光屈性におけるオーキシン輸送制御
酒井 達也・芳賀 健
新潟大学大学院自然科学研究科
Control of auxin transport in the phototropic response
Tatsuya Sakai and Ken HagaGraduate School of Science and Technology, Niigata University
要旨 : Phototropism allows plants to change their growth direction in response to the location of the light source. Asymmetric distribution of the phytohormone auxin occurs in response to a phototropic stimulus, and causes differential growth on the two sides of the plant organ and con-sequent organ bending. Recently, molecular genetic analyses of Arabidopsis have begun to shed light on the molecular mechanisms underlying this response system, including phototropin blue light photoreceptors, phototropin signaling components, auxin transporters and others. This re-view highlights some of the recent progress on the control of auxin transport in the phototropic response.
植物化学調節学会■ The Japanese Society for Chemical Regulation of Plants
はじめに 光屈性とは,光照射の光源方向に応じて茎や根の成 長方向を変化させる植物の成長運動である.茎や胚軸を屈 曲させて地上部を光源方向に向けるのは光合成に必要な光 を十分に受けるため,根の光を避ける性質は発芽後に大地 へと根を伸長させるため重力屈性とともに働く性質だと考 えられている.19 世紀に Darwin が行った単子葉植物芽生 え幼葉鞘の光屈性の観察は,光受容部位が幼葉鞘先端に存 在し,屈曲が生じる作用部位と離れていること,この間に 何らかのシグナルのやりとりがあることを明らかにした (Darwin 1880).20 世紀に入ってこのシグナル伝達物質が 植物成長ホルモンオーキシンとして同定された後,「光屈 性,重力屈性における偏差成長屈曲は,オーキシン横移動 によるオーキシン濃度勾配形成によって生じる」というコ ロドニー・ウエント説が確立した(Holland et al. 2009 ; Whippo and Hangarter 2006).
光屈性反応においてオーキシン濃度勾配が形成されるこ とは,複数の実験で証明されている.Briggs らはトウモロ コシ幼葉鞘先端に横から光を照射すると陰側にオーキシン 活性物質が蓄積すること,幼葉鞘先端に薄い障害物を差し 込んで物質の横移動を阻害するとこの蓄積がおきないこと を示した(図 1A : Briggs et al. 1957).飯野は内在性オー キシン・インドール-3-酢酸(IAA)量を実際に定量し, 光屈性反応時のトウモロコシ幼葉鞘陰側に IAA がより多 く蓄積していることを明らかにした(図 1B : Iino 1991). 光屈性誘導に必須のシグナル伝達因子 NPH3(後述)が欠 損したイネ突然変異体 cpt1 ではこのオーキシン濃度勾配 が形成されないことを,当時飯野研究グループに所属して いた芳賀(本稿著者)が明らかにした(図 1C : Haga et al. 2005).Liscum ら研究グループは,シロイヌナズナと同じ アブラナ科植物ヤセイカンラン(Brassica oleracea)の芽 生え胚軸の光屈性反応を観察し,陰側に IAA が多く蓄積 していることを明らかにした(Esmon et al. 2006).このオー キシン濃度勾配は各種オーキシン誘導遺伝子に加え,細胞 壁伸展性を高めるエキスパンシン酵素をコードする遺伝子 の発現を誘導することが示され,オーキシン濃度勾配と偏 差成長誘導の因果関係を強く示唆した(図 1D).また簡便 な方法として,オーキシンレポーター遺伝子を利用した オ ー キ シ ン 分 布 パ タ ー ン の 解 析 も 行 わ れ て い る. DR5rev:GFPオーキシンレポーター遺伝子を導入したシロ イヌナズナ芽生えに横から光を照射すると,オーキシン濃 度勾配を示すように GFP 蛍光強度が陰側により強く観察 される(図 1E).これらの結果は,どれも光屈性反応時に はオーキシンが幼葉鞘や胚軸の陰側に多く蓄積することを 示している. それでは,光照射後,オーキシンはどのようにして陰側 へと蓄積するのだろうか ? 図 2 で示すように,5 つのオー キシン濃度勾配形成機構が作業仮説として考えられる (Sakai and Haga 2012).図 2 の ④ および ⑤ の機構は可能 性としては考えられるものの,これまで解析が行われてお らず本稿では触れない.コロドニー・ウエント説の骨子は ① に当たる.本稿は ①,②,③ の光によるオーキシン輸 送制御の分子機構について最近の知見を紹介する. 1 光屈性を誘導するフォトトロピン光受容体複合体 光屈性反応は一方向からの青色光照射によって誘導 される.光屈性が異常になったシロイヌナズナ突然変異体 の分子遺伝学的解析が Briggs ら研究グループによって行 われ,光屈性を誘導する青色光受容体フォトトロピン
植物の生長調節 Vol. 47, No. 2, 2012 86
(phot)が発見された(Christie 2007 ; Holland et al. 2009 ; Sakai and Haga 2012).phot はシロイヌナズナにおいては phot1,phot2 の 2 分子種存在する.それぞれ約 120 kDa, 約 110 kDa の分子量を示すタンパク質が,N 末側に存在す る LOV と命名された 2 つのドメインに青色光吸収色素フ ラビンモノヌクレオチド(FMN)をそれぞれ一つずつ結 合して,青色光受容体として機能する.C 末側に AGCVIII ファミリーに属するセリン・スレオニン型タンパク質キ ナーゼドメインを有する.不活性状態では LOV ドメイン がキナーゼ活性を抑制しているが,FMN が青色光を吸収
13
図1
図 1 光屈性におけるオーキシン濃度勾配形成を示す実験.(A)Briggs らの実験(1957).(B)飯野の実験(1991). (C)芳賀らの実験(2005).(D)Esmon らの実験(2006).(E)オーキシンレポーター遺伝子発現解析.14
図2
図3
図4
図5 図6
図 2 胚軸光屈性におけるオーキシン濃度勾配形成機構の 5 つの作業仮説.暗条件下における双子葉芽生えにおいて は,茎頂や子葉といった若い組織でオーキシンが多く,基部に向かって濃度が薄くなるように勾配が形成される(赤 色).オーキシンは主に維管束を基部に向かって極性輸送されているが,表皮組織でも基部に向かった極性輸送が行 われていると思われる(赤色矢印).一方向からの青色光照射によって,胚軸光照射側のオーキシンが減少し,陰側 のオーキシンが増加する(赤色 : ①∼⑤).このオーキシン濃度勾配形成機構には,① オーキシン横輸送(紫色矢印), ② 維管束組織から放射軸方向へのオーキシン輸送制御(光照射側における輸送阻害[青]),③ 茎頂から基部へのオー キシン極性輸送制御(光照射側における輸送阻害[緑]),④ オーキシン生合成制御および ⑤ オーキシン代謝制御(光 照射側におけるオーキシン生合成阻害もしくは代謝促進によるオーキシン減少),が考えられる.The Japanese Society for Chemical Regulation of Plants (JSCRP)
すると LOV ドメイン内のシステイン残基と一過的な共有 結合を引き起こし,これがタンパク質の構造を変化させて LOVドメインによるキナーゼ活性抑制が解除される.phot のキナーゼ活性は自己リン酸化を誘導して phot 光受容体 のさらなる構造変化を引き起こし,光屈性を誘導するシグ ナルを流すと考えられている.筆者ら研究グループは, phot1と phot2 が光強度依存的に機能分担をしており,光 強度が 1 µmol m−2 sec−1以上と高いときには phot1 も phot2
も光屈性に働くが,それ以下の光強度では phot1 のみに よって光屈性が誘導されることを明らかにした(Sakai et al. 2001). phot1,phot2 光受容体には,それぞれに結合して働く複 合体サブユニットが存在する.タンパク質相互作用に働く BTBドメインを N 末側に,coiled-coilドメインを C 末側 に持つ NPH3 アダプタータンパク質は,phot1,phot2 どち らにも結合し,光屈性に必須の役割を担うシグナル伝達因 子である(Holland et al. 2009 ; Sakai and Haga 2012).NPH3 と同じ NRL(NPH3/RPT2)タンパク質ファミリーに属す る RPT2 は,phot1 および NPH3 への結合能を示して phot1 依存的な光屈性反応の調節を行うが,phot2 への結合は観 察されず,遺伝学的にも phot2 に依存した光屈性には必要 ないことを筆者らは明らかにしている(Inada et al. 2005). さらにシロイヌナズナで 4 分子種存在する PKS ファミ リータンパク質群,PKS1,PKS2,PKS3,PKS4 も光屈性 に 働 く. こ の う ち 少 な く と も PKS1,PKS2 が phot1, phot2,NPH3 と in vivo で結合していることが明らかになっ ている(de Carbonnel et al. 2010 ; Lariguet et al. 2006).こ のようなタンパク質複合体が原形質膜内側でどのような生 化学的機能を持ち,光屈性を誘導しているのかは未だほと んど明らかになっておらず,光屈性研究の最重要課題の一 つになっている. 2 光屈性に関与するオーキシン輸送体 PIN3 の発見 植物細胞において 4 種類のタンパク質ファミリーが オーキシン輸送活性を示す膜タンパク質として明らかに なっている.オーキシン排出活性を示す植物特異的な輸送 体 PIN-FORMED(PIN)ファミリー,原核生物から動植 物まで広く存在する ATP 結合カセット B 型サブファミ リー輸送体(ABCB)ファミリー,オーキシン流入活性を 示すアミノ酸パーミアーゼ様輸送体 AUX1/LAX ファミ リー,さらに最近明らかになったオーキシン排出体 PIN
-LIKES(PILS) フ ァ ミ リ ー で あ る(Barbez et al. 2012 ; Sakai and Haga 2012).シロイヌナズナゲノム中には,8 つ の PIN 遺伝子ファミリー,22 の ABCB 遺伝子ファミリー, 4 つの AUX1/LAX 遺伝子ファミリー,7 つの PILS 遺伝子 ファミリーがそれぞれ存在する.
光屈性に関与することが初めて報告されたオーキシン輸 送体は PIN3 である.Friml ら研究グループは PIN3 遺伝子
が異常になった突然変異体を単離し表現型を観察した.そ の結果,pin3 突然変異体は胚軸の光屈性,重力屈性,さら に根の重力屈性に異常を示すことが明らかになった(Friml et al. 2002).根冠コルメラ細胞の原形質膜に一様に存在し ていた PIN3 は,重力刺激を根に与えると,オーキシン濃 度勾配を形成させるように重力方向へと細胞内局在を変化
14
図2
図3
図4
図5 図6
図 3 青色光による PIN3 細胞内局在制御.暗条件下,胚 軸維管束を介して基部へのオーキシン輸送が行われている (赤色矢印).維管束組織に豊富に存在するオーキシンは(赤 色),内皮組織,皮層組織へと放射軸方向にオーキシンが 拡散する.暗条件では,内皮細胞で発現している PIN3 は 原形質膜全体に局在してオーキシン排出に働く(青色). 胚軸の横から青色光が照射されると,光照射側の内皮細胞 における PIN3 の細胞内局在が変化し,維管束側に不等分 布する.この細胞内局在変化によって,維管束から光照射 側へと拡散するオーキシンが,PIN3 の働きによって維管 束組織に排出されて戻され,結果として光照射側へのオー キシン流出が止まることが予想される.14
図2
図3
図4
図5 図6
図 4 胚軸光屈性における PIN3 および ABCB19 の働き. PIN3 は茎頂から若干離れた内皮細胞で,維管束から放射 状に拡散するオーキシンを光照射側で抑制しオーキシン量 を減少させる.ABCB19 は茎頂に近い領域で,基部へのオー キシン極性輸送を抑制し,間接的に陰側のオーキシン増加 を誘導すると思われる.コロドニー・ウエント説で提案さ れたオーキシン横輸送はこのオーキシン増加誘導に働くと 思われるが,そこで働くオーキシン輸送体はまだ発見され ていない(unknown).植物の生長調節 Vol. 47, No. 2, 2012 88 させた.この結果から,PIN3 は胚軸の屈性反応において も刺激感知後に細胞内局在を変化させてオーキシン横輸送 を行う分子であることが推測された. Frimlら研究グループは,PIN3 の光屈性における機能に ついて重要な研究成果を最近発表した(Ding et al. 2011). 胚軸内皮細胞の原形質膜に一様に分布していた PIN3 が, 横からの光照射によって,phot1 依存的に光照射側の内皮 細胞でのみ,内皮細胞維管束側への特異的な不等分布を示 した(図 3).DR5rev:GFP オーキシンレポーター遺伝子を 用いてオーキシン分布パターンを調べてみると,オーキシ ンは野生型では陰側により多く蓄積していることが示唆さ れたが,phot1 および pin3 突然変異体ではこのような不等 分布が抑制されていた.以上の結果は,phot1 の活性化が 光照射側内皮細胞における PIN3 の細胞内局在を変化させ, 維管束系から放射軸方向へと流れるオーキシンの排出を光 照射側で抑制することによって,陰側へのオーキシン蓄積 を誘導していることを示唆した(図 3,図 4).
ただし,pin3 および pin3 pin7 突然変異体の胚軸光屈性は, 野生型に比べて,それぞれおよそ 13%,17% の屈曲度減 少というわずかな表現型しか示さなかった(Ding et al. 2011).PIN3 および PIN7 の光屈性への関与を示すと同時 に,他の分子機構のさらなる機能重複性によってオーキシ ン濃度勾配形成および光屈性が誘導されていることを示唆 する結果だった. Christieら研究グループは,双子葉植物黄化芽生えがフッ クを維持したままでいると光屈性誘導時における光照射方 向が個体毎に異なり屈曲度の差が大きいことを問題視し た.彼らは光を照射してフックを解消し子葉が開いたシロ イヌナズナ脱黄化芽生えを作成し,これを暗条件に戻して 暗順化したのち光屈性反応を観察する方法を考案した (Christie et al. 2011).この方法で光屈性を観察すると, pin3 変異体は茎頂に近い領域でのみ野生型よりも強い光 屈性を示し,屈曲度としては野生型よりも促進するという 結果を得た.一方,pin7 は野生型よりも胚軸屈曲が若干抑 制される表現型を示した.これらの結果は,芽生えの状態 に応じて PIN3 と PIN7 の光屈性における貢献度が変化す ることを示唆する一方,PIN ファミリーの光屈性における 重要性についてはやはり疑問が残る結果だった. 3 光屈性を制御するオーキシン輸送体 ABCB19 の発 見 オーキシン輸送阻害剤 1-ナフチルフタラミン酸(NPA) への結合タンパク質として同定された ABCB19 は,Mur-phy ら研究グループによる逆遺伝学的解析により光屈性へ の関与が明らかにされた(Noh et al. 2003).abcb19 突然変 異体は胚軸における基部へのオーキシン極性輸送に異常を 示し,光屈性および重力屈性の促進を示した.ABCB19 に 依存した基部へのオーキシン極性輸送が失われると,オー キシン横輸送が間接的に促進され,結果として偏差成長が 促進するのではないかと予想された. Christieらは,abcb19 突然変異体の光屈性反応時のオー キシンレポーター遺伝子発現を観察し,維管束組織に存在 するオーキシンが減少し,陰側のオーキシン不等分布が促 進されることを示唆する結果を得た(Christie et al. 2011). さらに,活性化した phot1 が ABCB19 を直接リン酸化し, そのオーキシン輸送活性を阻害することを明らかにした. 以 上 の 結 果 は,phot1 に よ る ABCB19 の リ ン 酸 化 が ABCB19の維管束組織におけるオーキシン極性輸送を抑制 し,胚軸上部のオーキシン蓄積を促進し,胚軸上部で働く オーキシン横輸送機構によって陰側にオーキシンが運ばれ て,オーキシン濃度勾配形成に働く,という光屈性分子機 構を推測させた(図 4).ただし,abcb19 機能欠失型突然 変異体が光屈性促進の表現型を示すことから,ABCB19 の 機能制御が光屈性に重要なのかどうかはまだよく分からな い.phot1 に よ る リ ン 酸 化 が お き な い ア ミ ノ 酸 置 換 ABCB19タンパク質を abcb19 機能欠失型突然変異体中で 発現させ,維管束組織での基部へのオーキシン極性輸送の 阻害がおきない芽生えを用いて,光屈性反応がどうなるか を今後検討する必要がある. 4 光屈性におけるオーキシン輸送体機能の再検討 これまで筆者らは光屈性制御機構を明らかにするた めに,phot 光受容体複合体の生化学的機能解析を中心に, 光屈性シグナル伝達経路上流の研究を行ってきた(酒井達 也 2010 ; Sakai and Haga 2012).光屈性研究のブレークス ルーを目指して,オーキシン輸送体の光屈性における機能 と制御という下流研究を 5 年ほど前に計画し,2007 年か ら 2010 年度までの科学技術振興会さきがけプログラムに 採択された.当時オーキシン輸送活性が報告されていた オーキシン輸送体をコードする遺伝子,PIN1∼PIN8 遺伝 子,AUX1 お よ び LAX1∼LAX3 遺 伝 子,ABCB1,ABCB4,
ABCB19遺伝子,が異常になった突然変異体を網羅的に収 集し,連続光照射によって誘導される二次正光屈性を観察 した.また各 PIN 遺伝子 cDNA をエストラジオール誘導 系プロモーター下流につなぎ,エクトピックに各 PIN 遺 伝子を発現させた時の形質転換体の表現型を観察した.こ れらの解析で弱いながらも光屈性に関与するオーキシン輸 送体が発見できれば,多重変異体を作成することによって オーキシン輸送体の機能重複が示され,phot シグナル伝達 経路の標的が明確になるのではないかと期待した.しかし, これらの解析では,すでに報告されていた PIN3,ABCB19 以外には,オーキシン輸送体の光屈性における関与を明ら かにすることはできなかった(酒井 未発表). 光屈性にオーキシン輸送活性が必要であることを示す一 つの根拠は,オーキシン輸送阻害剤処理を行うとシロイヌ ナズナ芽生え胚軸の光屈性が強く阻害されることである. The Japanese Society for Chemical Regulation of Plants (JSCRP)
植物の生長調節 Vol. 47, No. 2, 2012 89 筆者らは,オーキシン極性輸送阻害剤 NPA が abcb19 突然
変異体の光屈性を阻害できないことを発見した(Nagashima et al. 2008).この結果は NPA が ABCB19 以外のオーキシ ン輸送体機能を直接阻害して光屈性抑制に働いているわけ ではないことを強く示唆した.しかし,abcb19 欠失突然 変異体は光屈性を示すことから,NPA が ABCB19 のオー キシン輸送活性を阻害するだけで光屈性が抑制されるとは 考えにくい.Murphy ら研究グループとの共同研究の結果, ABCB19は PIN ファミリーと結合して,PIN の輸送活性を 制御していることを明らかにした(Blakeslee et al. 2007). この結果を考慮すると,ABCB19 タンパク質は NPA と結 合することによって,自分自身のオーキシン輸送活性ばか りでなく,PIN オーキシン輸送体機能も抑制し,光屈性に 必要な陰側でのオーキシン極性輸送も失わせて光屈性を阻 害することが示唆された. 作用機序の明らかな新しいオーキシン輸送阻害剤で処理 すると,光屈性はどうなるだろうか ? 岡山理科大学林謙 一郎博士らグループは当時,オーキシン受容体には結合せ ずにオーキシン輸送体の穴にのみ入り込み,オーキシン輸 送を阻害する試薬の開発を行っていた.彼らの開発した人 工オーキシン・ナフタレン酢酸にベンジル基を修飾した Bz-NAA は,酵母で発現した PIN,ABCB19,AUX1,す べてのオーキシン輸送体のオーキシン輸送活性を減少させ た.一方,Bz-NAAはオーキシン派生体であっても TIR1 オーキシン受容体には結合せず,TIR1 シグナル伝達経路 には影響を与えなかった(Tsuda et al. 2011).筆者らは林 博士から Bz-NAAを分譲してもらい,光屈性反応の阻害 効果を調べることにした.驚いたことに,連続光照射によ る二次正光屈性には Bz-NAAは全く影響を与えなかった
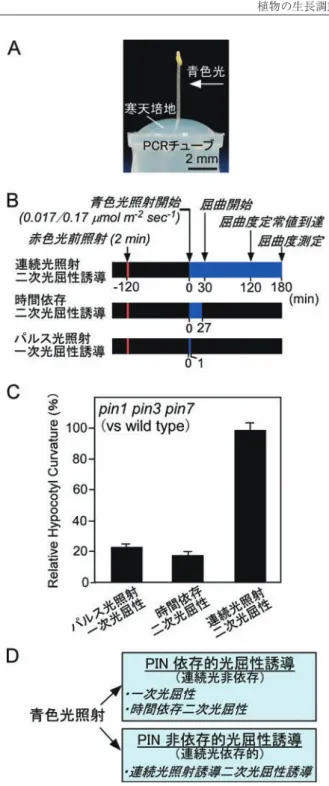
(Tsuda et al. 2011).Bz-NAAは シ ロ イ ヌ ナ ズ ナ 野 生 型, abcb19突然変異体,どちらの胚軸重力屈性も阻害効果を 示すことから,胚軸重力屈性に関与するオーキシン輸送体 の活性は阻害していることが示唆された.これらの結果は, オーキシン輸送体機能の異常や機能抑制が重力屈性ほどに は光屈性に影響を与えないことを示唆した.さきがけプロ グラム計画当初に予想していた光屈性誘導機構とは全く異 なる方向へ研究が展開した. 5 PIN に依存した光屈性誘導機構の解明 ここで改めて,光屈性の植物生理学的知見を紹介し ておきたい.光屈性応答の生理学的解析では,光屈性応答 は短いパルス光照射で誘導される光屈性と,ある程度の時 間(10 分前後)以上に連続させた光照射による光屈性反 応には違いが観察され,それぞれを一次正光屈性,二次正 屈性とわけている(図 5 : Iino 2001 ; Poff et al. 1994).一 次正屈性反応は,横軸を光量(光強度×時間),縦軸を胚 軸屈曲角度で示した場合,釣り鐘型の反応曲線を示す.こ れはある一定以上の光量になると,光寛容とよばれる現象 が生じて,芽生えが光に応答しなくなるためと考えられて いる.しかしながら光寛容の現象は光照射の時間が延びる と解消され,光量よりも照射時間に依存して屈曲度が増す 二次正光屈性反応へと転ずる.幼葉鞘を用いた植物生理学 的解析においては,低い光強度の光を短時間(パルス光) 照射して光屈性を誘導する一次正光屈性観察がよく行われ た. し か し 一 次 正 光 屈 性 も 二 次 正 光 屈 性 も,phot1 や NPH3といったシグナル伝達因子は同じように働いている ことがその後の分子遺伝学的解析によって明らかになり, シロイヌナズナ突然変異体を用いた光屈性解析では連続光 照射を用いた二次正光屈性を観察するのが一般的になっ た. 芳賀は,機能重複を示すことが示唆されるオーキシン輸 送体の突然変異体解析には厳密な一次正光屈性反応観察が ふさわしいと考え,pin 突然変異体の一次正光屈性を観察 した.胚軸屈曲が寒天培地との接触によって阻害されない ように PCR チューブを用いた芽生え培養を行い(図 6A : チューブ法),光屈性を誘導する青色光照射より 2 時間前 に赤色光を黄化芽生えにパルス照射(赤色光前照射)して 一次正光屈性の屈曲度を促進し,光屈性を誘導する青色光 照射は 1 分間,phot1 のみ働くことが予想される低い光強 度で行い,その後 3 時間暗条件に戻したのち,そこで生じ る胚軸屈曲度を測定した(図 6B). このような実験条件で各 pin 突然変異体の胚軸光屈性を 観察してみると大変興味深い結果が得られた(Haga and Sakai 2012).明らかに pin1 および pin7 の突然変異体が
pin3同様に一次正光屈性に異常を示した.一方,他の pin 突然変異体は有意な差を示さなかった.そこで pin1 pin3 pin7三重変異体を作成し,その一次正光屈性を観察すると, 期待通り著しい光屈性異常を示した(図 6C).この結果は, パルス光照射の一次正光屈性においては,PIN1,PIN3, PIN7オーキシン輸送体が機能を重複してオーキシン濃度 勾配を形成し,光屈性を誘導することを強く示唆する結果 だった.
14
図2
図3
図4
図5 図6
図 5 光量と胚軸光屈性の関係.The Japanese Society for Chemical Regulation of Plants (JSCRP)
植物の生長調節 Vol. 47, No. 2, 2012 90 6 PIN に依存しない光屈性誘導機構の発見 次にチューブ法で連続光照射によって二次正光屈性 を観察すると,野生型胚軸が光照射開始およそ 30 分後か ら屈曲を始め,およそ 120 分後には光源方向に完全に曲
が っ て 定 常 値 に 達 し た( 図 6B : Haga and Sakai 2012).
pin3,pin7 シングル突然変異体は 3 時間の連続光照射の二 次正光屈性には全く異常を示さず,屈曲の時間的変化も野 生型とほぼ同様であった.pin1 突然変異体は光屈性応答の 遅延を示したが,これは光屈性異常よりも胚軸伸長の遅延 に起因することが示唆された.驚いたことに,pin1 pin3 pin7三重変異体で観察しても,連続光照射の二次正光屈性 には全く異常が観察されなかった(図 6C).この結果は, PIN に依存した一次正光屈性誘導機構とは全く異なる分子 機構が連続光照射によって誘導される二次正光屈性には働 いていることを強く示唆した. それでは,屈曲が開始する前の 27 分の光照射後に暗条 件に戻して,屈曲度を測定するとどのようになるだろう か ? この光照射条件では,野生型胚軸では一次正光屈性, 光寛容を越えた二次正光屈性反応が現れる(時間依存二次 正光屈性とよぶ).このような光条件下では,pin1 pin3 pin7多重変異体は著しい光屈性異常を示した(図 6C :
Haga and Sakai 2012).この結果は,時間依存二次正光屈性 においては,パルス光照射誘導の一次正光屈性同様,PIN に依存した光屈性誘導機構が働くことを示した. 我々の研究結果は,光屈性反応が少なくとも 2 つの分子 機構によって生じていることを明らかにした(図 6D).第 一の分子機構はパルス光照射によって活性化しうる PIN1, PIN3,PIN7 依存的な光屈性誘導機構である.一次正光屈 性および時間依存二次正光屈性はこちらの分子機構によっ て主に誘導される.第一の分子機構は,光照射方向を記憶 することが可能で,胚軸屈曲が実際に生じる時間には青色 光が照射されていなくても光屈性を誘導することが可能で ある.パルス光によって PIN1,PIN3,PIN7 の細胞内局在 がひとたび変化すると,しばらくの間は光に非依存的にそ の細胞内局在変化を維持し,オーキシン濃度勾配を暗条件 下で形成し,胚軸の偏差成長を誘導することが推測される. 第二の分子機構は連続光照射を必要とする PIN1,PIN3, PIN7非依存的な光屈性誘導機構である.これは胚軸屈曲 が生じる間,常に光屈性誘導刺激となる青色光照射を必要 とすることが示唆される. 7 phot シグナル伝達経路による光屈性誘導機構 PINに依存した光屈性誘導機構は phot によってど のように制御されているのだろうか ? PIN1 および PIN7 が青色光照射によって phot 依存的に細胞内局在が変化す るかの検証はこれからの課題である.抗体を用いた観察に より,PIN1 が phot1 によって細胞内局在を変化させるこ とが報告されているが,抗体の特異性が疑問視されており, 今後さらに検証が必要である(Blakeslee et al. 2004 ; Sakai and Haga 2012).Ding らは,青色光照射が phot 依存的に PIN3 リン酸化に働くタンパク質キナーゼをコードした
PI-NOID(PID)遺伝子の転写を抑制し,結果として PIN3 の
図 6 チューブ法を用いた pin 突然変異体の光屈性応答観 察(Haga and Sakai 2012).(A)PCR チューブを用いたシ ロイヌナズナ黄化芽生え胚軸の光屈性応答の観察.(B) 光照射条件.(C)野生型胚軸の光屈曲度と比較した pin1 pin3 pin7三重変異体の胚軸屈曲度の割合.野生型胚軸は パルス光照射一次正光屈性,時間依存二次正光屈性,連続 光照射二次正光屈性,それぞれ約 46 度,68 度,93 度の屈 曲度を示した.(D)光屈性誘導に働く 2 つの分子機構.
The Japanese Society for Chemical Regulation of Plants (JSCRP)
リン酸化が抑制され,光照射側の内皮細胞における PIN3 の細胞内局在の変化を誘導するというモデルを提案してい る(Ding et al. 2011).我々はこのモデルを検証するため, 一次正光屈性誘導時の PID 発現量を定量的 RT-PCR法に よって測定した.予想に反し,青色光照射では PID 発現 量は減少せず,我々が観察している一次正光屈性では PID 発現制御は関係ないことを明らかにした(Haga and Sakai 2012).むしろ光屈性誘導前の赤色光前照射によっておよ そ 1/3 ほどに PID 転写産物量が減少したことから,青色光 照射による PID 転写産物減少は,phot 依存的でなく,青 色光照射によっても活性化する赤色光受容体フィトクロー ムの働きによって誘導された反応なのではないかと我々は 予想している. 青色光照射による phot を介した PIN3 細胞内局在変化の 分子機構について考えられる仮説をあげておく.第一に, PID ファミリータンパク質(PID,PID2,WAG1,WAG2 の 4 分子)の機能発現制御の可能性がある.phot による PID タンパク質安定化制御や,PID キナーゼ活性の調節な どの可能性である.PID はカルシウムイオンによって活性 が抑制されること,phot が細胞内カルシウムイオン濃度の 一過的上昇を誘導することが明らかになっていることか ら,phot による細胞内カルシウムイオン濃度制御による PID活性調節の可能性は十分考えられる(Sakai and Haga 2012).第二に,phot による PIN 脱リン酸化酵素の機能発 現制御の可能性がある.PIN1 の細胞内局在は,PP6 タン パク質脱リン酸化酵素の働きによる脱リン酸化によって, PIDによるリン酸化制御とは対立的に調節されていること が明らかになっている(Dai et al. 2012).PIN3 も同様に PP6による脱リン酸化制御が存在し,phot は PP6 のタンパ ク質発現や活性調節を行っている可能性がある.第三に, photによる PIN タンパク質の直接的なリン酸化の可能性 がある.In vitro の実験では,phot1 組換えタンパク質は PIN3のリン酸化を行わないことは示されているが(Ding et al. 2011),in vivo におけるリン酸化の可能性は残ってい る. 一方,連続光照射を必要とする光屈性誘導機構は今のと ころ作業仮説を立てる段階に至っていない.連続光照射を 行うと pin3 変異体においても野生型同様に DR5:GFP 発現 が光照射側と陰側の間で差を観察できること(Haga and Sakai 2012),NPA が連続光照射誘導の光屈性を完全に抑 制することから(Nagashima et al. 2008),PIN1,PIN3,PIN7 非依存的光屈性誘導機構によってもオーキシン濃度勾配が 形成されているのではないかと予想している.これが他の オーキシン輸送体機能によるのか,オーキシン生合成・代 謝制御によるのかは今後の解析を行う必要がある.また オーキシンとは異なる細胞伸長促進因子,または伸長抑制 因子の働きも否定できない. おわりに コロドニー・ウエント説で提案された図 2 ① の光 屈性誘導機構は,PIN3 が働く図 2 ②,ABCB19 が働く図 2③ のオーキシン輸送制御を加えるように修正された(図 3).図 2 ① で実際に働くオーキシン輸送体は未同定だが, 双子葉植物芽生え茎頂近傍,単子葉植物幼葉鞘先端,とも にオーキシン横輸送の存在は強く示唆されている(Christie et al. 2011 ; Matsuda et al. 2011).我々は少なくとも連続光 を必要としない光屈性においては,PIN1,PIN3,PIN7 が 極めて重要な役割を演じていることを明らかにした. PIN1,PIN7 はどのような調節によって光屈性に働くのか, 茎頂に近い位置でのオーキシン横輸送に働くオーキシン輸 送体の正体は何か,そしてこれらのオーキシン輸送体の機 能を phot 青色光受容体複合体はどのように制御している のかについて,今後明らかにしていきたい. 本稿を締めくくるにあたり,光屈性誘導には光照射側で の細胞伸長抑制因子の蓄積が働くという伸長抑制因子仮説 について意見を述べておきたい.古くは Blaauw が提唱し (Whippo and Hangarter 2006),国内では筑波大学長谷川宏 司博士ら研究グループがブルインスマ・長谷川説としてこ の仮説を強く支持し,高校の教科書およびインターネット 上でまことしやかに「光屈性におけるオーキシンの働きを 疑問視する最近の学説」として紹介されている(東郷重法 2011 ; 長谷川・Tin 2011).伸長抑制因子仮説において決 定的に欠けているのは,1)光屈性を誘導する phot の活性 化と伸長抑制因子の蓄積の因果関係(phot および nph3 突 然変異体では伸長抑制因子は蓄積しないか ?),2)伸長抑 制因子蓄積と光屈性の因果関係(伸長抑制因子を蓄積でき ない突然変異体は光屈性反応に異常が生じるか ?),の二 点の分子遺伝学的証拠である.本稿で述べたようにオーキ シン濃度勾配による光屈性誘導の分子機構は,この二点が 証明されている.オーキシンの働きに重複して働く他の成 長制御因子は今後発見される可能性があるが,光屈性の主 要な制御因子として働くオーキシンを否定することは今後 も極めて難しいと結論する. 謝辞 共同研究者の皆様,並びにさきがけ研究領域「代謝 と制御」および新学術領域「植物環境応答」の研究総括を はじめとする関係者の方々のご支援に厚く御礼申し上げま す. 文 献 酒井達也(2010) 光屈性におけるオーキシンシグナリング.「植物 のシグナル伝達」,柿本辰男・高山誠司・福田裕穂・松岡信編著, pp. 77-83,共立出版,東京. 東郷重法(2011) 高校生物教科書の現状と問題点.「植物生理化学」, 長谷川宏司・広瀬克利編著,pp. 1-22,大学教育出版,岡山. 長谷川剛・Tin, WWT(2011) 光屈性.「植物生理化学」,長谷川宏司・
植物の生長調節 Vol. 47, No. 2, 2012 92
広瀬克利編著,pp. 51-84,大学教育出版,岡山.
Barbez, E, Kubeš, M, Rolčík, J, Béziat, C, Pěnčík, A, Wang, B, Ros-quete, MR, Zhu, J, Dobrev, PI, Lee, Y, Zažímalovà, E, Petrášek, J, Geisler, M, Friml, J and Kleine-Vehn, J (2012) A novel putative
auxin carrier family regulates intracellular auxin homeostasis in plants.
Nature 485 : 119-122.
Blakeslee, JJ, Bandyopadhyay, A, Lee, OR, Mravec, J, Titapi-watanakun, B, Sauer, M, Makam, SN, Cheng, Y, Bouchard, R, Ada-mec, J, Geisler, M, Nagashima, A, Sakai, T, Martinoia, E, Frim, J, Peer, WA and Murhy, AS (2007) Interactions among PIN
-FORMED and P-glycoprotein auxin transporters in
Arabidop-sis. Plant Cell 19 : 131-147.
Blakeslee, JJ, Bandyopadhyay, A, Peer, WA, Makam, SN and Murphy, AS (2004) Relocalization of the PIN1 auxin efflux facilitator plays a
role in phototropic responses. Plant Physiol 134 : 28-31.
Briggs, WR, Tocher, RD and Wilson, JF (1957) Phototropic auxin
re-distribution in corn coleoptiles. Science 126 : 210-212.
Christie, JM (2007) Phototropin blue-light receptors. Annu Rev Plant
Biol 58 : 21-45.
Christie, JM, Yang, H, Richter, GL, Sullivan, S, Thomson, CE, Lin, J, Titapiwatanakun, B, Ennis, M, Kaiserli, E, Lee, OR, Adamec, J, Peer, WA and Murphy, AS (2011) phot1 inhibition of ABCB19
primes lateral auxin fluxes in the shoot apex required for phototropism.
PLoS Biol 9 : e1001076.
Dai, M, Zhang, C, Kania, U, Chen, F, Xue, Q, Mccray, T, Li, G, Qin, G, Wakeley, M, Terzaghi, W, Wan, J, Zhao, Y, Xu, J, Friml, J, Deng, XW and Wang, H (2012) A PP6-type phosphatase holoenzyme
di-rectly regulates PIN phosphorylation and auxin efflux in Arabidopsis.
Plant Cell 24 : 2497-2514.
Darwin, C and Darwin, F (1880) The Power of Movement in Plants,
John Murray, London.
de Carbonnel, M, Davis, P, Roelfsema, MR, Inoue, S, Schepens, I, Lar-iguet, P, Geisler, M, Shimazaki, K, Hangarter, R and Fankhauser, C (2010) The Arabidopsis PHYTOCHROME KINASE
SUB-STRATE2 protein is a phototropin signaling element that regulates leaf flattening and leaf positioning. Plant Physiol 152 : 1391-1405.
Ding, Z, Galván-Ampudia, CS, Demarsy, E, Łangowski, L, Kleine
-Vehn, J, Fan, Y, Morita, MT, Tasaka, M, Fankhauser, C, Offringer, R and Friml, J (2011) Light-mediated polarization of the PIN3
aux-in transporter for the phototropic response aux-in Arabidopsis. Nature
Cell Biol 13 : 447-452.
Esmon, CA, Tinsley, AG, Ljung, K, Sandberg, G, Hearne, LB and Lis-cum, E (2006) A gradient of auxin and auxin-dependent
transcrip-tion precedes tropic growth responses. Proc Natl Acad Sci USA
108 : 236-241.
Friml, J, Wiśniewska, J, Benková, E, Mendgen, K and Palme, K (2002)
Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 415 : 806-809.
Haga, K and Sakai, T (2012) PIN auxin efflux carriers are necessary for
pulse-induced but not for continuous light-induced phototropism in
Arabidopsis. Plant Physiol 160 : 763-776.
Haga, K, Takano, M, Neumann, R and Iino, M (2005) The rice
COLE-OPTILE PHOTOTROPISM1 gene encoding an ortholog of Arabidopsis NPH3 is required for phototropism of coleoptiles and lateral
transloca-tion of auxin. Plant Cell 17 : 103-115.
Holland, JJ, Roberts, D and Liscum, E (2009) Understanding
phototropism : from Darwin to today. J Exp Bot 60 : 1969-1978.
Iino, M (1991) Mediation of tropisms by lateral translocation of
endoge-nous indole-3-acetic acid in maize coleoptiles. Plant Cell Environ
14 : 279-286.
Iino, M (2001) Phototropism in higher plants. In Photomovement, Ed.
Häder, D and Lebert, M, pp. 659-811, Elsevier, Amsterdam.
Lariguet, P, Schepens, I, Hodgson, D, Pedmale, UV, Trevisan, M, Kami, C, de Carbonnel, M, Alonso, JM, Ecker, JR, Liscum, E and Fankhauser, C (2006) PHYTOCHROME KINASE SUBSTRATE 1
is a phototropin 1 binding protein required for phototropism. Proc
Natl Acad Sci USA 103 : 10134-10139.
Matsuda S, Kajizuka T, Kadota A, Nishimura, T and Koshiba, T (2011)
NPH3- and PGP-like genes are exclusively expressed in the apical tip region essential for blue-light perception and lateral auxin transport in
maize coleoptiles. J Exp Bot 62 : 3459-3466.
Nagashima, A, Uehara, Y and Sakai, T (2008) The ABC subfamily B
auxin transporter AtABCB19 is involved in the inhibitory effects of N
-1-naphthylphthalamic acid on the phototropic and gravitropic responses
of Arabidopsis hypocotyls. Plant Cell Physiol 49 : 1250-1255.
Noh, B, Bandyopadhyay, A, Peer, WA, Spalding, EP and Murphy, AS
(2003) Enhanced gravi- and phototropism in plant mdr mutants
mis-localizing the auxin efflux protein PIN1. Nature 423 : 999-1002.
Poff, KL, Janoudi, AK, Rosen, ES and Orbović, V (1994) The
physiol-ogy of tropisms. In Arabidopsis, Ed. Meyerowitz, EM and Somer-ville, CR, pp. 639-664. Cold Spring Harbor Laboratory Press, New
York.
Sakai, T and Haga, K (2012) Molecular genetic analysis of phototropism
in Arabidopsis. Plant Cell Physiol 53 : 1517-1534.
Sakai, T, Kagawa, T, Kasahara, M, Swartz, TE, Christie, JM, Briggs, WR, Wada, M and Okada, K (2001) Arabidopsis nph1 and
npl1 : blue light receptors that mediate both phototropism and chloro-plast relocation. Proc Natl Acad Sci USA 98 : 6969-6974.
Tsuda, E, Yang, H, Nishimura, T, Uehara, Y, Sakai, T, Furutani, M, Ko-shiba, T, Hirose, M, Nozaki, H, Murphy, AS and Hayashi, K (2011)
Alkoxy-auxins are selective inhibitors of auxin transport mediated by
PIN, ABCB and AUX1 transporters. J Biol Chem 286 : 2354-2364.
Whippo, CW and Hangarter, RP (2006) Phototropism : Bending
to-wards enlightenment. Plant Cell 18 : 1110-1119.
連絡先 : 〒 950-2181 新潟県新潟市西区五十嵐 2 の町 8050
新潟大学大学院自然科学研究科 酒井 達也
TEL/FAX : 025-262-7880
E-mail : [email protected]-u.ac.jp
The Japanese Society for Chemical Regulation of Plants (JSCRP)