グ ラ ム 陽 性 通 性 嫌 気 性 球 菌 で あ るStreptococcus
gordoniiは,口腔常在菌叢で優位を占めるMitis-group
の1菌種であり1),口腔内のいたるところに生息してい るが2),歯面初期定着細菌としてプラークバイオフィル ム形成の促進に関与するとともに,細菌性心内膜炎のよ うな日和見感染との関連が示唆されている細菌でもある
3, 4).グラム陽性細菌の宿主への感染には菌体表層タン
パク質や(リポ)タイコ酸などの菌体表層成分が重要な 役割を果たしていると考えられているが,S. gordoniiの 感染にかかわる菌体表層成分についてはあまり解明され てきていない.
S. gordoniiを含むグラム陽性球菌の菌体表層タンパ ク質の存在様式には7種類の結合様式が報告されてい
る5, 6).このうちタンパク質が細菌細胞壁に共有結合す
る様式は1種類のみで,この種のタンパク質はLPXTG タンパク質と総称され,近年ブドウ球菌やレンサ球菌を 中心に解析が進められてきている7, 8).これまでの報告 によると,LPXTGタンパク質は細菌感染に関わる重要 な機能を担っていることが明らかにされている.例え ば,組織や細胞への付着機能や細菌の凝集機能をもつタ ンパク質,付着・定着後の栄養獲得に関わる酵素タンパ
ク質,宿主免疫の抵抗性に関わるタンパク質などが報告 されている.また,グラム陽性菌は細菌種により保有す
るLPXTGタンパク質の種類と数が異なっている7, 8).
LPXTGタ ン パ ク 質 は 細 胞 壁 結 合 に 関 わ るsorting signal(細胞壁アンカー領域)と呼ばれる特徴的な配列 をC末端側にもつ7, 8).Sorting signalは5アミノ酸残基 のLPXTG motif, 25-30アミノ酸残基の疎水性領域,4-6 アミノ酸残基の荷電末端から構成されている.Sorting
signalをもつタンパク質の細胞壁への結合はsortaseと
呼 ば れ るcysteine protease-transpeptidaseに よ っ て 触
媒される7, 9).細胞膜上に局在するsortaseは,分泌中
に一時的に細胞膜にとどまったLPXTGタンパク質の LPXTG motifを認識し,T (threonine) とG (Glycine) の 間のペプチド結合を切断し,生じたT残基を細胞壁の 架橋構造をなすpentaglycine残基に共有結合する10).こ の反応によってLPXTGタンパク質は細胞壁に共有結合 した形で細胞表層に局在し,さまざまな機能を発揮する.
口腔レンサ球菌のLPXTGタンパク質の研究は,ヒ トのう蝕原性細菌であるStreptococcus mutansを中心 に展開されてきている.S. mutansは7種のLPXTGタ ンパク質 (PAc, WapA, WapE, GbpC, DexA, FruA, Cnm)
原 著
Streptococcus gordonii の菌体表層 nuclease の分子生物学的解析
高橋 真和,五十嵐 武
要旨:Streptococcus gordoniiは歯面初期定着細菌としてプラークバイオフィルム形成の促進に関与し,う 蝕や歯周病といった口腔感染症の誘発に深く関わるとともに,細菌性心内膜炎などの全身疾患との関連が 示唆されている細菌である.本研究ではS. gordoniiの菌体表層に局在するnucleaseの分子生物学的解析に ついて報告する.S. gordonii ChallisのゲノムデータベースからLPXTG motifをもつタンパク質を検索し,
その1つがnuclease (Nuc)ホモログであることを見出した.このnuclease遺伝子 (nuc)は2,340塩基対 で,780アミノ酸残基からなり84.5 kDaのタンパク質をコードし,その等電点は5.37であった.Nucの推 定アミノ酸配列は,N-末端にシグナルペプチド,分子中央に活性部位を含むpfam03372配列,そしてC-末 端に細胞壁アンカー領域 (sorting signal)を保持していた.Nucタンパク質はnuc遺伝子をpGEX-4T2ベク ターにクローニング後,単一の組換え体 (102 kDa) として精製した.この精製NucはCa2+ とMn2+ または Mg2+の存在下でDNA分解活性を示し,その至適pHは5.5であった.さらにNucに部位特異的変異を導入 すると,N362, E401, D578, H654の4種の変異体でDNA分解活性が消失した.また,菌体のDNA分解活性は,
野生株の抗Nuc血清処理で優位に抑制され,さらにnuclease欠損変異株で消失した.また,抗Nuc血清を
用いたWestern blotは,野生株の菌体表層でペプチドグリカン結合型Nucを検出したが,sortase欠損変異
株では検出しなかった.さらにS. gordoniiのNucホモログはMitis-groupのStreptococcus mitis, Streptococcus sanguinis, Streptococcus parasanguinis にも存在していた.以上の結果はS. gordonii Challisの菌体表層に局在
するnucleaseの分子生物学的性質を明らかにし,さらに,この酵素が環境中のDNAの利用に関与している
可能性を示唆した.
昭和大学歯学部口腔微生物学教室(主任:五十嵐 武教授)
(2010年8月30日受付;2010年9月14日受理)
をもち,このうち6種の機能が解析され,いずれもS.
mutansの感染に関わる重要な役割を担っていることが
明らかにされた.すなわち,PAc, WapA, GbpC, Cnmは 付 着・定 着・凝 集 に11〜14),DexAとFruAは 栄 養 獲 得
に15, 16),GbpCは免疫抵抗性に寄与している17).また,S.
mutansは sortaseを欠損するとこれらのタンパク質を
細胞表層に保持できなくなり,その結果,それらの機能 が消失することが明らかにされた16, 18〜22).
一方,近年公表されたS. gordonii Challisのゲノムデー タベース (http://www.oralgen.lanl.gov/oralgen/bacteria/
sgor)は,sortaseと推定LPXTGタンパク質の存在を示 唆している.S. gordoniiの推定LPXTGタンパク質もま た,S. mutansやその他のグラム陽性菌のそれと同様に 感染に関わる重要な機能をもつと推測されるが,機能未 知のものが多く,現在までに機能解析されたタンパク質 はSspA/Bの2種類にすぎない23).
我々はS. gordonii ChallisのLPXTGタンパク質の1つが,
Streptococcus pyogenes24), Streptococcus suis25)のnucleaseと相 同性が高いことを見出した.しかし,これらの菌種にお
けるnucleaseの生理的機能は未だ明らかにされていない.
そこで本研究では,S. gordonii Challisのnucleaseホモ ログの性質を分子生物学的に解析し,その生理機能を推 測した.
材料と方法 使用菌株と培養方法
供試菌株にはStreptococcus gordonii Challisを野生株と して用いた.S. gordoniiはTodd-Hewitt broth (TH broth)
(Difco Laboratories, Detroit, MI, USA) または1.5%寒天を 含むTH寒天平板を用い,37℃,嫌気的条件下 (80%N2, 10%H2, 10%CO2)で培養した.Escherichia coli JM109 と BL21 は Luria-Bertani broth (LB broth)(Invitrogen,
Carlsbad, CA, USA)またはその寒天平板を用い,37℃,
好気的条件下で培養した.各培地には必要に応じて ampicilin (50 µg/ml)ま た はerythromycin (10 µg/ml)
を添加した.
DNAの抽出
S. gordonii Challis の染色体DNAはGenElute Bacterial genomic DNA kit (Sigma-Aldrich, St. Louis, MO, USA) を 用いて抽出した.E. coliからのプラスミドDNA抽出は Wizard miniprep purification kit (Promega, Madison, WI,
USA) を用いて行った.
S. gordonii 変異株の作製
S. gordonii Challis のnuclease 欠損変異株(nuc欠損 変異株)ならびにsortase欠損変異株(srt欠損変異株)
は,Fig. 1に示すように各遺伝子の相同組換えによる 挿入不活化により作成した26).すなわち,標的遺伝子
(nuc とsrt)の5′および3′側の遺伝子断片をP1F-P2R お
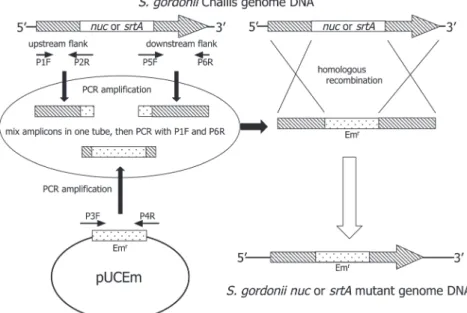
Fig. 1 Preparation of the nuclease- and the sortase-deficient mutants of S. gordonii Challis. Emr gene franking 5′- and 3′-regions of the target gene (nuc gene or srt gene) was introduced into S. gordonii Challis cells. The mutant generated by homologous recombination was screened on TH agar plate containing erythromycin. Replacement of target gene (the nuc gene or the srt gene) and Emr gene in the generated mutant was confirmed by PCR.
よびP5F-P6R primer pairで増幅した.一方,pUCEm内 のerythromycin耐 性 遺 伝 子(Emr)をP3F-P4R primer pairで増幅した26).P2Rの3′側とP5Fの5′側はそれぞ れEmr遺伝子の一部を含むように設計した.また,P3F とP4RはそれぞれP2RとP5Fの相補鎖として設計した.
得 ら れ た3つ の 増 幅 断 片(P1F-P2R断 片, P5F-P6R断 片,P3F-P4R断片)を混合後,P1FとP6RでPCR増幅 し,標的遺伝子の一部を両端に持つEmr遺伝子断片を得 た.次に,この増幅断片をPerryらの方法27)に準じてS.
gordonii Challisに導入後,erythromycinを含むTH寒天 平板上で選別し,nuc欠損変異株ならびにsrt欠損変異 株を得た.得られたEmr株のnucまたはsrt遺伝子の欠 損はPCRで確認した.その際の確認用PCRプライマー
はP1F-P4RおよびP3F-P6Rを使用した.使用したプラ
イマー配列をTable 1に示す.
組換えNucタンパク質の精製
S. gordonii ChallisのゲノムDNAを鋳型としてnuc遺 伝子をLA Taq DNA polymeraseを用いたLA-PCRで増 幅した.増幅にはShuttle PCR条件を使用し,その時の 増幅primers (P1F-fullnuc P2R-fullnuc)の配列をTable 1 に示す.Shuttle PCRの条件は,反応液 (50 µl)を94℃,
1 min熱変性させた後,98℃,20 sの熱変性と68℃,2
minのアニーリングと伸長反応を1サイクルとし,これ を28 cycle繰り返して,DNAを増幅した28).得られた
nuc遺伝子はpGEX-4T2 発現ベクターにクローニング後,
E. coli BL21に形質転換した.組換えNucタンパク質は
Glutathione-Sepharoseを用いたGST purification system
(GE Healthcare UK, Little Chalfont, UK)により精製し
た.精製Nucタンパク質は酵素活性の解析に使用する とともに,抗Nucウサギ血清作製のための抗原として 使用した.
Nuclease活性測定法
Nuc活性はpUC18プラスミドDNAまたはS. gordonii ゲノムDNAの分解能により検出した.反応液は1 µg pUC18, 1 mM CaCl2, 1 mM MnCl2, 0.1 µg Nucを含む混 合液をリン酸緩衝液pH 5.5で全量10 µlとした.反応液 中でのリン酸緩衝液の最終濃度は50 mMとした.この 反応液を37℃で60分間保温後,1.0%アガロースゲルで 泳動分離し,臭化エチジウムでDNA断片を染色した.
また,pUC18の代わりにS. gordonii ゲノムDNAを基質 とする場合は,50 ngを使用した.
金属イオン要求性の検討では,上記の反応をもとに CaCl2, MnCl2, MgCl2の濃度を0 mMまたは 1 mMとし,
その組合せによりDNA分解活性の有無を検討した.
至適pHの測定では上記の反応をもとに,酢酸緩衝液
(pH 3.5 6.0)またはリン酸緩衝液 (pH 5.5 8.0)を用い,
それぞれのpH範囲でDNA分解活性を測定した.反応 液中での緩衝液の最終濃度はいずれも50 mMとした.
S. gordonii菌体のnuclease活性の測定
S. gordonii Challisと nuc欠 損 変 異 株 をTH broth で 37℃,一晩,嫌気培養した後,遠心分離により集菌し た.その菌体をTH broth で懸濁し,OD600=0.1に調整後,
37℃で3時間嫌気培養した.その後,培養液を遠心分離 し,集菌した菌体を50 mMリン酸緩衝液pH 5.5に懸濁 後,その菌懸濁液の一部を酵素活性測定に使用した.ま た,必要に応じて抗Nuc血清で処理したChallis菌体を Table 1 Oligonucleotide primers and plasmid used in this study
Primers/plasmid Sequences/relevant characteristics1 References
Primers
P1F-nuc CCAATTACGGCAGGACTCAGATTG This work
P2R-nuc GAGAATATTTTATATTTTTGTTCATGGAACAGCCAAGGAAAGCAAAGGT This work
P3F-nuc ACCTTTGCTTTCCTTGGCTGTTCCATGAACAAAAATATAAAATATTCTC This work
P4R-nuc AGAAACCAAAGGCTACCCAATGCCTTATTTCCTCCCGTTAAATAATAGA This work
P5F-nuc TCTATTATTTAACGGGAGGAAATAAGGCATTGGGTAGCCTTTGGTTTCT This work
P6R-nuc GCTAGGCACCTATCTTCAAACAAACTC This work
P1F-srt AAGGCCGTGGTGGTAAAGGGATTA This work
P2R-srt GAGAATATTTTATATTTTTGTTCATTTCTTGGCTCTTCGTGATCCTCTG This work
P3F-srt CAGAGGATCACGAAGAGCCAAGAAATGAACAAAAATATAAAATATTCTC This work
P4R-srt CGGTGTTTCTTTATATGGAGTTTCGCTTATTTCCTCCCGTTAAATAATAGA This work
P5F-srt TCTATTATTTAACGGGAGGAAATAAGCGAAACTCCATATAAAGAAACACCG This work
P6R-srt AGTTCAGACAGAGTCATGTGACGG This work
P1F-fullnuc TTTGAATTCCCATGAAGAAACAAGCTTATCAAAAGCG This work
P2R-fullnuc TTTCTCGAGTTACTCTAGGTTAGATGAC This work
Plasmid
pUCEm pUC18 carrying Emr 26
1 Emr(erythromycin resistance). Single underline, EcoRI site; double underline, XhoI site.
酵素活性測定に使用した.この際,抗Nuc血清での処 理時間は4℃,3時間とし,50 mMリン酸緩衝液pH 5.5 で洗浄後,使用した.
Nuclease変異体の作製
Nucへの部位特異的変異の導入にはQuikChange site- directed mutagenesis kit (Stratagene, La Jolla, CA, USA)
を使用し,その方法に従ってNuc分子内の標的アミノ 酸を置換し,変異体を作製した28, 29).
細胞壁結合型nucleaseの抽出と解析
S. gordonii Challis 菌体を50 mM リン酸緩衝液pH 6.0 で洗浄後,同緩衝液に再懸濁し,フレンチプレスで菌体 を2回破砕した.その後,遠心分離により沈殿を回収し,
lysozyme(最終濃度10 mg/ml)とmutanolysin(最終 濃度200 units/ml)を含む50 mM Tris-HCl, pH 8.0に懸 濁後,56℃,40分間処理し30),遠心分離により沈殿を 得た.この沈殿を細胞壁抽出画分の試料とした.細胞壁 結合型Nucの検出は,抽出試料を7.5% SDS-PAGEで分 離後,抗Nuc血清を用いたWestern blot法で解析した.
S. gordonii nucleaseホモログの検索
S. gordonii nucleaseホモログの検出は nuclease遺伝子 の全長をプローブとした Southern hybridizationにより 行った.検索には次のレンサ球菌10菌種を使用した.S.
mutans GS5, S. sobrinus ATCC27607, S. downei NCTC11391, S. gordonii Challis, S. gordonii ATCC10558, S. sanguinis ATCC10556, S. mitis ATCC9811, S. oralis ATCC35037, S.
salivarius JCM5707, S. pyogenes ATCC12388.
また,ゲノムデータベースが公開されている菌種につ いては,Nucの推定アミノ酸配列を基にデータベース検 索を行った.
結 果
S. gordonii のnuclease遺伝子ホモログの解析 S. gordonii Challisのゲノムデータベース(http://www.
oralgen.lanl.gov/oralgen/bacteria/sgor)か らLPXTG motif をもつタンパク質を検索したところ,24種類が検出さ
れ,その1つが推定nuclease ホモログであることを
見 出 し た.こ のnuclease遺 伝 子(nuc)(accession no.
SGO1651)は2,340塩基対で,780アミノ酸残基からな
り84.5 kDaのタンパク質をコードし,その等電点は5.37
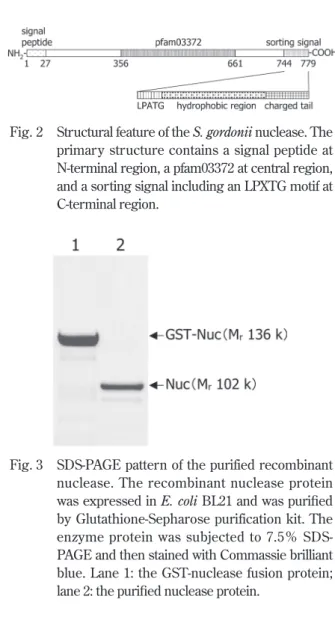
であった.Fig. 2に示すようにNucの推定アミノ酸配列 は,N-末端にシグナルペプチド(27アミノ酸残基),分 子中央には酵素活性に関連すると推測されるpfam03372 領域(306アミノ酸残基),C-末端にはLPXTG motifを含 むsorting signal(36アミノ酸残基),すなわち細胞壁ア ンカー領域を保持していた.
組換えnucleaseタンパク質の精製
Fig. 3に精製組換えNucタンパク質のSDS-PAGEパ ターンを示す.Nucタンパク質は単一のバンドとして精 製され,その分子量は102 kDaを示した (Fig. 3-lane 2).
この分子量は推定アミノ酸配列から算出した推定分子量
84.5 kDaよりも若干大きい値を示した.
Nucleaseの金属イオン要求性と至適pH
精製NucをpUC18プラスミドDNAと反応させたが,
DNA分解活性は検出されなかった(Fig. 4-lane 1).既
知のnucleaseはその活性発現に2価の金属陽イオンを
必要とすることから,S. gordonii のNucについても金 属イオン要求性を検討した.Fig. 4-lanes 2, 3, 4に示す ようにNucはCa2+, Mn2+, Mg2+の単独添加では,有意 なDNA分解能を示さなかった.一方,2種のイオンを 添加した場合は,Ca2+とMn2+およびCa2+とMg2+の 添加でDNAが完全に分解されたが (Fig. 4-lanes 5, 6),
Mn2+ とMg2+ の添加では,有意な分解活性を示さなかっ た (Fig. 4-lane 7).
Nucの金属イオン要求性の結果をもとに,Ca2+と
Mn2+ の存在下でNucの至適pHを検討した.酢酸緩衝
Fig. 2 Structural feature of the S. gordonii nuclease. The primary structure contains a signal peptide at N-terminal region, a pfam03372 at central region, and a sorting signal including an LPXTG motif at C-terminal region.
Fig. 3 SDS-PAGE pattern of the purified recombinant nuclease. The recombinant nuclease protein was expressed in E. coli BL21 and was purified by Glutathione-Sepharose purification kit. The enzyme protein was subjected to 7.5% SDS- PAGE and then stained with Commassie brilliant blue. Lane 1: the GST-nuclease fusion protein;
lane 2: the purified nuclease protein.
液を用いたpH 3.5 6.0の範囲では,pH 5.5とpH 6.0で DNA分解活性が最も強く,リン酸緩衝液を用いたpH 5.5 8.0の範囲では,pH 5.5で最も強いDNA分解活性を 示した(data not shown).また,Ca2+ とMg2+ の存在下 での至適pHは,Ca2+ とMn2+ の存在下の場合と同様な 結果を示した (data not shown).
以上の結果から,S. gordoniiのNucはCa2+とMn2+
またはMg2+要求性のDNA分解酵素で,その至適pH は5.5であることが明らかになった.
Nucleaseの活性部位の解析
Nuc酵素の活性部位を解析するために,pfam03372領 域をもち既に結晶構造解析が報告31)されているSchizo- saccharomyces pombe の synaptojanin (SPsynaptojanin;
inositol-5-phosphatase)の活性部位の立体構造を基に,S.
gordonii のNucで相同活性部位を検索し,活性に重要と
考えられる7種のアミノ酸残基,N362, E401, H525, H545,
D578, D621, H654を選んだ.これらのアミノ酸残基の酵
素活性への関与を明らかにするために,各アミノ酸に対 応するコドンに部位特異的変異を導入し,Glycine (G) に 置換した変異体,すなわちN362G, E401G, H525G, H545G, D578G, D578E, D621G, H654Gを 作 製 し た.Fig. 5に 各 変異体のDNA分解活性を示した.その結果,変異体 N362G, E401G, D578G, D578E, H654GでNuc 活性が消失 し (Fig. 5-lanes 2, 3, 6, 7, 9),その他の変異体では活性に影 響がなかった.以上の結果から,N362, E401, D578, H654 の4種のアミノ酸残基がNuc活性に重要であることが明 らかになった.
Nucleaseの細胞局在性の解析
Nuc分子はそのC-末端にLPXTG motifを含むsorting signalを保持することから (Fig. 2),Nucはsortaseの 触媒作用で細胞壁に結合して存在している可能性が強 く示唆された.これを明らかにするために,S. gordonii をlysozymeとmutanolysinで処理後,細胞表層画分を 抽出し,抗Nuc血清を用いたWestern blotでNucを検 出した.Fig. 6で示すようにChallis野生株では高分子
量のNuc-Peptidoglycan複合体が複数のラダーバンドと
して検出されたが (Fig. 6-lane W),srt欠損変異株では それらのラダーバンドが検出されず,Nucのみが検出 された (Fig. 6-lane S).また,nuc欠損変異株ではNuc- Peptidoglycan複合体のみならず,Nucも検出されなかっ Fig. 4 Requirement of divalent metal ions in the nuclease
activity of S. gordonii. Nuclease activity was measured at pH5.5 under the presence of divalent cations such as Ca2+, Mn2+, and Mg2+. Each divalent metal ion was used at a final concentration of 0 mM (−) or 1 mM (+). The enzyme activity was analyzed by 1.0% agarose gel stained with ethidium bromide.
Fig. 6 Localization of the S. gordonii nuclease. The strain Challis, the sortase mutant, and the nuclease mutant of S. gordonii were treated with lysozyme and mutanolysin. The surface components of these cells were extracted with sample buffer and then analyzed by 7.5% SDS- PAGE and subsequent Western blotting with anti- Nuc serum. Lane M: molecular weight marker;
lane W: strain Challis; lane S: sortase-deficient mutant; lane N: muclease-deficient mutant.
Fig. 5 Analysis of essential amino acid residues in the active site of nuclease. An amino acid residue in the active site was converted to glycin (G) by site-directed mutagenesis. The resultant mutants were incubated with DNA (pUC18) under the presence of Ca2+ and Mn2+ and the enzyme activity was analyzed by 1.0% agarose gel stained with ethidium bromide. Lane M: size marker;
lane 1: wild type; lane 2: N362G; lane 3: E401G;
lane 4: H525G; lane 5: H545G; lane 6: D578G;
lane 7: D578E; lane 8: D621G; lane 9: H654G;
lane 10: pUC18 DNA.
た (Fig. 6-lane N).加えて,Challis野生株とnuc欠損変 異株の菌体が持つpUC18 DNA分解能を調べたところ,
ChallisはpUC18 DNAを分解したが,nuc欠損変異株 では分解能が消失していた (data not shown).さらに,
ChallisのDNA分解活性は,Challis菌体の抗Nuc血清 処理により有意に抑制された (data not shown).
S. gordonii nuclease ホモログの検索
S. gordonii nuclease ホモログをSouthern hybridization 法とデータベース検索の2つの方法で調べた.Nuc遺 伝子全長をプローブとしたSouthern hybridizationでは 調べたレンサ球菌10菌種のうち,Mitis-groupに属す る S. mitis ATCC9811, S. sanguinis ATCC10556, S. gordonii Challis, S. gordonii ATCC10558の3菌種4菌株でホモログ が検出された (data not shown).一方,Nucの推定アミノ 酸配列を基にしたデータベース検索では,Mitis-group で はS. sanguinis SK36 (83%ホモロジー)と S. parasanguinis ATCC15912 (71%), 一 般 細 菌 で はS. pyogenes M1 GAS
(63%), S. suis SC84 (66%), Bacillus anthracis CI (57%), Bacillus cereus AH1271 (57%)でホモログが検索された.
考 察
Streptococcus gordoniiは口腔常在菌として口腔内に幅広
く分布し1, 2),歯面初期定着細菌としてプラークバイオ
フィルム形成の促進に関与するとともに,細菌性心内膜 炎との関連が示唆されている細菌である3, 4).S. gordonii の表層タンパク質は口腔粘膜細胞や唾液をコートした歯 面ならびに血管内皮細胞などへの付着に重要な役割を 果たしており,種々の解析が進められている.例えば,
SspA/BとCshA/Bは唾液アグルチニン (gp340)と結合 し4),AbpAは唾液amylaseに結合する32).さらにGspB とHsaは,いずれもserineに富むglycoproteinであり,
唾液中のMUC7, 分泌型IgA, gp340のシアル酸分子に結
合することが知られている33, 34).S. gordoniiにおけるこ れらの表層タンパク質の存在様式は様々で,SspA/Bの
みがLPXTGタンパク質として細胞壁に共有結合してい
る23).近年,S. gordonii DL1 のsortase欠損変異株は口
腔keratinocyteや唾液コートした表層への結合が優位に
減少するとの報告がなされた35).この結果は,Bolken らや Nobbsらの報告,すなわちS. gordoniiのsortase 欠損変異株はfibronectinや唾液agglutininとの結合が 減少し,さらにマウスの口腔内へのコロニー形成能が 減少したという報告と一致している23, 36).このことは
LPXTGタンパク質がS. gordoniiの口腔内への付着・定
着に重要な役割を果たしていることを示唆しているが,
LPXTGタンパク質に関する研究はほとんど進展してい
ないのが現状である.
本研究で我々はS. gordonii Challisのゲノムデータベー
ス上でLPXTG motifをもつタンパク質を検索し,24種
の推定LPXTGタンパク質を見出した.そのうちの1つ
がS. suis SC84 やS. pyogenes M1 GASのnucleaseとそれ ぞれ66%, 63%の相同性を示すnuclease (Nuc) ホモログ であることを見出した.遺伝子解析の結果,S. gordonii
Nuc ホモログはそのアミノ酸一次構造において,N末端
にsignal peptideをもち,分子中央にはNuc活性に関連 すると推測されるpfam03372領域,そしてC末端には細 胞壁への結合に関わるsorting signalを有していた (Fig.
2).Sorting signalの存在は,Nucがsortaseにより細胞 壁に結合されて細胞表層に局在する可能性を強く示唆す る.この可能性はlysozymeと mutanolysin処理したS.
gordonii菌体からのNuc-Peptidoglycan複合体の検出に より確認された(Fig. 6).すなわち,Nuc-Peptidoglycan
複合体はChallis野生株から検出され,srtA欠損変異株
からは検出されなかった.この結果はsortaseがNucの 細胞壁への結合を触媒していることを示している.さら
にChallis株を抗Nuc血清で処理すると,Nuc活性が優
位に抑制された(data not shown)ことも,Nucが細胞 表層に局在していることを示唆している.これまでにレ ンサ球菌における細胞壁結合型nucleaseの存在は,S.
suis25)とS. pyogenes24)で報告されているが,口腔レンサ 球菌における報告例はなく,本研究が最初の報告となる.
本実験で作製したnuc欠損変異株はpUC18 DNA分 解能が消失していたことから(data not shown),見か け上,NucがS. gordonii Challis株の菌体表層に局在す る唯一のDNA分解酵素のように思われた.しかし,S.
gordonii Challisのゲノムデータベース検索結果は,Nuc
以外にLPXTG motifをもち細胞表層に局在すると考え
られる推定5′-nucleotidase (accession no. SGO1247)の 存在を示した.また,DaviesらはS. gordoniiのsortase 欠損変異株では推定5′-nucleotidaseの局在性が細胞表 層から培養上清へと変化したと報告した35).この結果 は,推定5′-nucleotidaseがsortaseの支配下にあり,細 胞壁結合型として局在していることを示唆している.す
なわちS. godoniiは2種類の細胞壁結合型DNA分解酵
素をもつことになる.おそらくそれぞれの酵素の機能 から推測すると,NucがDNAを分解して生じたヌクレ
オチドを5′-nucleotidaseがさらに分解し,菌体内に取
り込んで利用するのではないかと推測される.Nucと
5′-nucleotidaseが機能連携するために,両酵素が細胞表
層に局在することは合理的なのかもしれない.両酵素の 連携をより明確にするには,今後の推定5′-nucleotidase の解析が必要とされる.Group A streptococciやGroup B streptococciを 含 む そ の 他 の レ ン サ 球 菌 は 菌 体 外
nucleaseをもち,さらに一部の細菌では推定上の細胞壁 結合型nucleaseをもつとの報告があるが24, 37),本研究 で示唆したような2種類の細胞結合型nucleaseをもつ 報告はなく,本研究が初めての例となる.
細菌の産生するnucleaseはその活性に2価の金属イオ ンを要求する.S. gordoniiのNucはCa2+とMn2+または Mg2+の存在下でのみDNA分解活性を示した(Fig. 4).
このことはNucが2種の金属イオン,すなわちCa2+と Mn2+またはCa2+とMg2+要求性の酵素であること示唆 した.この結果は,S. suisのnucleaseがCa2+とMg2+
の2種の金属イオンを要求する酵素であることと類似し ていた25).
S. gordonii Nucはその分子中央にNuc活性に関連す ると推測されるpfam03372領域を保持していた(Fig.
2).pfam03372領 域 はexonuclease, endonuclease, phos-
phataseに共通して存在することが知られている.我々
はS. gordonii のNuc活性に重要なアミノ酸を調べるため
に,既に結晶構造解析31)されているSchizosaccharomyces pombe の synaptojanin (inositol-5-phosphatase)(SPsyna-
ptojaninと呼ばれる)の活性部位の立体構造を基に,S.
gordonii Nucのpfam03372配列で相同活性部位を検索し,
活性に重要と考えられる7種のアミノ酸残基,N362, E401, H525, H545, D578, D621, H654を選び,これらのア ミノ酸をG (Gly)に置換した変異体,すなわちN362G, E401G, H525G, H545G, D578G, D621G, H654Gを作製し,
そのNuc活性を測定した.その結果,変異体N362G, E401G, D578G, H654GでNuc 活性が消失したことから,
N362, E401, D578, H654の少なくとも4種のアミノ酸残 基がNuc活性に重要であることが明らかになった(Fig.
5).細胞壁結合型Nuc酵素の活性に重要なアミノ酸残基
を明らかにしたのは本研究が初めての報告となる.
S. gordoniiを含むMitis-groupの口腔レンサ球菌はプ ラークバイオフィルム形成を促進し,プラーク構成細菌 として優位を占める.その結果,プラーク内はそれら の細菌の主要代謝産物である有機酸(ギ酸,酢酸,乳 酸など)の蓄積により酸性(pH 5.0 6.0)環境になる.
S. gordoniiのNucの至適pHは5.5 (data not shown)で あることから,プラーク内の酸性環境はNucの作用に 適しているものと考えられる.プラークの成熟は酸素 分圧やpHなどの内部環境の変化をもたらし,プラー ク内細菌叢もまたその環境に適応して変化する.S.
gordonii のnuclease ホモログはS. mitis, S. sanguinis, S.
parasanguinis といったプラーク細菌叢で優位を占める
Mitis-group の細菌種に分布する(data not shown)こと から,これらのnucleaseは環境に適応できずに死滅し たプラーク細菌由来のDNAを分解し,栄養源として利
用しているのかもしれない.ある種の溶血性レンサ球菌 ではnucleaseがoligonucleotidesの栄養供給源であると の報告があり38),我々の推測を示唆している.これとは 別に,S. gordoniiのnucleaseは外来遺伝子による形質 転換能に影響を与えているのかもしれない.S. gordonii を含むある種の口腔細菌は外来遺伝子を取り込み,新た な性質を獲得する.多菌種が高密度で存在するプラーク 内では,このような形質転換の機会が多いと考えられ る.そのため菌体表層に局在するnucleaseは,外来遺 伝子を分解し,菌体内への不必要な遺伝子の侵入を防御 しているのかもしれない.我々は予備実験でS. gordonii
Challis株とnuc欠損変異株に対するプラスミドDNAの
形質転換効率を比較したが,両菌株間で顕著な差は見ら れず,この推測を示唆するには至らなかった.
本研究では口腔常在菌の1菌種であるS. gordoniiの
菌体表層nucleaseを分子生物学的に解析し,その性質
を明らかにした.Nucleaseを菌体表層にもつ細菌種の 報告が少ないこともあって,その生理学的役割につい ては未だ明らかにされていない.上述のように,口腔レ ンサ球菌ではS. gordoniiを含むMitis-groupの細菌種が
菌体表層nucleaseをもつことから,今後はそれらの菌
種の菌体表層nucleaseの性質を明らかにするとともに,
その生理学的役割を解明していく必要がある.
文 献
1) Schachteie CF, Nobbs A, Zhang Y, Costalonga M, Herzberg MC: Oral streptococci: commensals and opportunistic pathogens. In Hakenbeck R, S Chhat- wal (eds): The Molecular Biology of Streptococci.
Norfolk, UK, 2007, Horizon Scientific Press, pp. 411 462
2) Aas JA, Paster BJ, Stokes LN, Olsen I, Dewhirst FE:
Defining the normal bacterial flora of the oral cavity. J Clin Microbiol, 43: 5721 5732, 2005
3) Kerrigan SW, Jakubovics NS, Keane C, Maguire P, Wynne K, Jenkinson HF, Cox D: Role of Streptococcus gordonii surface proteins SspA/SspB and Has in platelet function. Infect Immun, 75: 5740 5747, 2007 4) Takamatsu D, Bensing BA, Cheng H, Jarvis GA, Siboo
IR, Lopez JA, Grifiss JM, Sullam PA: Binding of the Streptococcus gordonii surface glycoproteins GspB and Has to specific carbohydrate structures on platelet membrane glycoprotein. Mol Microbiol, 58:
380 392, 2005
5) Chhatwal GS: Anchorless adhesions and invasions of Gram-positive bacteria: a new class of virulence factors. Trends Microbiol, 10: 205 208, 2002
6) Ton-That H, Marraffini LA, Schneewind O: Protein sorting to the cell wall envelope of Gram-positive bacteria. Biochim Biophys Acta, 1694: 269 278, 2004 7) Navarre WW, Schneewind O: Surface proteins of
gram-positive bacteria and mechanisms of their targeting to the cell wall envelope. Microbiol Mol Biol Rev, 63: 174 229, 1999
8) Marraffini LA, DeDent AC, Schneewind O: Sortase and the art of anchoring proteins to the envelopes of gram-positive bacteria. Microbiol Mol Biol Rev, 70:
192 221, 2006
9) Ton-That H, Liu G, Mazmanian SK, Faull KF, Schne- ewind O: Purification and characterization of sortase, the transpeptidase that cleaves surface proteins of Staphylococcus aureus at LPXTG motif. Proc Natl Acad Sci USA, 96: 12424 12429, 1999
10) Paterson GK, Mitchell TJ: The biology of Gram- positive sortase enzymes. Trends Microbiol, 12: 89 95, 2004
11) Lee SF, Boran TL: Roles of sor tase in sur face expression of the major protein adhesin P1, saliva- induced aggregation and adherence, and cariogeni- city of Streptococcus mutans. Infect Immun, 71: 676 681, 2003
12) Qian H, Dao ML: Inactivation of the Streptococcus mutans wall-associated protein A gene (wapA) results in a decrease in sucrose-dependent adherence and aggregation. Infect Immun, 61: 5021 5028, 1993 13) Sato Y, Yamamoto Y, Kizaki H: Cloning and sequence
analysis of the gbpC gene encoding a novel glucan- binding protein of Streptococcus mutans. Infect Immun, 65: 668 675, 1997
14) Sato Y, Okamoto K, Kagami A, Yamamoto Y, Igarashi T, Kizaki H: Streptococcus mutans strains harboring collagen-binding adhesion. J Dent Res, 83: 534 539, 2004
15) Burne RA, Penders JEC. Characterization of the Streptococcus mutans GS5 fruA gene encoding exo-β- d-fructosidase. Infect Immun, 60: 4621 4623, 1992 16) Igarashi T, Asaga E, Goto N: Roll of Streptococcus
mutans dextranase anchored to the cell wall by sortase. Oral Microbiol Immunol, 19: 102 105, 2004 17) Nomura R, Nakano K, Ooshima T: Contribution of
glucan-binding protein C of Streptococcus mutans to bacteremia occurrence. Arch Oral Biol, 49: 783 788, 2004
18) Igarashi T, Asaga E, Goto N: The sortase of Strepto- coccus mutans mediates cell wall anchoring of a surface protein antigen. Oral Microbiol Immunol, 18:
266 269, 2003
19) Igarashi T, Asaga E, Sato Y, Goto N: Inactivation of srtA gene of Streptococcus mutans inhibits dextran- dependent aggregation by glucan-binding protein C.
Oral Microbiol Immunol, 19: 57 60, 2004
20) Igarashi T: Deletion in sortase gene of Streptococcus mutans Ingbritt. Oral Microbiol Immunol, 19: 210 213, 2004.
21) Murai C, Igarashi T, Inoue M, Sasa R: Streptococcus mutans sortase catalyzes cell wall anchoring of WapA and FruA. Ped Dent J, 15: 127 133, 2005
22) Haseba Y, Morisaki H, Takahashi M, Igarashi T: Role of sortase in biofilm formation of Streptococcus mutans. Dentistry in Japan, 43: 16 29, 2007
23) Nobbs AH, Vajna RM, Johnson JR, Zhang Y, Erlandsen DL, Oli MW, Kreth J, Brady LJ, Herzberg MC: Consequences of a sortase A mutation in Strepto- coccus gordonii. Microbiol, 153: 4088 4097, 2007 24) Janulczyk R, Rasmussen M: Improved pattern for
genome-based screening identifies novel cell wall- attached proteins in Gram-positive bacteria. Infect Immun, 69: 4019 4026, 2001
25) Fontaine MC, Perez-Casal J, Willson PJ: Investigation of a novel DNase of Streptococcus suis serotype 2.
Infect Immun, 72: 774 781, 2004
26) Arimoto T, Igarashi T: Role of prolipoprotein diacyl- glyceryl transferase (Lgt) and lipoprotein-specific signal peptidase II (LspA) in localization and physio- logical function of lipoprotein MsmE in Streptococcus mutans. Oral Microbiol Immunol, 23: 515 519, 2008 27) Perry D, Kuramitsu HK. Genetic transformation of
Streptococcus mutans. Infect Immun, 32: 1295 1297, 1981
28) Yano T, Fukamachi H, Yamamoto M, Igarashi T:
Characterization of l -cysteine desulfhydrase from Prevotella intermedia. Oral Microbiol Immunol, 24:
485 492, 2009
29) Igarashi T, Morisaki H, Yamamoto Y, Goto N: An essential amino acid residues for catalytic activity of the dextranase of Streptococcus mutans. Oral Micro- biol Immunol, 17: 193 196, 2002
30) Nadkarni MA, Martin FE, Hunter N, Jacques NA:
Methods for optimizing DNA extraction before quan- tifying oral bacterial numbers by real-time PCR. FEMS Microbiol Lett, 296: 45 51, 2009
31) Chi Y, Zhou B, Wanf W-Q, Chung S-K, Kwon Y-U, Ahn Y-H, Chang Y-T, Tsujishita Y, Hurley JH, Zhang A-Y:
Comparative mechanistic and substrate specificity study of inositol-5-phosphatase Schizosaccharomyces pombe synaptojanin and SHIP2. J Biol Chem, 279:
44987 44995, 2004
32) Rogers JD, Haase EM, Brown AE, Douglas CWI, Gwynn JP, Scannapieco FA: Identification and analysis of a gene (abpA) encoding a major amylase-binding protein in Streptococcus gordonii. Microbiol, 144:
1223 1233, 1998
33) Takahashi Y, Konishi K, Cisar JO, Yoshikawa M:
Identification and characterization of hsa, the gene encoding the sialic acid-binding adhesion of Strepto- coccus gordonii DL1. Infect Immun, 70: 1209 1218, 2002
34) Takamatsu D, Bensing BA, Prakobphol A, Fisher SJ, Sullam PM: Binding of the Streptococcal surface glyco- proteins GspB and Has to human salivary proteins.
Infect Immun, 74: 1933 1940, 2006
35) Davies JR, Svensater G, Herzberg CH: Identification of novel LPXTG-linked surface proteins from Strepto-
coccus gordonii. Microbiol, 155: 1977 1988, 2009 36) Bolken TC, Franke CA, Jones KF, Zeller GO, Jones CH,
Dutton EK, Hruby DE: Inactivation of the srtA gene in Streptococcus gordonii inhibits cell wall anchoring of the surface proteins and decreases in vitro and in vivo adhesion. Infect Immun, 69: 75 80, 2001
37) Ferreira BT, Benchetrit LC, De Castro AC, Batista TG, Barrucand L: Extracellular deoxyribonucleases of
streptococci: a comparison of their occurrence and levels of production among beta-hemolytic strains of various serological groups. Zentbl Bakteriol, 277:
493 503, 1992
38) Wilson AT: Nucleic acid derivatives as growth factors for certain group A hemolytic streptococci. Proc Soc Exp Biol Med, 58: 249 257, 1945
Molecular Analysis of the Nuclease Anchored to the Cell Wall of Streptococcus gordonii
Masakazu Takahashi and Takeshi Igarashi
Department of Oral Microbiology, Showa University School of Dentistry 1 5 8 Hatanodai, Shinagawa-ku, Tokyo, 142 8555 Japan
(Received August 30, 2010;Accepted for publication September 14, 2010)
Abstract:Streptococcus gordonii is related to the formation of plaque biofilm as an early colonizer and is closely associated with the induction of oral infectious disease. It has been also suggested that S.
gordonii is responsible for systemic disease such as bacterial endocarditis. In this study, we describe molecular analysis of a nuclease located on the surface of S. gordonii.
We searched the proteins with an LPXTG motif on the genome database of S. gordonii Challis and found a nuclease (Nuc) homologue. The nuclease gene (nuc) was 2,340-bp, which encoded a Nuc protein consisting of 780 amino acids with a molecular size of 84.5 kDa and a pI 5.37. The deduced amino acid sequence showed a signal peptide at the N-terminus, a pfam03372 sequence containing an active site at central and a sorting signal (cell wall anchoring region) at the C-terminus. Nuc was purified as a single protein band with molecular mass of a 102 kDa after the nuc gene was cloned into pGEX-4T2 vector. The purified protein showed nuclease activity under the presence of Ca2+ and Mn2+ or Mg2+ and it was highly active at pH 5.5. Replacement of N362, E401, D578 and H654 with Gly (G) resulted in complete disappearance of nuclease activity, implying that these amino acids are essential for the enzyme activity. In addition, nuclease activity of the strain Challis was apparently reduced by the treatment of anti-Nuc serum, and the nuclease-deficient mutant did not show the enzyme activity. Western blot analysis with anti-Nuc serum exhibited that the Nuc-Peptidoglycan complex was detected from the surface of the strain Challis, but not that of the sortase-deficient mutant, indicating that nuclease was anchored to the cell wall by sortase. The nuclease homologue of S. gordonii distributed to Streptococcus mitis, Streptococcus sanguinis and Streptococcus parasanguinis of Mitis-group.
These results clarified molecular characteristics of the nuclease anchored to the cell wall of S.
gordonii Challis and suggested that the nuclease might be responsible for utilization of environmental DNA.
Key words:LPXTG protein, nuclease, sortase, sorting signal, Streptococcus gordonii.