1 はじめに 人間活動による生息地の縮小・分断化は多くの生物の個体数を減少させている。なかで も著しく個体数を減らしたものは、希少種や絶滅危惧種と呼ばれ、その保全が急がれてい る。一般に希少種の有効な保全策を考える上では、複数の空間スケールから集団構造を 把握し、その成因を探る必要がある。また、生息地の個体数を向上させる具体的方策を 明らかにするには、生息地の環境条件を特定する必要がある。サドガエルは2012 年に 新種記載されたばかりの佐渡島固有種である(Sekiya et al 2012)。そのため、本種はト キと並んで佐渡島における重要な保全対象種であり、環境保全型農業の指標種としての 価値も高い。本種の生息地は、佐渡島中央部の平野部(国中平野)や丘陵部で多く発見されて いるが、分布パターンは一様ではなく不連続になっている(図 1)。こうした分布パターンは、 保全策を考える上で個体群の空間構造を考慮する必要があることを示唆している。また、 本種の生活史は他の多くの日本産カエルと異なり、オタマジャクシで一年を過ごし、翌 年変態・上陸する。そのため、周年を通した水の存在が生息の条件と考えられる。本研 究では、サドガエルを対象に、遺伝子解析から地域個体群の特定と地域個体群内の生息 地の連結性を評価し、集団構造の推定を行うとともに、分布調査から個体数を制約する 環境要因を推定する。
2 材料と方法 1. 遺伝子解析 サンプル採取 サドガエルの生息が確認されている佐渡島の国中平野を中心に、2010、2012、2013、2014 年の6 月(繁殖期)および 2014 年の 8 月に 29 集団 481 個体から遺伝子サンプルを採取した(図 2、表 1)。そのうち、90 個体は宇留間ら(2010)が採取したものである。成体 461 体から指 の先を、幼体20 体からは、尾の先を 5 ㎜ほど採取し、エタノールにつけて保管した。すべ ての個体はサンプル採取後、捕獲した場所に放した。集団のサンプルサイズは5 から 35 の 範囲である(表 1)。 マイクロサテライト座位の解析
各個体の組織からQIAGEN 社の DNeasy® Blood & Tissue Handbook と FAVORGEN 社のTissue Genomic DNA Extraction Mini Kit を用いて DNA を抽出した。マイクロサテ ライト座位は、(Lian et al 2006) の手法を用い、7 座位(4_109、4_114、6_167、6_234、 6_603、6_571、4_472)を新たに開発した。ポリメラーゼ連鎖反応(Polymerase Chain Reaction: PCR)では、プライマーミックスを混合した 4.5μl の反応溶液に 0.5μl の各個体 の DNA を加えた。反応溶液は 1 サンプルにつき、プライマーミックス 0.5μl のほか、 Type-it® Microsatellite PCR Kit を使用し、Type-it Multiplex PCR Master Mix, 2x を 2.5 μl、Q-Solution®, 5x を 0.5μl、反応溶液の量を調整する H2O を 1.0μl 含んでいる。 PCR
機は、TaKaRa PCR Thermal Cycler DiceTM Version 3 Model TP650 を用い、マイクロサ
テライトDNA は Applied Biosystems 3130xl Genetic Analyzers を用いて断片長を解析し た。
集団内の遺伝的多様性については、解析対象の集団内で任意交配が行われていることを 仮定したときに期待されるヘテロ接合個体の割合を示すヘテロ接合度の期待値(He) (井鷺 &陶山 2013)を用いて評価した。HeはGenAlEx 6.2 (Peakall and Smouse 2006)を用いて
3 算出した。
遺伝子を用いた空間構造の解析には、2 つの手法を用いた。はじめに、遺伝構造の差異か ら地域個体群、すなわち保全管理単位を特定するため、STRUCTURE v. 2.3.4 (Pritchard et al 2000)で解析を行った。次に、集団数が多い国中平野の 22 局所個体群(生息地)を対象に、 生息地のネットワーク構造を推定した。Population Graph (Dyer and Nason 2004) を用い て推定した。 2. 個体数情報の解析 個体数調査 佐渡島の国仲平野を中心とした水田を対象に、水田3 から 9 枚をひとつの調査地とし て42 か所の水田で調査を行った(図 3)。サドガエルは水田の畔および水田内の畔付近で おもに観察されるため、2014 年の繁殖期のピークである 6 月に、上旬と下旬でそれぞ れ成体の個体数調査を行った。調査は、昼間に水田の畔を歩き(平均 1779.8m (730.4m ∼3461.3m))、畔や水田の縁から水田内に飛び込む成体の個体数を目視で記録した。 環境要因 個体数調査時、および10 月に、局所要因として、各調査水田で水田に隣接している 水路の深さ(cm)、排水路のタイプ(土もしくはコンクリート)、水田の湛水面積率、江の 割合、畦の草丈を計測した。水路の深さは、水田1 枚ごとに計測し、浅い水路であれば レフュジアとして利用できると考え、水田群内で最も浅い場所の値を用いた。排水路の タイプについては、調査水田群内の水路に隣接する排水路の材質を土かコンクリートか に分けて記録した。水田の水環境をあらわす指標として水田の湛水面積率、水田群内に おける江を設置してある水田の割合を用いており、目視で記録した。湛水面積率は水田 ごとに記録したものを平均して水田群の湛水面積率とした。また、湛水期にも陸域を維
4
持する畦の微環境の指標として、畦の草丈を水田ごとに6 か所測定し、平均して利用し た。
GIS Analysis
景観要因として、地理情報システム(Arc GIS)を用いて、各水田群の地形的湿潤指標 (Topographic wetness index :TWI)、森林率を評価し、各生息地間の距離から連結 性を推定した。TWI は、国土地理院で得られる 50mメッシュの標高データをラスター データに変換し、1 つのセルの大きさを 66m として算出した。TWI は斜面位置によっ て生じる土壌水分傾度を表す指標である (Sørensen and Seibert 2007) 。森林率は、各 水田群の中心から、宇留間ら(2012)の結果を元に、サドガエルで最も AIC が低かった 100m の円形バッファを発生させ、その内に含まれる森林面積の割合とした。空間自己 相関の効果を考慮するため、生息地間の連結性を先行研究(Hanskii 1994)に見られる式 を用いて算出した。 統計解析 各調査地のサドガエル個体数を目的変数とした一般化線形モデルで解析を行った。不 在データが多かったため(42 か所中 17 か所)、ゼロ過剰モデル(負の二項分布)を用いて いる(Martin et al 2005) 。また、密度に与える影響を推定するため、生息地内の調査歩 行距離を常にオフセット項に含んでいる。説明変数に水路の深さ、水田の湛水面積率、 江の割合、凹凸度、森林率、生息地間の連結性を用いた。 解析では、まず、すべての変数の有無を考えた総当たりのモデル選択を AIC 基準で 行った。次に、各説明変数の評価に、各モデルのAkaike weight とパラメータ推定を用 いてモデル平均を行った。そして、それらのパラメータ推定の 95%CI(信頼区間)から、 影響している環境要因を決定した。 GLM に関しては R 3.0.2(R core Team 2013)を使用した。
5 結果 1. マイクロサテライト座位の解析 (1) STRUCTURE 解析による遺伝構造 すべての個体を用いてSTRUCTURE 解析を行った結果、lnP(D)は K=1 から K=5 に かけて増加した (図 4A)。また、⊿K では K=5 のとき最大となった(図 4B)。また、K=6 以降ではクラスター数の増加による遺伝構造の情報に変化がほとんどないため(図 5)、 K=5 が最適なクラスターであるとした。遺伝構造の成因を考えるため、K=2 から K=5 にかけての遺伝構造の変化を調べた。まず、K=2 の場合、一方のクラスターに佐渡島西 部および東部に位置する個体群が属し、中央の国仲平野の個体群は2 つのクラスターか らなっていた。一方、K=5 の場合、東部の個体群が他の個体群と明らかに異なるクラス ターに属していた。西部の個体群は、ひとつのクラスターのみに属し、国仲平野の個体 群は2 つのクラスターが混合する結果となった。 (2) 各生息地の遺伝的多様性 マイクロサテライト7 座位において、24 集団 373 個体すべてで遺伝的多型が検出さ れた。集団内の遺伝的多様性を示すヘテロ接合度期待値(He)は表 1 のようになった。 He が最も小さいのは MK と RO の 0.334 であり、最も大きいのは US の 0.593 である。 表1 から、西部の 2 集団と東部の 3 集団においては、遺伝的多様度が低いのがわかる。 (3) 生息地間の遺伝的ネットワーク Population Graph をもとに、最大の地域個体群である国仲グループ(22 局所個体群、 もしくは生息地)における、生息地の遺伝的なネットワークを作成した(図 6、図 7)。図 7 から、国仲平野においては、完全に孤立している局所個体群はみられなかった。
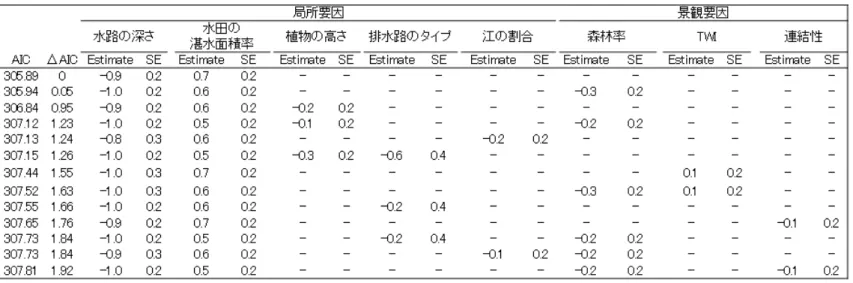
6 2. 個体数に影響を与える環境条件の推定 個体が記録された水田は42 調査地のうち、25 か所であった。モデル選択の結果、上 旬・下旬ともに⊿AIC<2 のすべてのモデルで水路の深さと水田の湛水面積率が選択さ れ、水路の深さに負の効果が、湛水面積率に正の効果がみられた(表 2)。他の説明要因 は、⊿AIC が 2 以内のモデルに稀にしか含まれなかった。 AIC を用いたモデル平均では、上旬・下旬のモデルで水路の深さと水田の湛水面積率 の推定値の値が大きく、また推定値の95%信頼区間が 0 をまたがなかった(図 8)。他の 説明要因は0 をまたいでいる(図 8)。したがって、水路の深さはサドガエルの局所個体 群密度に負の効果を、湛水面積率に正の効果を与えていたが、水路がコンクリートかど うかといったことや、江の割合、景観要因は効果を与えていなかった。 これらの要因との散布図を描いた結果、上旬・下旬ともに水路の深さにおいては、 30cm を境に観察された個体数は減少していた(図 9)。湛水面積率は上旬・下旬ともに湛 水面積率が高いほどサドガエル個体数が多かった (図 9)。その他の要因では、個体数と の間に傾向は見られなかった。
7 考察 1. 遺伝子による空間構造の解析 (1) 地域個体群の特定 佐渡島東部に位置する個体群(MK、RO、SD)と西部の個体群(OG、TS)の He の平 均は、佐渡島中央にある国仲平野のサドガエル個体群のHe の平均に比べてそれぞ 小さく、これらの個体群は個体群サイズが小さいことから、ボトルネックの影響を 受けていると考えられる。地理的距離や地形も考慮すると、サドガエルは東部・西 部・国仲平野の3 つのグループに分けられると言える。 STRUCTURE 解析の結果、サドガエルは遺伝的に 3 つのクラスターに分けられ た(K=5、図 5)。東部の個体群は固有のクラスターに属していたため、独立したグ ループとして考えられる。西部と国仲の局所個体群は共通のクラスターに属してい たが、西部の局所個体群がほぼ単一のクラスターにのみ属し、国仲平野の個体のみ が属すクラスターがあることから、サドガエルは東部、西部、国仲平野の3 つの地 域個体群に分けるのが妥当であると考えられる。地域個体群間の遺伝構造の差異は、 山地地形と汽水湖を挟んでみられた。水辺を好むサドガエルにおいても、山地地形 が個体の分散の障壁になったと考えられる。 (2) 国仲地域個体群内の生息地の連結性 Population Graph により、国仲地域個体群における生息地間の連結性を評価し た結果、ネットワークの一部の生息地が環状構造を示した(図 10)。このような構造 を示した理由として、谷津田地形と汽水湖の存在が考えられる。佐渡島の汽水湖の 西部は谷津田地形が広がっており、谷津田を流れる小河川は汽水湖(加茂湖)に流れ 込んでいる。サドガエルは下流の汽水湖ではおそらく生存することができないので、 異なる谷津田へは低地部を介した分散はできないにちがいない。一方、それぞれの 谷津田は、上流の台地を通しての分散が可能と考えられ、それがネットワークの環
8 状構造をつくりあげていると考えられる。 2. 生息に適する環境条件の推定 個体数を目的とした統計モデルの解析の結果、水田周辺の水路の深さと水田の湛水率 のみが説明変数として選択され、水路が浅いほど、または水田の湛水率が高いほどサド ガエルの個体数が増える傾向が認められた。水路は、水田の非湛水期における幼生や成 体のレフュジアになると考えられ、特に幼生が一年を経て成体となるまでの生息地とし て不可欠であろう。しかし、幼生が成体になるまで水路で生存したとしても、サドガエ ルの成体の指には吸盤がないため、コンクリートの壁面を登ることができないと推測さ れる。水田の湛水面積率がある。湛水面積率の変化に応じて個体数が変化する第一の理 由として、個体の移動が考えられる。水田では、例年6 月の半ばごろから中干しにより 水が減るので、サドガエルは水のある水路などへ移動している可能性が高い。 一方、景観要因であるTWI や凹凸度は効果が見られなかった。TWI が高い場所は集 水域が広く、水が溜まりやすいため、本来はサドガエル生息地としての価値が高いはず である。しかし、そのような場所は平野部に位置し、集約的農法の水田が広がっている。 このような水田では農業の近代化による乾田化が進んでおり、サドガエルに不適な環境 に改変されてしまったため、TWI は効いていなかったと考えられる。 3. 保全への提言 ここでは、本研究の結果を踏まえ、環境保全型農業を利用したサドガエルの具体的 な保全策について考察する。 まず、STRUCTURE 解析による遺伝子型解析から、サドガエルは 3 つの地域個体群 に分けられた。3 つの地域個体群は、地形的、地理的に現在は個体の交流がほぼ不可能 なことから、それぞれ異なる保全管理単位と、独立して管理することが適切である。
9 地域個体群間は各々の環境に局所適応している可能性があるため、生息地の保全や再 生を考える際には、地域個体群をまたぐ個体の移入は避けるべきである。特に、東部 と西部では単一のクラスターから形成されているので、個体導入による再生を考える 場合には、同じ地域個体群の個体に限定すべきである。 サドガエル生息地の個体数を向上させるには、レフュジアの環境整備と、繁殖場所 の質の向上が必要な条件であると考える。本研究の結果を踏まえて、3つの具体的方 策を提言する。1 つ目に、水田横の水路と田面間のギャップを減少させることである。 深い水路を浅い水路に改変することは現実的ではないため、カエルが水田と水路を行 き来できるように水路にスロープを設置することや、段差のついた開けた魚道を設置 することがあげられる。 2 つ目に、通年湛水状態の江の設置を提案する。本研究では、江の割合による個体数 の変化がみられなかった。その理由として、現在の江は、大部分の江で水田と同時期 に水が抜かれてしまうため、レフュジアや産卵の場として機能していない可能性が高 い。しかし、先行研究ではサドガエルの個体数に江が正に効いていた(宇留間ら 2010)。 これは今回よりも丘陵地で行われたため、常に湛水状態の江が多い水田が含まれてい たためだろう。江の設置は、証制度の条件の中でも比較的実施しやすいと考えられ、 個体群の維持や復元にとって重要な役割を果たすに違いない。
10 謝辞 本研究を行うにあたって、東京工科専門学校の小林頼太先生には、野外調査に全面的に ご協力していただき、調査手法などについてご意見をいただきました。野外調査では、サ ドガエル発見者である新潟大学の関谷國男教育支援員にも、手厚いご支援をいただきまし た。朱鷺・自然再生学センターの職員の方々には、サドガエルの貴重な分布情報の提供の ほか、調査道具を貸していただくなど、大変お世話になりました。また、トキ交流会館の 職員の方々には、調査の際の宿泊場所や、調査用車の保管場所の提供などのご協力をいた だきました。 以上の皆様、ならびに、近隣農家の皆さま、地域の皆さまには、この場を借りて厚く御 礼申し上げます。
11 引用文献
Dyer R, Nason J (2004) Population graphs: the graph theoretic shape of genetic structure. Mol. Ecol.
井鷺裕司, 陶山佳久 (2013) 第 4 章 遺伝情報をどのように解釈するか―種内の解析. 生態学者が書いた DNA の本 メンデルの法則から遺伝情報の読み方まで(初版), 97-144 文一総合出版
Lian CL, Wadud MA, Geng Q, et al (2006) An improved technique for isolating codominant compound microsatellite markers. J Plant Res 119:415–417. doi:
10.1007/s10265-006-0274-2
Martin TG, Wintle B a, Rhodes JR, et al (2005) Zero tolerance ecology: improving ecological inference by modelling the source of zero observations. Ecol Lett 8:1235–46. doi: 10.1111/j.1461-0248.2005.00826.x
Pritchard JK, Stephens M, Donnelly P (2000) Inference of Population Structure Using Multilocus Genotype Data. Genetics 155:945–959.
Sekiya K, Miura I, Ogata M (2012) A new frog species of the genus Rugosa from Sado Island, Japan (Anura, Ranidae). Zootaxa 62:49–62.
Sørensen R, Seibert J (2007) Effects of DEM resolution on the calculation of topographical indices: TWI and its components. J Hydrol 347:79–89. doi:
10.1016/j.jhydrol.2007.09.001
宇留間悠香, 小林頼太, 西嶋翔太, & 宮下直. (2012). 空間構造を考慮した環境保全型農業の影響 評価: 佐渡島における両生類の事例. Japanese Journal of Conservation Ecology, 164, 155–164.
12 表1 サドガエル個体群のサンプル数と遺伝的変異 n:核採取地におけるサンプリング数および遺伝的解析に用いた個体数。 He:ヘテロ接合度期待値 地域 ID n He 西部(W) OG 13 0.377 AH 14 0.397 TS 14 0.433 国中平野 (K) DM 20 0.565 DMD 16 0.549 SG 20 0.561 KT 16 0.568 KD 17 0.537 NG(P) 19 0.471 YY 16 0.587 NG(R) 35 0.534 ST 13 0.443 MT 19 0.532 US 13 0.593 AT 14 0.542 ON 14 0.535 KB 19 0.45 GS 17 0.59 JK 14 0.568 SS 14 0.548 TY 15 0.548 KG 18 0.522 UM 5 0.500 GZ 17 0.520 MG 17 0.409 HN 15 0.465 東部(E) MK 22 0.334 RO 16 0.334 SD 19 0.363
13 図1 サドガエルの分布図
14 図2 サドガエルの遺伝子サンプル採取地。29 か所で採取した。 サンプリング地点以外の凡例は図1 と同様である。
OG
TS
SD
RO
MK
SG
DM
DM
NG
US
NG
SS
K
ON
KD
K
TY
A
KB
S
Y
MT
GS
J
AK
HN
MG
UM
15 図3 個体数調査地:42 か所
Forest
Paddy fields
Salt water
Others
16 -9000 -8500 -8000 -7500 -7000 -6500 0 5 10 15 20 25 30 0 20 40 60 80 100 120 140 160 0 5 10 15 20 25 30 図4 STRUCTURE 解析の結果 (A)LnP(D) および(B)⊿K
LnP(D)
クラスター数(A)
⊿
K
(B)
クラスター数17 図5 STRUCTURE 解析によるバープロット 1 本の線がそれぞれどの程度の割合で祖先クラスターに属しているのかを示す。黒い線はサ ンプリング集団の境目。
K=2
K=5
K=3
K=4
K=6
18 図6 Population Graph を用いて作成した、国仲地域個体群内の生息地ネットワーク ノードの大きさは遺伝的変異性(遺伝子座多型の数、アリルの数、ヘテロ接合度)を表 し、大きいほど変異性が高いことを意味する。リンクの長さは遺伝的類似度を表し、 類似度が高いほどリンクは短くなる。 図7 Population Graph を用いたネットワーク構造を地図上に当てはめたもの ノードの大きさ・色が示すものは図9 と同様である。 US SG DM DMD NG(R) SS KT ON KD KG TY AT KB ST YY MT GS JK NG(P) HN MG GZ
OG
TS
SD
RO
MK
SG
DM
DMD
NG(R)
US
NG(P)
SS
KT
ON
KD
KG
TY
AT
KB
ST
YY
MT
GS
JK
GZ
HN
MG
19
表2 モデル選択の結果。水路の深さと水田の湛水率はベストモデルから⊿AIC2 以内のモデル すべてで選ばれている。
20 図8 モデル平均した係数の推定値
21 図9 散布図および回帰線 (A)は水路の深さと密度の関係を示し、(B)は水田の湛水面積率と密度の関係を示している。