Prolyl Hydroxylaseの投与がヒラメ筋の筋線維組成と代謝酵素活性に及ぼす影響
6
0
0
全文
(2) ?t%i@k%%Y%S ( $ % ? ~ %61& ~ ~ ) %2 3 Journal of Hokkaido University of Education (Natural Sciences) Vol. 61, No.2. 2 February, 2011. %i%23+. Effects of Prolyl Hydroxylase Inhibitor on Fiber Type Composition and Metabolic Enzyme Activities in Rat Soleus Muscle SUZUKI Junichi Laboratory of Exercise Physiology, Health and Sports Sciences, Course of Sports Education, Department of Education, Iwamizawa Campus, Hokkaido University of Education. Prolyl Hydroxylase O@%iFk 7 3 fi%Ofi%#~$E&LE 2 f'Z&f@%%'r%KE IT-j-83B $. *. j$. -. ABSTRACT A family of 02-dependent prolyl hydroxylase domain-containing exzymes has been identified as a cellular oxygen sensing mechanism. Reduced prolyl hydroxylase activity facilitates an accumulation of hypoxiainducible factor (HIF)-la. The present study was designed to examine the effects of prolyl hydroxylase inhibitor, ethyl-3,4 dihydroxybenzoate (EDHB), administration on fiber type distribution and metabolic enzyme activities in rat soleus muscle. Sixteen (12 weeks old) female Wistar rats were used in the present study. Rats received interperioneal injection of 100 mgkg EDHB (n=8) or vehicle (n=8), dirnethylsulfoxide, three consecutive days per week lasted for 3 weeks. According to irnmunohistocheical identification of type I1 fiber, EDHB treatment significantly increased a proportion of type I1 fiber and concomitantly decreased that of type I fiber (R0.05). Total number of muscle fibers did not change after the treatment. Hexokinase activity was significantly enhanced (P<0.05), whereas the activities of citerate synthase, B-hydroxyacyl-CoA dehydrogenase, lactate dehydrogenase, and phosphofructokinase did not change after EDHB treatment. The present results suggest that prolyl hydroxylase inhibitor enhances type I1 fiber distribution and glycolytic metabolism in the rat soleus muscle. [Keywords] hypoxia inducible factor, prolyl hydroxylase inhibitor, skeletal muscle, fiber type, metabolic enzyme INTRODUCTION Inadequate oxygen supply can lead to cellular dysfunction and even cell death in aerobic organisms. However, exposure to the oxygen must be limited because of damaging effects of reactive oxygen species on cellular macromolecules. Thus the major mammalian physiological systems has complex homeostatic mechanisms that are designed to maintain the oxygen concentration with a narrow range. Recently a family of oxygen-dependent prolyl hydroxylase domain-containing enzymes (PHD) have been identified as a cellular oxygen-sensing mechanism. (Ivan et al. 2001, Jaakkola et al. 2001, Bruick & McKnight 2001). Hypoxia inducible factors (HIFs) are transcription factors whose activity is regulated through oxygen-dependent proteolysis of the a subunit. HIF-la is constitutively expressed and rapidly degraded under normoxic conditions. The first step in degradation is hydroxylation of a proline residue by one of three HIFPHDs that requires oxygen, iron and 2-oxyglutarate as cofactors (Ivan et al. 2001). Under hypoxic conditions, PHDs are inactivated, and HIF-la accumulates and translocates to the nucleus where it activates HIF.
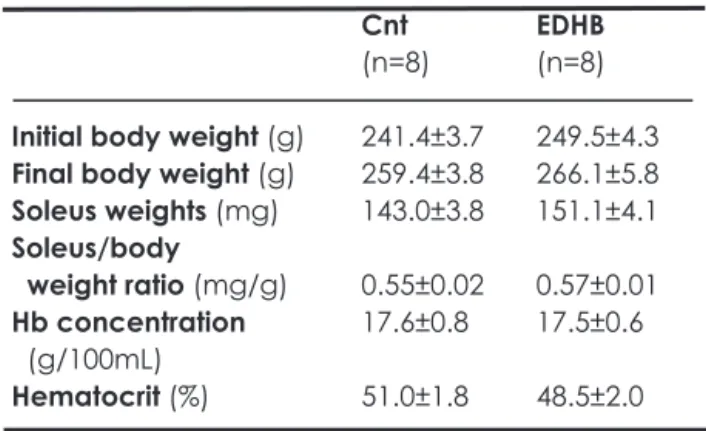
(3) responsive genes (e.g. erythropoietin, glucose transporter (GLUT)-1 and vascular endotherial growth factor(VEGF)). The PHD inhibition with ethyl-3,4-dihydroxybenzoate (EDHB) induced the GLUT-1, nitric oxide synthase (NOS)-2, and the antioxydant heme oxygenase in cardiomyocytes (Wright et al. 2003). EDHB administration for 7 days markedly enhanced endurance running performance under hypoxic (8%O2) environment (Kasignesan et al. 2007). Many studies have been conducted on the adaptation of muscle fiber types in skeletal muscle in response to hypoxia. Recent finding indicates that hypoxic exposure may cause a type-shift of muscle fibers from slowtwitch oxidative (SO) to fast-twitch oxidative glycolytic (FOG) in the soleus muscle (Itoh et al. 1988). Ishihara et al. (1995) reported that hypoxic exposure does not change the muscle, but rather inhibits the type-shift of muscle fibers from FOG to SO during postnatal growth in the rat soleus muscle. In the present study, experiments were designed to examine whether PHD inhibition with EDHB treatment affects fiber-type distribution and metabolic enzymes in soleus muscle. The present results demonstrate that EDHB administration enhanced distribution of type I1 fiber and glycolytic enzyme activity. MATERIALS AND METHODS This study was approved by the Animal Care and Use Committee of Hokkaido University of Education and performed in accordance with the "Guiding Principles for the Care and Use of Animals in the Field of Physiological Sciences" of the Physiological Society of Japan. Animals, experimental conditions and muscle samples Sixteen (10-week-old) female Wistar rats were purchased from Charles River Japan Inc. (Tokyo, Japan). After the rats were kept for 2 weeks to allow adaptation to the new environment, they were randomly divided into non-treated (Cnt, n=8) and EDHB-treated (ED, n=8) groups. The rats received interperioneal injection of 100 mglkg EDHB or vehicle, dimethylsulfoxide, three consecutive days per week lasted for 3 weeks. All rats were given commercial laboratory chow (CE-2, Clea Japan Inc.) and tap water ad libitum. Under light anesthesia with ether, the rats were anaesthetized with alpha-chloralose (0.06 g K g i. p.) and urethane (0.7 gikg i. p.). A toe pinch response was used to validate adequate anesthesia. Then, the left soleus. (SOL) muscles were excised and weighed. The muscles were fixed at the length measured when the knee joint was maximally extended and the tibiotarsal joint was fixed at 90 deg. The tissues were placed in embedding medium, an Optimal Cutting Temperature (O.C.T.) compound (Sakura Finetechnical, Tokyo, Japan), and fiozen rapidly in isopentane cooled to its melting point (-160°C) with liquid nitrogen. The right SOL muscles were excised and frozen rapidly liquid nitrogen, and used for biochemical analyses. All samples were treated in a similar fashion. The tissue samples were stored at 80°Cuntil analysis. Histological analyses For immunodetection of type I1 fibers, tissue cross-sections, 6-pm-thick, were obtained using a cryotome (CM-1500; Leica Japan, Tokyo, Japan) at -20°C. The sections were fixed in cold acetone (4°C) for 5 min and then rinsed for 5min in phosphate-buffered saline with 0.05% Tween-20 (PBST). Endogenous peroxide activity was blocked by incubation in 0.3% hydrogen peroxide in methanol for 30 min and then rinsed with PBST. The sections were incubated with primary antibody, monoclonal antiskeletal myosin (fast) clone MY-32 (Sigma, M4276). The primary antibody reaction was amplified with a Histofine MAX-PO (MULTI) (Nichirei, Japan). The sections were exposed to 3,3-diarninobenzidine (DAB) to obtain a brown reaction product using a DAB staining kit (Nichirei). The images of incubated sections were digitized using a digital microscope camera (PDMC ley Polaroid, USA) attached to a light microscope (BX-50, Olympus, Tokyo, Japan) and were stored on computer disk. Non-overlapping microscopic fields were selected fiom each muscle sample. During the measurements, the observer was blind as to the source (groups) of each slide. Biochemical analyses Tissue homogenates were -. Table 1 Body and organ weights. Initial body weight (g) Final body weight (g) Soleus weights (mg) Soleus/body weight ratio (mglg) Hb concentration ( g l lOOmL) Hematocrit (%). Values are means k SE.. Cnt (n=8). EDHB (n=8). 241.4k3.7 259.4k3.8 143.0k3.8. 249.5k4.3 266.1k5.8 151.1k4.1. 0.55k0.02 17.6k0.8. 0.57k0.01 17.5*0.6. 51.0k1.8. 48.5*2.0.
(4) Prolyl Hydroxylase Inhibitor and Muscle Function. EDHB. Figure 1 Irnmunohistochemical identification of type I (light) and type I1 (dark) fibers in the soleus muscle. Horizontal bar represents 100pm.. obtained fiom approximately 50 mg of frozen tissue homogenized with three interrupted 15-s bursts with a Polytron homogenizer in ice-cold medium (10 mM HEPES buffer, pH 7.3; 0.1% Triton X-100; 11.5% (wlv) sucrose; and 5% (vlv) protease inhibitor cocktail (P2714, Sigma)). After centrifugation at 1,500 xg for 10 min at 4OC, the supernatant was used for enzyme activity analysis. Total protein was measured using PRO-MEASURE protein measurement solution (iNtRON Biotechnology, Korea). The activities of Bhydroxyacyl-CoA-dehydrogenase (HAD) and lactate dehydrogenase (LDH) were assayed according to the method of Bass et al. (1969). The activities of citrate synthase (CS) was determined according to the method of Srere (1969). Hexisokinase (HK) and phosphofiuctokinase (PFK) were determined according to the method of Passonneau & Lowry (1993). All measurements were carried out at 25OC with a. 0%. Cnt. spectrophotometer (U-2001, Hitachi Co., Tokyo, Japan). Statistical Analyses All values are expressed as SE. Differences between the groups were means analyzed using Student's t-test. Differences were considered statistically significant at P<0.05.. *. RESULTS Mean body mass and muscle mass at the end of experiment was not significantly different between the groups (Table 1). Intermittent administration of EDHB did not affect hematocrit and Hb concentration. To investigate the effects of EDHB treatent on fiber type distribution, immunohistochemical identification of type I1 fiber was performed on tissue cross-sections. Figure 1 shows representative micrographic images of sections that demonstrate type I (light) and type I1 (dark) fibers. EDHB treatment significantly increased the proportion of type I1 fibers and concomitantly decreased that of type I fibers (P<0.05; Fig. 2). Total number of muscle fibers did not change after the treatment (Fig. 3).. ED. Figure 2 Changes in fiber-type composition (%) after EDHB treatment in the soleus muscle. * ,significantly different from the Cnt group at P<0.05.. Cnt. ED. Figure 3 Total number of muscle fibers in the soleus muscle..
(5) SUZUKI Junichi. 0.150. -. 0.113. -. 0.075. -. 0.038. -. 0. -. T. Figure 3 Enzyme activity after EDHB treatment in the soleus muscle. *, significantly different from the Cnt group at P<O.O5.. HAD. PFK Thus, EDHB treatment enhanced type I1 fiber distribution in the SOL muscle. Metabolic enzyme activities were determined to investigate the effects of EDHB administration on muscle metabolism (Fig. 4). EDHB treatment significantly increased the HK activity, while it did not affect the CS, HAD, PFK and LDH activities. Therefore, EDHB treatment facilitated glycolytic metabolism in the SOL muscle.. DISCUSSION The present study demonstrated that proryl hydroxylase inhibition with EDHB enhanced hexokinase activity in the soleus muscle (Fig 3). Since hexokinase is one of the HIF-1 target genes (Iyer et al. 1998), it is confirmed that the present administration of EDHB stimulated the PHD oxygen-sensing pathway, i.e., inhibiting proryl hydroxylase and stabilizing HIF-la in skeletal muscle. HIF-la activates the expression of genes that mediate a myriad of compensatory responses that include the upregulation of anaerobic glycolytic metabolism. The findings that PHD hydroxylating activity is the signaltransduction mechanism that senses oxygen concentrations provided the rationale for the use of inhibitors of the PHDs to selectively activate cellular. LDH. hypoxic response in the absence of metabolic perturbation with their attendant signalling responses. Several studies have reported that PHD inhibitors recapitulate various cellular responses to hypoxia. These include HIF- 1a stabilization, the induction of HIF- 1a targeted genes, stimulation of angiogenesis, and protection against metabolic stress (Warnecke et al. 2003, Wright et al. 2003, Asikainen et al. 2005, Sridharan et al. 2006, Chen et al. 2009). In the present study, proryl hydroxylase inhibition with EDHB increased distribution of type I1 fiber in the SOL. Although many studies have been conducted to clarify the adaptation of muscle fiber type to hypoxia, conflict results have been obtained. Sillau & Banchero (1977) reported no change in the fiber type distribution in the soleus muscle of developing rats after hypoxic exposure at a simulated altitude of 5,100m for 6 weeks. In contrast, An increase in the percentage of typeIIAB fibers (intermediate type between typeIIA and type IIB) was observed in the extensor digitorurn longus muscle and in the deep portion of plantaris muscle in rats after 14 weeks of hypoxic (4,000111) exposure (Bigard et al. 1991). These controversial results may be due to the difference of muscles, animal species, age of animals andlor hypoxic conditions..
(6) Prolyl Hydroxylase Inhibitor and Muscle Function. Ishihara et al. (1995) examined the effect of hypoxic exposure, equivalent to 4,000m, on the fiber type of soleus muscle using developing rats. A growth-related type shift of muscle fibers from FOG to S O was found in the normoxic rats (35% and 19% of FOG fibers at 5 and 12 weeks of age, respectively). However, the hypoxic rats at 12 weeks of age had 29% of FOG fibers. There is no difference in total number of fibers between normoxic and hypoxic rats. Moreover, hypoxic exposure did not cause a de novo synthesis of type IId (type IIx) myosin heavy chain isoform. These authors concluded that hypoxic exposure inhibits the growth-related type shift of fibers from FOG to SO in the rat soleus muscle.. In the present study, significant increase in the percentage of type I1 fiber (Fig. 2) with no change in total number of fibers (Fig. 3) after intermittent EDHB treatment for 3 weeks. Thus, the present EDHB administration may inhibit the growth-related type shift of fibers in soleus muscle. The results of the present study suggest that inhibition of prolyl hydroxylase and accumulation of HIF-la plays a major role in inhibiting growth-related fiber type shift during hypoxic exposure. In the present study, rats at 12 weeks of age were subjected to EDHB treatment for 3 weeks. After 5-week exposure to simulated hypoxia (4,000m) from 5 and 10 weeks of age, the growth-related type shift of muscle fibers from FOG to SO was inhibited regardless of age (Ishihara et al. 1995). However, the inhibition did not occur when the hypoxic exposure was started at 15 weeks of age, because the postnatal type shift of muscle fibers was completed (Ishihara et al. 1995). I n summary, i n t e r m i t t e n t E D H B t r e a t m e n t significantly enhanced a proportion of type I1 fiber and hexokinase activity in soleus muscle. The present results suggest that the present administration of EDHB stimulates the PHD oxygen-sensing pathway, and inhibits growth-related fiber type shift in rat soleus muscle.. ACKNOWLEDGEMENT This work was supported by the JSPS KAKENHI (20500571). REFERENCES Asikainen TM, Ahmad A, Schneider BK, Ho WB, Arend M, Brenner M, Giinzler V, White CW (2005) Stimulation of HIF-lalpha, HIF-2alpha, and VEGF by prolyl 4hydroxylase inhibition in human lung endothelial and epithelial cells. Free Radic Biol Med 38,1002-1013 Bass A, Brdiczka D, Eyer P, Hofer S, Pette D (1969) Metabolic differentiation of distinct muscle types at the level of enzymatic organization. Eur J Biochem. 10:198-206 Bigard AX, Brunet A, Guezennec CY, Monod H (1991) Skeletal muscle changes after endurance training at high a1titude.J Appl Physiol71,2114-2121 Bruick RK, McKnight SL. (2001) A conserved family of prolyl-4-hydroxylases that modify HIF. Science 294, 1337-1340 Ishihara A, Itoh K, Hirofuji C, Hayashi H (1995) Effects of hypobaric hypoxia on histochemical fibre-type composition and myosin heavy chain isofonn component in the rat soleus muscle. Pflugers Arch 429:601-606 Itoh K, Itoh M, Taguchi S, Ishihara A (1988) Effects of hypobaric-hypoxia on the total number and histochemical properties of the soleus muscle fibers and motoneurons in the rat. J Physiol SOCJpn 50, 163-168 Ivan M, Kondo K, Yang H, Kim W, Valiando J, Ohh M, Salic A, Asara JM, Lane WS, Kaelin WG Jr. (2001) HIFalpha targeted for VHL-mediated destruction by proline hydroxylation: implications for 0 2 sensing. Science 292,464-468 Iyer NV, Kotch LE, Agani F, Leung SW, Laughner E, Wenger RH, Gassmann M, Gearhart JD, Lawler AM, Yu AY, Semenza GL (1998) Cellular and developmental control of 0 2 homeostasis by hypoxia-inducible factor 1 alpha. Genes Dev 12,149-162 Jaakkola P, Mole DR, Tian YM, Wilson MI, Gielbert J, Gaskell SJ, Kriegsheim Av, Hebestreit HF, Mukherji M, Schofield CJ, Maxwell PH, Pugh CW, Ratcliffe PJ. (2001) Targeting of HIF-alpha to the von Hippel-Lindau ubiquitylation complex by 02-regulated prolyl hydroxylation. Science 292,468-472. Kasiganesan H, Sridharan V, Wright G (2007) Prolyl hydroxylase inhibitor treatment confers whole-animal hypoxia tolerance. Acta Physiol (Oxf) 190,163-169 Passonneau JV and Lowry OH (1993) Enzymatic analysis: A practical guide, Humana press, New Jersey. Sillau AH, Banchero N (1979) Effect of hypoxia on the capillarity of guinea pig skeletal muscle. Proc Soc Exp Biol Med 160,368-373 Srere PA (1969) ditrate synthase. Meth Enzyrn 13:3-5 Sridharan V, Guichard J, Bailey RM, Kasiganesan H, Beeson C, Wright GL (2007) The prolyl hydroxylase oxygensensing pathway is cytoprotective and allows maintenance of mitochondrial membrane potential during metabolic inhibition. Am J Physiol Cell Physiol292, C719-C728 Warnecke C, Griethe W, Weidemann A, Jiirgensen JS, Willam C, Bachmann S, Ivashchenko Y, Wagner I, Frei U, Wiesener M, Eckardt KU (2003) Activation of the hypoxia-inducible factor-pathway and stimulation of angiogenesis by application of prolyl hydroxylase inhibitors. FASEB J 17,1186-1188 Wright G, Higgin JJ, Raines RT, Steenbergen C, Murphy E. (2003) Activation of the prolyl hydroxylase oxygen-sensor results in induction of GLUT1, heme oxygenase-1, and nitric-oxide synthase proteins and confers protection from metabolic inhibition to cardiomyocytes. J Biol Chem 278,20235-20239 Chen X, Wan C, Rarnaswamy G, Mavalli M, Wang Y, Duvall CL, Deng LF, Guldberg RE, Eberhart A, Clemens TL, Gilbert SR (2009) Prolyl hydroxylase inhibitors increase neoangiogenesis and callus formation following femur fracture in mice. J Orthop Res DOI, 10.1002/jor.20886. (%RR@lEBB).
(7)
図

関連したドキュメント
This study was therefore designed to examine the induc- tion of CYP2E1 in genetically obese Zucker rats fed a normal diet (OB) and its effect on the disposition kinetics of CZX and
小田25)は「デトラヨ■一ドフエノールフタレンナ
そこでこの薬物によるラット骨格筋の速筋(長指伸筋:EDL)と遅筋(ヒラメ筋:SOL)における特異
[r]
Amount of Remuneration, etc. The Company does not pay to Directors who concurrently serve as Executive Officer the remuneration paid to Directors. Therefore, “Number of Persons”
いられる。ボディメカニクスとは、人間の骨格や
敷地と火山の 距離から,溶 岩流が発電所 に影響を及ぼ す可能性はな
敷地と火山の 距離から,溶 岩流が発電所 に影響を及ぼ す可能性はな
