九州大学学術情報リポジトリ
Kyushu University Institutional Repository
Porphyromonas gingivalisとAggregatibacter
actinomycetemcomitansの混合感染における相互作用 に関する研究
髙﨑, 敬
九州大学大学院歯学研究院口腔機能修復学講座歯周病学分野
https://doi.org/10.15017/19952
出版情報:Kyushu University, 2010, 博士(歯学), 課程博士 バージョン:
権利関係:
1
緒言
歯周病は歯周組織の破壊を主たる病態としており、歯肉縁下プラーク中の歯周病 原細菌に起因する感染症である(1、2)。プラークはその70% 以上が細菌の塊で、
その中では数百種類を超える細菌がバイオフィルムを形成し存在している。歯周病 は歯周病原細菌が歯周組織局所に付着し、バイオフィルムを形成して成長すること で、発症し進行していく(3)。菌体外多糖からなるグリコカリックスによって保護 されたバイオフィルム中の細菌は、抗菌物質や貪食細胞、免疫グロブリンに対して 抵抗性を示して局所に長期に留まり、感染の慢性化、難治化を招くと考えられてい る(4、5)。近年では単独バイオフィルムに関する研究だけでなく、複数菌種と形 成する複合バイオフィルムに関する研究も注目され始めたところである。
歯周病 の原 因菌と して 、こ れま でに十 数 種類の 好気 性細菌 (Actinomyces viscosus、Actinomyces naeslundii など)や嫌気性細菌(Prevotella intermedia、
Aggregatibacter actinomycetemcomitans 、 Porphyromonas gingivalis 、 Treponema denticola、 Fusobacterium necrophorum など)が発症に深く関わっ ていると言われている(6)。なかでもPorphyromonas gingivalis は歯周炎の主要 な原因菌として知られており、生理学的特性や病原因子などについて数多くの研究 が報告されている(7‐11)。
グラム陰性嫌気性細菌であるP. gingivalis は、細胞表面に線毛やリポ多糖(LPS)
を有し、菌体表面や菌体外に強力なタンパク分解酵素を産生する特異な細菌であり、
中でも線毛とジンジパインは本菌の付着やバイオフィルム形成において重要な役
2
割を果たしていることが報告されている(12‐16)。P. gingivalis は、線毛の主要 構成タンパクの一つであるフィンブリリン(FimA)の構成遺伝子 fimA の塩基配 列の違いによりI~V型、およびIb型の6 型に分類されており、fimA遺伝子型と 病原性との関連について多くの研究がされている(17、18)。一方、ジンジパイン
は P. gingivalis が産生する総プロテアーゼ活性の少なくとも 85% を占めるシス
テインプロテアーゼであり(19)、そのペプチド結合切断特異性から、アルギニン 残基のC末端側を特異的に切断するArg-gingipain(Rgp)とリジン残基のC末端 側を特異的に切断する Lys-gingipain(Kgp)の2つの酵素群に分類される(20、
21)。ジンジパインはその酵素活性による歯周組織の破壊に関与するだけでなく、
赤血球凝集能や線毛形成等にも関与し、上皮細胞への付着や他の細菌との共凝集に も関与するとの報告もある(22、23)。本菌のバイオフィルム形成因子に関しては、
これまでに線毛やジンジパインが関連しているといういくつかの報告があるもの の、未だ意見が分かれているところである(12‐16)。また、他の菌種との複合バ イオフィルム形成における P. gingivalis の相互作用や関連因子についてはほとん ど未解明の状態である。
Aggregatibacter actinomycetemcomitans も重要な歯周病原細菌であり、これ までに多くの研究報告が得られている(24)。A. actinomycetemcomitans は外膜 構成成分であるLPSのO抗原多糖の構造の違いによりaからfの6つの血清型に 分類されており、この血清型の違いが病原性の違いに関わると示唆されている(25)。 A. actinomycetemcomitans もP. gingivalis と同様に、in vitroの実験において非 生体面に単独のバイオフィルムを形成することがこれまでに多く報告されている が、複合バイオフィルム形成に関する報告はほとんど得られていない(26、27)。
3
そこで本研究では、まず始めに P. gingivalis の単独バイオフィルム形成能につ いて、特に線毛とジンジパインの関与について検討した。更に、P. gingivalis と A. actinomycetemcomitans との共培養下での複合バイオフィルム形成において、
そのバイオフィルムの構造とジンジパインの役割について検討した(第一章)。 バイオフィルム形成過程においては、表面への浮遊菌の付着から始まり、成熟す ると菌同士がシグナルを出して増殖をコントロールしたり、チャネルを介して栄養 源を取り込んだり、他菌の代謝産物などを利用することが知られている(4)。そこ で本研究では、A. actinomycetemcomitans とP. gingivalis との混合感染において 複合バイオフィルムを形成する際の、P. gingivalis の増殖への影響について検討す るため、A. actinomycetemcomitans のP. gingivalis 増殖促進への関与について調 べた(第二章)。
尚、本研究では単一細菌で培養して形成されたバイオフィルムを「単独バイオフ ィルム」、共培養により形成されたバイオフィルムを「複合バイオフィルム」と表 記した。
4
第 一 章 : P. gingivalis の 単 独 バ イ オ フ ィ ル ム 形 成 お よ び
A. actinomycetemcomitans との共培養下での複合バイオフ ィルム形成におけるジンジパインの役割について
1‐1:序論
歯周炎患者の歯周ポケット内には歯周病原細菌がプラーク(バイオフィルム)を 形成し棲息している。プラークは歯周動的治療における歯面清掃やスケーリング・
ルートプレーニングあるいは歯周外科治療により機械的に除去される。しかしなが らこのような治療後、メインテナンス期に移行したにもかかわらず、歯周病の再発 が見られることも多い。これまでに歯周動的治療後のメインテナンス期間中におけ る細菌学的背景についてはいくつか研究報告されており(28‐31)、歯周病の再発 には口腔内に残存した歯周病原細菌の歯周ポケット内への再定着やそれに続くバ イオフィルム形成が重要であると考えられる。バイオフィルム形成・成熟過程にお いて、複数種の細菌間で共生的相互作用や競合的拮抗作用が起きていると考えられ ている(32‐34)。歯周ポケット内において、初期付着能を有する2種類以上の細 菌が同時に存在する状況で、特に共凝集をしない細菌同士であれば、歯面に優位に バイオフィルムを形成するために競合的拮抗作用を示す可能性が考えられるが、こ のような歯周病原細菌の競合的拮抗作用に関する報告はほとんど得られていない。
歯周病原細菌は、初期定着菌群(Early Colonizers)の後からバイオフィルムを 形成するため、後期定着菌群(Late Colonizers)であると考えられている。初期定 着菌群としては Staphylococcus aureus や Streptococcus 属の細菌種などが知ら
5
れているが、これら以外の細菌種でも、潜在的にバイオフィルム形成能をもつ細菌 であれば、環境状態によっては初期に細菌バイオフィルムを形成する可能性がある と考えられる。P. gingivalis も後期定着菌であると言われているが(35)、単独で 非生体面へのバイオフィルム形成能を持つことがこれまでに多く報告されている ことから、歯周動的治療によって一旦感染バイオフィルムが除去された後、浮遊菌 として残存した P. gingivalis が直接歯根面に定着し、さらに他菌に対し拮抗的な 関係を示せば、本菌の優勢なバイオフィルムを形成するかもしれないという可能性 が推察される。
これまでに、P. gingivalis の単独バイオフィルム形成能については、久保庭らが 線毛遺伝子型と関係すると報告している(12)。一方ジンジパインも単独バイオフ ィルム形成に影響するとの報告もあって意見が分かれているところであり(14‐
16)、どちらが本菌のバイオフィルム形成に重要な因子として働くかについては未 だに明らかにされていない。
P. gingivalis と A. actinomycetemcomitans と の 相 互 関 係 に つ い て は 、 PeriasamyらはP. gingivalis ATCC33277株とA. actinomycetemcomitans JP2株
(血清型b)とは相互共生して複合バイオフィルムを形成することを報告している
が(36)、鈴木らはA. actinomycetemcomitans の血清型cはP. gingivalis と共凝 集をするものの、他の血清型を持つA. actinomycetemcomitans とは共凝集しない ことを示した(37)。臨床的な研究では、P. gingivalis が検出される歯周ポケット からは A. actinomycetemcomitans があまり検出されないと示されているものの
(38、39)、in vitro の研究で両菌が拮抗的に作用していると明らかにされた報告 はない。
6
本章では、上記の推察について検討するために、P. gingivalis の初期バイオフィ ルム形成能について調べるとともに、本菌とA. actinomycetemcomitans 菌株との 複合バイオフィルム形成における競合的拮抗作用について調べた。
7 1‐2:材料及び方法
1.使用菌株と培養条件
本研究で用いたP. gingivalis 株(表1に示す)およびA. actinomycetemcomitans
ATCC29523株は九州大学大学院歯学研究院口腔機能修復学講座歯周病学分野で継
代保存されているものを用いた。P. gingivalis 臨床分離菌株は九州大学病院歯周病 科を受診した慢性歯周炎患者から採取され、以前に本研究室で分離され PCR 法に より線毛タイプが同定されたものである(40、45)。
P. gingivalis は羊血液寒天培地(5% defibrinated laked sheep blood、4% tryptic soy agar、0.5% brain heart infusion、0.1% cysteine、hemin 5 μg /ml、menadione 1 μg /ml)あるいはhemin 5 μg /ml、 menadione 1 μg /ml を含むenriched BHI broth(A)(3.7% brain heart infusion、0.5% yeast extract、0.05% cysteine)を 用いて酸素吸収剤であるアネロパック・ケンキ(三菱ガス化学株式会社)を使用し、
嫌気的条件下にて37℃で培養した(46)。
A. actinomycetemcomitans ATCC29523 株の培養には TSB 寒天培地(3%
trypticase soy broth、0.3% yeast extract、5% heat-inactivated horse serum、
1.5% agar)あるいはenriched BHI broth(A)からhemin、menadione を除い たenriched BHI broth(B)を用い、炭酸ガス発生剤であるアネロパック・CO2 (三 菱ガス化学株式会社)を使用し、5% CO2下、37℃にて培養した(47)。
本研究における全ての菌株の一次培養はenriched BHI broth(A)または(B)
を用いて、それぞれ嫌気下あるいは5% CO2下で行い、対数増殖期まで培養したも
8 のを二次培養に用いた。
なお、P. gingivalis ATCC33277株のジンジパイン変異株(KDP133・KDP129・
KDP136)及び線毛欠損株(KDP150)は長崎大学大学院医歯薬学総合研究科新興 感染症病態制御学専攻 感染免疫学講座口腔病原微生物学分野 中山浩次教授より 恵与された。
9 表 1 P. gingivalis 使用菌株
Strain Characteristics Source/reference
ATCC33277 Wild type (fimA type I) ATCC
MPW1-01 Clinical isolate (fimA type I) (40)
ATCC53977 Wild type (fimA type II) ATCC
MPW2-01 Clinical isolate (fimA type II) (40) MPW2-03 Clinical isolate (fimA type II) This study
W50 Wild type (fimA type IV) (41)
W83 Wild type (fimA type IV) (42)
MPW1b-01 Clinical isolate (fimA type Ib) This study KDP133 Rgp-null derivative of ATCC33277
(rgpA::Tcr rgpB::Emr)
(43)
KDP129 Kgp-nullderivative of ATCC33277 (kgp::Cmr)
(43)
KDP136 Rgp/Kgp-nullderivative of ATCC33277 (rgpA::Tcr rgpB::Emr kgp::Cmr)
(43)
KDP150 fimbria-less derivative of ATCC33277 (fimA::Emr)
(44)
10
2.P. gingivalis 単独バイオフィルム形成とバイオフィルム形成量の測定
ガラス試験管内の3 ml enriched BHI broth(A)に各P. gingivalis の細菌数が
3~5 × 107 個 / mlになるように一次培養液をそれぞれ懸濁し、二次培養(嫌気下、
37℃)を24時間行い、ガラス試験管壁にバイオフィルムを形成させた。培養後バ
イオフィルムが形成されたガラス試験管壁から非付着菌を除去するため、ボルテッ クスミキサーを用いて 5 秒間振盪した。いずれの菌株も培養液中に吸光度が 590 nm で約1.0まで増殖していることを確認した後、3 ml PBS(pH 7.4)にて試験 管内壁を2回洗浄した。洗浄後、3 ml 0.1% クリスタルバイオレットにて10分間 染色を行い、観察した。乾燥後、3 ml 95% エタノールにて色素抽出を行い、可視 光度計(Colorwave CO7500 Colorimeter、 Biochromlimited、 Cambridge、 UK ) を用い吸光度590 nmで測定しバイオフィルム形成量を測定した(48)。さらにガ ラス試験管内にあらかじめ入れておいたカバーガラス(Micro Cover Glass、 Matsunami Glass Industries Limited、 Osaka、 Japan)上にも同じ培養条件で バイオフィルムを形成させた。カバーガラス上の非付着菌はPBS中で1000 μl 容 量のピペッティング操作を 10 回行うことで除去した。通法に従いグラム染色を行 い、光学顕微鏡(BX50-DP70、 Olympus、 Tokyo、 Japan)にて菌体の付着の 確認を行った。
図1と2の結果は、再現性の得られた3回以上の評価の中で代表的なものを示し ており、グラフはトリプリケイトで行った実験で得られた値の平均値から算出した ものである。
11 3.ジンジパイン活性の測定
前述のP. gingivalis 単独バイオフィルム形成と同じ培養条件で各 P. gingivalis 菌株を二次培養し、培養後に菌体を遠心分離(6,000 × g、4℃、15分)にて回収し た。回収した菌体は3 ml 緩衝液(1 mM CaCl2、 50 mM Tris-HCl : pH 7.4)で2 回洗浄し、再度 3 ml 緩衝液に懸濁した後、タンパク量を PROTEIN ASSAY
(BIO-RAD、 Hercules、 CA、 USA)を用いて測定した(49)。標準タンパク としてウシ血清アルブミンタンパクを用い、菌体懸濁液が50 μg / mlのタンパク濃 度になるように緩衝液を追加して調整した。Arg-ジンジパイン(Rgp)活性とLys- ジンジパイン(Kgp)活性は合成基質BAPNA(Nα-benzoyl-DL-Arg-p-nitroanilide)
およびTGPLPNA(N-p-Tosyl-Gly-Pro-Lys-p-nitroanilide) (Sigma、 Aldrich、
St Louis、 MO、 USA)を用いて測定した(50)。Microtiter plate上で、タンパ ク量を調整した菌体懸濁液40 μlに対し、160 μlの反応液(0.25 mM BAPNAまた はTGPLPNA、10 mM CaCl2、 100 mM Tris-HCl : pH 8.5)を添加し、37℃で10 分間インキュベート後、50% 酢酸にて反応を停止させた。反応後、分光光度計
(Jasco Ubest-30 UV/VIS Spectrophotometer、 Jasco、 Tokyo、 Japan)を用
い405 nmにて吸光度を測定し、Rgp活性及びKgp活性を求めた。
図3の結果は、再現性の得られた3回以上の評価の中で代表的なものを示してお り、グラフはトリプリケイトで行った実験で得られた値の平均値から算出したもの である。
12
4.A. actinomycetemcomitans 単独あるいはP. gingivalis との共培養下における 複合バイオフィルム形成とバイオフィルム形成量の測定
P. gingivalis 株は単独バイオフィルム形成能およびジンジパイン活性の違いか
らATCC33277株、ATCC53977株、KDP136株(Rgp・Kgp欠損株)の3菌株を 選択した。A. actinomycetemcomitans 株はP. gingivalis 株と同じ培養条件下でガ ラス試験管壁にバイオフィルムを形成するATCC29523株を用いた。
それぞれ細菌数が3~5 × 107 個 / mlになるように3 ml enriched BHI broth(A)
に一次培養液を懸濁し、P. gingivalis の培養条件に合わせて18 時間の二次培養(嫌 気下、37℃)を行い、A. actinomycetemcomitans 単独あるいは各P. gingivalis 株 との共培養下でガラス試験管壁にバイオフィルムを形成させた。培養後、前述と同 じ手順でボルテックスミキサーを用いて非付着菌の除去を行い、試験管内壁をPBS で洗浄し、クリスタルバイオレットにて染色後、エタノールにて色素抽出を行い、
吸光度を測定した。なお、各 P. gingivalis 株の単独バイオフィルム形成は前述と 同じ手順で行った。
図4の結果は、再現性の得られた3回以上の評価の中で代表的なものを示してお り、グラフはトリプリケイトで行った実験で得られた値の平均値から算出したもの である。
13
5.P. gingivalis とA. actinomycetemcomitans の単独あるいは複合バイオフィル ム中の構成細菌数の計測
P. gingivalis ATCC33277株、ATCC53977株及びKDP136株(Rgp・Kgp欠損 株)をそれぞれA. actinomycetemcomitans ATCC29523株との共培養に用いた。
あらかじめカバーガラスを入れた細胞培養用滅菌シャーレ(Tissue Culture Dish style 60×15 mm style、 Falcon、 Becton Dickinson、 USA)内のenriched BHI broth(A)にそれぞれ細菌数が3~5 × 107 個 / mlになるように一次培養液を懸濁 し、二次培養(嫌気下、37℃)を18時間行った。培養後、シャーレ内の培養液中
で1000 μl 容量のピペッティング操作を10回行い、カバーガラスから非付着菌を
除去した。カバーガラスを取り出して、ピペットを用い1 ml PBSで3回洗浄した 後、形成されたバイオフィルムをセルスクレーパー(Sarstedt、 Newton、 NC、
USA)で回収した。バイオフィルム中の菌体をPBSにて段階希釈して、嫌気下の
羊血液寒天培地あるいは5% CO2下のTSB寒天培地で5日間培養した。
培養後、 羊血液寒天培地上で 黒いコロニーを形成したものを P. gingivalis
ATCC33277株あるいはATCC53977株の細菌数、TSB寒天培地上に形成したコロ
ニーを A. actinomycetemcomitans の細菌数とし、それぞれ単位面積あたりの
colony forming units(CFU)を算出した。黒色コロニーを形成しないP. gingivalis KDP136株については200 µg /ml カナマイシン含有のP. gingivalis 選択培地を用 いて行った。同様に培養液1 mlあたりの各浮遊菌数についても細菌数を算出した。
表2と図5・7に示した値は、それぞれ独立した3回以上の評価の中で再現性の ある代表的なものの中から選ばれ、表2はその中から3つの結果の平均を示した。
14
6.P. gingivalis 培養上清による A. actinomycetemcomitans ATCC29523株の バイオフィルム剥離試験
各P. gingivalis 株の細菌数が3~5 × 107 個 / mlになるようにガラス試験管内 の3 ml enriched BHI broth(A)に一次培養液を懸濁し、18時間の二次培養(嫌 気下、37℃)を行った。培養後、二次培養液を遠心分離(6,000 × g、4℃、15分)
し、各P. gingivalis 株の培養上清を回収した。
A. actinomycetemcomitans ATCC29523株の細菌数が3 × 107 個 / mlになるよ うにガラス試験管内の3 ml enriched BHI broth(B)に懸濁し、18時間の二次培 養(5% CO2下、37℃)を行い、ガラス試験管壁にバイオフィルムを形成させた。
A. actinomycetemcomitans ATCC29523株のバイオフィルムが形成された試験 管に、各P. gingivalis 株の培養上清をそれぞれ3 ml分注し、37℃で60分間イン キュベートした。インキュベート後、試験管内壁を PBS にて洗浄し、剥離された 菌体を除去した後、前述に従いクリスタルバイオレットにて染色し、剥離されずに 試験管内壁に残存したバイオフィルム量の測定を行った。また P. gingivalis 培養 上清中のタンパク分解活性の関与を探るため、P. gingivalis ATCC33277株の培養 上清を回収し、最終濃度が500 μg /ml になるようにProteinase K(和光純薬工業 株式会社)を加えて処理(37℃、60 分)したもの、あるいは熱処理(100℃、10 分)を行ったものを用いた。
図6の結果は、再現性の得られた3回以上の評価の中で代表的なものを示してお り、トリプリケイトで行った実験で得られた値の平均値から算出したものである。
15 7.統計解析
各P. gingivalis 菌株とA. actinomycetemcomitans ATCC29523株の共培養中の 構成細菌数の比較(表2)はスチューデントのt‐検定によって統計学的有意差の 有無を検定した。
16 1‐3:結果
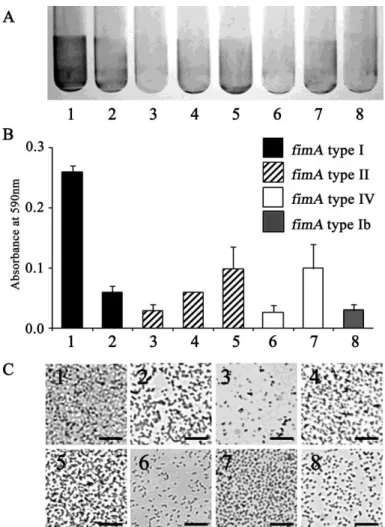
1)P. gingivalis 線毛遺伝子型の単独バイオフィルム形成への関与
研究室保存株及び臨床分離株(線毛タイプ I 型・II 型・IV 型・Ib 型を含む)
8菌株全てがガラス面に付着していることが確認できた(図1C)。バイオフィルム 形成量は株間で差があり、特に ATCC33277 株のバイオフィルム形成量が多く、
ATCC53977 株が最も少なかった(図1A・B)。また、ATCC33277 株と同じ線毛
タイプI型であるMPW1‐01とを比較するとバイオフィルム形成量に差があった。
同様に、II型線毛型株間およびIV型線毛型株間でも、バイオフィルム形成量は異 なり、本菌のバイオフィルム形成量と線毛遺伝子型は非依存的であることが示され た。
17
Scale bars represent 10 µm
図 1 各P. gingivalis 株のバイオフィルム形成量の比較
(A)クリスタルバイオレット染色、(B)定量、(C)グラム染色像 1:ATCC33277、 2:MPW1‐01、 3:ATCC53977、 4:MPW2‐01 5:MPW2‐03、 6:W50、 7:W83、 8:MPW1b‐01
全ての株でガラス面への付着を認めた。ATCC33277株のバイオフィルム形成量 が最も多く、ATCC53977株の形成量が最も少なかった。バイオフィルム形成量は 線毛遺伝子型には非依存的だった。
18
2)P. gingivalis ジンジパインの単独バイオフィルム形成への関与
単独バイオフィルム形成量の多かったATCC33277株について、ジンジパイン変 異株を用いて単独バイオフィルム形成へのジンジパインの関与について調べたと ころ、野生株である ATCC33277 株と比較して、KDP129 株(Kgp 欠損株)のバ イオフィルム形成量は減少しなかったものの、KDP133 株(Rgp欠損株)は減少し、
KDP136株(Rgp・Kgp欠損株)はバイオフィルムをほとんど形成しなかった(図
2)。一方、KDP150 株(線毛欠損株)は野生株と同等量の単独バイオフィルムを 形成した(データ省略)。このことから、本菌のバイオフィルム形成にはジンジパ イン特に Rgp が関与していることが示唆されたため、ジンジパインのプロテアー ゼ活性とバイオフィルム形成量との関与について調べたところ、膜結合型ジンジパ インのタンパク分解活性とバイオフィルム形成量との間に相関性は認めなかった
(図3)。
19
図 2 各ジンジパイン変異株のバイオフィルム形成量の比較
(A)クリスタルバイオレット染色、(B)定量
1:ATCC33277(野生株)、 2:KDP133(Rgp欠損株)、
3:KDP129(Kgp欠損株)、 4:KDP136(Rgp・Kgp欠損株)
野生株と比較してKDP129株のバイオフィルム形成量は減少しなかったが、
KDP133株のバイオフィルム量は減少し、KDP136株はバイオフィルムをほとん
ど形成しなかった。
20
Rgp 活性
Kgp 活性
図 3 各P. gingivalis 株の膜結合型ジンジパインのRgp活性、Kgp活性の比較 1:ATCC33277、 2:MPW1‐01、 3:ATCC53977、 4:MPW2‐01
5:MPW2‐03、 6:W50、 7:W83、 8:MPW1b‐01
ジンジパイン活性と単独バイオフィルム形成量との間に相関性はなかった。
21
3)P. gingivalis とA. actinomycetemcomitans の単独あるいは複合バイオフィル ム形成量の比較
A. actinomycetemcomitans ATCC29523株はP. gingivalis ATCC33277株のよ うに視覚化できる単独バイオフィルムを形成することが確認された(図4A)。図1 と 2より、単独バイオフィルム形成能の高い株(ATCC33277 株)と低い株
(ATCC53977 株)、およびバイオフィルムをほとんど形成しないジンジパイン欠 損株(KDP136株)を用い、A. actinomycetemcomitans との複合バイオフィルム 形成量を比較した。複合バイオフィルム形成量はいずれも単独バイオフィルムに比 べ形成量が多かった。A. actinomycetemcomitans ATCC29523株の単独バイオフ ィルム形成量と比較して、ATCC33277株やKDP136株との複合バイオフィルム形 成量はさほど変わらなかったが、ATCC53977株との複合バイオフィルムはあまり
形 成 さ れ な か っ た ( 図 4B )。 こ れ ら の 結 果 よ り 、P. gingivalis と
A. actinomycetemcomitans の間でバイオフィルム形成における何らかの競合的
拮抗作用が起きている可能性が考えられた。
22
図 4 各P. gingivalis (P. g )株とA. actinomycetemcomitans (A.a )
ATCC29523株の単独あるいは複合バイオフィルム形成量の比較
(A)クリスタルバイオレット染色、(B)定量 1:A. a ATCC29523株 単独
2:P. g ATCC33277株 単独、3:P. g ATCC33277株とA. a ATCC29523株 複合 4:P. g ATCC53977株 単独、5:P. g ATCC33277株とA. a ATCC29523株 複合 6:P. g KDP136株 単独、 7:P. g KDP136株とA. a ATCC29523株 複合
単独バイオフィルム形成量に比べ、いずれも複合バイオフィルム形成量の方が多
かった。A. a ATCC29523 株の単独バイオフィルム形成量と比較して、P. g
ATCC33277 株や KDP136 株との複合バイオフィルム形成量はさほど変わらなか
ったが、ATCC53977株との複合バイオフィルムはあまり形成されなかった。
23
4)P. gingivalis とA. actinomycetemcomitans との共培養下における浮遊細菌数 と複合バイオフィルム中の構成細菌数の比較
表2は共培養18時間後の浮遊細菌数とバイオフィルム細菌数をP. gingivalis と A. actinomycetemcomitans それぞれについて調べた結果である。
3)と同様に、3種のP. gingivalis 菌株を用いて3種類のコンビネーション毎に解 析した。浮遊細菌数を比較すると、いずれの場合においても P. gingivalis と A.
actinomycetemcomitans の細菌数に差はなかった。しかしながら、単独バイオフ
ィルム形成能が高い ATCC33277 株を用いたコンビネーション#1において、複合 バイオフィルム中の構成細菌数は有位に P. gingivalis が多く、その割合は98.9%
を占めていた。コンビネーション#2では、A. actinomycetemcomitans の細菌数 はコンビネーション#1と同様に少なかったものの、単独バイオフィルム形成能の
低い ATCC53977 株の細菌数も少ないままであった。一方、コンビネーション#3
においては複合バイオフィルム中の A. actinomycetemcomitans 細菌数が圧倒的
に多く、KDP136株の占める割合は1.8% と有位に少なかった。以上より、共培養
下の複合バイオフィルム形成においては、P. gingivalis のジンジパインにより
A. actinomycetemcomitans バイオフィルム形成が阻害されるような競合的拮抗
作用が起きていること、更に P. gingivalis の単独バイオフィルム形成能が高いと
P. gingivalis 優勢の複合バイオフィルムが形成されることが示唆された。
24
表 2 各P. gingivalis 菌株とA. actinomycetemcomitans ATCC29523株の 共培養中の構成細菌数の比較;浮遊細菌およびバイオフィルム細菌について
*
Determined by Student’s t test. P values of < 0.01 are considered significant.Strain 浮遊細菌数
( Log10 CFU/ml ± SD)
バイオフィルム細菌数
( Log10 CFU/cm2 ± SD) Combination #1
P. gingivalis ATCC33277 9.68 ± 0.21 7.90 ± 0.05 A. actinomycetemcomitans ATCC29523 9.39 ± 0.16 6.19 ± 0.23
*
Combination #2
P. gingivalis ATCC53977 9.32 ± 0.12 6.42 ± 0.21 A. actinomycetemcomitans ATCC29523 9.45 ± 0.11 6.58 ± 0.26 Combination #3
P. gingivalis KDP136 8.88 ± 0.17 5.78 ± 0.15 A. actinomycetemcomitans ATCC29523 9.35 ± 0.12 7.72 ± 0.26
*
25
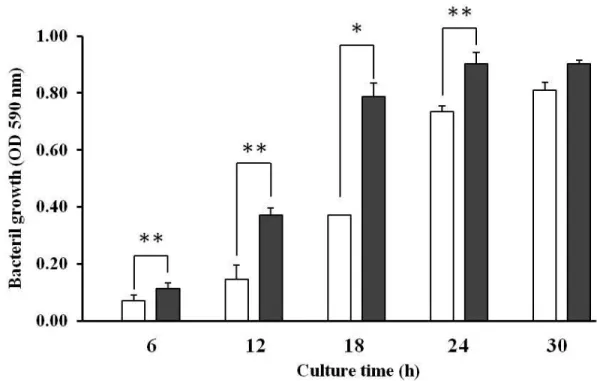
5)P. gingivalis とA. actinomycetemcomitans のバイオフィルム中の構成細菌数 の経時的変化;単独培養時と共培養時の比較
図5はP. gingivalis ATCC33277株とA. actinomycetemcomitans ATCC29523 株の単独培養時と共培養時において、バイオフィルム形成細菌数について経時的に 調べ、比較したものである。共培養下においては、A. actinomycetemcomitans は 培養初期からバイオフィルムの形成が抑制されており、6時間頃をピークに細菌数 が減少した。一方、P. gingivalis は単独培養時よりも共培養時でバイオフィルム形 成能が増加した。この結果より、A. actinomycetemcomitans の方が先にバイオフ ィルムを形成し始めるものの、P. gingivalis との共培養下ではその形成が抑制され ており、複合バイオフィルム形成初期からP. gingivalis 優勢の競合的拮抗作用が 起きていることが示唆された。
26
図 5 バイオフィルム中の各構成細菌数の経時的変化;単独バイオフィルムと複合 バイオフィルムの比較;単独培養時と共培養時の比較
P. gingivalis(P. g ):ATCC33277株、
A. actinomycetemcomitans(A. a ):ATCC29523株
それぞれ単独培養および共培養下で複合バイオフィルムを形成させた。
バイオフィルム中のP. gingivalis 細菌数は、単独培養に比べると共培養下で増 加したが、A. actinomycetemcomitans 細菌数は共培養下で減少した。
27
6)各P. gingivalis 株培養上清のA. actinomycetemcomitans バイオフィルム剥 離効果の比較
4)と5)でみられた競合的拮抗作用のメカニズムについて追求するために、あ らかじめA. actinomycetemcomitans ATCC29523株単独で形成させたバイオフィ ルムに対し、各P. gingivalis 株の培養上清で処理を行い観察した(図6)。
ジンジパイン産生能を有する ATCC33277 株、ATCC53977 株の培養上清は A. actinomycetemcomitans ATCC29523 株のバイオフィルムを剥離した。また
ATCC33277 株の培養上清にあらかじめ熱処理やProteinase K処理を行った培養
上清では剥離しなかった。更にジンジパイン変異株において、KDP133 株(Rgp 欠損株)や KDP129 株(Kgp 欠損株)の培養上清では剥離量が減少し、KDP136 株(Rgp・Kgp欠損株)の培養上清では剥離しなかった。また、剥離された細胞の 生死を確認したところ、細胞は死滅していないことを確認した(データ省略)。
以上の結果から、培養上清中のジンジパインがA. actinomycetemcomitans の剥 離に関与していることが示唆された。
28
図 6 各P. gingivalis 培養上清によるA. actinomycetemcomitans バイオフィル ムの剥離
(A)クリスタルバイオレット染色、(B)定量
A. actinomycetemcomitans ATCC29523株のバイオフィルムを各P. gingivalis 培 養上清を用いて、37℃にて60分間処理し、洗浄後に染色および定量を行った。
1:fresh BHI(コントロール)、 2:ATCC33277(野生株)、 3:ATCC53977、
4:KDP133(Rgp 欠損株)、 5:KDP129(Kgp 欠損株)、 6:KDP136(Rgp・
Kgp欠損株)、 7:proteinase K-treated ATCC33277、 8:heat-treated ATCC33277 A. actinomycetemcomitans ATCC29523 株のバイオフィルムは P. gingivalis
ATCC33277株とATCC53977株の培養上清により剥離された。また、KDP133株
と KDP129 株の培養上清では剥離量が減少し、KDP136 株やタンパク変性処理を
行ったATCC33277株の培養上清では剥離されなかった。
29 1‐4:考察
本研究では「メインテナンス期に浮遊菌として存在するP. gingivalis が他菌と 競合して直接歯根面に定着するかもしれない」という推察に対する検討を行った。
図1の結果より、本実験に使用した全ての研究室保存株および臨床分離株の8菌 株はカバーガラスに付着する能力があることが示された。このことはP. gingivalis が初期付着菌になりうるポテンシャルを有していることを示唆した。しかしバイオ フィルム形成量は株間で差があり、線毛遺伝子型に非依存的であった。一方、久保 庭らは以前の報告で線毛遺伝子型がバイオフィルム形成に関連すると示唆してい る(12)。しかしながら、彼らの研究では同一線毛遺伝子型で単一菌株しか用いて なかった。また稲葉らは同じ線毛遺伝子 II 型の株間でマウスの膿瘍形成能に差が あることを報告している(51)。これらのことより、本菌のバイオフィルム形成能
を含む phenotype の違いは同じ線毛遺伝子を有していても、株間で差があること
が推測された。
そこでジンジパインとの関連について検討したところ、ジンジパイン、特にRgp が単独バイオフィルム形成に関与することが示唆された(図2)。図3からジンジ パイン活性との関連は明らかでなかったが、KDP136株(Rgp・Kgp欠損株)がほ とんどバイオフィルムを形成しなかったことから、ジンジパインが本菌の他の膜タ ンパク質や分泌タンパク質のプロセシングを介して、間接的に関与しているものと 推察された。Grenier らも本研究と同様にジンジパイン欠損株を使った実験から、
ジンジパインと単独バイオフィルム形成との関連を示しており、これは本研究の結 果を支持するものである(15)。
30
複合バイオフィルム形成時における P. gingivalis の競合的拮抗作用について調 べるため、共培養下におけるA. actinomycetemcomitans との複合バイオフィルム
への P. gingivalis 単独バイオフィルム形成能やジンジパインの関与について検討
した。図1~3の結果より、バイオフィルム形成能が高くジンジパイン活性を持つ ATCC33277 株 、 バ イ オ フ ィ ル ム 形 成 能 は 低 い が ジ ン ジ パ イ ン 活 性 を 持 つ
ATCC53977 株、バイオフィルム形成能がほとんどない KDP136 株(Rgp・Kgp
欠損株)の3菌株を選択し、各株とA. actinomycetemcomitans との共培養を行い 比較した。A. actinomycetemcomitans 株はガラス試験管壁表面に単独バイオフィ ルム形成能を持ち、またP. gingivalis と共凝集しない血清型aのATCC29523株 を選択した。図4より、ジンジパインがA. actinomycetemcomitans とP. gingivalis の競合的拮抗作用に関与することが考えられた。更に表2より、ジンジパインと単 独バイオフィルム形成能の両方を兼ね備えれば、P. gingivalis が優勢の複合バイオ フィルムを形成する可能性が示された。図5より P. gingivalis 優勢な競合的拮抗 作用はバイオフィルム形成初期から起きており、A. actinomycetemcomitans のバ イオフィルム形成阻害と P. gingivalis のバイオフィルム形成促進が起きている可 能性が示され た。この P. gingivalis 優勢に なるメカニ ズムに つ いて は、A.
actinomycetemcomitans 剥離が複合バイオフィルム形成中に起きており、その剥
離にジンジパインが関与することが示唆された(図6)。濃縮されたP. gingivalis の 培養上清が A. actinomycetemcomitans のバイオフィルムを形成阻害することも 確認されているが(データ省略)、ジンジパインが剥離やバイオフィルム形成阻害 にどのように働いているかについては、今後さらに検討すべき課題である。
これまでに A. actinomycetemcomitans の付着因子あるいはバイオフィルム形
31
成因子について、バイオフィルム基質中のタンパクや線毛、菌体外多糖および菌体 外 DNAなど多くの因子が関わっているという報告があり(26、27、48)、今回の 複合バイオフィルム形成においてはこのうちのタンパクに対してジンジパインが 影響し、A. actinomycetemcomitans 剥離が起こった可能性が考えられる。未発表 データではあるが、A. actinomycetemcomitans ATCC29523株の単独バイオフィ ルムがProteinase K処理で剥離されることからも、A. actinomycetemcomitans 剥 離に関与するのはジンジパインのプロテアーゼ活性である可能性が示唆された。
図5よりP. gingivalis よりもA. actinomycetemcomitans の方が先にバイオフィ ルムを形成するものの、ジンジパインはそれを剥離することで、P. gingivalis は
A. actinomycetemcomitans に対し競合的優位性を有する複合バイオフィルムを
形成することが推測された。我々の知る限り、本研究は両細菌の競合的拮抗作用お
よびP. gingivalis の競合的優位性についての初めての報告である。
スケーリング・ルートプレーニングなどの歯周動的治療後における再感染におい ては、口腔内に残存して浮遊している複数菌種が同時に根面への再定着をチャレン ジする機会であると考えられるが、本研究の結果から P. gingivalis の株によって は歯周ポケット内で P. gingivalis 優勢のバイオフィルムを形成しうる可能性が示 唆された。これは、P. gingivalis 再定着がメインテナンス期間中の早期に始まると いう報告(52)とも一致すると考えられるが、菌株によってこの現象および臨床症 状に違いを与える可能性が考えられた。
本研究の結果より、ジンジパインは P. gingivalis の単独バイオフィルム形成能 に関与するとともに、A. actinomycetemcomitans との複合バイオフィルム形成に
おいてP. gingivalis が競合的優勢を得ることにも関与することが示唆された。
32
第 二 章 : A. actinomycetemcomitans と の 共 培 養 下 で の
P. gingivalis の増殖におけるジンジパインの役割について
2‐1:序論
P. gingivalis は種々の病原因子を有するが、その病原性を発揮する上で本菌の増
殖は重要なファクターの一つとして挙げられる。P. gingivalis は糖発酵能を持たず、
栄養・エネルギー源をもっぱら外界のタンパク質やペプチドに依存しており(53
‐55)、その発育にヘミンとビタミン K であるメナジオンを要求する(56)。これ までに、特定の細菌種の代謝副産物であるビタミン K 合成に関連したナフトキノ
ン(57)やヘミンの代わりに増殖促進因子として働くコハク酸が Treponema
denticola などから産生され、P. gingivalis の増殖促進に関わるという細菌間相互 関係が示されてきた(56、58‐60)。また、YonedaらはマウスにP. gingivalis と
Tannerella forsythia を混合感染させると膿瘍形成が相乗的に増大することを示
し(61)、さらにその相乗効果をもたらすメカニズムの可能性の一つとして、
T. forsythia の細胞構成成分中のタンパク質が P. gingivalis の増殖促進に働くこ とを示した(62)。同様にChenらもP. gingivalis とA. actinomycetemcomitans の 混合感染がマウスにより大きな膿瘍形成をもたらすことを報告している(63)。
しかしA. actinomycetemcomitans のP. gingivalis に対する増殖促進への関与は これまでに報告されていない。
第一章では本菌のバイオフィルム形成に注目し、A. actinomycetemcomitans と の共培養下での複合バイオフィルム形成におけるジンジパインの作用について述
33
べた。この際、単独培養時と共培養時のバイオフィルム細菌数と浮遊細菌数につい て経時的な変化を観察したところ、浮遊細菌中の P. gingivalis は単独培養時に比 べ A. actinomycetemcomitans との共培養下でその増殖増強効果が見られる傾向 にあった(図7)。
そこで本章ではA. actinomycetemcomitans の細胞構成成分がP. gingivalis の 増殖促進に何らかの影響を与えているのではないかと考え、P. gingivalis の増殖実 験を行った。
34
図 7 P. gingivalis(P. g )ATCC33277株の浮遊細菌数の経時的変化;単独培養 下とA. actinomycetemcomitans(A. a )ATCC29523株との共培養下での比較
P. gingivalis は単独培養した時に比べ、A. actinomycetemcomitans との共培養 下において増殖増強効果が見られた。
35 2‐2:材料及び方法
1.使用菌株と培養条件
各P. gingivalis 株あるいは A. actinomycetemcomitans 株の培養条件は第一章 に準じて行った。
2.P. gingivalis のenriched BHI broth(A)濃度による増殖量の測定
P. gingivalis の増殖量の差をより明白にする目的でenriched BHI broth(A)を PBS で希釈していき、100% 濃度で定常期まで培養した時の約半分の増殖量を示 すenriched BHI broth(A)濃度を模索した。
PBSにて各濃度(100%、 80%、 60%、 40%、 20%)に希釈した3 ml enriched BHI broth(A)に、P. gingivalis ATCC33277 株の細菌数が3 × 107 個/ mlにな るように一次培養液を懸濁し、二次培養(嫌気下、37℃)を30時間行った。培養 後、可視光度計(Colorwave CO7500 Colorimeter、 Biochromlimited、 Cambridge、
UK )を用い吸光度590 nmで測定し増殖量を測定した。
図8の結果は、再現性の得られた3回以上の評価の中で、トリプリケイトで行っ た実験で得られた値の平均値から算出したものである。
36
3.A. actinomycetemcomitans のcell extractsの回収とタンパク変性処理
A. actinomycetemcomitans ATCC29523株はenriched BHI broth(B)で二次 培養(5% CO2下、37℃)を18時間行った後、遠心分離(6,000 × g、 4℃、15分)
にて菌体を回収した。残存する培養上清を除去するために PBSにて菌体を2回洗 浄した。菌体を再度 PBS に懸濁し、ガラスビーズを用いてビーズ式ホモジナイザ ー(MINI-BEADBEATERTM、BIOSPEC)にて4℃で冷却しながら1分間の菌体 粉砕を、冷却のため2分間の休止をおきながら合計3回行った。その後菌体粉砕液 を遠心分離(6,000 × g、 4℃、10分)し、さらにその上清をポアサイズ0.2 μmの フィルター(Minisart
®
sartorius stedim、biotech)にてろ過滅菌し、cell extracts として回収した。cell extracts のタンパク量は PROTEIN ASSAY(BIO-RAD、Hercules、 CA、 USA)を用いて測定した(49)。標準タンパクとして既知濃度
のウシ血清アルブミンタンパクを用いた。
また、A. actinomycetemcomitans のcell extractsのタンパク変性による増殖促 進効果の違いについて調べるために、cell extracts のタンパク濃度を測定した後、
熱処理(80℃、10分)によりcell extractsのタンパク変性処理を行った。
37
4.P. gingivalis 増殖促進におけるA. actinomycetemcomitans のcell extracts 中のタンパクの影響および経時的増殖量の変化
回収したA. actinomycetemcomitans のcell extractsを各タンパク濃度(0、10、
20、30、40、50、100、400、800 μg /ml )になるように添加した3 ml 40% 濃度 のenriched BHI broth(A)にP. gingivalis ATCC33277 株の細菌数が3 × 107 個 / mlになるように一次培養液を懸濁し、二次培養(嫌気下、37℃)を18時間行っ た。培養後、吸光度を測定し増殖量を測定した。
経時的増殖量の変化はA. actinomycetemcomitans のcell extractsのタンパク 濃度が50 μg /ml になるように添加した3 ml 40% 濃度のenriched BHI broth(A)
にて、6 時間毎に 30 時間までの二次培養(嫌気下、37℃)を行い、経時的な増殖 量の変化を観察した。
P. gingivalis 増殖促進のメカニズムについて検討するため、タンパク変性処理を
行ったcell extracts のタンパク濃度が50 μg /ml になるように添加した3 ml 40%
濃度のenriched BHI broth(A)にて、18時間の二次培養(嫌気下、37℃)を行 った。標準タンパクとしてタンパク濃度50 μg /ml のウシ血清アルブミンタンパク
(BSA)も用いた。
さらにジンジパインの関与について調べるために、各ジンジパイン変異株をcell extracts のタンパク濃度が 50 μg /ml になるように添加した 3 ml 40% 濃度の
enriched BHI broth(A)にて、18時間の二次培養(嫌気下、37℃)を行った。
図9~12の結果は、再現性の得られた3回以上の評価の中で、トリプリケイト で行った実験で得られた値の平均値から算出したものである。
38 5.統計解析
P. gingivalis 増殖促進におけるA. actinomycetemcomitans のcell extracts中 のタンパクの影響および経時的増殖量の変化(図 10~12)の結果についてスチュ ーデントのt‐検定によって統計学的有位差の有無を検定した。
39
2‐3:結果
1)各BHI濃度におけるP. gingivalis 増殖量
P. gingivalis の増殖量の差をより明白にする目的で、enriched BHI broth(A)
を PBS で希釈していき、定常期を示す 30 時間の二次培養後、100% 濃度の enriched BHI broth(A)での培養と比較してその約半分の増殖量を示すenriched
BHI broth(A)濃度を模索した。その結果、40% 濃度での増殖量が100% 濃度で
のおよそ半分の増殖量を示した(図8)。従って、以降の実験は40% 濃度のenriched BHI broth(A)を用いて行った。
40
図 8 各濃度のenriched BHI broth(A)でのP. gingivalis 増殖量の測定
PBSにて各濃度に希釈したenriched BHI broth(A)でP. gingivalis を30時間培 養した。
40% 濃度enriched BHI broth(A)で培養すると、100% 濃度で培養した時の 約半分の増殖量を示した。
41
2)A. actinomycetemcomitans cell extracts タンパク濃度によるP. gingivalis 増殖量の変化と経時的増殖量の変化
A. actinomycetemcomitans のcell extracts 中のタンパクによるP. gingivalis 増殖への影響について調べるため、A. actinomycetemcomitans のcell extractsを 添加した40% 濃度のenriched BHI broth(A)を用いてP. gingivalis を18時間 培養した。その結果、タンパク濃度依存的にP. gingivalis の増殖が促進された(図
9)。また50 μg /ml で増強効果がほぼピークを示したため、以降の実験はタンパ
ク濃度50 μg /ml になるようにcell extractsを添加して行った。
さらにP. gingivalis の増殖量の経時的変化について調べたところ、培養6時間
後からコントロールとの差が認められ、18時間後に最も有意差を認めた(図10)。 よって以降の実験は培養時間を18時間に設定して行った。
42
図 9 A. actinomycetemcomitans のcell extracts(CEs)タンパク濃度における P. gingivalis 増殖量の比較
各タンパク濃度のA. actinomycetemcomitans のcell extractsを添加した40% 濃 度のenriched BHI broth(A)にてP. gingivalis ATCC33277株を18時間培養し た。
A. actinomycetemcomitans cell extractsのタンパク濃度依存的にP. gingivalis の増殖量は増加し、50 μg /ml のタンパク濃度で増強効果がほぼピークを示した。
43
□
:40 % BHI■
:40 % BHI with 50 μg /ml CEs from A. actinomycetemcomitans* P < 0.01、** P < 0.05
図 10 各培養時間によるP. gingivalis 増殖量の経時的変化
タンパク濃度50 μg /ml A. actinomycetemcomitans のcell extracts(CEs)を添 加した40% enriched BHI broth(A)で6時間毎に培養を行った。
18時間後にコントロールとの間で最も有意差が認められた。
44
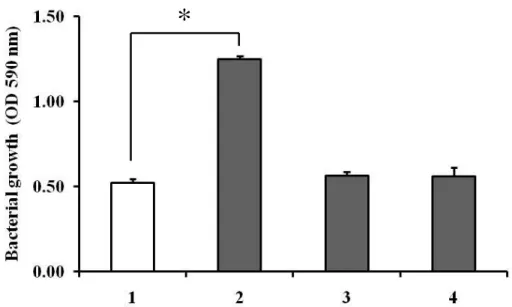
3)A. actinomycetemcomitans cell extracts中のタンパク変性によるP. gingivalis 増殖への影響
A. actinomycetemcomitans のcell extracts中のどのような成分がP. gingivalis の増殖促進に働いているのかを検討するため、あらかじめ熱処理(80℃、10分間)
したcell extracts を加えたところ、P. gingivalis の増殖促進効果は認められなくな った(図11)。また同じタンパク濃度になるようにBSAを加えても、P. gingivalis は増殖促進しなかったため、この増殖促進にはA. actinomycetemcomitans のcell
extracts 中の特別なタンパク構造が関与している可能性が示唆された。
45
*
P < 0.01図 11 A. actinomycetemcomitans のcell extracts(CEs)中のタンパク変性によ るP. gingivalis 増殖促進効果への影響
40% enriched BHI broth(A)で18時間培養を行った。
1:コントロール(40% enriched BHI broth(A)のみ)、 2:50 μg /ml A. actinomycetemcomitans CEs、
3:熱処理(80℃で10分間)した50 μg /ml A. actinomycetemcomitans CEs、
4:50 μg /ml BSA
コントロールと比較して、熱処理後のA. actinomycetemcomitans CEsと 50 μg /ml BSAではP. gingivalisの増殖促進を認めなかった。
*
46
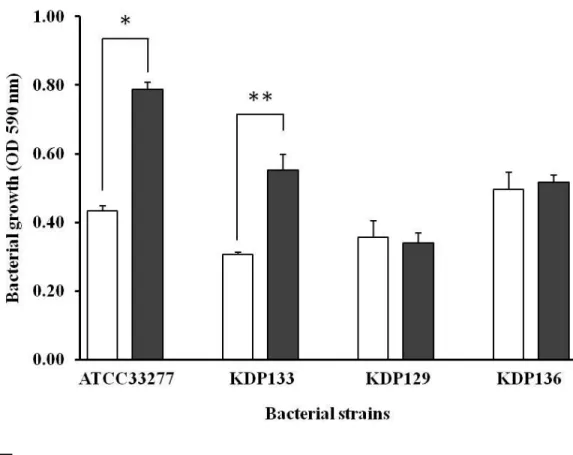
4)A. actinomycetemcomitans cell extractsによるP. gingivalis 増殖促進とジン ジパインの関与
A. actinomycetemcomitans がP. gingivalis の増殖を促進するメカニズムを調 べるために、ジンジパインの関与について検討した。各ジンジパイン変異株を用い てA. actinomycetemcomitans cell extractsによる増殖への影響を調べたところ、
KDP133(Rgp欠損株)は野生株と同じように増殖を促進したが、KDP129(Kgp
欠損株)とKDP136(Rgp・Kgp欠損株)では増殖促進しなかった(図12)。この ことからA. actinomycetemcomitans cell extracts中のタンパクの獲得にはKgpの 関与が示唆された。
47
□
:40% BHI■
:40% BHI with 50 μg /ml CEs from A. actinomycetemcomitans* P < 0.01、** P < 0.05
図 12 ジンジパイン変異株の増殖量の比較
50 μg /ml タンパク濃度のA. actinomycetemcomitans のcell extracts(CEs)を 添加した40% enriched BHI broth(A)で18時間培養を行った。
ATCC33277株(野生株)とKDP133株(Rgp欠損株)は有意に増殖促進を認
めたが、KDP129株(Kgp欠損株)とKDP136株(Rgp・Kgp欠損株)の両菌株 は増殖促進されなかった。
48
2‐4:考察
本章ではA. actinomycetemcomitans ATCC29523株のcell extractsを用いて、
A. actinomycetemcomitans との共培養下におけるP. gingivalis 増殖促進のメカ ニズムについて検討した。
P. gingivalis はタンパク質をペプチドやアミノ酸に分解し、取り込むことで栄養
源としているが(53‐55)、P. gingivalis の増殖促進に関わる細菌間相互作用にタ ンパクが関与するという報告はこれまでにほとんど得られていない。そこで、
enriched BHI broth中にA. actinomycetemcomitans のcell extractsを添加した ところ、そのタンパク濃度依存的に P. gingivalis の増殖を促進することが分かっ た(図9)。さらに、図10で得られた経時的なP. gingivalis 増殖量の変化は、図 7で示した混合培養時の P. gingivalis 細菌数の経時的増加曲線とよく似た傾向を 示しており、混合感染時においても同様の増殖促進効果が起きている可能性が示唆 された。
また、熱処理を行ったA. actinomycetemcomitans のcell extractsや同じタンパ ク濃度のBSAを添加した場合では、P. gingivalis の増殖を促進しなかったことか ら(図11)、A. actinomycetemcomitans のcell extracts 中のタンパク成分は本菌 の増殖において、単に栄養源として利用されているわけではなく、増殖促進因子と してシグナリングタンパクのような働きをしている可能性が示唆された。
ジンジパイン変異株への影響を調べたところ、Kgp 欠損株や Rgp・Kgp 欠損株 では増殖促進しなかったことから(図12)、A. actinomycetemcomitans の細胞構 成成分中のP. gingivalis 増殖促進因子の獲得には、Kgpが関与している可能性が
49
示唆された。Kgpは赤血球凝集やヘムの獲得などにも関与しているが、P. gingivalis がタンパク質をペプチドに分解する過程に必要であると考えられており、上記のメ カニズムの一つとして、Kgpのタンパク分解活性がA. actinomycetemcomitans の
cell extracts中の特別なタンパクを分解し、増殖促進因子として働くシグナリング
タンパク分子が作用することで本菌の増殖が促進される可能性が示唆された。
本実験で示されたA. actinomycetemcomitans によるP. gingivalis の増殖促進 効果は、歯周ポケット内における両菌の混合感染時、限られた栄養環境の中で P. gingivalis が優位に増殖することを示唆するものである。更に、P. gingivalis が 競合的優勢なバイオフィルム形成を行って、本菌の病原性をより発揮できる可能性 も考えられた。今後、増殖促進因子として働くA. actinomycetemcomitans の細胞 構成成分中のタンパクがどこに存在しているのか、どのようにして Kgp が働き作 用しているのか、また他の A. actinomycetemcomitans 菌株の cell extracts や
P. gingivalis 菌株においても同様の増殖促進効果が得られるか、等について検討し
ていく予定である。
50
総括
第一章ではP. gingivalis の単独バイオフィルム形成にジンジパイン特にRgpが 関与していること、またジンジパインがA. actinomycetemcomitans のバイオフィ ルムを剥離し、その形成を抑制することで P. gingivalis 優勢な複合バイオフィル ムを形成する可能性が示唆された。
第二章ではP. gingivalis のKgpがA. actinomycetemcomitans の細胞構成成分 の獲得に関わっており、本菌の増殖促進に働いている可能性が示唆された。この結 果からも、混合感染においては P. gingivalis 優勢な複合バイオフィルムを形成す る可能性が示唆された。
以上のことから、歯周動的治療後の歯周ポケット内において、P. gingivalis と A. actinomycetemcomitans の混合感染時では、Rgp およびKgp が付着、剥離、
増殖促進といった相互作用に関与し、P. gingivalis 優勢な複合バイオフィルムが形 成される可能性が示唆された。
51
謝辞
本研究は九州大学大学院歯学研究院口腔機能修復学講座歯周病学分野 前田勝 正教授の御指導のもとに行われたものであり、先生の御懇篤なる御指導と御助言に 深く感謝いたします。本研究を遂行するにあたり、終始御指導さらに本論文の御校 閲を賜わりました口腔機能修復学講座歯周病科助教 三浦真由美博士ならびに口 腔機能修復学講座歯周病科講師 藤瀬 修博士に謹んで御礼申し上げます。
最後になりましたが、本研究に御指導、御協力下さいました九州大学大学院歯学 研究院口腔機能修復学講座歯周病学分野の皆様に、心から感謝いたします。
52 引用論文
1. Winkelhoff. A. J., and Graaff. J. Microbiology in the management of destructive periodontal disease. J Clin Periodontol 1991; 18: 406-410.
2. Jeffcoat. M. K., and Reddy. M. S. Progression of probing attachment loss in adult periodontitis. J Periodontol 1991; 62: 185-189.
3. Darveau. R. P., Tanner. A., Page. R. C. The microbial challenge in periodontitis. Periodontology 2000 1997; 14: 12-32.
4. Marsh. P.D. Dental Plaque as a Microbial Biofilm. Caries Res 2004; 38:
204-211.
5. Stewart. P.S., Costerton. J.W. Antibiotic resistance of bacteria in biofilms.
The Lancet 2001; 358: 135-137.
6. Nguyen. K-A., DeCarlo. A. A., Paramaesvaran. M., Collyer. A. C., Langley.
B. D., and Hunter. N. Humoral responses to Porphyromonas gingivalis gingipain adhesin domains in subjects with chronic periodontitis. Infect Immun 2004; 72 : 1374-1382.
7. Haffajee. A. D., Socransky S. S. Microbial etiological agents of destructive periodontal disease. Periodontology 2000 1994; 5: 78-111.
8. Holt. S. C., Kesavalu. L., Walker. S., Genco. C. A. Virulence factors of Porphyromonas gingivalis. Periodontology 2000 1999; 20: 168-238.
9. Abe . N., Kadowaki. T., Okamoto. K., Nakayama. K., Ohishi. M., and Yamamoto. K. Biochemical and functional properties of Lysine-specific
53
cysteine proteinase (Lys-gingipain) as a virulence factor of Porphyromonas gingivalis in periodontal disease. J Biochem. 1998; 123: 305-312.
10. Mcgraw. W. T., Potempa. J., Farley. D., and Travis. J. Purification,
characterization, and sequence analysis of a potential virulence factor from Porphyromonas gingivalis, peptidylarginine deiminase. Infect Immun 1999; 67: 3248-3256.
11. Mooney. J., Adonogianaki. E., Riggio. M. P., Takahashi. K., Haerian. A., and Kinane. D. F. Initial serum antibody titer to Porphyromonas gingivalis influences development of antibody avidity and success of therapy for chronic Periodontitis. Infect Immun 1995; 63: 3411-3416.
12. Kuboniwa. M., Amano. A., Inaba. H., Hashino. E., Shizukuishi. S.
Homotypic biofilm structure of Porphyromonas gingivalis is affected by FimA type variations. Oral Microbiol Immunol 2009; 24: 260-263.
13. Lin. X., Wu. J., and Xie. H. Porphyromonas gingivalis minor fimbriae are required for cell-cell interactions. Infect Immun 2006; 74: 6011-6015.
14. Kuboniwa. M., Amano. A., Hashino. E., Yamamoto. Y., Inaba. H., Hamada N., Nakayama K., Tribble. G. D., Lamont. R. J., and Shizukuishi. S. Distinct roles of long/short fimbriae and gingipains in homotypic biofilm development by Porphyromonas gingivalis. BMC Microbiol 2009; 9:105-117.
15. Grenier. D., Roy. S., Chandad. F., Plamondon. P., Yoshioka. M., Nakayama.
K., and Mayrand. D. Effect of inactivation of the Arg-and/or Lys-gingipain gene on selected virulence and physiological properties of Porphyromonas
54
gingivalis. Infect Immun 2003; 71: 4742-4748.
16. Yamanaka. A., Kouchi. T., Kasai. K., Kato. T., Ishihara. K., Okuda K.
Inhibitory effect of cranberry polyphenol on biofilm formation and cysteine proteases of Porphyromonas gingivalis. J Periodont Res 2007; 42: 589-592.
17. Dickinson. D. P., Kubiniee. M. A., Yoshimura. F., and Genco. R. J. Molecular cloning sequencing of the gene encoding the fimbrial subunit protein of Bacteroides gingivalis. J Bacteriol 1988; 170: 1658-1665.
18. Amano. A. Molecular interaction of Porphyromonas gingivalis with host cells: implication for the microbial pathogenesis of periodontal disease. J Periodontol 2003; 74: 90-96.
19. DeCarlo. A. A., Paramaesvaran. M., Yun. P. L. W., Collyer. C., and Hunter. N.
Porphyrin-mediated binding to hemoglobin by the HA2 domain of cysteine proteinases (gingipains) and hemagglutinins from the periodontal pathogen Porphyromonas gingivalis. J Bacteriol 1999; 18: 3784-3791.
20. Nakayama. K. Construction and characterization of Arginine-specific cysteine proteinase (Arg-gingipain)-deficient mutants of Porphyromonas gingivalis. J Biol Chem 1995; 270: 23619-23626.
21. Okamoto. K., Nakayama. K., Kadowaki. T., Abe. N., Ratnayake. D. B., and Yamamoto. K. Involvement of a Lysine-specific cysteine proteinase in hemoglobin adsorption and heme accumulation by Porphyromonas gingivalis. J Biol Chem 1998; 273: 21225-21231.
22. Pavloff. N., Potempa. J. Pike. M. C., Prochazka. V., Kiefer. M. C., Travis. J.,
55
and Barr. P. J. Molecular cloning and structural characterization of the Arg-gingipain proteinase of Porphyromonas gingivalis. J Biol Chem 1995;
270: 1007-1010.
23. Ito. R., Ishihara K., Shoji. M., Nakayama. K., Okuda K.
Hemagglutinin/Adhesin domains of Porphyromonas gingivalis play key roles in coaggregation with Treponema denticola. FEMS Immunol Med Microbiol 2010; 60: 251-260.
24. Asikainen. S., Chen. C., Oral ecology and person-to-person transmission of Actinobacillus actinomycetemcomitans and Porphyromonas gingivalis.
Periodontol 2000 1999; 20: 65-81.
25. Kaplan. J. B., Perry. M. B., MacLean. L. L., Furgang. D., Wilson. M. E., Fine.
D. H. Structural and genetic analyses of O polysaccharide from
Actinobacillus actinomycetemcomitans serotype f. Infect Immun 2001; 69:
5375-5384.
26. Kachlany. S. C., Planet. P. J., DeSalle. R., Fine. D. H., Figurski. D. H. Genes for tight adherence of Actinobacillus actinomycetemcomitans: from plaque to plague to pond scum. Trends Microbiol 2001; 9: 429-437.
27. Wang. Y., Chen. C. Mutation analysis of the flp operon in Actinobacillus actinomycetemcomitans. Gene 2005; 351: 61-71.
28. Wennström. J. L., Dahlén. G., Svensson. J., Nyman. S. Actinobacillus actinomycetemcomitans, Bacteroides gingivalis and Bacteroides intermedius: predictors of attachment loss? Oral Microbiol Immunol 1987;
56 2: 158-193.
29. Lavanchy. D. L., Bickel. M., Baehni. P. C. The effect of plaque control after scaling and root planing on the subgingival microflora in human periodontitis. J Clin Periodontol 1987; 14: 295-299.
30. Ebersole. J. L., Cappelli. D., Steffen. M. J., Willmann. D. E., O’Dell. D. S.
Host response assessment in recurring periodontitis. J Clin Periodontol 1996; 23: 258-262.
31. Ehmke. B., Beikler. T., Haubitz. I., Karch. H., Flemming. T. F. Multifactorial assessment of predictors for prevention of periodontal disease progression.
Clin Oral Investig 2003; 7: 217-221.
32. Saito. Y., Fujii. R., Nakagawa. K-I., Kuramitsu. H. K., Okuda. K., Ishihara.
K. Stimulation of Fusobacterium nucleatum biofilm formation by Porphyromonas gingivalis. Oral Microbiol Immunol 2008; 23: 1-6.
33. Marsh. P. D. Dental plaque as a biofilm and a microbial community implications for health and disease. BMC Oral Health 2006; 6: S14.
34. Kolenbrander. P. E., Andersen. R. N., Blehert. D. S., Egland. P. G., Foster. J.
S., and Palmer. R. J. Jr. Communication among oral bacteria. Microbiol Mol Biol Rev 2002; 66: 486-505.
35. Kuboniwa. M., Lamont. R. J. Subgingival biofilm formation. Periodontology 2000 2010; 52: 38-52.
57
36. Periasamy. S., and Kolenbrander. P. E. Mutualistic biofilm communities develop with Porphyromonas gingivalis and initial, early, and late
colonizers of enamel. J Bacteriol 2009; 191: 6804–6811.
37. Suzuki. N., Nakano. Y., Kiyour. Y. Characterizing the specific coaggregation between Actinobacillus actinomycetemcomitans serptype c strains and Porphyromonas gingivalis ATCC33277. Oral Microbiol Immunol 2006; 21:
385-391.
38. Ali. R. W., Lie. T., Skaug. N. Early effects of periodontal therapy on the detection frequency of four putative periodontal pathogens in adults. J Periodontol 1992; 63: 540-547.
39. Riggio. M. P., Macfarlane. T. W., Mackenzie. D., Lennon. A., Smith. A. J., Kinane. D. Comparison of polymerase chain reaction and culture methods for detection of Actinobacillus actinomycetemcomitans and Porphyromonas gingivalis in subgingival plaque samples. J Periodontal Res 1996: 31: 496–
501.
40. Miura. M., Hamachi. T., Fujise. O., Maeda. K. The prevalence and pathogenic differences of Porphyromonas gingivalis fimA genotypes in patients with aggressive periodontitis. J Periodontal Res 2005; 40: 147-152.
41. Lewis. J. P., Macrina. F. L. IS195, an insertion sequence-like element associated with protease genes in Porphyromonas gingivalis. Infect Immun 1998; 66: 3035-3042.
42. Curtis. M. A., Aduse-Opoku. J., Slaney. J. M., rangarajan. M., Booth. V.,