制御機構の解明
著者 五十嵐 信智
雑誌名 星薬科大学紀要
号 59
ページ 13‑21
発行年 2017‑12‑10
URL http://id.nii.ac.jp/1240/00000823/
1
近年、 高齢化に伴う腸管機能の低下などに伴い、 便秘 症の患者が増加傾向にある。 また、 麻薬性鎮痛薬である モルヒネを服用している患者ではほぼ100%に便秘が認 められるなど、 薬剤性便秘症も深刻な問題となっている。
現在、 このような便秘症患者に対しては、 第一選択薬と して浸透圧性下剤 (硫酸マグネシウム; MgSO4や酸化 マグネシウム) が処方され、 効果が認められない場合に は大腸刺激性下剤 (ビサコジルやセンノシドA) など、
作用機序が異なる瀉下剤が併用される。 しかしながら、
その治療効果は十分なものとはいえず、 この現状を打破 しようと、 製薬企業では様々な作用機序を呈した瀉下剤 が開発されるに至っている。 このように、 瀉下剤に関す る研究開発が活発に行われているものの、 生体内での便 の濃縮機構については不明な点が多く残っていた。
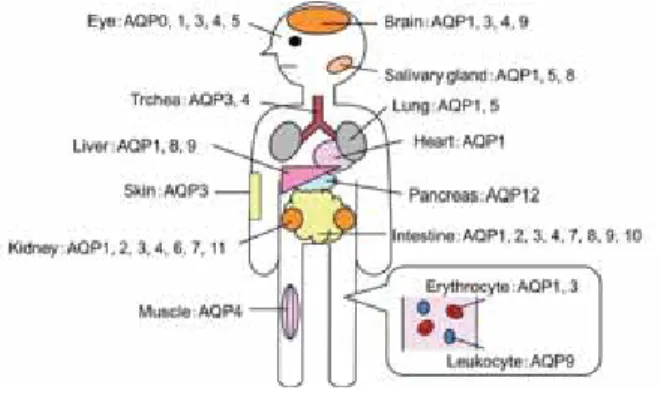
このような中、 近年、 体内での水輸送にアクアポリン (AQP) と呼ばれる水チャネルが重要な役割を担ってい ることが明らかとなってきた。 ヒトにおいては現在、
AQP0
からAQP12
までの13種類のAQP
が同定されて おり、 様々な臓器に発現・分布していることが見出され ている (Fig. 1)1)。 また、 最近では、AQP
の発現量や 活性の変化が種々の疾患の発症に関与していることも明 らかとなってきており、 その機能の重要性がクローズアッ プされているとともに、AQP
をターゲットとした疾患 治療薬の開発を目指した基礎研究も積極的に行われてい る2-6)。腸管には多数の
AQP
ファミリーの発現が認められて おり、 少なくともAQP1、 AQP2、 AQP3、 AQP4、 AQP7、
AQP8、 AQP9、 AQP10
の8
種類の存在が知られてい る (Fig. 1)7-10)。 便の水分量を最終的にコントロールし ている大腸では、 主にAQP1、 AQP2、 AQP3、 AQP4
および
AQP8
が発現している7, 10-12)。 特に、AQP3
はヒ トの大腸粘膜上皮細胞に強く発現しており12)、 便の濃縮 に重要であると考えられている。 しかしながら、 これま で大腸AQP3
の生理的役割やその発現制御機構の詳細 についてはほとんどわかっていなかった。 著者は、 上述 した点について明らかになれば、 便秘や下痢の新しい治 療法や予防法が見出されるのではないかと考え、 基礎研 究を行った。 本稿では、 便の水分量をドラスティックに 変化させる瀉下剤 (MgSO4およびビサコジル) をツー ルとして用いて見出した大腸AQP3
の機能解析の成果 を紹介する。2 MgSO
4AQP3
132-1. MgSO
4便秘症の第一選択に用いられる浸透圧性下剤は、 消化 管ではほとんど吸収されず、 腸管内の浸透圧を上昇させ るため、 水を血管側から管腔側へと移動させ、 瀉下作用 を示すと考えられている14)。 そこで、 ラットに
MgSO
4 を投与し、 大腸における浸透圧調節関連遺伝子 (so-dium myo-inositol transporter; SMIT) の mRNA
発3 !"#$%&'()!*#+
五十嵐 信 智
星薬科大学 薬動学教室
Elucidation of the function and the regulatory mechanism of aquaporin-3 in the colon
Nobutomo IKARASHI
Department of Clinical Pharmacokinetics, Hoshi University
Fig. 1 AQP
現量を指標15, 16) とし、
MgSO
4の瀉下作用と大腸内浸透 圧との関係について調べた。 その結果、 糞中水分量はMgSO
4投与2
時間目から有意に増加し、 投与4
時間後 から8
時間後にかけて、 重度の下痢が発生することが わかった (Fig. 2A)。 これに対して、 大腸内浸透圧は2
時間後にはピークに達し、 この浸透圧の変動パターンと 下痢発生のパターンが異なることが明らかとなった (Fig. 2B)。これらのことより、
MgSO4
による瀉下作用は、 従来 から考えられている腸管内浸透圧の上昇のみでは説明で きないことがわかった。2-2. MgSO
4AQP3
ラットの大腸粘膜上皮細胞には、 ヒトと同様に、
AQP3
が優位に発現していた (Fig. 3)。 そこで、MgSO
4投与後のラット大腸における
AQP3
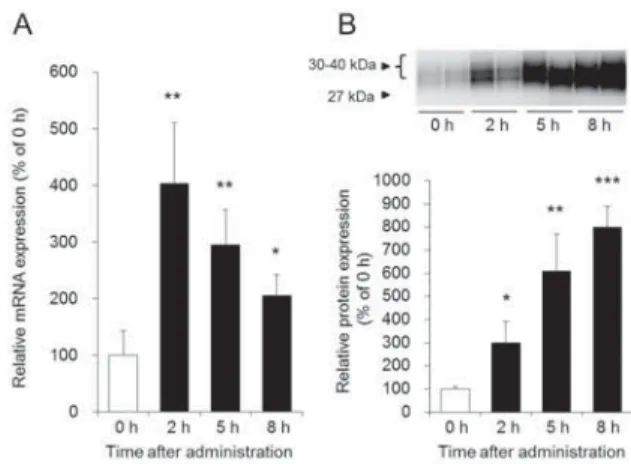
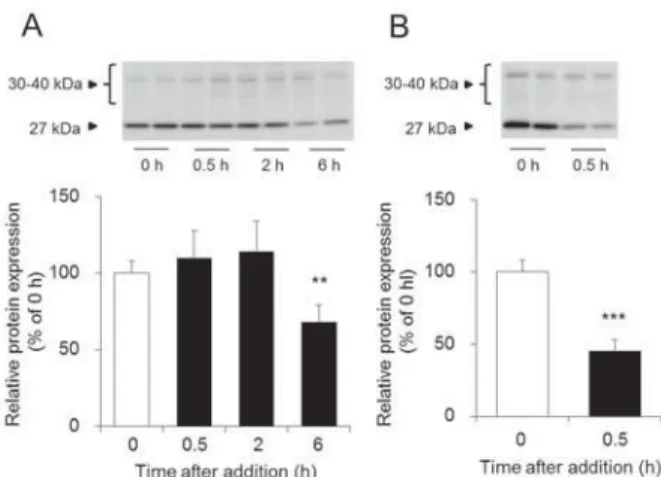
の発現量を解析し、瀉下作用との関係について調べた。 その結果、
AQP3
のmRNA
発現量は、MgSO
4投与2
時間後に最も高値を示 し、 その後、 徐々に減少することがわかった (Fig. 4A)。一方、 タンパク質発現量は
MgSO
4投与2
時間目以降か ら経時的かつ著明に増加した (Fig. 4B)。 また、 このAQP3
タンパク質の発現パターンは、 糞中水分量の経時 変化と符合していた (Fig. 2A and 4B)。以上の結果から、
MgSO
4による瀉下作用は、 次に示 すように生じているものと考えられた。 生理的条件下では、 水は浸透圧が低い管腔側から血管側に
AQP3
を介 して輸送される。MgSO
4投与時には、 浸透圧が逆転す るため、 水は血管側から管腔側に移動する。 この際、MgSO
4投与2
時間後の時点では、AQP3
の発現量が十 分ではなかったため、 水の移動量はそれほど多くなかっ た。 それに対して、 投与2
時間目以降ではAQP3
が著 明に増加したため、 大量の水が管腔側に移動し、 下痢が 発症したものと考えられた (Fig. 5)。 このように、MgSO
4による瀉下作用は、 従来から考えられている腸 管内浸透圧の上昇のみによってもたらされるものではな く、 大腸粘膜上皮細胞のAQP3
の発現増加を伴って、極めて合理的に生じている可能性が示唆された。
3 MgSO
4AQP3
3-1. AQP3 !"
一般に、
AQP
は高浸透圧刺激によって発現が増加す ることが知られている18, 19)。 一方、MgSO
4は水溶液中 でMg
2+とSO
42-に解離し、 浸透圧を上昇させる。 そこ で、
MgSO
4によるAQP3
の発現増加が浸透圧の増加に よって引き起こされたものであるかどうかをヒト結腸癌 由来細胞株HT-29
細胞を用いて調べた。0.75 mM
のNaCl
(282 mOsm) およびMgSO
4 (280 mOsm) 添加Fig. 2 SMIT
MgSO
4MgSO
4! A "#$
SMIT mRNA %& ! B "#'( 13 )*+,
Fig. 3 AQP3 -
./ AQP3 0/1#'( 13 )*+,
Fig. 4 AQP3 MgSO
4MgSO
4AQP3 mRNA
! A "23456! B "%& #'( 13 )*+,
Fig. 5 MgSO
4789: AQP3 ;<
時 の
AQP3
のmRNA
発 現 量 は 、 い ず れ もControl
(279 mOsm) との間に有意な差は認められなかった。同様に、
7.5 mM
のNaCl
添加時のAQP3
の発現量も、Control
と比較して有意な差は認められなかった。 これ に対して、7.5 mM
のMgSO
4添加時においては、 浸透 圧は7.5 mM
のNaCl
と同等であったにもかかわらず、AQP3
の発現量はControl
と比べて約2
倍有意に高い 値を示した (Table 1)。以上の結果から、
MgSO
4によるAQP3
の発現増加は、浸透圧の上昇に起因したものではないことが明らかとなった。
3-2. AQP3
MgSO
4によるAQP3
発現増加作用が、Mg
2+に起因す るものであるのか、 それともSO
42-に起因するものであ るのかを種々のマグネシウム塩および硫酸塩を用いて検 討した。
Mg
2+濃度の等しいMgSO
4とMgCl
2をHT-29 細 胞 に 添 加 し た と こ ろ 、AQP3
の 発 現 量 は い ず れ もControl
に比べて有意に増加し、 この増加率は両者でほ ぼ等しかった (Fig. 6A)。 これに対して、SO
42-の濃度
が等しい
MgSO
4、Na
2SO
4およびK
2SO
4添加実験では、MgSO
4 添 加 時 の み にAQP3
の 発 現 増 加 が 見 ら れ た (Fig. 6B)。これらのことから、
MgSO
4によるAQP3
の発現増加 には、SO
42-は関与せず、
Mg
2+が重要な役割を担ってい ることが示唆された。3-3. MgSO
4AQP3
Mg
2+は大腸粘膜上皮細胞の刷子縁膜に存在するMg
輸送トランスポーターtransient receptor potential melastatin
(TRPM)6/TRPM7
複合体により、 細胞内 に取り込まれる20)。 細胞内に取り込まれたMgは、 アデ ニル酸シクラーゼを活性化する21-23)。 また、 アデニル酸 シクラーゼの活性化により産生されるcAMP
は、 プロ テ イ ン キ ナ ー ゼA
を 活 性 化 し 、 転 写 因 子cAMP- response element-binding protein
(CREB) のリン酸 化を亢進する24, 25)。 一方、AQP
はCREB
のリン酸化に より転写および発現量が調節されている26, 27)。 そこで、MgSO
4によるAQP3
発現増加メカニズムについて、 こ れ ら の シ グ ナ ル 経 路 を 調 べ た 。 そ の 結 果 、MgSO
47.5 mM
添加10分後にはすでに、 細胞内Mg
濃度が約1.4倍有意に増加していることがわかった (Fig. 7A)。
また、
MgSO
4添加20分後におけるアデニル酸シクラー ゼ活性およびプロテインキナーゼA活性は、 それぞれ約1.5倍 お よ び1.6倍 有 意 に 増 加 し 、 添 加 1
時 間 後 に はCREB
のリン酸化も亢進していることが確認できた (Fig. 7B-7D)。以上の結果から、
MgSO
4は細胞内Mg
濃度を増加さ せることにより、 アデニル酸シクラーゼおよびプロテイ ンキナーゼAを活性化し、CREB
のリン酸化の亢進を介 して、AQP3
の発現量を増加させたものと考えられた。4 !"#$%&'( AQP3 )*
28, 29+4-1. AQP3 !
ビサコジルは大腸刺激性の瀉下剤に分類され、 単独あ るいは浸透圧性下剤との併用により、 便秘症の改善に広 く用いられている。 ビサコジルは、 大腸の
prostaglan- din E
2 (PGE2) の産生を促進し、Na
+, K
+-ATPaseの活
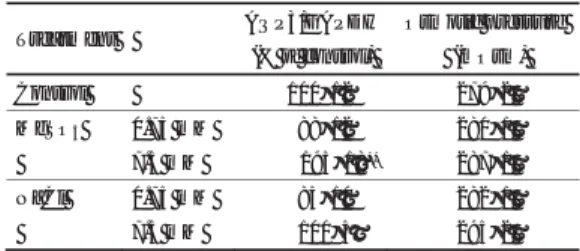
性を阻害する。 その結果、 腸管内浸透圧が高まり、 腸管Table 1 AQP3
ߩ⊒ߦ߷ߔᶐㅘߩᓇ㗀Treatment AQP3/GAPDH
(% of control)
Osmotic pressure (mOsm) Control 100±12㩷 㩷 279±2 MgSO
40.75 mM 88±12㩷 㩷 280±1 7.5 mM 195±18
**287±1 NaCl 0.75 mM 85±10㩷 㩷 282±1 7.5 mM 100±5㩷 㩷 295±2
Table 1 AQP3
HT-29 MgSO
4NaCl
AQP3 mRNA 17
Fig. 6 AQP3
A HT-29 MgSO
4MgCl
2!"#$ AQP3 mRNA % &'#() B HT-29 MgSO
4$ Na
2SO
4K
2SO
4!"#$ AQP3 mRNA % &'#()*+ 17 ,-./
Fig. 7 0 Mg 12$3456789:;<$=>?
@ABC9: A ;< CREB DAE MgSO
4HT-29 MgSO
4!"#$0 Mg 12F A G$34
56789:;<F B G$=>?@ABC9: A ;<F C G
HI#()J($ CREB DAEF P-CREB GK CREB
LAM7N% &'#$OPF P-CREB/CREB G
QR#(F D G)*+ 17 ,-./
側から血管側への水の吸収を減弱させるため、 瀉下作用 を発現すると考えられている30, 31)。 しかしながら、 大腸 における水の移動がどのように行われているのかは全く 不明であった。 そこで、 ビサコジルの瀉下作用における 大腸
AQP3
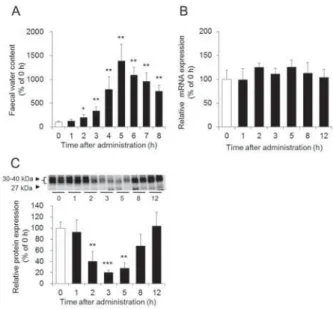
の役割を調べた。 ビサコジルをラットに経 口投与した結果、 ビサコジルはMgSO
4投与時とは異な り、 大腸内浸透圧を変化させずに、 投与4
時間後から 激しい下痢を引き起こすことが確認できた (Fig. 8Aand 8B)。 一方、 大腸の AQP3
のタンパク質発現量は 投与2
時間後から著明に低下し、 この低下と下痢発生 の経時変化が相関していた (Fig. 8C)。以上の結果から、 ビサコジルは大腸の
AQP3
を低下 させることにより、 管腔側から血管側への水の移動を抑 制し、 瀉下作用を示している可能性が示唆された (図9)。4-2. AQP
上述のように、 ビサコジルによる瀉下作用は、
AQP3
の発現低下が起因していることが示唆されたが、 大腸AQP
の発現量が低下すると水の移動が抑えられ、 下痢 が発症するかどうかについてはこれまで全くわかってい なかった。 そこで、 このことを確認する目的で、AQP3
の機能が阻害され、 水の移動が抑制された際に、 糞中水 分量が上昇するのかどうかを調べた。AQP3
の機能を阻 害することが知られている塩化水銀 (HgCl2)32) あるい は硫酸銅 (CuSO4)33) をラットの大腸内に投与した結 果、 投与1
時間以内に下痢が発生することがわかった。この際、
HgCl
2およびCuSO
4を投与しても、 大腸内浸 透圧およびAQP3
の発現量に影響を及ぼさないことが わかった (data not shown)。以上の結果から、 大腸粘膜上皮細胞に局在する
AQP3
の機能を阻害すると、 下痢が発生することが明らかとなっ た。 また、 生理的条件下では、AQP3
を介して、 水は管 腔側から血管側に輸送されており、 ビサコジルの瀉下作 用においてAQP3
の発現低下が重要であることも確認 できた (Fig. 9)。5 AQP3 !"#
$%
28&HT-29細胞を用いた実験から、 ビサコジルが直接、 大
腸粘膜上皮細胞に作用し、AQP3
の発現量を低下させた 可能性は低いことがわかった (data not shown)。 そ こで次に、 ビサコジルによる間接的な作用について検討 を行った。ビサコジルは大腸マクロファージを活性化することが 知られている34, 35)。 マクロファージが活性化すると、
tumour necrosis factor-
(TNF-) などの炎症性サイ トカインの発現および分泌が亢進し、cyclooxygenase-2
(COX-2) の発現増加を介してPGE2が分泌される36, 39)。一方、
TNF-
40. 42)およびPGE
243, 44) は、AQP
の発現量を低下させることが報告されている。 そこで、 この経路 によりビサコジルが
AQP3
の発現量を低下させた可能 性について調べた。5-1. '()*+,-.
ビサコジルが直接マクロファージを活性化し、
TNF-
およびPGE
2を分泌させるかどうかをマウスマクロファー ジ 由 来 細 胞 株Raw264.7
細 胞 を 用 い て 調 べ た 。Raw264.7
細胞にビサコジルを添加した結果、TNF-
の分泌量の増加、COX-2
の発現量の増加およびPGE
2の分泌量の増加が認められた (Fig. 10)。 一方、 ラット にビサコジルを投与し、 下痢が発生した際の大腸におい ても、
TNF-
およびCOX-2
の現増加、 ならびにPGE
2の分泌増加が認められた (Fig. 11)。 さらに、 この
COX-2の発現増加はマクロファージにおいて特異的に
見られた (Fig. 12)。以上のことから、 ビサコジルはマクロファージを活性
Fig. 8 SMIT AQP3
!"#$"%& A '(
)% SMIT mRNA & B ' AQP3
*+,-.& C '/0"%(12 28 3456
Fig. 9 789: AQP3 ;<
化させること、 ならびに
TNF-
およびPGE
2の産生お よび分泌を亢進させることがわかった。5-2. AQP3 TNF- PGE
2TNF- および PGE
2がパラクライン因子として大腸 粘膜上皮細胞に作用し、AQP3
の発現を低下させている かどうかを調べた。 その結果、HT-29
細胞にTNF-
を 添加しても、 添加2
時間後ではAQP3
の発現量に変化 は見られず、6
時間後になってはじめて、 低下すること がわかった (Fig. 13A)。 これに対して、 下痢が発生し たビサコジル投与2
時間後において、 ラット大腸のTNF-
の増加とAQP3
の低下がほぼ同時に見られた (Fig. 8 C and 11A)。 したがって、 マクロファージから分泌された
TNF-
は、 オートクライン因子として、マクロファージの
COX-2
の発現増加には関わっている ものの、 パラクライン因子として大腸粘膜上皮細胞に作 用し、AQP3
の速やかな発現低下を引き起こした可能性 は低いと考えられた。 一方、HT-29
細胞にPGE
2を添 加すると、AQP3
の発現量が著明に低下した。 また、 こ の発現低下は速やかに生じ、PGE
2添加後30分以内には 起きることがわかった (Fig. 13B)。以上のことから、 マクロファージから分泌された
PGE
2 は、 パラクライン因子として大腸粘膜上皮細胞に作用し、AQP3
の発現量を低下させたと考えられる。5-3. COX
次に、
COX
阻害剤インドメタシンを前処置し、PGE
2の分泌を抑制した際に、 ビサコジルによる瀉下作用およ び大腸
AQP3
の発現がどのように変化するかを調べた。その結果、 インドメタシンを前処置することにより、 ビ サコジルの瀉下作用が抑制されるとともに、 大腸AQP3 の発現低下も抑制されることがわかった (Fig. 14)。
Fig. 10 Raw264.7 TNF-
COX-2 PGE
2Raw264.7 TNF-
! A " COX-2 mRNA ! B " PGE
2! C "#$ %&'( 28 )*+,
Fig. 11 -. TNF- COX-2
PGE
2/0123 -. TNF- mRNA
! A " COX-2 mRNA ! B " PGE
2! C "#$ %&'( 28 )*+,
Fig. 12 234/01-. COX-2
56
789:;<=>?8 COX-2 @8A&'( 28 )*+,
Fig.13 AQP3 TNF- PGE
2HT-29 TNF- ! A " PGE
2! B " AQP3 BCD:EFG %&'( 28 )*+,
Fig. 14 HIJK-.L PGE
2M
N AQP3 OIPCQRBSC
PCQRBSCTUL23 %/01VW2
3 X Y5! A "-.L PGE
2! B " AQP3
BCD:E! C "#$ %&'( 28 )*+,
以上のことより、 ビサコジル投与による大腸
AQP3
の急速かつ著明な発現低下には、PGE
2が関与している ことが確認できた。 加えて、 非ステロイド性抗炎症薬 (non-steroidal antiinflammatory drugs;NSAIDs) とビサコジルを併用すると、 ビサコジルの瀉下作用が減 弱することがわかった。6 MgSO
4 45臨床において、 重度な便秘症患者には、 第一選択薬と して浸透圧性下剤 (MgSO4ほか) が処方され、 効果が 認められない場合には大腸刺激性下剤 (ビサコジルほか) など、 作用メカニズムが異なる瀉下剤が併用される46, 47)。 しかし、 瀉下剤の併用により、 瀉下作用が増強するか どうかについての明確なエビデンスはない。 そこで、
MgSO
4およびビサコジルを併用した場合、 瀉下作用が 増強するか否かを検討した。 その結果、MgSO
4とビサ コジルを併用しても、 糞中水分量の変動パターンおよび 変化率には、 両薬剤の相加効果あるいは相乗効果は見ら れず、 ビサコジル単独投与時のそれらとほぼ同様である ことが明らかとなった (Fig. 15)。 一方、 併用投与した 際の大腸内浸透圧はControl
群に比べて高く、MgSO
4単独投与時とほぼ同程度であることがわかった (Fig. 1
6A)。 また、 併用投与時の大腸 AQP3
の発現量は、Con- trol
群に比べて有意に低く、 この低下率はビサコジル単 独投与時とほぼ同程度であることが明らかとなった (Fig. 16B)。以上のことから、
MgSO
4とビサコジルを併用した際 の瀉下作用は、MgSO
4単独投与時よりも減弱し、 ビサ コジル単独投与時の場合とほぼ同程度であることが明ら かとなった。 さらに、MgSO
4とビサコジルを併用した 場合の大腸AQP3
の発現パターンが、 ビサコジル単独 投与時とほぼ同様になることがその理由として考えられ た。7
本研究の結果から、 浸透圧性下剤および大腸刺激性下 剤の瀉下作用において、 大腸の
AQP3
の発現量の変化 が重要な役割を担っていることが明らかとなった。 加え て、 大腸AQP3
の機能を阻害すると、 下痢が発生する ことも明らかとなった。AQP3
はヒトの大腸において最 も多く発現しているAQP
である。 今後、 大腸AQP3
の 発現および機能と水の移動についてさらなる研究を展開 することにより、AQP
をターゲットとした新たな瀉下 剤や止瀉剤の開発が可能になるものと考える。さらに、 本研究により、 瀉下剤の併用が必ずしも瀉下 作用を増強しないことが明らかとなった。 薬物の服用数 の増加は、 薬物間相互作用の増加につながるため、 安易 な併用は避けるべきである。 今後、 瀉下剤に関しても、
その治療効果に対するエビデンスを明確にし、 適正使用 を図ることが必要であると考える。
8
本研究を遂行するにあたり、 平成28年度星薬科大学 大谷記念研究奨励金を賜りましたことに対し、 理事長の 大谷卓男先生ならびに学長の田中
治先生に深く感謝申 し上げます。 また、 本研究の遂行のために多大なるご指 導を賜りました星薬科大学薬動学教室の杉山 清教授 (現;食品動態学研究室・特任教授) に心より御礼申し 上げます。 最後に、 本研究を進めるにあたり、 多大なる ご協力をいただきました薬動学教室の教室員の皆様に深 謝いたします。
開示すべき利益相反はない。