着し共存することとなる(Fig.4).このようにわれわ れの免疫システムは 免疫のセントラルドグマであ る異物排除という役割においても,必要に応じては それを制御して異物を受け入れる融通性を発揮して いる.粘膜免疫については 腸内フローラのセグメ ン ト 細 菌 (segmentedfilamentous bacteria: SFB)と呼ばれる細菌群が逆に自己免疫に関与する Th17細胞を誘導して関節炎などをきたす8)ことも 報告されている.腸内フローラにおいて細菌の種類 の異常な偏向がメタボリック症候群の発症と関連す ることもよく知られている. -7-おわりに 今まで見てきたように,われわれの免疫システム は, non-selfを排除するという観点からは,非常に厳 格にできているわけではない.末梢性トレランスの 維持は,感染などにより自己ペプチドの提示細胞が 補助刺激分子の発現を上昇すれば,破綻する可能性 がある.その危険を Tregcellsが押さえ込むように できている.すなわち,いかに免疫が自身の反応を 制御するかが,われわれの免疫システムが機能する か否かを決めている.この制御により non-selfであ る微生物の共存を許し お互いに利益を得ている.1
9
6
0
年代ないし7
0
年 代 に は 免 疫 シ ス テ ム はidio-type networkとかnetworkand curicuitと い う 概
念が提唱されていた.種々の免疫担当細胞はお互い に制御し合い,免疫システムは一つの大きな平衡系 を作っていることは近年より明確になってきてい る. 文 献 1) Medzhitov R, Preston-Hurlburt P, Janeway CA Jr:A human homologue of theDrosophilaToll pro
-teinsignalsactivationof adaptive immunity. Na -ture 388: 394-397, 1997
2) Kawai T, Akira S: Toll-like receptors and their crosstalk with other innate receptors in infection
and immunity. Immunity 34: 637-650, 2011
3) Franchi L, Warner N, Viani K et al: Function of Nod-like receptors in microbial recognition and host defense. Immunol Rev 227: 106-128, 2009
4) Abbas AK, Lichtman AH, Pober JS :自然免疫.
「分子細胞免疫学
J
,松島綱治ほか監訳, pp299,エルゼピアジャパン,東京 (2008)
5)八 木 淳 ニ:T細 胞 の 活 性 化 機 構 .
I
Annual review呼吸器
J
,pp20-21,中外医学社,東京 (2007) 6) Kronenberg M, Rudensky A: Regulation ofimmu-nity by self-reactiveT cells. Nature 435: 598-604, 2005 7) Kunisawa J, Kiyono H: Peaceful mutualism in the gut:revealing key commensal bacteria for thecrea -tion and maintenance of immunological homeosta -sis. Cell Host Microbe 9: 83-84, 2011 8) Wu HJ, Ivanov 11, Darce J et al: Gut-residing seg -mented filamentous bacteria drive autoimmune ar -thritis via T helper 17 cells. Immunity 32: 815-827, 2010 免 疫 ー 掲 載 予 定 一 執筆者 所属 テーマ 掲載号 八木淳二 微生物学免疫学 1. 自然免疫と獲得免疫 85(1) 有 賀 淳 先端生命医科学研究所 2.腫蕩と免疫ーがんワクチンー 85(2) 勝又康弘 リウマチ内科 3.自己免疫性疾患の最近の知見 85(3) 清水京子 消化器内科 4. IgG4関連疾患 85(4) 5.アレルギー疾患と免疫異常 槍垣裕子 皮膚科 ①アトピー性皮膚炎 85(5) 野 中 学 耳鼻咽喉科 ②アレルギー性鼻炎,副鼻腔炎,気管支端息 85(5) 田中柴ー リウマチ内科 6.免疫学的治療法としての生物学的製剤 85(6)