Instructions for use
Title FUS R495X 変異によるALS発症機構の解明
Author(s) 河原, 大貴
Citation 北海道大学. 博士(薬科学) 甲第13958号
Issue Date 2020-03-25
DOI 10.14943/doctoral.k13958
Doc URL http://hdl.handle.net/2115/80625
Type theses (doctoral)
File Information Daiki̲KAWAHARA.pdf
Hokkaido University Collection of Scholarly and Academic Papers : HUSCAP
博士学位論文
FUS R495X
変異によるALS
発症機構の解明2020
年 3 月北海道大学大学院生命科学院
生命科学専攻 生命医薬科学コース 神経科学研究室
河原 大貴
- 1 -
目次
1. 略語一覧・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・p. 2
2. 序論・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・p. 4
2.1 筋萎縮性側索硬化症(ALS) 2.2 Fused in Sarcoma (FUS) 2.3 FUS R495X変異
2.4相転移
2.5局所的翻訳機構 本研究の目的
3. 研究結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・p. 8
FUS R495X構造体形成機構の解明
構造体形成による細胞内変化の解明
4. 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・p. 15
5. 実験手法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・p. 19
6. 参考文献・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・p. 28
7. 図表・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・p. 35
8. 謝辞・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・p.69
2
1.略語一覧
a.a.: amino acid
ALS: Amyotrophic Lateral Sclerosis b. p.: base pair
cDNA: complementary deoxyribonucleic acid CDS: cording sequence
CLIP: UV-crosslink and immunoprecipitation DMEM: Dulbecco modified Eagle medium Dox: Doxycycline
ECL: enhanced chemiluminescence
EDTA: ethylene diamine N, N, N, N-tetraacetic acid EGFP: enhanced green fluorescent protein
ES cell: Embryonic stem cell EB: Embryonic body FUS: Fused in Sarcoma FBS: fetal bovine serum FL: full length
GDNF:Glial cell-derived neurotrophic factor
HITS-CLIP: High throughput sequencing of RNA isolated by CLIP HEPES: 2- [4- (2-hydroxyethyl)-1-piperazinyl] ethanesulfonic acid HEK:Human Embryonic Kidney
IgG: immunoglobulin G IP: immunoprecipitation kDa: kilo Dalton
LC: Low-complexity LB: Luria-Bertani
LIF:Leukemia Inhibitory Factor PBS: phosphate-buffered saline PCR: polymerase chain reaction PLA: Proximity Ligation Assay RGG: Arginine-Glycine-Glycine RRM: RNA recognition motif RT: reverse transcription
SDS-PAGE: sodium dodecyl sulfate polyacrylamide gel electrophoresis SQGY: Serine-Glutamine-Glycine- Tyrosine
3 TBS-T: Tris-bufferd saline with tween 20 Tris: tris (hydroxymethyl) aminomethane
Triton X-100: polyoxyethylen (10) octhylphenyl ether Tween-20: polyoxyethlene (20) solbitan monolaurate UTR: untranslated region
WB: western blot WT: wild type ZnF: Zinc-Finger
4
2.序論
2.1筋萎縮性側索硬化症
筋萎縮性側索硬化症 (Amyotrophic Lateral Sclerosis ,ALS ) は上位、下位の運動ニューロ ンが選択的かつ進行性に変性し、筋萎縮や筋力低下を生じる神経変性疾患である。症状の進 行に伴い、構音障害、嚥下障害、寝たきりになり、発症後3~5 年には呼吸筋の麻痺により 死に至る重篤な疾患であり、厚生労働省指定難病の一つに登録されている。患者数は日本だ けでも8000人を超え、加齢とともに頻度は増し、60歳以降の罹患危険率は300人に一人 とも言われおり、決して珍しい疾患でない。
ALS の発症者の 5~10%は遺伝性である (Figure 1A)。家族性 ALS の遺伝学的解析によ
り、1993年、第21番染色体にコードされているCu/Zn superoxide dismutase (SOD1) が 原因遺伝子として初めて発見された (Rosen DR et al.,1993) 。その後も解析は行われ、現 在では約50種類もの遺伝子に変異が同定されている (Boylan K ,2015)。注目すべき点とし て家族性患者の約半数を占める割合で、TAR DNA-binding protein 43 (TDP-43), Fused in Sarcoma (FUS), Senataxin, Angiogenin (ANG) , hnRNPA2B1, hnRNPA1などの RNA結合 タンパク質及び RNA 代謝に関わるタンパク質をコードする遺伝子に変異が同定されてお り (Figure 1B) 、RNA機能や代謝がALS発症に寄与する可能性が示唆されている。
また、神経病理学的特徴として、家族性、孤発性患者両者に共通して運動神経特異的にユ ビキチン免疫染色に陽性な異常構造体が形成される (Figure 2)。この異常構造体の構成タ ンパク質がTDP-43 やFUS等であることが同定され、現在、原因解明の突破口となってい る。
しかし、ALS発症機構は未解明な点が多く残されており、有効な治療法を確立する上で、
詳細なALS発症機構の解明が求められている。私はこれらの原因因子のうちFUSに着目し 解析を行った。
2.2 Fused in Sarcoma (FUS)
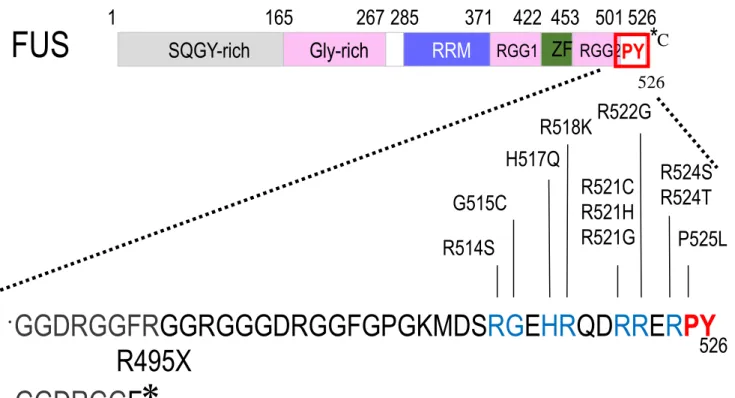
FUSは、526個のアミノ酸からなるRNA結合タンパク質であり、粘液性脂肪肉腫の癌化
を誘導する因子として同定された。EWS (Ewing’s Sarcoma) や TAF15 (TATA-binding protein-associated factor 15) らと共にTET familyに属し (Tan AY et al., 2009) 、N末端 から順に Serine-Glutamine-Glycine- Tyrosine (SQGY) rich ドメイン,Glycine-rich ドメイ ン, RNA recognition motif (RRM) ,Arginine -Glycine-Glycine (RGG) ドメイン,zinc-finger モチーフ,そして非古典的な核局在化シグナルであるPY-NLSをC末端に持つ (Figure 3A) 。 これらドメインのうちGly-rich, RRM, RGG, ZnF ドメインはRNA及びDNAと結合能を
5 有する (Law WJ et al., 2006) 。
FUSは全身で発現し、主に核内に局在する (Andersson et al., 2008 ; Morohoshi F et al.,
1996) 。しかしながら、核/細胞質間をシャトリングし、核内・細胞質内どちらにおいても
RNAと結合することで様々な機能を担う。現在までに報告されたFUSの機能としては、核 内におけるDNA修復、転写やスプライシングの制御 (Kuroda M et al., 2000 ; Meissner M et al., 2003 ; Wang X et al., 2008) 、細胞質内における翻訳過程やストレス負荷時のStress Granuleの形成 (Sama RR et al., 2013) 等が挙げられ、多岐にわたる (Figure 3B)。
2.3 FUS R495X変異
FUSの家族性ALS変異は現在までに多数報告があり、C末のNLSやその周囲に存在す
ることが知られている (Lagier-Tourenne C and Cleveland DW 2009 ; Figure 4) 。FUSは、
核内輸送体TransportinがFUSのNLSを認識・結合することで、核内へと輸送される。C 末に変異がある場合、Transportinとの結合が弱まり輸送が損なわれるため、細胞質局在が 増加する (Dormann D et al., 2010) 。
家族性変異の一つであるR495X変異は2010年に家族性ALSの遺伝子変異として同定さ れた (Bosco DA et al., 2010) 。495番目のアルギニンをコードするコドンがストップコド ンに変異したミスセンス変異であり、C末端の核移行シグナルを含む32アミノ酸が欠失し ている。他の変異 (R521G) を持つ患者群が平均 40 歳で発症し約 27 ヶ月後に死に至る (Kwiatkowski TJ Jr et al., 2009) のに対して、R495X変異を持つ患者群は、平均35歳に発 症し、約16ヶ月後に死に至る。したがって、R495X変異はALSを発症するFUSの遺伝子 変異の中でもきわめて重篤な変異だと考えられる。また、所属研究室では、N末端にEGFP を付加したR495X変異体をマウス ES細胞由来神経細胞に発現させた場合、神経細胞の細 胞質において局所的に集積し構造体を形成することを観察している。
従って、FUS R495X変異によるALS発症機構として、FUSの本来の核内での機能消失
によるLoss of function説と、細胞質内での構造体形成によるToxic gain of function説の二 つの説が有力であった。しかし、FUS KOゼブラフィッシュやマウスにおいてALS様の症 状を示さないこと等 (Rogelj B et al., 2012; Svetlana Lebedeva et al., 2017) から、FUSの
Loss of functionによる発症の可能性が低いことが示唆されている。従って、FUS R495X変
異によるALS発症には、構造体形成によるGain of toxic functionが寄与すると考えられる。
以上の経緯からFUS R495X変異体によるALSの発症機構として次のような機構が考え られる (Figure 5) 。
①R495X変異による細胞質局在の増加
②細胞質におけるR495X構造体の形成
6
③構造体形成により生じるgain of toxic functionによる神経細胞死の誘引
しかしながら、FUS R495X 変異体がどのように構造体を形成するのか、また構造体形 成により細胞内でどのような変化が引き起こされるのか、は未解明である。
2.4 相転移
真核生物の細胞において観察されるRNA顆粒 (RNA granule) は、主にRNA、RNA結 合タンパク質から構成される膜を持たない構造体である。RNA顆粒として、生殖細胞に見 られるgerm granuleやpolar granule, 体細胞に見られるstress granuleやProcessing-body 等の存在が知られている (Anderson P et al.,2006) 。これらのRNA顆粒はストレスなどの 環境変化に応じ、mRNA の輸送、貯蔵、分解などを行い遺伝子発現を制御すると考えられ ている。しかしながら、RNA顆粒は非常に動的な構造体であり、その形成の仕組み及び内 部の構造等について検証がなされていなかった。RNA 顆粒の構成タンパク質である RNA 結合タンパク質の多くは RNA結合ドメインと LC (Low-complexity) 配列と呼ばれる配列 をもつ。LC配列はアミノ酸の著しく偏った配列であり、決まった構造をもたずフレキシブ ルな状態を保っている。このLC配列の本来の機能について長年未解明であった。
近年、LC 配列を有するタンパク質は濃度、温度依存的に相転移を引き起こし liquid
droplet と呼ばれる流動性を保った構造体を形成する性質を持つことが相次いで報告され
(Kato M et al., 2012; Molliex A et al., 2015) 、RNA顆粒の形成において重要な役割を担っ ていることが示唆されている。FUSもその他RNA結合タンパク質と同様LC配列を持ち、
濃度、温度依存的に相転移し、liquid dropletを形成する (Murakami T et al., 2015; Patel et al., 2015) 。
これらの経緯から、FUS の構造体の形成に相転移が関与する可能性が示唆されている。
しかし、相転移に関する多くの報告は精製系を用いた実験により得られた結果であり、相転 移と疾患との関連性については明らかでない。
2.5 局所的翻訳機構
神経細胞は長い軸索と複数の樹状突起を持つ、高度に発達した極性細胞であり、神経軸索 と標的細胞の樹状突起間でシナプスを形成し、細胞間の情報伝達を行う。極性の形成や維持 には、各々の部位に必要なタンパク質が局在する必要がある。神経細胞では、ほとんどのタ ンパク質は細胞体で合成されるため、モータータンパク質やカーゴ受容体を介した軸索輸 送に焦点を当て解析が行われてきた (Hirokawa et al ., 2010)。
しかしながら、細胞体と切り離した軸索においてもタンパク質翻訳が行われ、かつ軸索成 長因子を添加した場合に軸索伸長が起こるという発見 (Campbell DS and Holt CE , 2001)
7
から、各々の部位で様々な刺激に応じてタンパク質合成が起こる局所的翻訳機構の存在が 示唆されるようになった。以降の解析により、樹状突起や軸索等で機能するタンパク質をコ ードする一部のmRNAは、mRNAとして細胞内輸送を受け、各々の機能部位に局在し、局 所的に翻訳されることが明らかとなった。
また2015年、Puromycin incorporation法とProximity Ligation Assay (PLA) 法を組み合 わせた手法であるPuro-PLA法が報告され (Susanne tom Dieck et al., 2015) 、着目遺伝子 の翻訳部位を可視化することが可能になった。その結果、現在14遺伝子が局所的に翻訳さ れる遺伝子として同定されている (Holt CE et al., 2019) 。
局所的翻訳を受けるRNAはRNA結合タンパク質とRNA顆粒を形成し、Kinesin等のモ ータータンパク質により輸送される (Kanai Y et al., 2004) 。輸送されるRNAは 3’UTR の 長さ、配列、構造に特徴を持つ (Tushev G et al., 2018) 。これらをRNA結合タンパク質が 認識することでRNA顆粒に取り込まれると考えられている。
ALSの原因因子であるTDP-43やFUSもmRNAの細胞内輸送に関わることが報告され ている (Birsa N et al ., 2019) ことから、細胞質での異常な構造体の形成によりmRNAの 輸送が変化する可能性が示唆されている。実際、FUSの家族性変異の一つであるP525L変 異体を発現させた場合に、構造体を形成した細胞ではDdr2とKif5b mRNAの輸送障害が 起こることが報告されている (Yasuda K et al ., 2017)。
しかしながら、FUSの構造体形成が、その他にどのような遺伝子のmRNA輸送を変化さ せるのか、そして局所的翻訳機構の破綻がALS発症に寄与するかについては未解明である。
本研究の目的
以上の背景から ALS発症機構解明及び新たな創薬ターゲットを探索する上で、現在未解 明であるFUS の構造体形成分子機構及び、構造体形成に伴う細胞内変化について明らかに することは、極めて重要であると考えられる。
そこで私は、所属研究室でのFUS R495X変異体が神経細胞において構造体を形成すると いう発見を基に、FUS R495Xの構造体形成分子機構、およびR495X構造体形成に伴う細胞 内変化を明らかにすることを目的に解析を行った。
8
3.研究結果
FUS R495X
構造体形成機構の解明1. FUS R495Xは神経細胞の細胞質において構造体を形成する。
まず、FUS R495X変異体が神経細胞において構造体を形成することを確認した。N末端 にEGFPを付加したEGFP-FUS WT及びR495X変異体を作製し (Figure 6A,B) 、マウス ES細胞由来神経細胞に発現させ細胞内局在を観察した。その結果、WTは主に核内に局在 していたのに対し、R495X 変異体では細胞質において局所的な集積が観察された (Figure 6C) 。この結果からR495X変異体が神経細胞細胞質において構造体を形成することが確認 できた。
2.R495X構造体形成に必須な領域の同定
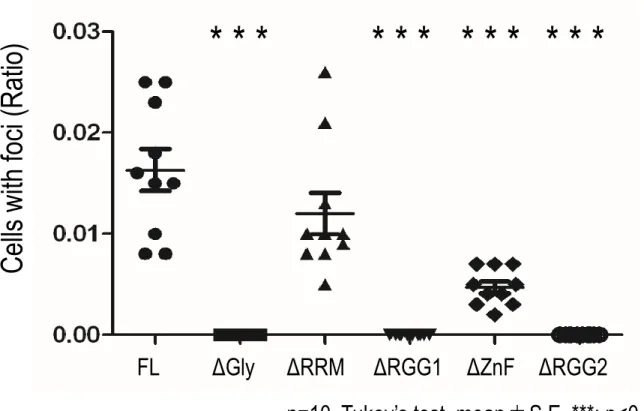
次に、R495X 構造体形成に必須な領域の同定を試みた。そこで、構造体形成に必要なド メインを明らかとするため、N末にEGFPを付加したR495X変異体を基に、各ドメインを 欠損したコンストラクトを作製し (Figure 7A) 、構造体形成能を検証した。各ドメインが 欠損されていることを、ウエスタンブロットを用い確認した (Figure 7B)。その後、各欠損 体をマウスES細胞由来神経細胞にウイルスを用い発現させたのち、細胞内局在を観察した。
その結果、Gly-rich, RGG1 及び RGG2 の欠損体では構造体の形成は観察されなかった
(Figure 7C) 。また、各種欠損体を発現している細胞の内、構造体を形成した細胞の割合を
算出した (Figure 7D) 。その結果、ZnFドメインの欠損体においても構造体形成が減少す ることが分かった。この結果よりGly-rich, RGG1, RGG2各ドメインが構造体形成に必須な ドメインであることが明らかとなった。また、ZnF ドメインも構造体形成に一部寄与する ことが明らかとなった。
3. R495X変異体における各RNA結合ドメインのRNA結合量への寄与
次に、各欠損変異体のRNA結合能について検証を行った。マウスES細胞由来神経細胞 に各欠損変異体を発現させ、UV-crosslink and immunoprecipitation (CLIP) 法を用いFUS- RNA複合体を回収した。その後 RNAをγ32P-ATPを用い放射標識したのち、タンパク質 をウエスタンブロット、放射標識された RNA を X 線フィルムへの感光を行い検出した (Figure 8A) 。各欠損体のRNAとの結合量は、IPサンプルにおけるFUS量に対するX線 フィルムで検出されたintensityを求めることで評価した。その結果、構造体を形成しない
Gly-rich, RGG1, RGG2ドメインの欠損変異体ではRNAとの結合量が約9割近く減少する
9
ことが分かった。また、ZnFドメインの欠損体においてもRNAとの結合量の減少が観察さ れた (Figure 8B, 8C) 。この結果は、各欠損体発現細胞での構造体形成細胞の割合を検証し た結果 (Figure 7D) と高い相関を示したことから、これらRNA結合ドメインのRNAとの 結合が構造体形成に強く寄与すると考えられた。
また、興味深いことにGly-rich, RGG1, RGG2ドメインの欠損変異体全てで、R495X変 異体と比較してRNAとの結合量が9割近く減少していた。この結果から、R495X変異体は
Gly-rich, RGG1, RGG2ドメイン一つごとにRNAを認識しているのではなく、3つのドメ
インが協調して構造的にRNAを認識すると考えられる。
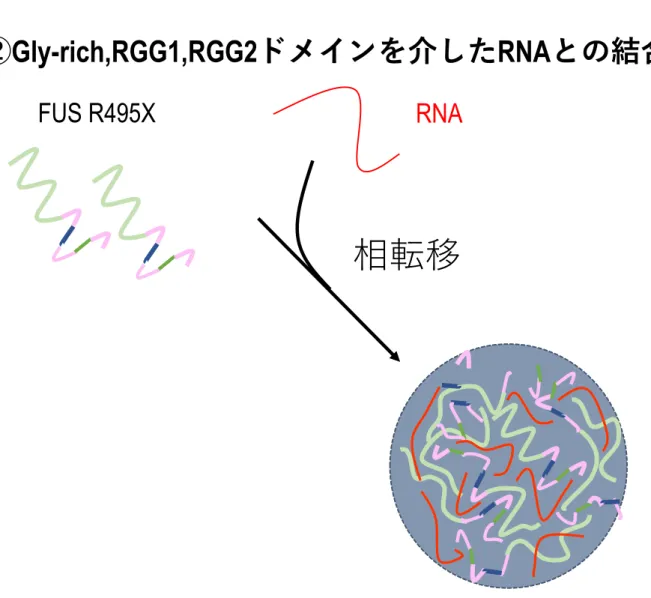
4.FUSの相転移にRNAが寄与する。
これまでの解析から、R495X 構造体の形成には RNA との相互作用が寄与することが示 唆されている。また、RNA 結合タンパク質の相転移にRNA が関与することが相次いで報 告されている (Saha S et al., 2016 ; Maharana S et al., 2018) 。以上の経緯から、RNAがFUS の相転移に寄与することで、R495X構造体の形成に寄与する可能性を考えた。そこでRNA がFUSの相転移に寄与するか検証を行った。
まず、RNAがFUSと共に相転移するのか検証をおこなった。具体的には、GST tagを用 い精製したFUS protein(3.0 μM)をTEV proteaseを用いGST切断したのち、Cy5標識 したRNAを添加し、FUSが形成するLiquid dropletとCy5蛍光を観察した (Figure 9A) 。 用いたRNAは、当研究室の解析によりすでに FUSが結合することが示されている遺伝子 であるCalm2のFUSと結合する配列 (GAA AAU UGA UGA AGU AAC AUG UUG CAU
GUG GCU UAC UCU GGC UAU)の5’末端にCy5を付加し標識したRNAプローブを用い
た。その結果、FUSが形成するliquid droplet 内にCy5の蛍光が観察された(Figure 9B)。
この結果から、RNAはFUS共に相転移すること、そしてLiquid droplet内にRNAが集積 することが分かった。
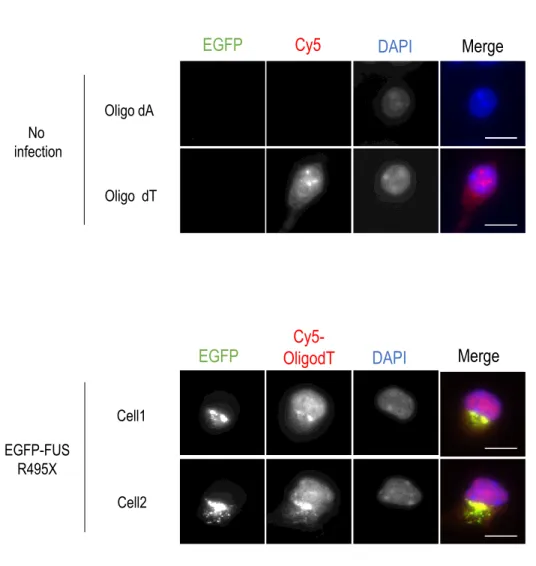
実際に細胞内においてR495X構造体とRNAが共に集積するか検証を行った。EGFP-FUS
R495XをマウスES細胞由来神経細胞に発現させたのち、mRNAの局在をCy5で標識した
Oligo dT probeを用いたFluorescence in situ hybridization (FISH)を行い観察した。その結 果、構造体とCy5-Oligo dTの共局在が観察された (Figure 9C )。この結果から、R495X変 異体はmRNAと共に集積すると考えられる。
次にFUSの相転移にRNAが寄与するか検証をおこなった。GST tagを用い精製したFUS protein(1.0 μM)をTEV proteaseを用いGST切断したのち、マウスES細胞由来神経細 胞から抽出した total RNA (1.5μg) を加え、構造体が形成された場合に、遠心した際に
Pellet画分に検出されるFUSの量を比較した。タンパク質の検出は、SDS-PAGE, CBB染
10
色を用いた (Figure 10A) 。その結果、RNAの添加により、Pellet画分に多くのFUSが検 出された (Figure 10B) 。この結果から、FUSの相転移をRNAが促進することが示唆され た。
以上の結果からFUSはRNAと共に相転移し、かつRNAがFUSの相転移を促進するこ とが示唆された。
5.R495X変異によるRNAとの結合量の変化
これまでの結果から、R495X 変異体は RNA との結合を介して構造体を形成することが 示唆されている。そこで、WTとR495X変異体とでRNAとの結合量に変化があるか検証 を行った。具体的にはマウスES細胞由来神経細胞にEGFP-FUS WT およびR495X変異 体を発現させ、Figure 8 と同様の手法を用い RNA 結合量を検証した。その結果、WT と R495X変異体とではRNAとの結合量に有意な差は検出されなかった (Figure 11A, 11B) 。 この結果から、R495Xの構造体形成には、RNAとの結合のみでなく、細胞質局在の増加が 必須であると考えられる。
FUS R495X構造体形成機構 小括
Fig1~24までの結果から、FUSはRNAと相互作用することで相転移が促進される性質を
もち、R495X変異体はGly-rich, RGG1, RGG2ドメインを介してRNAと相互作用するこ とにより相転移が促進された結果、構造体を形成すると考えられる。しかし、WTとR495X 変異体とでは RNA との結合量に変化は観察されたなかったことから、R495X の構造体形 成には①細胞質への局在の増加②細胞質でのRNAとの結合、の2つの要素が必要であるこ とが示唆される。想定された分子機構をFigure 12に示した。
11
R495X
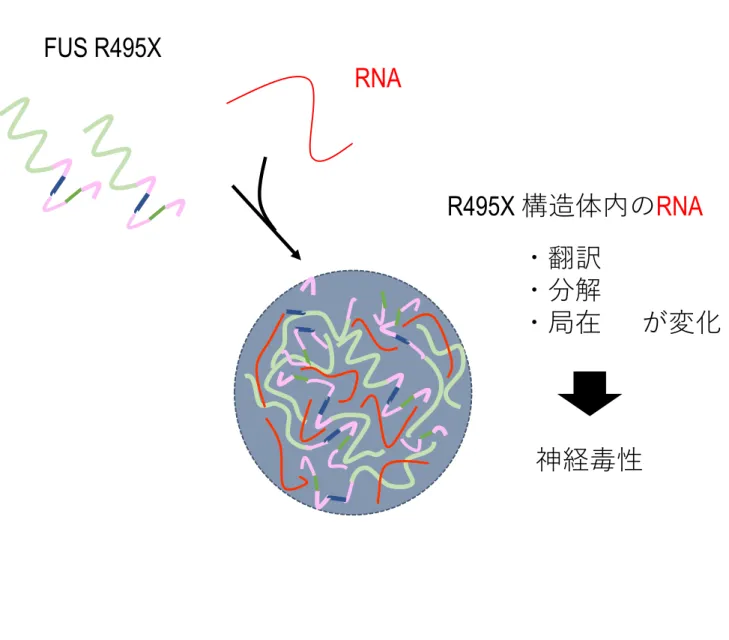
構造体形成に伴う細胞内変化の解明次に、R495X 構造体形成に伴う細胞内変化を明らかにするため検証を行った。これまで の解析から R495X 構造体内には R495X変異体と RNAが共に集積すると考えられる。そ こで、構造体内に特定のRNA分子が取り込まれることで、取り込まれたRNA分子の局在、
翻訳、分解等を変化させることで神経毒性および神経細胞死を引き起こしている可能性を 考えた (Figure 13) 。この可能性を検証するため、本研究ではFUS分子内のRNA結合ド メイン及びR495X結合RNAに着目し解析を行った
各RNA結合ドメインに着目した解析
1.神経細胞死にRRMドメインが寄与する。
まず、各RNA結合ドメインに着目し細胞死への寄与について検証した。マウスES細胞 由来神経細胞に各RNA結合ドメインの欠損EGFP-R495X変異体を発現させ、24時間おき に同一細胞のEGFPの蛍光観察を行った。その結果、構造体を形成しないGly-rich, RGG1, RGG2の欠損体を発現させた細胞では蛍光の消失は観察されず、構造体を形成するZnF の 欠損体を発現させた細胞では蛍光の消失が観察された。しかし、予想に反して、RRMの欠 損体では構造体を形成するにも関わらず、蛍光の消失は観察されなかった (Figure 14) 。こ の結果より構造体が誘引する神経細胞死に RRM ドメインが関与していることが示唆され た。
2.構造体が誘引する神経細胞死にはRRMドメインのRNAとの結合が関与する。
RRMドメインは、ドメイン内の305, 341, 359, 368番目のフェニルアラニンがRNAとの 結合に重要であることが知られている (Daigle JG et al.,2013) 。そこで、RRMドメインの RNAとの結合が神経細胞死に関与しているか検証するため、4 つのフェニルアラニンをロ イシンに置換したEGFP-FUS R495X 4FL変異体を作製し (Figure 15A) 、マウスES細胞 由来神経細胞に発現させ、構造体形成能及び神経細胞死への寄与について検証した。
まず、EGFP-FUS R495X 4FLの構造体形成能について検証した。マウスES細胞由来神
経細胞にEGFP-FUS R495X 4FLを発現させ、EGFPの蛍光を観察した。その結果、RRM
ドメインの欠損体同様、EGFP蛍光の局所的な集積が観察され、細胞質において構造体の形 成が観察された (Figure 15B) 。
次に、4FL変異による細胞死への寄与について検証した。EGFP-FUS R495X 4FLを神経 細胞に発現させ24時間おきに同一細胞の観察を行った。その結果RRMの欠損体同様、発 現誘導後120時間以内に蛍光の消失は観察されなかった (Figure 15C) 。
12
この結果より構造体が誘引する神経細胞死にはRRMドメインのRNAとの結合が関与し ていることが示唆された。
R495X変異体結合RNAに着目した解析
次に、R495X 変異体の結合 RNA に着目し解析を行った。当研究室ではすでに FUS R495X変異体が結合するRNAについてCLIP-RNAseq法 (Figure 16A) を用いて網羅的に 明らかにしている (Nakaya T and Maragkakis M , 2018) 。そこで、この網羅的解析データ
を用いR495X構造体が引き起こす細胞内変化について解明を試みた。
3.FUS R495Xは主にCDS, 3’UTR 領域に結合する。
まず、R495X 変異体が遺伝子のどの領域に結合するのか検証するため、R495XCLIP tag の遺伝子領域ごとの分布ついて検証した。具体的には遺伝子の各領域5’UTR (untranslated region) , CDS (coding sequence) , 3’UTR, Intron の1kbあたりのR495X CLIP tagの数を 算出し比較した。その結果、R495X変異体は最も 3’UTR に結合し、次いでCDSの遺伝子 領域に結合することが分かった (Figure 16B) 。
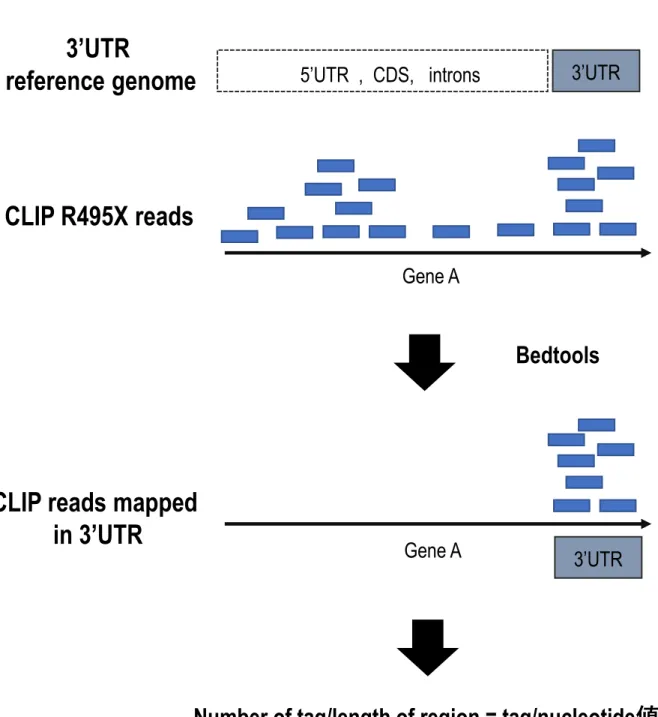
4.各領域のCLIP tagの抽出,
次に、R495X変異体がどのような遺伝子の3‘UTRもしくはCDSに結合するか明らかに するため、各領域にマッピングされるR495X CLIP readの抽出を行った。具体的には、各 領域ごとのリファレンスゲノムを作製し、その領域にマッピングされたreadをBEDTools (Quinlan AR and Hall IM. 2010) を用いることで抽出した (Figure 17A) 。各遺伝子領域の みを抽出できていることをUCSCのGenome browserにて確認した (Figure 17B)。その後、
得られた各領域ごとのreadをカウントし、Number of tag/length of region = tag/nucleotide 値を算出した。
5.各領域ごとの結合RNAのGene enrichment解析
次に、各領域ごとのR495X結合RNAのGene Ontology enrichment (GO) 解析をおこな った。具体的には 3’UTR、CDSごとにtag/nucleotide値のTop50の遺伝子を抽出し (Figure 18A, 18B) 、Gene Ontology enrichment解析を行った。
その結果、R495X変異体は細胞の形態形成及び軸索形成に関わる遺伝子群の3’UTR に結 合することが分かり、リボソームの構成に関わる遺伝子群のCDS領域に結合することが分 かった (Figure 18C) 。
13
6.各領域ごとのR495X変異体の結合モチーフ
次に、各領域ごとのR495X変異体の結合モチーフについて検証を行った。各領域におけ
る R495X 変異体の結合モチーフは Figure 19A, 19B のようになった。興味深いことに、
3’UTR での結合モチーフであるUGUGUGUGモチーフは、FUS同様ALSの原因因子であ
る TDP-43 (Figure 20) の結合モチーフであることが知られている (Tollervey JR et al.,
2011)。さらに、マウス脳を用いた結合RNA網羅的解析により、FUSとTDP-43は3’UTR
で特に共通した結合領域を持つことが示されている(Lagier-Tourenne C et al ., 2012)。以上 から、FUSとTDP-43は3’UTR を介した共通の機構によりALS発症に寄与している可能 性が考えられた。そこで、3’UTR に着目し解析を行うこととした。
7.R495X変異体は局所的に翻訳される遺伝子の3’UTR に強く結合する。
これまでの解析から、R495X 変異体は細胞の形態形成及び軸索形成に関わる遺伝子群の 3’UTR に結合することが示唆されている (Figure 18D) 。
近年、樹状突起や軸索等で機能するタンパク質をコードする一部のmRNAは、mRNAと して細胞内輸送を受け、各々の機能部位に局在し、局所的に翻訳されることが報告されてい る。局所的翻訳を受けるRNAは 3’UTR の長さや配列、構造に特徴を持ち、これらをRNA 結合タンパク質が認識することでRNA顆粒に取り込まれ輸送される (Figure 21A)。
そこで、R495X変異体が3’UTRを介して結合する遺伝子群である、細胞の形態形成及び 軸索形成に関わる遺伝子群について着目すると、軸索および軸索末端で局所的翻訳される 遺伝子であるActb, Mapt (Holt CE et al., 2019) の2遺伝子が含まれていた (Figure 21B) 。 現在、局所的翻訳が示された遺伝子が14遺伝子のみであることから考えると極めて高い割 合だと考えられる。また、APP, GAP43のmRNAは神経細胞において細胞内輸送を受ける ことが知られており(Rivera D et al .,2015 ; Fallini C et al ., 2016) 、局所的翻訳を受ける 可能性が考えられる。
さらに、その他局所翻訳されることが報告されている12遺伝子において3’UTR とCDS での結合量 (tag/nucleotide値) 比を底2で対数を取った値を算出した。この値は、値が0
の場合、3’UTR とCDSに均等に結合することを指す。その結果、Figure 21C の様になり、
Rapgef以外の11 遺伝子は正の値をとることが分かった。従って、R495X 変異体はこれら
局所的翻訳される遺伝子の3’UTR を認識しやすい性質をもつことが分かった。
以上の知見から、R495X変異体は局所的翻訳を受けるRNA分子の特徴的な 3’UTR 領域 を、認識し結合することで、局所的翻訳機構の破綻を引き起こすという新規分子機構の存在 が示唆された (Figure 21C)。また、軸索で機能する遺伝子であるActb, Maptの2遺伝子と
14
特に結合していたことから、軸索での局所的翻訳機構を破綻させている可能性が考えられ た。
8.Actb, Mapt mRNAとの結合にRRMドメインが寄与する。
次に、Actb, Mapt mRNAとの結合に、構造体形成及びRRMドメインが寄与するか明ら
かにするため、RNA-IP法 (Figure 22A) を用い検証を行った。マウスES細胞由来神経細 胞にEGFP-FUS WT, EGFP-FUS R495X、EGFP-FUS R495X ΔGly-rich, EGFP-FUS R495X 4FL の4変異体を発現させ、RNA-IP法を用いFUS-RNA複合体を回収した。その後RNA を抽出しRT-PCRを行うことで、各変異体とActb, Mapt mRNAとの結合量を検証した。
その結果、4FL変異体では、R495X変異体と比較してActb ,Mapt mRNAとの結合量の 減少が観察された。また、ΔGly-rich変異体では、Mapt mRNA のみ結合量の減少が観察さ れた (Figure 22B, 22C) 。これら結果より、R495X変異体はRRMドメインを介して、局 所的翻訳されるRNAであるActb, Maptと結合することが分かった。また、Mapt mRNA との結合にはGly-richドメインも一部寄与することが示唆された。
小括 構造体形成による細胞内変化の解明
Figure 13~22 の結果から、R495X構造体はRRMドメインを介したRNAとの結合を介
して神経細胞死を引き起こすこと、R495X変異体は局所的翻訳されるRNAの3’UTR を認 識しやすい傾向にあること、さらに軸索で局所的に翻訳される RNA 分子である Mapt,
Actb mRNAとの結合にRRMドメインが寄与することが分かった。
以上の結果から、今後さらなる検証が必須であるがR495X構造体はRRMドメインを介 して局所的翻訳機構を破綻させることで神経細胞死を引き起こす、新規分子機構の存在が 示唆された(Figure 23)。
15
考察
本研究はFUS R495X変異によるALS発症機構の解明に向け、R495X変異体の構造体形
成分子機構の解明とR495X構造体形成による細胞内変化について明らかにすることを目的 に解析を行った。
R495X構造体形成分子機構に関しては以下の点を明らかにした。
1) 各RNA結合ドメインの欠損変異体を用いた解析から、Gly-rich,RGG1,RGG2ド メインが構造体形成に必須であることを示した。また、ZnFドメインが構造体形成 に一部寄与することを示した。さらに、CLIP法を用いた解析から、構造体を形成 しない欠損体であるΔGly-rich, ΔRGG1, ΔRGG2ではR495X変異体と比較して RNAとの結合量が約9割減少することを示した。
2) 精製系を用いた解析から FUS は RNA と共に相転移し、さらに FUS の相転移を RNAが促進することを示した。この結果より、R495X変異体はRNAとの相互作 用を介して相転移が促進された結果、構造体を形成することが示唆された。
以上の結果から、FUSはRNAと相互作用することで相転移する性質をもち、R495X変
異体はGly-rich, RGG1, RGG2ドメインを介してRNAと相互作用することにより相転
移が促進された結果、構造体を形成すると考えられる。
本研究の解析から、FUS R495X変異体の構造体形成にRNAとの結合が寄与することを 示した。しかしFUS WTとR495X変異体とではRNAとの結合量には有意な変化は観察さ れなかった(Figure 11)。この結果から、R495Xの構造体形成には、①変異による細胞質の 局在増加、そして②細胞質におけるRNAとの相互作用、の2点が重要であることが示唆さ れる。細胞質への局在増加がR49X構造体形成に寄与する機構として、いくつか可能性が考 えられる。
まず、結合するRNAの違いにより相転移への影響が異なる可能性が考えられる。これま で所属研究室では、WTは核内にて主にpre-mRNAのイントロン領域に結合し、R495X変 異体は細胞質にてmRNAの3’UTR およびCDSと結合することを示している (Nakaya T and Maragkakis M. 2018)。これら結合RNAの違いによりFUSの相転移に与える影響が 異なる可能性が考えられる。実際、Stressを受けた際に形成されるStress granuleにおいて、
特異的に排除されるRNA分子が存在することも報告されており (Fay MM. and Anderson
PJ.2018)、RNA の構造、配列によって相転移に与える影響が異なる可能性は十分に考えら
16
れる。今後、細胞分画法等を用い核、細胞質内ごとのRNAを抽出しFUS の相転移への影 響について検証することで、明らかにすることができると予想される。
また、その他の可能性として、核内と細胞質において FUSのRNAとの結合様式が異な る可能性も考えられる。CLIP法を用いた結合RNA量解析の結果、興味深いことにGly-rich,
RGG1, RGG2ドメインの欠損変異体全てで、R495X変異体と比較して RNAとの結合量
が9割近く減少していた。この結果から、R495X変異体はGly, RGG1, RGG2ドメイン一 つごとにRNAを認識しているのではなく、3つのドメイン全てを介して構造的にRNAを 認識すると考えられる。現時点ではこのRNAの結合様式が構造体形成にどのように寄与す るのかは未知である。しかし、今後FUS 全長を基にした各RNA結合ドメインの欠損変異 体を用いた同様の解析を行い、核内と細胞質とでRNAとの結合様式が大きく異なっていた 場合、RNAの結合様式の違いが構造体形成に寄与する可能性も考えられる。
また、RNA以外の因子により相転移が制御されている可能性も考えられる。相転移の制 御に関する報告は近年盛んに研究が行われており、様々な因子の関与が示唆されている。例 を挙げると、本研究で着目したRNA以外にもATPや塩濃度、温度等が寄与することが報 告されており (Kang J et al., 2018 )、これら因子の差異により細胞質においては環境的に相 転移が起こりやすい場が構築されている可能性も考えられる。
また、反対に核内においては何等かの因子が相転移を抑制している可能性も考えられる。
ALSの原因因子であるTDP-43, FUS , hnRNPA等のRNA結合タンパク質は、共通して天 然変性領域をもち、相転移する性質をもつ(Franzmann TM and Alberti S. 2019)。これら分 子は、主に核内に存在するため、核内ではそれらタンパク質が密集して存在すると予想され る。相転移は天然変性領域を持った分子が局所的に高密度に存在することで引き起こされ ることから (Patel A et al., 2015) 、核内は極めて相転移が起こりやすい場が形成されてい ると考えられる。しかし、DNA損傷時等を除き、これらタンパク質の構造体は通常観察さ れない。従って、核内においては何等かの因子が抑制的に相転移を制御する可能性が考えら れる。実際、核内輸送受容体Kapβ2が相転移を抑制することが報告されており (Guo L et
al ., 2018)、このような分子の存在により核内においては不必要な相転移を抑制する機構が
存在する可能性も考えられる。
17
R495X構造体形成による細胞内変化については以下を明らかにした。
R495X構造体内にはR495X変異体とRNAが共に集積すると考えられることから、集積
したmRNAの翻訳、分解、局在等に異常が起こる可能性を考えた。そこで本研究ではFUS 分子内のRNA結合ドメイン及びR495X結合RNAに着目し解析を行った。
1) 各 RNA 結合ドメインの欠損変異体および 4FL 変異体を用いた解析から、構造体
を形成しない Gly-rich, RGG1, RGG2 の欠損変異体は神経細胞死を誘引しないこ と、RRMの欠損変異体及び4FL変異体は形成するにも関わらず、神経細胞死を誘 引しないことを示した。この結果より、R495X 構造体は RRM ドメインを介した RNAとの結合を介して神経細胞死を引き起こすことが示唆された。
2) R495X 変異体が結合する RNAについて CLIP-RNAseq法を用い得られた網羅的
データを用い解析を行った。その結果、R495X 変異体は、細胞の形態形成及び軸 索形成に関わる遺伝子群の3’UTR に結合することが分かり、リボソームの構成に 関わる遺伝子群のCDSに結合することが分かった。その後、3’UTR に着目した解 析から、神経細胞の軸索や樹状突起内で局所的に翻訳される遺伝子の3’UTR を好 んで結合することが分かった。さらに、RNA-IP法を用い局所的翻訳される遺伝子
であるActb, Maptとの結合にRRMドメインが寄与することを明らかにした。以
上の結果から、R495X変異体は軸索でRRMドメインを介してActb, Mapt mRNA と結合することで、軸索における局所的翻訳機構の破綻を引き起こす可能性が考 えられる。
各 RNA 結合ドメインの欠損変異体および4FL 変異体を発現させた細胞の経時的な観察 結果から、R495X構造体はRRMドメインを介したRNAとの結合を介して神経細胞死を引 き起こすことが示唆されている。また、構造体を形成しない欠損変異体 (ΔGly-rich,Δ
RGG1,ΔRGG2) を発現させた細胞では、RRMドメインは存在するにも関わらず神経細胞
死が誘引されなかった。この結果から、R495X変異により神経細胞死を誘引する機構には、
構造体形成と RRM ドメインを介した RNA との結合の 2 点が必要であることが示唆され た。精製系を用いた解析からFUSはRNAと共に相転移することが示唆されていることか
ら、R495X変異体がRRMドメインを介して結合するRNAと共に集積し構造体内に捕捉す
ることで神経細胞死を引き起こすと推測される。
CLIP法を用いた解析から、RRMドメインの欠損変異体において、R495X変異体と比較 して RNA との結合量に有意な減少は観察されなかった (Figure7) ことから、神経細胞死 に関わるRNAは極めて限局的であると予測される。しかし、RNA-IP法を用いた解析の結 果、軸索において局所的翻訳を受ける代表的な遺伝子である Mapt と Actb との結合には
18
RRMドメインが強く寄与することが示唆された (Figure 22)。この結果から、RRMドメイ ンは局所的翻訳されるRNAに特徴的な3’UTR を認識しやすい性質をもつ可能性が考えら れる。今後、他の遺伝子についても検証を行いたいと考えている。
局所的翻訳機構がALS発症に寄与するかは現時点では不明である。しかし、ALS病態で の主要な変性部位である脊髄の運動神経は極めて長い神経細胞であることが知られている。
そのため、細胞体から遠く離れた軸索中および末端部での局所的翻訳機構が担う役割は他 の細胞と比較して大きいと予想される。よって、局所的翻訳機構の破綻が神経細胞死、ALS 発症に寄与する可能性は考えられるが、今後より詳細に解析を行う必要がある。
また、現在報告されている局所的に翻訳される遺伝子は14種類である。軸索や樹状突起 に存在するRNAは、細胞分画法とRNA-seq法を用いた解析により、2550遺伝子も同定さ れている (I.J. Cajigas et al ., 2012) 。この数から考えると、現在までに同定された14遺伝 子という数字はあまりにも少ない。
本研究の結果、R495X変異体は局所的翻訳されるRNAの3’UTR を好んで結合すること が示唆された。この結果から、R495X変異体の3’UTR への結合量が多い遺伝子群の中には、
現時点では未同定の局所的翻訳される遺伝子が含まれている可能性が示唆される。
今後Puro-PLA法等を用いR495X変異体が3’UTR を好んで結合する遺伝子について順
に検証を行うことで、新規局所的翻訳遺伝子の発見、そして局所的翻訳機構の破綻による ALS発症機構のより詳細な機構を明らかできると期待できる。
R495X変異体の 3’UTR における結合モチーフであるUGUGUGUGモチーフは、TDP-
43の結合モチーフであることが知られている(Tollervey JR et al., 2011)。TDP-43はALS 患者のうち約 9 割の患者で凝集体が観察され、最大の原因因子であると考えられている。
TDP-43とFUSの構成ドメインは類似しているが、RNAとの結合様式はFUSとは異なる
と考えられる。CLIP法の結果より、FUSはGly-rich, RGG1, RGG2ドメインを介して大部 分のRNAと結合していたのに対して、TDP-43 は主としてRRM1 とRRM2 の2 領域が RNAとの結合に寄与すると考えられている(Buratti E et al., 2001)。しかし、本解析の結果、
興味深いことに3’UTR領域おいてはR495X変異体の結合モチーフはTDP-43のモチーフ
と同様のUGUGUGUGモチーフが検出された。この結果から、FUSとTDP-43は3’UTR
領域においては共通の RNA を認識する可能性が考えられる。従って、本研究での 3’UTR に着目した解析により示唆された、局所的翻訳機構の異常を介したALS発症機構は、FUS
とTDP-43 に共通したより普遍的な ALS発症機構の解明につながる可能性が考えられる。
19
5.実験手法
Plasmid
以下のplasmidは当研究室保有のものを使用した。
EGFP/pSLIK EGFP-FUS /pSLIK EGFP-FUS R495X/pSLIK
なおplasmidの精製にはFastGene Plasmid mini kit (日本ジェネティクス) を使用した。配 列の確認には BigDye Terminator Cycle Sequencing に基づきサンプル調製を行い、3100 Data Collection (Applied Biosystem) を使用した。
作製した各constructについて以下に手法を示す。
pSLIK vectorの作製
Entry vectorであるpEN vectorに目的配列を組み込み、LR recombination (Gateway ○R
LR Clonase○RⅡEnzyme mix ,Life Technologies) を用いpSLIK vectorに目的配列を導入し た。以下pEN vectorへの目的配列の挿入はMega primer法を用いて行った。
各鋳型plasmidを用いてAccuprimePfx によりPCR反応をおこなった。その後、PCR産 物をアガロースゲル電気泳動によって分離した後、目的産物に相当する断片をゲルから切
り出し、FastGene ゲル/PCR抽出キット (日本ジェネティクス)を用いてDNAを抽出した。
得られた1st PCR productsをmegaprimerとしてもちい、それに対応するprimerを加え、
2回目のPCR 反応をおこなった。1回目と同様にDNA 抽出をおこない、次いで、insert,
vevtorにたいしてそれぞれ制限酵素による切断をおこなった。反応液は各制限酵素1 L、
推奨buffer 4 l、DNA溶液を加え、DDWでtotal 40 lになるように調整した。37℃、1hr
incubate することで制限酵素による切断をおこなった後、先述の通りにアガロースゲル電
気泳動による分離、目的DNAの抽出をおこなった。
ついで、vector 0.5 l、insert 3.0 l、2x ligation mix (ニッポンジーン) を3.5 lを混合 し、16℃、30 min、incubate することで ligation 反応をおこなった。本反応液に 50l の competent E. coli (DH5) を加え、on iceで5 min incubateし、42℃で30秒間incubateし たあと、on iceで2分間incubateした。LB (Geneticin) plateに播種し、37℃で16hr培養 した。Single colonyからplasmidを精製し、配列を確認した。
以下に上記手法で作製した各construct、制限酵素 (5’側/3’側)、 使用した鋳型plasmid、
megaprimerの作製に用いたprimerの塩基配列、 2回目のPCR反応に用いたprimerの塩 基配列を記す
EGFP-FUS R495XΔGly-rich /pEN (Spe1/ Not1)
20 1st PCR (megaprimer)
1. 鋳型plasmid EGFP /pEN
Primer for : cccgggactagtcaccatggtgagca rev : atcgtttgaggccatcttgtacagctc 2. 鋳型plasmid FLAG- FUSΔRGG1/pEN Primer for : gagctgtacaagatggcctcaaacga rev : tacgcggccgctcagaagcctccacggtccc 2nd PCR
Primer for : cccgggactagtcaccatggtgagca rev : tacgcggccgctcagaagcctccacggtccc
EGFP-FUS R495XΔRRM /pEN (Spe1/ Not1) 1st PCR (megaprimer)
1. 鋳型plasmid EGFP /pEN
Primer for : cccgggactagtcaccatggtgagca rev : atcgtttgaggccatcttgtacagctc 2. 鋳型plasmid FLAG- FUSΔRRM/pEN
Primer for : gagctgtacaagatggcctcaaacga rev : tacgcggccgctcagaagcctccacggtccc 2nd PCR
Primer for : cccgggactagtcaccatggtgagca rev : tacgcggccgctcagaagcctccacggtccc
EGFP-FUS R495XΔRGG1 /pEN (Spe1/ Not1) 1st PCR (megaprimer)
1. 鋳型plasmid EGFP /pEN
Primer for : cccgggactagtcaccatggtgagca rev : atcgtttgaggccatcttgtacagctc 2. 鋳型plasmid FLAG- FUSΔRGG1/pEN Primer for : gagctgtacaagatggcctcaaacga rev : tacgcggccgctcagaagcctccacggtccc 2nd PCR
Primer for : cccgggactagtcaccatggtgagca rev : tacgcggccgctcagaagcctccacggtccc
EGFP-FUS R495XΔZnF /pEN (Spe1/ Not1)
21 1st PCR (megaprimer)
1. 鋳型plasmid EGFP /pEN
Primer for : cccgggactagtcaccatggtgagca rev : atcgtttgaggccatcttgtacagctc 2. 鋳型plasmid FLAG- FUSΔZnF/pEN
Primer for : gagctgtacaagatggcctcaaacga rev : tacgcggccgctcagaagcctccacggtccc 2nd PCR
Primer for : cccgggactagtcaccatggtgagca rev : tacgcggccgctcagaagcctccacggtccc
EGFP-FUS R495XΔRGG2 /pEN (Spe1/ Not1) 1st PCR (megaprimer)
1. 鋳型plasmid EGFP /pEN
Primer for : cccgggactagtcaccatggtgagca rev : atcgtttgaggccatcttgtacagctc 2. 鋳型plasmid FLAG- FUSΔRGG3/pEN Primer for : gagctgtacaagatggcctcaaacga rev : tacgcggccgctcagaagcctccacggtccc 2nd PCR
Primer for : cccgggactagtcaccatggtgagca rev : tacgcggccgctcagaagcctccacggtccc
EGFP-FUS R495X 4FL /pEN (Spe1/ Not1) 1st PCR (megaprimer)
1. 鋳型plasmid EGFP /pEN
Primer for : cccgggactagtcaccatggtgagca rev : atcgtttgaggccatcttgtacagctc 2. 鋳型plasmid FLAG- FUS 4FL/pEN
Primer for : gagctgtacaagatggcctcaaacga rev : tacgcggccgctcagaagcctccacggtccc 2nd PCR
Primer for : cccgggactagtcaccatggtgagca rev : tacgcggccgctcagaagcctccacggtccc
22
細胞培養
1.HEK293T細胞
HEK293T細胞は56℃で30分間処理し非動化したFBS (MP) を10% (v/v) 、含むDMEM (Wako) 中で37℃、5% CO2の条件下で培養した。
2.マウスES細胞
マウスES細胞はFeederlessの状態で、ES培地(200 mL MEM, 37.5 mL ES cell tested fetal bovine serum (FBS, SH30070.03,Hyclone) , 2.5 mL of 200 mM L-glutamine (25030- 081,Invitrogen) ,180 µL of diluted 2-mercaptoethanol (diluted 1/100 in PBS , M- 7522 ,Sigma) , and 25 µL of LIF/ESGRO (ESG1107, Chemicon) ) 中で37℃、5% CO2の条 件下で培養した。プレートはGelatinでコートしたものを用いた。
3.神経細胞への分化
1 日目、ES 細胞を ADFNK 培地 (250 mL of Advanced DMEM/F12 (12634-010, Invitrogen) . 250 mL of Neurobasal Medium (21103-049, Invitrogen) , 57 mL of knockout- SR(10828-028, Invitrogen), 5.7 mL of Pen/Strep,5.7 mL of 200 mM L-glutamine, and 400µL of diluted 2-mercaptoethanol (diluted 1/100 in PBS) ) に播種し37℃、5% CO2の条件下で 培養した。2日目、浮遊しているEmbryonic body (EB) を再度ADFNK培地に播種した。
3日目、EBをレチノイン酸含有 (5 µM) ADFNK 培地に播種し3日間培養した。6日目、
培地を神経細胞用 (ADFNB+GDNF) 培地 (24 mL of Advanced DMEM/F12 ,24 mL of Neurobasal medium , 0.5 mL of Pen/Strep , 0.5 mL of L-glutamine, 1 mL of B27 50x supplement (17504-044, Invitrogen) , 0.5 µL of GDNF (5 ng/mL final concentration) ) に 置換し、7日目にEBを分散して、神経細胞を得た。EBの分散は以下にして行った。EBを 回収し、PBSでwash後、0.05% Trypsine/EDTAで6 min incubateし、100 µL FBS を添加 させることで反応を停止した。培地をDNase/L-15に置換後、ピペットを用いて分散させ、
5min incubateし、上清に含まれる分散した細胞を回収し、神経細胞用培地に置換してpoly-
L-lysine/Laminin コートしたプレートに播種した。
Lentivirus
を用いた発現方法1.LentiVirusの作製
6 well dish (Corning) にHEK293T細胞を1.25 cells/wellの濃度で播種した。翌日、作製 したpSLIK plasmid (2.0 µg) 、packaging vectorである psPAX2 (1.5 µg) , pMD2G (0.5 µg) plasmidを 60 µlのOPTI-MEM で希釈した溶液と、6 µlのLipofecamine 2000 (Invitrogen) を 100 µlのOPTI-MEMに希釈し室温で5 min incubateした溶液を混和し、20 min室温で
23
incubateした。これを1.8 mLのDMEM / 10% FBS培地に添加しtransfectionした。翌日 培地を交換し、24時間後、48時間後に培地を回収した。得られた培地をfiltrate後、100,000
×g で90 分遠心し濃縮した。沈殿したウイルスパーティクルを ADFNB 培地で懸濁し、- 80℃で保存した。
LentiVirusを用いた遺伝子導入及び発現誘導
pSLIK vectorを用い作製したLentiVirusの目的配列の発現調節は、Tet-on systemを利用 しており、Doxycyclineの添加により発現が誘導される。播種した神経細胞にLentiVirusを 加え、翌日 4 µg/mL のDoxycyclineを含む神経細胞用培地に置換して発現誘導した。この 置換を24時間おきに行い、目的タンパク質を発現させた神経細胞を得た。
各種実験手法
Western blotting (WB)
5x Tris-Glycine Sample buffer (final conc. 1x) を加え 熱処理 (100℃、5 min) したサン プ ル を Tris-Glycine gel に ア プ ラ イ し 、 電 気 泳 動 に よ り タ ン パ ク 質 を 分 離 し 、Pure Nitrocellulose Blotting Membrane (Pall corporation) へ転写した。Membrane を5% skim milk / TBS-Tで1 hr, R.T. でincubateすることでblockingをおこなった。TBS-Tで洗浄 後、各種一次抗体と一晩、4℃でincubateした。MembraneをTBS-Tで洗浄後、各種二次 抗体と 1 hr, R.T.で incubate した。TBS-T で洗浄後、ECLTM Western blotting detection regent (GE Healthcare) またはClarityTM Western ECL Substrate (Bio-Rad) を用いて化 学発光させ、LAS-4000 mini (富士フィルム株式会社) または Hyperfilm (GE Healthcare) で検出をおこなった。定量はLAS-4000 miniで取得した画像にたいしmulti gage (富士フィ ルム株式会社) によっておこなった。
WBに用いた一次抗体とその希釈率
rabbit anti-GFP polyclonal antibody (MBL) : 希釈率 1/5000
mouse anti-Tubulinβ-Ⅲ (TuJ1) monoclonal antibody (R&D systems) :希釈率1/500 WBに用いた二次抗体とその希釈率
anti-mouse IgG HRP-linked species-specific whole antibody (GE Healthcare) : 希釈率 1/5000
anti-rabbit IgG HRP-linked species-specific whole antibody (GE Healthcare) : 希釈率 1/5000