Introduction 8
Historically, Echinacea has been used against infectious diseases and snake bites (Miller et al., 2004; Barrett, 2003). Among these modifications, a related P450s, F3'H, catalyzes B-ring hydroxylation, a common process in plant secondary metabolism. The F3'H gene was first isolated and characterized from petunia (Petunia hybrida) by Brugliera et al., 1999.
As, for example, in soybean, a single base deletion in the F3'H gene resulted in deletion of the heme-binding domain of F3'H and a change in pubescence color from brown to gray (Toda et al. 2002).
Materials and methods 10
- Plant materials 10
- Cloning of the full-length cDNA of EpF3'H 10
- Phylogenetic analysis 11
- Comparative and bioinformatic analysis 11
- Expression profile analyses 11
Quantification of gene expression was performed using the comparative Ct method (Bogs et al., 2006; Muller, Spontaneous mutations of the flavonoid 3'-hydroxylase gene conferring reddish flowers in the three morning glory species. Importance of the proline-rich region following signaling anchor sequence in forming the correct conformation of microsomal cytochrome P-450.
Virgin' is one of its characteristic selections in the Netherlands by planter Piet Oudolf in 2006. Whittal et al (2006) noted that the co-regulation of the DFR and ANS genes eliminated flower pigmentation in Aquilegia (Whittal et al., 2006). The expression levels of the structural genes EpDFR and EpANS were analyzed by semi-quantitative RT-PCR.
These results suggested that the difference in their respective colors was due to the difference in the levels of the anthocyanins. The PCR showed that none of the derived transcripts were differentially expressed between the EpDFRs in the wild type and 'Virgin'. I thought of this insertion sequence as a transposable element. The insertion sequence was a non-autonomous transposable element as it was too small to encode full-length transposons, which may have been like the insertion position of dTph3s in the RT gene (Kroon et al, 1994; Nakajima et al, 2005).
The insertion currently at 'Virgin' can no longer transpose due to the structural defect in the terminal reverse region. The PCR analysis (Fig. 3.2) showed that EpDFR was detected in the petals of the wild type and 'Virgin', and their amino acid sequences were completely identical. In the anthocyanin biosynthetic pathway, ANS is located downstream of the DFR (Fig 1.1), affects pathway efficiency and affects flower color depending on the species (Smith et al., 2012).
Several studies have also shown that the expression pattern of the ANS gene is consistent with the accumulation of anthocyanins (Gong et al., 1997; Saito et al., 1999; Shimada et al., 2005).

Results 12
- Isolation of the Full-Length cDNA of EpF3'H Gene 12
- Homology and structural characters of EpF3'H 12
- Comparison of EpF3'H sequences 13
- Gene expression of EpF3'H 13
Discussion 13
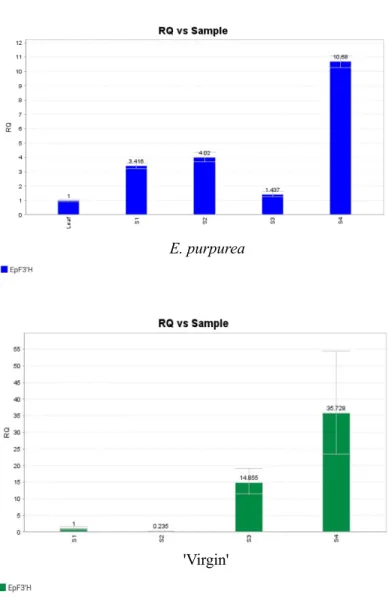
This was similar to the expression levels of LvF3'H in the lily cultivar 'Vivian' which was higher in the pigmented petals than in unpigmented petals (Yin et al., 2020). The possible explanation for this anomaly could be that EpF3'H is probably regulated by post-transcriptional regulation, such that its activation is inhibited, or like CHS in the petunia (Saito et al., 2006) which has different splicing forms. For example, F3'H has been hypothesized to be involved not only in anthocyanin biosynthesis but also in flavone biosynthesis (Kitada et al., 2001).
The results showed that the F3'H gene has different temporal and spatial expression patterns in different varieties, tissues and growth stages (Brugliera et al.,1999; Huang et al., 2007).
The possible explanations for this could be the unavailability of the relevant substrates in specific tissues, or the high transcriptional activity of EpF3'H in 'Virgin' exceeded the threshold level that triggers post-transcriptional silencing mechanisms (Wassenegger and Pélissier, 1998). Identification of the Arabidopsis thaliana Flavonoid 3'-Hydroxylase gene and functional expression of the encoded P450 enzyme. The identification of flavonoids and the expression of genes of anthocyanin biosynthesis in the chrysanthemum flowers.
Ectopic expression of apple F3'H genes contributes to anthocyanin accumulation in the Arabidopsis tt7 mutant grown under nitrogen stress. Functional analysis of Antirrhinum kelloggii flavonoid 3'-hydroxylase and flavonoid 3',5'-hydroxylase genes; critical role in flower color and evolution in the genus Antirrhinum. Differential expression of two cytochrome P450s involved in the biosynthesis of flavones and anthocyanins in chemovariety forms of Perilla frutescens.
Identification of the Arabidopsis thaliana flavonoid 3'hydroxylase gene and functional expression of the encoded P450 enzyme. Cloning, functional identification and sequence analysis of flavonoid 3'-hydroxylase and flavonoid 3',5'-hydroxylase cDNA reveal independent evolution of flavonoid 3',5'-hydroxylase in the Asteraceae family. Anti-inflammatory effect, total polysaccharide, total phenolic content and antioxidant activity of the aqueous extract of three basidiomycetes.
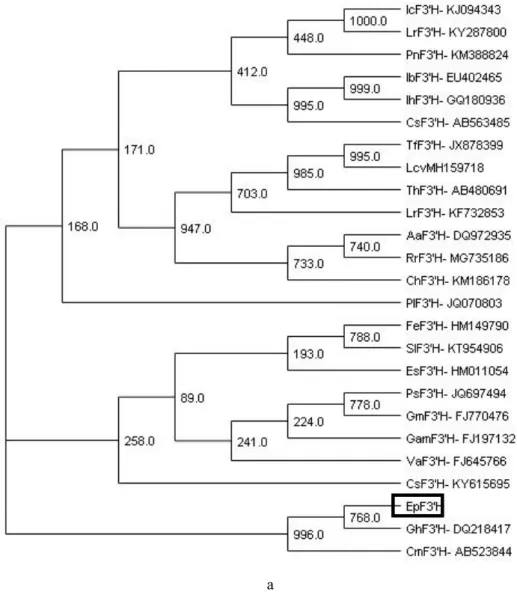
Integrated metabolic profiling and transcriptome analysis of pigment accumulation in diverse petal tissues in the lily cultivar 'Vivian'. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to derive the phylogenetic tree.
Introduction 31
Flower color is one of the most important traits of ornamental plants, which is influenced by different types of plant pigments, such as flavonoids, carotenoids, and betalains (Tanaka et al., 2008; Tripathi et al., 2018). There are more than 8,000 flavonoid derivatives (Veitch and Grayer, 2011) that act as UV protection and pigmentation in plants (Shirley, 1996; Fini et al., 2011). The most widespread group of flavonoids are anthocyanins, which are closely related to flower development (Weiss et al., 2000) and are responsible for most of the red, pink, purple, and blue colors of plants (Field et al., 2001; Honda and Saito, 2002; Miyagawa et al., 2015).
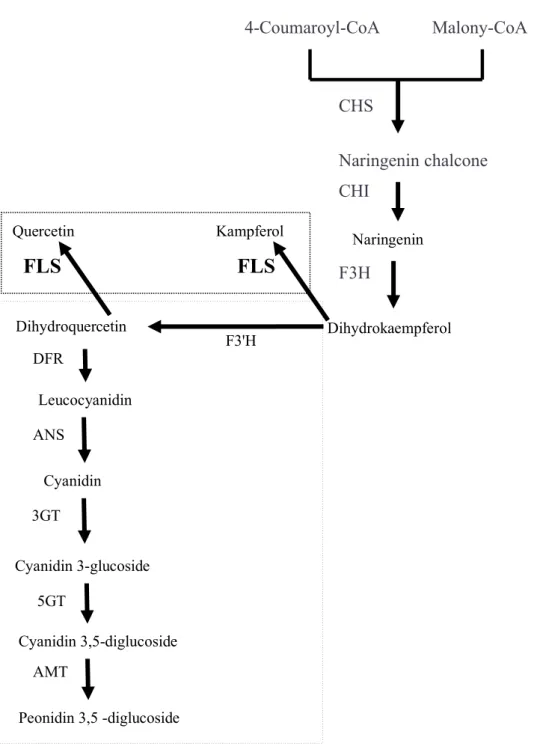
Many structural genes encoding enzymes in the anthocyanin biosynthetic pathway (Fig. 1.1, Holton and Cornish, 1995; .. Simhon et al., 2015) have been extensively studied and characterized in many plant species. The blockage in the early steps of flavonoid biosynthesis results in the formation of white flowers due to accumulation of colorless pigments, while the later steps are blocked by mutations resulting in the formation of differently colored flowers due to accumulation of a. Durbin et al. (2003) showed that color phenotypes in plants were closely associated with changes in the regulation of gene expression.
For example, due to the lack of F3′5′H, roses, carnations, chrysanthemums, lilies, and many other plant species do not have blue flowers. Mutations in the genes encoding flavanone 3'-hydroxylase (F3'H), DFR or ANS caused the accumulation of colorless flavonoids in the petals of white flower phenotypes A. In this study, I wanted to investigate the cause of white flower coloration in 'Virgin' and revealed their different functions involved in the flavonoid biosynthetic pathway.
To my knowledge, this is the first report documenting the molecular analysis of flower color mutations among E.
Materials and methods 32
- Plant materials 32
- UPLC analysis of flavonoid 32
- Total RNA Extraction 33
- Cloning of the full-length cDNA of EpDFR and EpANS 33
- Semi-quantitative RT-PCR 34
- Quantitative RT-PCR analysis 34
- Bioinformatics Analysis 35
A full-length cDNA sequence of EpDFR and EpANS was deduced by pooling the products of 3′-RACE,5′-RACE. The specific primers used to amplify the anthocyanin biosynthesis genes and their corresponding PCR primers are listed in Table 1. BLAST sequence analysis of the product showed that it was highly homologous to DFRs from other plant species.
In addition, flower color change by genetic engineering of the DFR gene has been reported for Rosa hybrida (Katsumoto et al., 2007), Dianthus caryophyllus (see review; . Tanaka et al., 1998), T. Insertion of the transposable element of EpANS in 'Virgin ' inhibited normal EpANS transcripts (Fig. 3.3) and caused low levels of incomplete EpANS transcripts (Fig. reported a non-autonomous DNA transposon insertion caused significant reduction in the transcription of the ANS gene, resulting in a pink color phenotype in onion A "white" anthocyanin-free pomegranate (Punica granatum L.) caused by an insertion in the coding region of the leucoanthocyanidin dioxygenase (LDOX; ANS) gene.
Cloning and structural analysis of the Rt anthocyanin pigmentation locus of Petunia hybrida: characterization of insertion sequences in two mutant alleles. Molecular analysis of instability in Antirrhinum majus flower pigmentation following isolation of the pallida locus by transposon tagging. The insertion in 'Virgin' is shown as a blue triangle and is located between positions 923–924 downstream of the ATG start codon.
My results contribute to the investigation of the possible function of the structural genes EpF3'H, EpDFR and EpANS from E. Test crosses between 'Virgin' and the wild type or other breeding lines should be performed to identify the function of the non-autonomous elements of transposable EpANS and used to breed more new color varieties.
Result 35
- Flavonoid analysis of E. purpurea and 'Virgin' 35
- Gene expression of EpDFR and EpANS 36
- DFR gene analysis 37
- Characterization of the EpANS sequence 37
Discussion 38
Based on both the sequence similarity of the homologue of TIR and the specific number of nucleotides comprising TSD caused by insertion of the element into the host genomic DNA, DNA transposable elements can be grouped into families (Kunze and Weil, 2002 ). One of the members is called MITEs, a heterogeneous group of small, non-autonomous elements, ranging from a few tens to a few hundred base pairs in size. The transposable element of EpANS in 'Virgin', a non-autonomous DNA transposon, does not contain ORFs encoding a transposase, it contains 8-bp terminal inverted repeats and 5-bp target site duplication as typical features of DNA transposons (Wicker et al. al., 2007), which may belong to MITEs.
2013) showed that using the transposases produced by active partners, non-autonomous elements without genes encoding active transposases can still transpose. The nucleotide sequences of the terminal ends of the transposable element of EpANS were imperfect inverted repeats, as there was one nucleotide difference between the TIR sequences. Such an imperfect TIR sequence could not be recognized by the transposase of the transposable element, so that the insertion could no longer be excised.
Hoshino et al (2003) also reported that a nonsense codon generated by the point mutations of F3'H in Ipomoea reportedly causes nonsense-mediated decay (NMD), affecting the stability of mature mRNA. Molecular cloning, substrate specificity of the functionally expressed dihydroflavonol 4-reductases from Malus domestica and Pyrus communis cultivars and the consequences for flavonoid metabolism. The gene encoding flavanone 3-hydroxylase is normally expressed in the pale yellow flowers of the Japanese morning glory that carries the speckled mutation that produces neither flavonol nor anthocyanin but accumulates chalcone, auron and flavanone.
Panels E and F show the specific accumulation of flavonols in petals of 'Virgin', the y-axes represent relative intensity of the spectrum, and the x-axes represent the mass-to-charge ratio. This suggested that EpF3'H are functional components of the coloration and may be crucial for anthocyanin B-ring hydroxylation pattern in E.
Summary 63
Future prospects and recommendations 65