Bach2 遺伝子欠損マウスを用いた続発性肺胞蛋白症
発症機構の解明
著者
渋谷 里紗
雑誌名
東北医学雑誌
巻
129
号
2
ページ
185-189
発行年
2017-12
URL
http://hdl.handle.net/10097/00128767
―
女子大学院学生奨励賞受賞記念講演
― 2017年 5 月 20 日 : 勝山館Bach2 遺伝子欠損マウスを用いた
続発性肺胞蛋白症発症機構の解明
東北大学大学院医学系研究科 呼吸器内科学分野 渋 谷 里 紗 略 歴 2008年 3 月 東北大学医学部医学科 卒業 2008年 4 月 水戸医療センター 初期研修医 2010年 4 月 仙台市立病院 後期研修医 2016年 3 月 東北大学大学院医学系研究科 博士過程修了186
―
女子大学院学生奨励賞受賞記念講演
―Bach2 遺伝子欠損マウスを用いた
続発性肺胞蛋白症発症機構の解明
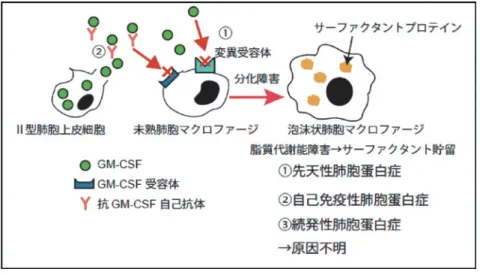
Identity Crisis of Alveolar Macrophages Leading to Pulmonary Alveolar Proteinosis 渋 谷 里 紗 東北大学大学院医学系研究科 呼吸器内科学分野 学術的背景 1) 肺胞蛋白症の病態生理 肺胞蛋白症は,肺胞内に肺胞サーファクタントが異 常に貯留する呼吸器疾患である.余剰な肺胞サーファ クタントは肺胞マクロファージによって貪食・代謝さ れることで全体量の調節が行われている1).肺胞蛋白 症の主な発症機序は,肺胞マクロファージの脂質処理 能力の異常である.肺胞マクロファージの脂質処理能 が正常に機能するためには,GM-CSFシグナルの下 流で転写因子 PU.1 遺伝子の発現が誘導され,さらに その下流で転写因子 Pparγ が作用することで分化が完 了し,脂質処理関連遺伝子の発現が誘導されることが 必要である.肺胞マクロファージの機能異常の背景の 1つには,このシグナルの破綻に起因する,肺胞マク ロファージの分化障害があることが明らかとなってい る2)(図 1).一方で,骨髄異形成症候群などの血液腫 瘍などに続発して起こる続発性肺胞蛋白症の原因は明 らかとなっていない3). 2) 転写因子 Bach2 の多様な機能
Bach (BTB and CNC homology) 因 子 は,MafK (musculoaponeurotic fibrosarcoma K) などの小 Maf と
2量 体 を 形 成 す る こ と で MARE (Maf recognition element : Maf認識配列)に結合し,遺伝子発現を抑 制する転写因子である(図 2).Bach2 は B 細胞にお ける形質細胞への分化や,クラススイッチ DNA 組み 換えによる抗体の多様性獲得の制御4,5),T 細胞におけ
るエフェクター T 細胞の分化に関わる遺伝子の発現 の抑制6),Common lymphoid progenitor や造血幹細胞
における,白血球分化のバランス制御を行っている7,8).
3) Bach2 遺伝子欠損マウスの肺胞蛋白症 筆者らは,Bach2 が肺胞マクロファージで発現して いること,Bach2 遺伝子欠損マウスは肺胞蛋白症を発 症していることを発見し,転写因子 Bach2 がマウス における肺胞蛋白症の発症に関与していることを発見 した.Bach2 遺伝子欠損肺胞マクロファージは貪食能 と脂質代謝能が低下していたが,GM-CSF経路は正 常であった.Bach2 遺伝子欠損マウスの肺胞マクロ ファージでは,脂質代謝,炎症応答および M2 マクロ ファージに関わる遺伝子群の発現に大きな変動を認め た.以上より,Bach2 遺伝子欠損マウスの肺胞蛋白症 の発症機序は,GM-CSF経路を介さない,肺胞マク ロファージの機能異常が原因であると考えられた.そ こで,野生型マウスの骨髄を Bach2 遺伝子欠損マウ スに移植したところ,肺胞蛋白症の発症を抑え,発症 後でも症状を改善させることに成功した9).以上の結 果から,Bach2 は肺胞マクロファージの機能的成熟と 維持に必要であり,GM-CSFとは別のシステムとし て,肺の恒常性を維持していると考えられた(図 3). 問 題 点 続発性肺胞蛋白症は背景となる疾患が多岐に渡り, その発症機序は不明であった.そのため,原疾患の治 療以外に有効な治療法は確立されていない. 研 究 目 的 Bach2遺伝子欠損マウスをモデルとし,肺胞マクロ ファージにおける Bach2 を中心とした遺伝子や細胞 のネットワークと,肺胞蛋白症の発症機序を明らかに することで,ヒトにおける続発性肺胞蛋白症の病態解 明と新たな治療戦略を考案する. 研 究 方 法 1) Bach2 遺伝子欠損肺胞マクロファージの遺伝子 発現パターンと,ミエロイド系細胞における Bach2 の標的遺伝子を網羅的に解析した. 2) 細胞特異的 Bach2 遺伝子欠損マウスの表現型 を体系的に解析した. 3) T 細胞を標的とした続発性肺胞蛋白症の新規治 療法について検討した. 研 究 結 果 1) 肺胞マクロファージにおける Bach2 の機能の 解明 Bach2遺伝子欠損マウスの肺胞マクロファージの遺 伝子発現パターンを,マイクロアレイを用いて網羅的 に解析したところ,肺胞マクロファージ特異的な遺伝 子発現パターンを失う一方で,脾臓赤脾髄マクロ ファージや腹腔マクロファージに特異的な遺伝子発現 図 2. 転写因子 Bach2 図 3. 背景模式図
188 渋谷 ─ Bach2 遺伝子欠損マウスを用いた続発性肺胞蛋白症発症機構の解明
パターンを示していた.このような遺伝子発現変化に 伴う機能の喪失が,肺胞蛋白症を発症させていると考 えられた.次に,肺胞マクロファージにおける Bach2 の標的遺伝子を明らかにするために,ミエロイド系細 胞における Bach2 の結合領域を Chromatin immuno-precipitation sequence (ChIP-seq) によって調べた.そ の結果,Bach2 は Il6 を含む多くの炎症関連遺伝子や, SpiCなどを含む脾臓赤脾髄マクロファージ特異的な 遺伝子に結合し,その発現を抑制していた.また,こ れまで Bach2 は転写抑制因子であると考えられてい たが,Bach2 は多くのヒストン遺伝子に結合し,それ ら遺伝子の発現を促進していることも明らかとなっ た10). 2) 肺胞蛋白症における細胞間相互作用の解明 Bach2遺伝子欠損マウスの肺胞洗浄液中に,リンパ 球の浸潤を認めた.T 細胞において Bach2 は Il4,Il6, Ifngなどの炎症性サイトカインの遺伝子発現を抑制し ている.従って Bach2 遺伝子欠損マウスの肺環境に おいて,Bach2 欠損 T 細胞で分泌されたこれらサイト カインが肺胞マクロファージの性質を変えている可能 性が示唆された.そこで,この可能性を検証するため に,複数の遺伝子改変マウスの表現型を解析した.ま ず,リンパ球を持たず Bach2 を欠損した Bach2/Rag2 二重欠損マウスを作成した.このマウスが肺胞蛋白症 を発症しなかったことから,次に,B 細胞特異的 Bach2遺伝子欠損マウスと T 細胞特異的 Bach2 遺伝 子欠損マウスを解析したところ,そのどちらでも肺胞 蛋白症を発症しなかった.しかし,T 細胞特異的 Bach2遺伝子欠損マウスの肺では,Bach2 遺伝子欠損 マウスと同様の炎症細胞の浸潤を認めた.これらの結 果から,肺胞マクロファージで発現している Bach2 が炎症のある環境下で機能し,性質の変化を食い止め ている可能性を考えた.さらに,筆者は,T 細胞特異 的 Bach2 遺伝子欠損マウスの肺胞マクロファージに おいて,Bach2 の発現が亢進していること,野生型マ ウスの肺胞マクロファージをインターフェロンγ で刺 激すると,Bach2 の発現が誘導されることを発見した. Bach2が,炎症関連遺伝子や脾臓赤脾髄マクロファー ジ特異的遺伝子の発現を抑制していることから,肺胞 マクロファージにおいて,Bach2 は,炎症反応に応答 した機能喪失を抑える guardian として機能し,細胞 間ネットワークを制御していることが明らかとなった (図 4)10). 3) 肺胞蛋白症の新規治療法の発見 筆者は,Bach2 遺伝子欠損マウスにおいて,抗体を 用いて CD4 陽性 T 細胞を枯渇させたところ,肺胞蛋 白症が治癒することを見出し,T 細胞を標的とした治 療が肺胞蛋白症の新規治療法となりうることを発見 した10). 図 4. 研究結果模式図
終 わ り に 本稿では,転写因子 Bach2 遺伝子欠損マウスを病 態モデルとした,続発性肺胞蛋白症の発症メカニズム の研究を紹介した.本研究から得た新たな知見をさら に活かし,免疫応答の異常に起因する呼吸器疾患の病 態解明や,新規治療法の考案,癌免疫療法における課 題の克服など,臨床における問題点の解決につながる ような研究を続けていきたい. 筆者は,本研究を遂行するまでに,二人の子供の妊 娠・出産を経験した.その都度,女性研究者が研究を 続けることの大変さと,周囲のサポートのありがたさ を実感するばかりであった.女性ならではのライフイ ベントとキャリア構築における課題は,以前から各分 野で取りあげられてはいるが,今後さらに,情報の共 有と理解,支援制度の充実がなされることを心から 願っている. 最後に,ご指導いただいた東北大学大学院医学系研 究科 呼吸器内科学分野 貫和敏博前教授,一ノ瀬正 和教授,生物化学分野 五十嵐和彦教授,並びにご協 力いただいた国内外の多くの先生方に,心から感謝申 し上げます. 文 献
1) Trapnell, B.C., Whitsett, J.A. and Nakata, K. (2003) Pulmonary alveolar proteinosis. N. Engl. J. Med., 349 (26), 2527-2539.
2) Schneider, C., Nobs, S.P., Kurrer, M., et al. (2014) Induction of the nuclear receptor PPAR-gamma by the
cytokine GM-CSF is critical for the differentiation of
fetal monocytes into alveolar macrophages. Nat. Immunol., 15(11), 1026-1037.
3) Cordonnier, C., Fleury-Feith, J., Escudier, E., et al.
(1994) Secondary alveolar proteinosis is a reversible cause of respiratory failure in leukemic patients. Am. J. Respir. Crit. Care Med., 149(3 Pt 1), 788-794.
4) Ochiai, K., Katoh, Y., Ikura, T., et al. (2006) Plasma-cytic transcription factor Blimp-1 is repressed by
Bach2 in B cells. J. Biol. Chem., 281(50), 38226
-38234.
5) Muto, A., Ochiai, K., Kimura, Y., et al. (2010) Bach2 represses plasma cell gene regulatory network in B cells to promote antibody class switch. EMBO J., 29 (23), 4048-4061.
6) Roychoudhuri, R., Hirahara, K., Mousavi, K., et al. (2013) BACH2 represses effector programs to
stabi-lize T (reg)-mediated immune homeostasis. Nature,
498(7455), 506-510.
7) Itoh-Nakadai, A., Hikota, R., Muto, A., et al. (2014)
The transcription repressors Bach2 and Bach1 promote B cell development by repressing the myeloid program. Nat. Immunol., 15(12), 1171-1180.
8) Itoh-Nakadai, A., Matsumoto, M., Kato, H., et al.
(2017) A Bach2-Cebp Gene Regulatory Network for
the Commitment of Multipotent Hematopoietic Progenitors. Cell Rep., 18(10), 2401-2414.
9) Nakamura, A., Ebina-Shibuya, R., Itoh-Nakadai, A., et
al. (2013) Transcription repressor Bach2 is required for pulmonary surfactant homeostasis and alveolar macrophage function. J. Exp. Med., 210(11), 2191
-2204.
10) Ebina-Shibuya, R., Matsumoto, M., Kuwahara, M., et
al. (2017) Inflammatory responses induce an identity crisis of alveolar macrophages leading to pulmonary alveolar proteinosis. J. Biol. Chem., 292(44), 18098