東北医科薬科大学 審査学位論文(博士)
氏名(本籍) オオウチ リュウスケ
大内 竜介(宮城県)
学位の種類 博士(薬学)
学位記番号 博薬学第9号
学位授与の日付 平成31年3月8日
学位授与の要件 学位規則第4条1項該当
学位論文題名
幼少期ストレスによる成人喘息発症機序の解明を目指した モデルマウスを用いた研究
-免疫寛容の誘導に対する幼少期ストレスの影響と成人喘 息発症-
論文審査委員
主査 教 授 井ノ口 仁 一
副査 教 授 丹 野 孝 一
副査 教 授 高 橋 知 子
幼少期ストレスによる成人喘息発症機序の解明を目指した モデルマウスを用いた研究
―免疫寛容の誘導に対する幼少期ストレスの影響と成人喘息発症―
東北医科薬科大学大学院 薬学研究科薬学専攻博士課程 病態生理学教室
大内竜介
目次
Ⅰ.研究背景 ・・・・・・・・・・・・・・・・・・・・・・・・・・ 1
Ⅱ.研究方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・ 4
Ⅲ.研究結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・11
Ⅳ.考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・15
Ⅴ.謝辞 ・・・・・・・・・・・・・・・・・・・・・・・・・・19
Ⅵ.参考文献 ・・・・・・・・・・・・・・・・・・・・・・・・・・20
Ⅶ.Figure ・・・・・・・・・・・・・・・・・・・・・・・・・・27
1
Ⅰ.研究背景
気管支喘息は、世界中で3億人以上の患者がいると推定される慢性炎症性疾 患である。1) 喘息の生理学的な特徴は、気道過敏性の亢進や可逆的な気道狭窄 であり、持続する気道炎症は気道閉塞やそれに引き続く組織リモデリングを惹 起し気流制限をもたらす。喘鳴、息切れ、胸部絞扼感などの喘息症状は2型ヘ ルパーT(Th2)細胞型免疫応答が優位となり、気管支平滑筋の収縮、粘膜浮腫、
粘液分泌・産生亢進による気道狭窄が生じることにより惹起される。2-6) 一般に 生体内に侵入した抗原は、樹状細胞(Dendritic cell: DC)などの抗原提示細胞に 貪食され、その一部がナイーブT(Th0)細胞に提示される。抗原提示を受け、
活性化したTh0はそれぞれT-bet、GATA3、Foxp3、RORγtを主要な転写因子と する1型ヘルパーT(Th1)細胞、Th2細胞、制御性T(Treg)細胞、17型ヘル パーT(Th17)細胞に分化する。Th2細胞が産生するサイトカインのうち、イン ターロイキン(Interleukin; IL)‐5は好酸球の骨髄での分化および増殖、循環血 中での活性化、気道局所への遊走に深く関与しており、IL-4およびIL-13はB 細胞に作用してIgE抗体産生を亢進する。7)
一方で、無害な抗原に対し過剰なあるいは不必要な免疫応答を抑制する機構 として免疫寛容がある。この機構ではTregが中心的な役割を担っており、8-11) その分化誘導にはIL-10やTGF-βなどが関連している。12, 13) 健常者と比べ、喘 息患者では末梢血中や喀痰中のTreg数の減少や活性の低下が認められる。14) さらに気管支肺胞洗浄液(Bronchoalveolar Lavage Fluids: BALFs)中のTreg数 においては小児喘息患者、成人喘息患者どちらも共通して低下している。15, 16)
2
Treg数の低下は免疫バランスの抑制性の制御が欠如を引き起こし、無害抗原に 対するTh2型免疫反応の亢進による気道炎症を惹起する。、12, 17, 18)
喘息の発症には個体因子と環境因子が相互に関連しており、個体因子として 遺伝子素因、アレルギー素因、性差など、環境因子としてアレルギー物質や感 染症、空気汚染物質、薬、そして精神的ストレスなどがあげられる。7, 58) 加え て、免疫寛容の障害はアレルギー性喘息罹患率の増加に寄与することが報告さ れている。例えば幼少期のライノウイルスやRSウイルスへの感染が成人喘息の 発症リスクとなることが示唆されており、59) 特に寛容誘導期にライノウイルス 感染することにより免疫寛容の誘導やTregの分化誘導が障害される報告がある。
19) また、RSウイルス感染によりTregの表現型がGATA3発現、Th2サイトカイ ン産生といったIL-4依存型に変化し、Tregの抑制性機能の減弱や免疫寛容の誘 導が阻害され、20) さらにTregへのIL-33暴露によって表現型がTh2様へ変化し、
結果として寛容誘導マウスに気道炎症を生じることが示されている。21) 加えて 寛容誘導マウスへのインドメタシン投与による PGI2 受容体シグナルの抑制は、
Tregに関係なくアレルギー物質誘因性の気道炎症を増大する報告がある。22) こ のように免疫寛容の障害によるアレルギー性喘息の罹患率増加には、複数の機 序や原因が関与することが示唆されている。
喘息発症の要因の一つである精神的ストレスについて、幼少期のストレス暴 露は成人喘息の発症リスク増加を伴うという報告がある。23, 24) 例えば、アフリ カ系アメリカ人女性において小児期や思春期の虐待体験が成人喘息発症と正の 相関があることが報告されている。25) さらに、小児期の虐待体験は喘息発症の
3
可能性を上昇させるという報告もされている。26) しかし、幼少期ストレスが成 人喘息を発症させる詳細な機序は不明である。
Tregによる免疫応答に対する制御の欠如について、さまざまな種類のストレ スがTregの分化誘導を障害することが示唆されている。例えば心的外傷後スト レス障害(Post Traumatic Stress Disorder: PTSD)患者の末梢血中Treg数は健常 人に比べ有意に低下しているという報告がある。27, 28) また、ヒトでの急性の実 験的ストレス要因は末梢血中のCD3+Foxp3+Treg数を減少させるという結果も出 ている。29) 私たちもこれまでに、成人期の精神的ストレスがTregの誘導を抑制 して、喘息の発症を引き起こすことをマウスモデルを用いて報告している。30)
以上のことより私は、幼少期の精神的ストレス暴露はTregの分化誘導を抑制 することにより免疫寛容の誘導を抑制し、Th2型への免疫反応の偏りとそれに引 き続くアレルギー喘息の発症を引き起こすと仮説をたて、検証のためアレルギ ー喘息モデルマウスをつかい、幼少期ストレスが免疫寛容に与える影響と喘息 発症について検討を行った。
4
Ⅱ.研究方法
1.マウス
生後4日齢(Postnatal Day 4: PND 4)のBALB/c雌性マウスを雌親と共に
CLEA Japan(Tokyo, Japan)から購入した。マウスは一定の室温下で、12 時間
ごとに明暗循環し滅菌済の飼料と水を自由に摂取できる環境下に収容した。す べての実験は東北医科薬科大学動物実験委員会の承認を得て実施し、実験動物 の苦痛や疼痛の緩和に充分配慮した。
2.母子分離
幼少期ストレスモデル作製のため、マウスに1日3時間の母子分離(Maternal Separation: MS)をPND 17からPND 22の6日間連続して行った(MSマウス)。 比較対象群としてMSを行わない群を作製した(非MSマウス)。はじめに雌親 をMSが終了するまでホームケージから別ケージへ移し、次に子マウスを別々 の容器に入れ孤立させた。MSの終了後、ホームケージには子マウスから戻し、
次に雌親を戻した。ヒトにおいて、ネグレクトなど幼少期のストレス体験は視 床下部‐下垂体‐副腎(HPA)軸の障害を引き起こし、不安うつ病などを惹起す るという報告があり、31, 32) 動物モデルにおいても、MSを含む幼少期ストレス の負荷はHPA軸の障害を引き起こし成長後のストレス負荷に対する応答能力の 低下や不安行動の増悪を惹起する。33, 34) したがって、MSはヒトにおける幼少 期ストレスを検討する目的で広く使用されているストレスモデルの一つである。
35)
5
3.ストレス暴露と寛容誘導、抗原感作と抗原暴露
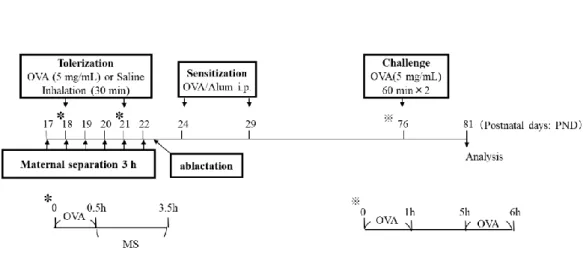
幼少期ストレスとして、マウスに1日3時間のMSを6日間連続(PND 17か
らPND 22)して負荷し、比較対象群としてMSを行わない群を作成した。免疫
寛容は卵白アルブミン(Ovalbumin: OVA)の吸入(PND 18とPND 21)により 誘導し、すべてのマウスはPND 23に離乳した。
PND 17よりストレス暴露と免疫寛容誘導を行い、その後抗原感作と抗原暴露
を行った(Fig. 1)。免疫寛容誘導は容器内で30分間エアロゾル化したOVA(5
mg/mL)を吸入をさせて行った。MS負荷群はOVA吸入後に負荷した。免疫寛
容誘導の比較対象群として生理食塩液のみの吸入群を作製した。作成した4群
(非MS/非寛容;喘息群、非MS/寛容;寛容群、MS/非寛容;ストレス群、MS/
寛容;ストレス+寛容群)すべてに抗原感作を行った。抗原感作は8 μgの OVAと4 mgの水酸化アルミニウム(Wako Pure Chemical Industries, Osaka,
Japan)の混合液をPND 24とPND 29に腹腔内投与した。抗原曝露はPND 76
にエアロゾル化したOVAを吸入させた。
4.BALF採取
Diethyl ether (和光純薬)麻酔下、眼窩より脱血後、マウスの胸部を開き気
管にカニューレを挿入、固定した後気管及び肺を一塊に摘出した。PBS(NaCl 137 mM, KCl 2.7 mM, Na2HPO4・12H2O 8.1 mM, KH2PO4 1.47 mM, pH 7.4)
(和光純薬) 0.25 mLをカニューレより注入し洗浄した後、回収してBALFと
6
した。BALFの回収は2回行った。回収したBALFは4℃、200 × gで3分間遠 心後、上清は量を測定して-80℃で保存した。上清回収後の細胞残渣は1% fetal bovine serum (FBS)(ICN Biomedicals, Icn., Aurora, OH, USA)含有PBS 0.2 mLで懸濁し、0.035 mLを0.1% trypan blue (和光純薬) 0.035 mLに添加し、
ヘモサイトメーターを用いて総白血球数を算定した。残りの0.165 mLをサイト スピン(Shandon, Runcorn, UK)にて400 rpmで5分間遠心し、細胞標本を作 製した。細胞標本を乾燥後、Diff-Quik染色液(国際試薬、神戸、日本)で染色 し、細胞分画を算定した(対物100倍で総細胞数200個以上観察)。
5.血漿中コルチコステロン測定
MS負荷前及びPND 17、 PND 19、 PND 22のMS負荷直後にMS負荷マウ ス、非MS負荷マウス共に血液を採取した。血液から分離した血漿は‐80℃で 保存しcorticosterone enzyme immunoassay kit(Enzo Life sciences, Farmingdale, NY)を用いて手順書に従いコルチコステロン濃度の測定を行った。検出限界は 32 pg/mlであった。
6.気道過敏性
気道過敏性の評価PND 81に行った。キシラジン(3 mg/kg)(Bayer Healthcare, Brussels, Belgium)とペントバルビタールナトリウム(40 mg/kg)
( Schering-Plough Animal Health Co, Kenilworth, N.J., USA)の混合物を腹腔 内投与することで麻酔し、咽頭部切開により気道を露出しカニューレを挿管し
7
た。その後マウスはFinePointe Resistance and Compliance (Buxco Electronics,
Troy, N.Y., USA)を使い、流速0.2 mL、呼吸数1分あたり150回の頻度で人工
的に呼吸させた。気道抵抗(RL)は、生理食塩液(ベースライン; 0 mg/mL)
および、メサコリン濃度0.625から5 mg/mLの範囲の吸入に対する反応を各3 分間測定し、各濃度における最大抵抗値により求めた。気道感受性は、ベース
ラインRL値が200 %上昇するのに必要なメサコリン濃度(PC200)を、用量反
応曲線を用いて算出することで検討した。
7.肺組織解析
PND 81に採取した肺を4%ホルマリン固定し、パラフィン(和光純薬)で包
埋した。包埋した肺を4 μmの厚さで切り出し、切片を作製した。切片は、脱パ ラフィン操作後、0.5%過ヨウ素酸水溶液で酸化し、Cold Schiff液(和光純薬)
で粘液を、ヘマトキシリン液にて核を染色し、脱水後、透徹、封入し、サンプ ルを得た。顕微鏡下対物40 倍の倍率で気道上皮細胞に占める粘液産生細胞の割 合を算定した。
8.OVA特異的IgE
96穴プレート(Nalge Nunc International, Neerijse, Belgium)にbicarbonate buffer(0.05 M, pH 9.7)で希釈したOVA (10 μg/mL)100 μL/wellを分注し4℃ で一晩静置した。その後、0.5% Tween20(SIGMA)/PBSで5回洗浄し、1%bov ine serum albumin(SIGMA)/PBS 200 μLを注入し、室温で1.5 時間静置、ブ
8
ロッキングを行った。0.5% Tween20/PBSで3回洗浄し、800 ~ 1600倍に希釈し た血清あるいは標準血清を100 μL/well添加した。37℃で2 時間静置し、0.5%
Tween20/PBSで5回洗浄後、10000倍希釈したhorseradish peroxidase-conjugated goat anti-mouse IgE antibody (Bethyl Laboratories Inc., Montogomery, TX, USA)を100 μL/well加え、室温で1.5時間静置した。5回洗浄後 tetramethylbenzidine (BioFX Laboratories, Owing Mills, MD, USA)を100 μL/
well添加30分後1N HCl 100 μL/wellで反応を停止させ、450 nmの吸光度を測 定した。OVAで感作したBALB/c系雌性マウスプール血清を標準血清として、
標準血清中に含まれるOVA特異的IgE量を1000 Uとし検量線を作成した。こ の検量線を用いて検体中のOVA特異的IgE量を決定した。
9.脾細胞培養
PND 76の抗原暴露前に脾臓を採取し、脾細胞を調整した。脾臓はステンレス
メッシュを用い細分化、50 mLコニカルチューブに回収し500 × g, 4℃で5分 間遠心し細胞を得た。その後、RBC Lysis Buffer(eBioscience, San Diego, CA)
を10 mL加えて正確に5分間室温で静置し、2% FBS RPMI1640 10 mLの添加 により溶血を止め、70 μmのナイロンメッシュに通し、2% FBS RPMI1640で洗 浄(500 × g, 4℃で5分間遠心)した。その後、10% FBS RPMI1640を10 mL で再懸濁し細胞数を計測した。
調整した脾細胞は1 × 106個ずつ平底96穴プレート(BD FALCON)に播種 し、OVA(200 μg/mL)と共に72時間培養した。その後、 water-soluble tetrazolium salt(WST-8)(nacalai tesque, Kyoto, Japan)を加え更に3時間培養
9
し、マイクロプレートリーダーを用いて、生成したホルマザン塩の吸光度(450
nm)を測定し、OVAに対する増殖活性を評価した。また、回収した上清はサ
イトカインの測定に使用した。
10.気管支リンパ節細胞調整
PND 22に気管支リンパ節(Bronchial Lymph Nodes: BLN)をすでに報告され ている方法で採取した。30) BLN細胞は後述のフローサイトメトリー解析を用い て解析した。
11.フローサイトメトリー解析
採取したBLN細胞の全量を10%FBSと50 μM 2‐メルカプトエタノール、
100 U/mLペニシリンG、100 U/mLストレプトマイシンを含むRPMI1640 1 mLに懸濁した。細胞内の染色のため、BLN細胞を5 μg/mL のphorbol 12-myristate 13-acetate(PMA)、500 ng/mLのionomycin、2 μMのmonesinで4 時間刺激した。刺激後、非特異的結合を低減するため、BLN細胞をCD16/CD32 抗体(FC gamma Ⅲ/Ⅱ receptor; BD Biosciences, Franklin Lanes, NJ)を添加し 培養した。死細胞を判別するためLIVE/DEAD Fixable Blue Dead Cell Stain Kit(Thermo Fisher scientific, Waltham, MA)を使用した。BLN細胞表面染色の ため、まずFITC標識したCD3ε抗体(clone 145-2C11; Miltenyi Biotec, Bergisch Gladbach, Germany)とAlexa Fluor 700標識したCD4抗体(clone RM4-5; BD biosciences)もしくはアイソタイプ抗体で染色した。細胞固定と透
10
過処理の後、細胞内染色のためPE-Cy7標識したFoxp3抗体(clone FJK-16s;
Thermo Fisher Scientific)もしくはアイソタイプ抗体で染色した。本研究では CD3+CD4+Foxp3+細胞をTregとして扱った。細胞カウントにはFACSAriaⅡ(BD Biosciences)を用い、解析にはFACSDiva software(BD Biosciences)を用いた。
12.サイトカイン量測定
BALF中及び脾細胞培養上清中のIL-4、IL-5、IL-13、IFN-γ濃度はELISA Kits(Thermo Fisher Scientific)を用い手順書に従い測定した。検出限界はそれ ぞれIL-4が2 pg/mL、IL-5が7 pg/mL、IL-13が1.5 pg/mL、IFN-γが8 pg/mL であった。
13.統計解析
データは平均値±標準偏差(Standard Deviation: SD)で示す。2群間の有意差 検定はノンパラメトリックなマンホイットニーのU検定で解析した。多群間の 比較の際はvariance and Bonferroni’s post-hoc検定を行った。これらの統計解析 はPrism 6(GraphPad Software, San Diego, CA)を用いて行った。P値は0.05 未満を有意差ありとした。
11
Ⅲ.研究結果
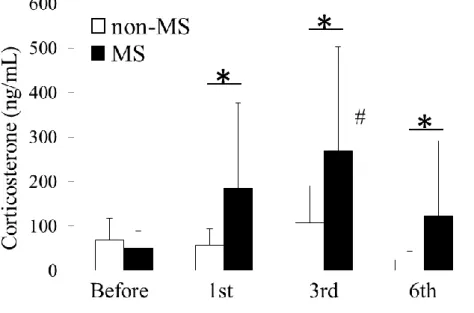
1.MSは血漿中コルチコステロン濃度を上昇させる
MSの効果を確認するため、PND 17のMS負荷前及びMS負荷直後とPND
19、 PND 22のストレス負荷直後にMS負荷マウス、非MS負荷マウスよりそ
れぞれ血液を採取し血漿中コルチコステロン濃度を測定した。MS負荷後のMS 負荷マウスの血漿中コルチコステロン濃度は非MS負荷マウスにくらべPND
17、 PND 19、 PND 22いずれにおいても有意に高値であった(PND 17: 185
± 191.1 [n=6] vs. 55.9 ± 38.1 [n=6], P < 0.05; PND 19: 267.8 ± 234.9 [n=6]
vs. 106.4 ± 83.9 [n=7], P < 0.05; PND22: 122.3 ± 168.7 [n=7] vs. 24.1 ± 19.8 [n=6], P < 0.05)。MS負荷前では、MS負荷マウスと非MS負荷マウスの血 漿中コルチコステロン濃度に有意な差は認められなかった(Fig.2)。
2.免疫寛容誘導は気道過敏性を低下させMSは免疫寛容誘導の効果を抑制する MSと免疫寛容誘導が、気道反応に与える影響を検討するためまずメサコリン 吸入による気道過敏性の測定を行った。予想した通り、寛容群(n=7)の気道反 応性(RL)は喘息群(n=6)に比べ有意に低値であった(methacholine concentrations of 0.625 mg/mL: 1.37 ± 0.21 vs. 2.01 ± 0.57, P < 0.05; 1.25 mg/mL: 1.57 ± 0.38 vs. 3.37 ± 1.24, P < 0.01; 2.5 mg/mL: 1.92 ± 0.46 vs.
6.79 ± 2.25, P < 0.01; 5 mg/mL: 3.17 ± 1.2 vs. 6.85 ± 1.96, P < 0.01)(Fig.
3A)。同様に、寛容群の気道過敏性(PC200)は喘息群に比べ有意に高値であっ た(3.60 ± 0.87 vs. 1.18 ± 0.25, P < 0.01)(Fig. 3B)。ストレス+寛容群(n=5)
の気道反応性と気道過敏性はどちらも寛容群に比べ有意に高値であり(RL, 1.25
12
mg/mL of methacholine: 3.15 ± 1.85 vs. 1.57 ± 0.38, P < 0.05; 2.5 mg/mL:
7.80 ± 4.24 vs. 1.92 ± 0.46, P < 0.01; 5 mg/mL: 6.13 ± 3.28 vs. 3.17 ± 1.20, P < 0.05; Fig. 3A; PC200; 1.44 ± 0.61 vs. 3.6 ± 0.87, P < 0.01; Fig. 3B)、MS は免疫寛容誘導による気道過敏性の抑制効果を完全に無効にした。さらに気道 反応性と気道過敏性はどちらも、ストレス+寛容群とストレス群(n=3)におい て統計的有意差はなかった。これは、MSの気道過敏性に対する効果は寛容誘誘 導への影響を介していることを示している。
3.免疫寛容誘導はOVA特異的IgE産生、気道粘液分泌を低下させMSは免疫寛 容誘導の効果を抑制する
MS、免疫寛容誘導がOVA特異的IgE産生に与える影響を検討するためPND
81の血漿中OVA特異的IgEの測定を行ったところ、寛容群では喘息群に比べ有 意に低値であった(627.2 ± 519.6 EU/mL [n=8] vs. 2987.8 ± 1049.7 EU/mL
[n=8], P < 0.001)。さらに、ストレス+寛容群では寛容群に比べ有意に高値であ
った(1501.3 ± 1004.0 EU/mL [n=6] vs. 627.2 ± 519.6 EU/mL [n=8], P <
0.05)。そのうえ、ストレス+寛容群とストレス群では統計的有意差は認められ
ず、喘息群とストレス群においても同様に統計的有意差はなかった(Fig. 4A)。
次に、MSと免疫寛容誘導が気道粘液分泌に与える影響を検討するためPAS 陽性細胞数の算定を行った。寛容群は喘息群に比べ有意に低値であり、ストレ ス+寛容群では寛容群に比べ有意に高値であった(48.6 ± 12.2% [n=6] vs.
27.6 ± 10.6% [n=8], P < 0.05)。ストレス+寛容群とストレス群では統計的有意 差は認められず、喘息群とストレス群でも同様に統計的有意差はなかった(Fig.
13
4B)。これらは、MSのOVA特異的IgE産生、粘液分泌に対する効果は免疫寛
容誘導への影響を介していることを示している。
4.免疫寛容誘導は気道炎症、Th2サイトカイン分泌を低下させMSは免疫寛容 誘導の効果を抑制する
MSと免疫寛容誘導が気道炎症、Th2サイトカイン分泌に与える影響を検討す
るためPND 81に採取したBALF中炎症性細胞数、サイトカイン含量について
検討を行った。予想した通り、寛容群では喘息群にくらべ総細胞数、好酸球数、
リンパ球数の有意な低下が認められた。免疫寛容誘導を行ったマウスでは好中 球の有意な増加が認められたが、非寛容誘導マウスでは増加は認められなかっ た。BALF中Th2サイトカインにおいても炎症性細胞数と同様の結果であった
( Fig. 6)。BALF中のIFN-γ濃度については4群とも差は認められず、ストレ ス群と喘息群の気道炎症、Th2サイトカイン産生ともに差はなかった。これらの 結果は、MSの気道炎症に対する効果は免疫寛容誘導への影響を介していること を示している。
5.MSは免疫寛容誘導の効果を抑制する
以上の結果から、MS負荷によって免疫寛容誘導マウスにおいても抗原反応性 の気管支喘息症状が惹起されることが分かったため、免疫寛容の誘導に対する MSの影響について検討を行った。全身性免疫を示す指標としてのin vitroでの OVA刺激による脾細胞の増殖性は免疫寛容誘導を行ったマウスで抑制された。
14
しかし、免疫寛容誘導期にMSを負荷したマウスでは脾細胞増殖性は非MS負荷 マウスに比べ有意に増加していた。また、ストレス+寛容誘導群と喘息群、ス トレス群では脾細胞増殖性に差は認められなかった。つまり、MSは免疫寛容非 誘導下では脾細胞の増殖性に影響を与えなかった(Fig. 7A)。
さらに、4群から採取した脾細胞でのin vitroでのOVA刺激によるサイトカ イン分泌(IL-4、IFN-γ)について検討を行った(Fig. 7B及び7C)。免疫寛容誘 導は非MS負荷の条件下において有意にIL-4分泌を低下させたがMS負荷によ り有意な増加回復が認められた。ストレス+寛容群及び免疫寛容を誘導しなか ったマウスではIL-4分泌は同程度であった(Fig. 7B)。免疫寛容誘導はMSの 有無にかかわらず、IFN-γ分泌へは影響を与えなかった(Fig. 7C)。非免疫寛容 誘導の条件下においては、MSはサイトカイン分泌に影響を与えなかった。
6.MSは気管支リンパ節中Tregを減少させる
これまでの結果よりMSは免疫寛容誘導を抑制することがわかったため、次 にBLN中のFoxp3+Tregについて調査を行った。BLN中のFoxp3+Tregは喘息群 に比べ寛容群では有意に増加がみられた。また、ストレス+寛容群の
Foxp3+Tregは寛容群に比べ有意に低下していた。ストレス群と喘息群では
Foxp3+Tregに違いは見られず、MSは非寛容下ではFoxp3+Treg分化誘導に影響
を与えなかった。加えて、ストレス群とストレス+寛容群では差は見られなか
った(Fig. 8)。以上より、MSはTregの分化誘導の抑制を介し免疫寛容誘導に
影響を与える。
15
Ⅳ.考察
本研究では、幼少期の精神的ストレスとしてのMSを免疫寛容誘導時期に負 荷することにより、成人期のアレルギー物質の吸入によってTh2型アレルギー 性気道炎症が引き起こされることを示した(Fig. 3-6)。まず、抗原特異的脾細胞 の増殖とIL-4分泌の測定によってMS負荷による免疫寛容誘導への影響を確か めたところ、どちらも寛容群にて抑制されていた。さらに、ストレス+寛容群 では寛容群に比べBLN中のTregの減少がみられた。したがって、免疫寛容誘導 期のMS負荷はTreg分化の抑制とそれに引き続くTh2型免疫応答への偏りを引 き起こすと考えられる。
幼少期の経粘膜的な抗原暴露により免疫寛容が誘導されることが知られてお り、例えばマウスモデルにおいてPND 7でのOVA吸入によって、抗原暴露後 のBALF中の顆粒球、IL-13を減少させ、成人期のOVA特異的IgE産生抑制を 示すという報告がある。36) またヒトにおいて、免疫寛容を含む生得的及び適応 性免疫は幼少期に成熟する37) とされており、その時期の精神的ストレス暴露は 免疫システムの発達に大きく影響を与える。38, 39) 本研究の結果でも、マウスで の幼少期ストレスは免疫寛容の誘導を抑制し気道炎症を増強することが明らか になった。ヒトにおいても幼少期ストレスとなる虐待は、厚生省が公表する「児 童相談所での児童虐待相談対応件数とその推移」をみると年々増加しているこ とがわかる。40) その主な増加要因としては心理的虐待に係る相談対応件数の増 加が指摘されており、虐待を受けた子どもの相談内容についても心理的虐待の
割合が54.0%ともっとも多かった。40) 今回幼少期精神的ストレスを負荷した17
日齢から22日齢マウスはヒト換算で1~2歳と考えられている。41) 厚生省の報
16
告をふまえて、幼少期ストレスが免疫寛容誘導に与える影響の検討は成人期喘 息発症のメカニズム解明の一助となりえる。
BALF中の炎症性細胞の結果より、寛容群に比べストレス+寛容で好中球の有 意な増加が見られた点について、in vitroにおいて活性化された好中球が好酸球 の遊走に関連するという報告がある。42, 43) したがって、本研究においてもスト レス+寛容群で増加した好中球が、好酸球の遊走に影響を与えた可能性がある。
また、主にTh17細胞によって産生されるIL-17Aは、重症喘息での好中球気性 道炎症の誘発において重要な役割を果たすことが実証されている。44, 45) 私たち も以前、拘束ストレスが好酸球のみならず好中球の気道浸潤を誘発し、また寛 容群に比べてストレス+寛容群IL-17AとTh17細胞の増加を伴い肺への好中球 の集積がみられたことを報告している。30) したがって、本研究でのストレス+
寛容群においてもMS曝露によるTh17細胞の分化誘導およびIL-17Aの産生亢 進が、好中球の浸潤に関与する可能性がある。
MSによるTreg分化誘導への抑制については、ヒトにおいて外傷的、精神的 ストレスが末梢血中Treg数を減少させるという報告があるものの、27, 29, 46) ス トレス暴露がTreg分化に影響するメカニズムははっきりしていない。精神的ス トレスに対する生体反応として、視床下部‐下垂体‐副腎軸の賦活化によるス トレスホルモンであるグルココルチコイド(GC)の分泌があり、この経路がス トレスの免疫寛容に対する抑制性の影響に重要な役割を果たしている可能性が ある。GCは抗アレルギー活性を有し、喘息の予防、治療に用いられている。
GCはT細胞と上皮細胞からのサイトカイン産生を制限し気道炎症と気道過敏性 の低下させ、また好酸球や他の炎症性細胞の動員、賦活化を抑制する。2, 47) 一
17
方で、GCはTh2型免疫反応の亢進作用を示すことが知られており、例えばGC が抗原提示細胞からのIL-12分泌を減少させる報告48) やGCによる直接的な Th1細胞の抑制作用によってTh2細胞分化を増強する報告、49) アレルギーモデ ルマウスの抗原感作期に一過性にGC投与を行うことによりTh2反応を増強す る報告がある。50) 加えて、寛容誘導期にGC投与を行うとTregからのIL-10分 泌に必要とされるIL-10産生樹状細胞の減少と抗原暴露によるTh2型免疫応答へ の偏りがみられる。51) 興味深いことに、抗原の吸入など経気道的な寛容誘導は GCの局所投与ではなく全身投与によって阻害されることが報告されている。52) これは本研究での、MSによって免疫寛容誘導が抑制されたマウスにおいて血 中GCが増加するという結果と一致している(Fig. 2)。さらに重要なことに、
ヒトおよびマウスへのGC全身投与は血中Foxp3+Tregの機能的・数的な抑制を 示し、53) 局所性のGC投与はアレルギー性炎症の抑制を示すという報告がある。
54) 加えて、私たちは以前に成人期のマウスに精神的ストレスとして拘束ストレ スを負荷するとGCの作用によりTreg分化誘導が阻害されるが、寛容誘導期の ストレス負荷前にGC受容体遮断薬を投与することによって精神的ストレス負 荷が免疫寛容誘導へ与える影響を抑制することを報告している。30)
免疫寛容の欠如に伴うアレルギー性喘息罹患率増加のメカニズムについての 研究について、幼少期のウイルス性呼吸器感染症が成人期の喘息発症のリスク 因子となることが知られている。マウスでは免疫寛容誘導と同時期のライノウ イルス感染による肺内樹状細胞でのOX40リガンド発現増加や、気道上皮細胞 においてIL-33および胸腺間質性リンパ球新生因子(thymic stromal
lymphopoietin: TSLP)の発現増加によって、結果として鼻腔内OVA投与による
18
免疫寛容の誘導が阻害されアレルギー性気道反応の増加が見られることが知ら れている。 19) また、OVA暴露した雌性マウスからの母乳摂取によってOVA寛 容化した子マウスにおいて幼少期に繰り返しRSウイルス感染することによっ
てFoxp3+TregがGATA-3発現、Th2サイトカイン産生といったTh2様に変化し、
抑制性の活性が消失して成人期のアレルギー性反応が増加するが、非感染のマ ウスではこのような反応は見られないという報告もある。20) さらに、感作時期 あるいは感作後の鼻腔内IL-33投与によってIL-33結合TregがGATA-3発現や Th2サイトカインの産生といったTh2様の性質を示し、抑制性の活性が消失し 抗原反応による気道炎症を惹起することが実験的に確認されている。21) 免疫寛 容に対する気道上皮細胞の影響については本研究とは別のアレルギーモデルマ ウスを用いて確認されており、例として気道上皮細胞でのNF-κB活性化は寛容 化マウスにおいて骨髄性樹状細胞の活性化を伴うアレルギー性気道炎症を引き 起こす報告がある。55) これらは、本研究のような精神的ストレス誘発の内分泌 やウイルス感染による炎症19, 20) が関与する、免疫寛容の抑制により惹起される 喘息に特徴的なメカニズムである可能性がある。
結論として、免疫の発達に重要な時期である幼少期に精神的ストレスとして のMSに暴露することによりTregの分化誘導が阻害され、経気道的な免疫寛容 の成立の抑制が生じ、結果としてTh2型免免疫応答への偏りによってアレルギ ー性喘息の罹患率を増加する。今後の研究では、ストレス暴露によるGC分泌に 関連した神経内分泌経路の解明や、ストレスによる喘息増悪でのオピオイドな どの神経ペプチドの作用56, 57) についての検討が必要となるだろう。
19
Ⅴ.謝辞
本研究を行うにあたり、終始御懇篤な御指導とご鞭撻を賜りました東北医科 薬科大学薬学部病態生理学教室 高橋知子教授ならびに同大学医学部医学教育 推進センター 大野勲教授に心より感謝申し上げます。また、研究に必要な知 識・技術から研究に対する姿勢まで幅広く御指導頂きました河野資先生に心よ り感謝いたします。東北医科薬科大学薬学部病態解析学教室 大河原雄一教授、
同大学薬学部病態生理学教室 宮坂智充先生をはじめ病態生理学教室の皆様か ら多くのご協力を頂きましたことに感謝いたします。
これまでに多くの方々のお力添えを賜り、ここに本研究の完成を迎えること ができました。今後もより一層研究に努め、患者さんの治療の助けとなるよう な研究者兼医療人となります。
20
Ⅵ.参考文献
1) To, T., Stanojevic, S., Moores, G., Gershon, A.S., Bateman, E.D., Cruz, A.A. &
Boulet, L.P. (2012) Global asthma prevalence in adults: findings from the cross-sectional world health survey. BMC Public Health, 12, 204.
2) Fahy, J.V. (2015) Type 2 inflammation in asthma — present in most, absent in many. Nat Rev Immunol, 15, 57-65.
3) Holgate, S.T., Wenzel, S., Postma, D.S., Weiss, S.T., Renz, H. & Sly, P.D. (2015) Asthma. Nat Rev Dis Primers, 1, 15025.
4) Kim, R.Y., Rae, B., Neal, R., Donovan, C., Pinkerton, J., Balachandran, L., Starkey, M.R., Knight, D.A., Horvat, J.C. & Hansbro, P.M. (2016) Elucidating novel disease mechanisms in severe asthma. Clin Trans Immunol, 5, e91.
5) Lambrecht, B.N. & Hammad, H. (2015) The immunology of asthma. Nat Immunol, 16, 45-56.
6) Rothenberg, M.E., Saito, H. & Peebles, R.S. (2017) Advances in mechanisms of allergic disease in 2016. Journal of Allergy and Clinical Immunology, 140, 1622-1631.
7) 喘息予防・管理ガイドライン2018
8) Martin-Orozco, E., Norte-Munoz, M. & Martinez-Garcia, J. (2017) Regulatory T Cells in Allergy and Asthma. Front Pediatr, 5, 117.
9) Palomares, O., Yaman, G., Azkur, A.K., Akkoc, T., Akdis, M. & Akdis, C.A.
(2010) Role of Treg in immune regulation of allergic diseases. Eur J Immunol, 40, 1232-1240.
10) Peterson, R.A. (2012) Regulatory T-cells: diverse phenotypes integral to immune homeostasis and suppression. Toxicol Pathol, 40, 186-204.
11) Sakaguchi, S., Miyara, M., Costantino, C.M. & Hafler, D.A. (2010) FOXP3+
regulatory T cells in the human immune system. Nat Rev Immunol, 10, 490-500.
21
12) Rivas, M.N. & Chatila, T.A. (2016) Regulatory T cells in allergic diseases. J Allergy Clin Immunol, 138, 639-652.
13) Palomares, O., Martin-Fontecha, M., Lauener, R., Traidl-Hoffmann, C.,
Cavkaytar, O., Akdis, M. & Akdis, C.A. (2014) Regulatory T cells and immune regulation of allergic diseases: roles of IL-10 and TGF-beta. Genes Immun, 15, 511-520.
14) Mamessier, E., Nieves, A., Lorec, A.M., Dupuy, P., Pinot, D., Pinet, C., Vervloet, D. & Magnan, A. (2008) T-cell activation during exacerbations: a longitudinal study in refractory asthma. Allergy, 63, 1202-1210.
15) Hartl, D., Koller, B., Mehlhorn, A.T., Reinhardt, D., Nicolai, T., Schendel, D.J., Griese, M. & Krauss-Etschmann, S. (2007) Quantitative and functional
impairment of pulmonary CD4+CD25hi regulatory T cells in pediatric asthma. J Allergy Clin Immunol, 119, 1258-1266.
16) Barczyk, A., Pierzchala, W., Caramori, G., Wiaderkiewicz, R., Kaminski, M., Barnes, P.J. & Adcock, I.M. (2014) Decreased percentage of
CD4(+)Foxp3(+)TGF-beta(+) and increased percentage of CD4(+)IL-17(+) cells in bronchoalveolar lavage of asthmatics. J Inflamm (Lond), 11, 22.
17) Fujita, H., Meyer, N., Akdis, M. & Akdis, C.A. (2012) Mechanisms of immune tolerance to allergens. Chem Immunol Allergy, 96, 30-38.
18) Lloyd, C.M. & Hawrylowicz, C.M. (2009) Regulatory T cells in asthma.
Immunity, 31, 438-449.
19) Mehta, A.K., Duan, W., Doerner, A.M., Traves, S.L., Broide, D.H., Proud, D., Zuraw, B.L. & Croft, M. (2016) Rhinovirus infection interferes with induction of tolerance to aeroantigens through OX40 ligand, thymic stromal lymphopoietin, and IL-33. J Allergy Clin Immunol, 137, 278-288.e276.
20) Krishnamoorthy, N., Khare, A., Oriss, T.B., Raundhal, M., Morse, C., Yarlagadda, M., Wenzel, S.E., Moore, M.L., Peebles, R.S., Jr., Ray, A. & Ray, P. (2012)
22
Early infection with respiratory syncytial virus impairs regulatory T cell function and increases susceptibility to allergic asthma. Nat Med, 18, 1525-1530.
21) Chen, C.C., Kobayashi, T., Iijima, K., Hsu, F.C. & Kita, H. (2017) IL-33
dysregulates regulatory T cells and impairs established immunologic tolerance in the lungs. J Allergy Clin Immunol, 140, 1351-1363.
22) Zhou, W., Goleniewska, K., Zhang, J., Dulek, D.E., Toki, S., Lotz, M.T., Newcomb, D.C., Boswell, M.G., Polosukhin, V.V., Milne, G.L., Wu, P., Moore, M.L., FitzGerald, G.A. & Peebles, R.S., Jr. (2014) Cyclooxygenase inhibition abrogates aeroallergen-induced immune tolerance by suppressing prostaglandin I2 receptor signaling. J Allergy Clin Immunol, 134, 698-705.
23) Ilmarinen, P., Tuomisto, L.E. & Kankaanranta, H. (2015) Phenotypes, Risk Factors, and Mechanisms of Adult-Onset Asthma. Mediators Inflamm, 2015, 514868.
24) Rosa, M.J., Lee, A.G. & Wright, R.J. (2018) Evidence establishing a link between prenatal and early-life stress and asthma development. Curr Opin Allergy Clin Immunol, 18, 148-158.
25) Coogan, P.F., Wise, L.A., O'Connor, G.T., Brown, T.A., Palmer, J.R. & Rosenberg, L. (2013) Abuse during childhood and adolescence and risk of adult-onset
asthma in African American women. J Allergy Clin Immunol, 131, 1058-1063.
26) Scott, K.M., Smith, D.A. & Ellis, P.M. (2012) A population study of childhood maltreatment and asthma diagnosis: differential associations between child protection database versus retrospective self-reported data. Psychosom Med, 74, 817-823.
27) Jergovic, M., Bendelja, K., Vidovic, A., Savic, A., Vojvoda, V., Aberle, N., Rabatic, S., Jovanovic, T. & Sabioncello, A. (2014) Patients with posttraumatic stress disorder exhibit an altered phenotype of regulatory T cells. Allergy Asthma Clin Immunol, 10, 43.
23
28) Loerbroks, A., Apfelbacher, C.J., Thayer, J.F., Debling, D. & Sturmer, T. (2009) Neuroticism, extraversion, stressful life events and asthma: a cohort study of middle-aged adults. Allergy, 64, 1444-1450.
29) Freier, E., Weber, C.S., Nowottne, U., Horn, C., Bartels, K., Meyer, S., Hildebrandt, Y., Luetkens, T., Cao, Y., Pabst, C., Muzzulini, J., Schnee, B., Brunner-Weinzierl, M.C., Marangolo, M., Bokemeyer, C., et al. (2010) Decrease of CD4(+)FOXP3(+) T regulatory cells in the peripheral blood of human
subjects undergoing a mental stressor. Psychoneuroendocrinology, 35, 663-673.
30) Kawano, T., Ouchi, R., Ishigaki, T., Masuda, C., Miyasaka, T., Ohkawara, Y., Ohta, N., Takayanagi, M., Takahashi, T. & Ohno, I. (2018) Increased
Susceptibility to Allergic Asthma with the Impairment of Respiratory Tolerance Caused by Psychological Stress. Int Arch Allergy Immunol, 177, 1-15.
31) Tarullo, A.R. & Gunnar, M.R. (2006) Child maltreatment and the developing HPA axis. Horm Behav, 50, 632-639.
32) Maniam, J., Antoniadis, C. & Morris, M.J. (2014) Early-Life Stress, HPA Axis Adaptation, and Mechanisms Contributing to Later Health Outcomes. Front Endocrinol (Lausanne), 5, 73.
33) Murgatroyd, C. & Spengler, D. (2011) Epigenetic programming of the HPA axis:
early life decides. Stress, 14, 581-589.
34) Nishi, M., Horii-Hayashi, N. & Sasagawa, T. (2014) Effects of early life adverse experiences on the brain: implications from maternal separation models in rodents. Front Neurosci, 8, 166.
35) Tractenberg, S.G., Levandowski, M.L., de Azeredo, L.A., Orso, R., Roithmann, L.G., Hoffmann, E.S., Brenhouse, H. & Grassi-Oliveira, R. (2016) An overview of maternal separation effects on behavioural outcomes in mice: Evidence from a four-stage methodological systematic review. Neurosci Biobehav Rev, 68, 489-503.
24
36) Wang, Y. & McCusker, C. (2006) Neonatal exposure with LPS and/or allergen prevents experimental allergic airways disease: development of tolerance using environmental antigens. J Allergy Clin Immunol, 118, 143-151.
37) Lloyd, C.M. & Marsland, B.J. (2017) Lung Homeostasis: Influence of Age, Microbes, and the Immune System. Immunity, 46, 549-561.
38) Bosquet Enlow, M., King, L., Schreier, H.M., Howard, J.M., Rosenfield, D., Ritz, T. & Wright, R.J. (2014) Maternal sensitivity and infant autonomic and
endocrine stress responses. Early Hum Dev, 90, 377-385.
39) McLaughlin, K.A., Sheridan, M.A., Tibu, F., Fox, N.A., Zeanah, C.H. & Nelson, C.A., 3rd (2015) Causal effects of the early caregiving environment on
development of stress response systems in children. Proc Natl Acad Sci U S A, 112, 5637-5642.
40) 厚生労働省 子ども虐待による死亡事例等の検証結果等について 第14 次報告
41) 佐藤徳光 動物実験の基本 新訂版
42) Kikuchi, I., Kikuchi, S., Kobayashi, T., Hagiwara, K., Sakamoto, Y., Kanazawa, M. & Nagata, M. (2006) Eosinophil trans-basement membrane migration induced by interleukin-8 and neutrophils. Am J Respir Cell Mol Biol, 34, 760-765.
43) Nishihara, F., Nakagome, K., Kobayashi, T., Noguchi, T., Araki, R., Uchida, Y., Soma, T. & Nagata, M. (2015) Trans-basement membrane migration of
eosinophils induced by LPS-stimulated neutrophils from human peripheral blood in vitro. ERJ Open Res, 1.
44) Chesne, J., Braza, F., Mahay, G., Brouard, S., Aronica, M. & Magnan, A. (2014) IL-17 in severe asthma. Where do we stand? Am J Respir Crit Care Med, 190, 1094-1101.
25
45) Ray, A. & Kolls, J.K. (2017) Neutrophilic Inflammation in Asthma and Association with Disease Severity. Trends Immunol, 38, 942-954.
46) Sommershof, A., Aichinger, H., Engler, H., Adenauer, H., Catani, C., Boneberg, E.M., Elbert, T., Groettrup, M. & Kolassa, I.T. (2009) Substantial reduction of naive and regulatory T cells following traumatic stress. Brain Behav Immun, 23, 1117-1124.
47) John, M., Lim, S., Seybold, J., Jose, P., Robichaud, A., O'Connor, B., Barnes, P.J.
& Chung, K.F. (1998) Inhaled corticosteroids increase interleukin-10 but reduce macrophage inflammatory protein-1alpha, granulocyte-macrophage
colony-stimulating factor, and interferon-gamma release from alveolar macrophages in asthma. Am J Respir Crit Care Med, 157, 256-262.
48) DeKruyff, R.H., Fang, Y. & Umetsu, D.T. (1998) Corticosteroids enhance the capacity of macrophages to induce Th2 cytokine synthesis in CD4+ lymphocytes by inhibiting IL-12 production. J Immunol, 160, 2231-2237.
49) Miyaura, H. & Iwata, M. (2002) Direct and indirect inhibition of Th1
development by progesterone and glucocorticoids. J Immunol, 168, 1087-1094.
50) Wiley, R.E., Cwiartka, M., Alvarez, D., Mackenzie, D.C., Johnson, J.R., Goncharova, S., Lundblad, L. & Jordana, M. (2004) Transient corticosteroid treatment permanently amplifies the Th2 response in a murine model of asthma.
J Immunol, 172, 4995-5005.
51) Stock, P., Akbari, O., DeKruyff, R.H. & Umetsu, D.T. (2005) Respiratory tolerance is inhibited by the administration of corticosteroids. J Immunol, 175, 7380-7387.
52) Kerzerho, J., Wunsch, D., Szely, N., Meyer, H.A., Lurz, L., Rose, L., Wahn, U., Akbari, O. & Stock, P. (2012) Effects of systemic versus local administration of corticosteroids on mucosal tolerance. J Immunol, 188, 470-476.
26
53) Sbiera, S., Dexneit, T., Reichardt, S.D., Michel, K.D., van den Brandt, J., Schmull, S., Kraus, L., Beyer, M., Mlynski, R., Wortmann, S., Allolio, B., Reichardt, H.M.
& Fassnacht, M. (2011) Influence of short-term glucocorticoid therapy on regulatory T cells in vivo. PLoS One, 6, e24345.
54) Olsen, P.C., Kitoko, J.Z., Ferreira, T.P., de-Azevedo, C.T., Arantes, A.C. &
Martins Mu, A. (2015) Glucocorticoids decrease Treg cell numbers in lungs of allergic mice. Eur J Pharmacol, 747, 52-58.
55) Ather, J.L., Foley, K.L., Suratt, B.T., Boyson, J.E. & Poynter, M.E. (2015) Airway epithelial NF-κB activation promotes the ability to overcome inhalational
antigen tolerance. Clin Exp Allergy, 45, 1245-1258.
56) Drolet, G., Dumont, E.C., Gosselin, I., Kinkead, R., Laforest, S. & Trottier, J.F.
(2001) Role of endogenous opioid system in the regulation of the stress response.
Prog Neuropsychopharmacol Biol Psychiatry, 25, 729-741.
57) Miyasaka, T., Dobashi-Okuyama, K., Takahashi, T., Takayanagi, M. & Ohno, I.
(2018) The interplay between neuroendocrine activity and psychological stress-induced exacerbation of allergic asthma. Allergol Int, 67, 32-42.
58) Rod NH, Kristensen TS, Lange P, Prescott E, Diderichsen F. (2012) Perceived stress and risk of adult-onset asthma and other atopic disorders: a longitudinal cohort study. Allergy, 67, 1408-1414
59) Jackson DJ, Gangnon RE, Evans MD, Roberg KA, Anderson EL, Pappas TE, Printz MC, Lee WM, Shult PA, Reisdorf E, Carlson-Dakes KT, Salazar LP, DaSilva DF, Tisler CJ, Gern JE, Lemanske RF Jr. (2008) Wheezing rhinovirus illnesses in early life predict asthma development in high-risk children. Am J Respir Crit Care Med, 178, 667-672
27
Ⅵ.
Figure
Figure 1. Schematic representation of the protocols for tolerization, stress exposure, sensitization, and antigen challenge.
28
Figure 2. Effects of MS on plasma corticosterone concentrations. Blood was collected from MS and non-MS mice before the 1st and after the 1st, 3rd, and 6th MS exposures, and plasma corticosterone levels were measured in enzyme immunoassays. Data are expressed as the mean ± SD (n = 3–11). #P < 0.05 compared to values before stress exposure. *P < 0.05 and **P < 0.01, compared to non-stressed mice on the same day.
29
Figure 3. Effects of MS on methacholine-induced airway hyperresponsiveness.
Airway reactivity (RL) (A) and airway sensitivity (PC200) (B) were measured 5 days after the last OVA challenge (PND 81). Data are expressed as the mean ± SD (n = 3–7 from 3 independent experiments). *P < 0.05 and **P < 0.01 compared with non-MS/non-tolerized mice. #P < 0.05 and ##P < 0.01, compared with non-MS/tolerized mice.
30
Figure. 4. Effects of MS on OVA-specific IgE and airway mucus secretion.
(A) Sera were obtained from mice of each treatment group 5 days after the last OVA challenge (PND 81). OVA-specific IgE in serum was measured by ELISA.
Data are expressed as the mean ± SD (n = 6–8 from 3 independent experiment s). *P < 0.05, and ***P < 0.001. (B) Lungs were isolated from mice of each treatment group 5 days after the last OVA challenge (PND 81). Early life stress Lung sections were prepared and stained with period acid–Schiff (PAS). Shown are representative images of lung sections (left, scale bar, 100 μm) and the percentage of PAS-positive cells (right) from the 4 mice groups. Data are expressed as the mean ± SD (n = 6–8 mice from 3 independent experiments).
*P < 0.05, and ***P < 0.001.
31
Figure. 5. Effects of MS on airway inflammation. BALFs were obtained from mice of each group 5 days after the last OVA challenge (PND 81). Total cells (Total), macrophages (MΦ), eosinophils (Eo), neutrophils (Neu), and lymphocytes (Lym) were counted. Data are expressed as the mean ± SD (n = 6–8 from 3 independent experiments). *P < 0.05, **P < 0.01, and ***P < 0.001.
32
Figure. 6. Effects of MS on BALF cytokine contents. BALFs were obtained from mice of each group 1 day after the last OVA challenge (PND 77). IL-4, IL-5, IL-13 and IFN-γ levels were measured by ELISA. Data are expressed as the mean ± SD (n = 7–11 from 4 independent experiments). *P < 0.05 and **P < 0.01.
33
Figure. 7. Effects of MS on sensitization. On PND 76, splenocytes were prepared from mice of each group of sensitized mice and stimulated in vitro with OVA. After a 72-h culture, cell proliferation was measured in WST-8 assays (A). The content of IL-4 (B) and IFN-γ (C) in culture supernatants were determined by ELISA. Data are expressed as the mean ± SD of triplicate cultures from 3 independent experiments. *P < 0.05, **P < 0.01, and ***P <
0.001.
34
Figure. 8. Effects of MS on Treg cells in BLNs. BLNs were collected from mice of each group after the final MS (PND 22). The percentages of CD3+CD4+ Foxp3+ cells in CD3+CD4+ cells were determined. Data from 4 independent experiments are expressed as the mean ± SD. *P < 0.05.