フィブリンゲル形成における糖-フィブリノーゲン相互作用の研究
135
0
0
全文
(2) <目次> 論文要旨. 1. 1章 序論. 4. 1-1. 1-2. 1-3. 1-4. 1-5. 1-6. 1-7.. 血液凝固とは フィブリノーゲン フィブリンゲル形成過程 これまでの研究動向 我々の研究成果 社会的意義 本論文の構成. 2章 理論的背景 2-1. 光散乱 2-2. 濁度測定 2-3. 共焦点レーザー顕微鏡. 11 16 19. 3章 フィブリンゲル形成に及ぼす糖の異性体効果 3-1. 3-2. 3-3. 3-4. 3-5.. 緒言 試料調製と測定法 単糖の添加効果 二糖の添加効果 結論. 21 23 32 40 49. 4章 糖類添加効果とフィブリノペプチド切断の関係 4-1. 4-2. 4-3. 4-4.. 緒言 試料調製と測定法 結果と考察 結論. 50 53 56 63.
(3) 5章 リコンビナントフィブリノーゲンのゲル化における糖の添加効果 5-1. 5-2. 5-3. 5-4.. 緒言 試料調製と測定法 結果と考察 結論. 65 67 80 101. 6章 総括. 103. 7章 今後の展望. 105. 参考文献. 107. 本研究に関連した論文及び発表. 113. 付録 Ⅰ. フィブリノーゲンのゲル化における NDSB の効果. 118. Ⅱ. リコンビナントフィブリノーゲンの精製. 126. 謝辞. 132.
(4) 論文要旨 出血を防ぐ血液凝固の仕組みは、 免疫とならぶ生体防御の重要なシステムの 1 つである。 高齢化社会を迎え、生活習慣病がますます重要な社会問題となった現代においては、 『血液 凝固機能のコントロール』を目標に、血液・血流に関する研究が盛んに行われている。 血液凝固過程の最終段階は、フィブリノーゲンのゲル化(フィブリンゲル形成)である。 近年、当研究室においては、種々の糖類がこのフィブリンゲル形成に与える影響を報告し てきた。これまでの研究により得られた結果は、糖がフィブリノーゲンと直接相互作用す ることを強く示唆した。本研究は、糖−フィブリノーゲン相互作用メカニズムの解明を目 的として研究を進めた。 まず、フィブリンゲル形成過程と糖との相関関係についてより詳細な知見を得るために、 これまでの研究でゲル化の遅延効果を示すことが明らかになっている単糖(グルコース)お よび二糖(マルトース)の立体異性体を用いて添加効果を検討した。単糖は、グルコースを 基準に、アルドヘキソースの立体異性体であるマンノース、ガラクトース、アロースの 4 種類を用いた。一方、二糖はマルトースと同様に、グルコース 2 分子からなるグルコシル −グルコース型の二糖(イソマルトース、セロビオース、トレハロース)を用いた。これら 糖の立体異性体の添加がフィブリンゲル形成に及ぼす影響について、散乱光強度の経時変 化、濁度測定、共焦点顕微鏡(CLSM)観察により検討した。また、フィブリノペプチド A, B (FPA, FPB)切断の有無については HPLC により確認した。単糖の立体異性体の添加はいず れもフィブリンのゲル化を遅延し、細くルーズなフィブリン繊維を形成することが明らか になった。二糖の立体異性体では、マルトース、セロビオース、トレハロースの添加がゲ ル化を遅延し、細い繊維が形成され、唯一、イソマルトースの添加はゲル化にもフィブリ ン繊維の凝集状態にも影響を与えないことが明らかになった。これらの単糖・二糖存在下 においても、フィブリノペプチドの遊離が HPLC により検出され、これらの糖類はフィブ リノーゲンと直接相互作用していることが示唆された。フィブリノーゲンのゲル化過程、 形成されるフィブリン繊維の凝集構造およびフィブリンゲルのネットワーク構造において、 単糖・二糖の立体構造に依存した影響(糖の異性体効果)を及ぼすことが明らかになった。. 1.
(5) また、FPA、FPB の両方を切断してフィブリンゲルを形成させるトロンビン系と、FPA のみを切断するレプチラーゼ系を比較することにより、フィブリノペプチド切断とフィブ リンゲル形成との関係を検討した。さらに、それぞれの系において、グルコースとデキス トランの添加効果を調べ、フィブリンゲル形成過程と糖類添加効果の関係を、動的光散乱 測定、HPLC 測定および CLSM による観察により比較検討した。レプチラーゼ系では、ト ロンビン系と同様にグルコースの添加でゲル化が遅延され、デキストランの添加でゲル化 が促進された。しかし、レプチラーゼ系においては、形成されたゲルのネットワーク構造 にグルコース添加による影響がほとんど観察されず、それらのネットワーク構造は、トロ ンビン系におけるグルコース添加時に類似した構造であった。一方、デキストラン添加時 のネットワーク構造は、トロンビン系におけるデキストラン添加の結果と酷似していた。 さらに、添加したグルコースとデキストランは、レプチラーゼの活性に影響を与えないこ とも明らかになり、どちらの系でも糖類はフィブリノーゲンと直接相互作用していること が強く示唆された。 これまでのすべての研究結果から、グルコースとデキストランでは、フィブリノーゲン への相互作用機構が異なると考えた。グルコースの作用は、グルコースがフィブリノーゲ ンのαC ドメインと相互作用することによりプロトフィブリルのラテラル凝集を抑制し、 一方、デキストランの作用は、デキストランが接着剤のように機能してプロトフィブリル の凝集を促進していると考え、それぞれの作用モデルを考案した。 そして、糖−フィブリノーゲン相互作用機構の解明と、考案した糖の作用モデルを検証 するために、糖との相互作用が考えられるフィブリノーゲンのαC ドメインに変異を導入 したリコンビナントフィブリノーゲンを作製し、濁度測定と形成されたフィブリンゲルの CLSM 観察により糖(グルコースおよびデキストラン)の添加効果を比較検討した。その結 果、フィブリノーゲンのゲル化過程、形成されるフィブリン繊維の凝集構造およびフィブ リンゲルのネットワーク構造に対して、2 種類の欠損型リコンビナントフィブリノーゲン (Aα508stop と Aα571stop)においては、グルコース添加による影響は観測されず、それら のコントロール(糖添加なし)が Native(ヒト血漿由来フィブリノーゲン)のグルコース添加 時の結果と良く一致した。この結果から、Aα508stop および Aα571stop が共通して欠損 しているαC ドメイン C 末端側のアミノ酸 40 残基(Aα571-610)の領域が、フィブリンゲ 2.
(6) ル形成過程において繊維を形成する段階、つまりプロトフィブリルのラテラル凝集に関与 し、さらにグルコースと相互作用する部位がこの領域に含まれることが明らかになった。 また、これらの結果は、考案したグルコースの作用モデルを支持するものであり、今後、 フィブリノーゲン Aα571-610 の領域に様々な変異を導入することで、さらなるグルコー ス相互作用部位の特定化が可能であることが本研究により示された。 本研究においては、糖−フィブリノーゲン相互作用のみならず、これまで構造・機能と もに未解明であったフィブリノーゲンαC ドメインの機能に関して、極めて有益な情報を 得ることに成功した。これらの結果は、血液凝固のメカニズムのより深い理解へとつなが るものであり、 『血液凝固機能のコントロール』へ向けた大きな進展が期待できる。. 3.
(7) 1章 序論 1-1. 血液凝固とは 止血(血栓形成)機構は、血小板凝集による血小板血栓の形成過程である一次止血と、こ の血小板血栓をより強固なものにするために引き続いて起きる二次止血(血液凝固)の 2 段 階に分類される(図 1.1)。 フィブリノーゲンのゲル (フィブリンゲル)は、赤血球や血小板などの血球. 血小板血栓. フィブリン網. を取り込みながらゲルを形成し、出血を完全に阻 止する役割を持つ。血液凝固機構は、循環器系の 生体防御機構として機能しており、 健常な人体(動. 一次止血. 物)においては状況に応じて血液凝固が促進また. 二次止血(血液凝固). 図 1.1 止血機構. は抑制され、生体内で精緻にコントロールされて いる。本研究で注目しているフィブリンゲル形成過程は、血液凝固機構において最も重要 な反応の1つである。血液凝固カスケードを図 1.2 に示す。体内で起きる反応は、図 1.2 に示したように非常に複雑であり、多くの血液凝固因子が関与している。. 内因系凝固系 XII. 体内での異物との接触など. XIIa XI. 抗凝固機構 XIa Ⅸa. Ⅸ Ca2+ リン脂質. アンチトロンビンⅢ Ⅷa. Ⅷa ヘパリン. Ⅶ. Ⅶa. 外傷など. Xa. X. 外因系凝固系. V. Va. Ca2+ リン脂質. トロンビン. プロトロンビン. フィブリノーゲン XIII. フィブリン. XIII a 凝固. ゲル化. 図 1.2 血液凝固カスケードの模式図. 4.
(8) 1-2. フィブリノーゲン 本研究の主たる対象であるフィブリノーゲンは、血液凝固第 I 因子の血漿タンパク質で、 フィブリノペプチド A、B(FPA, FPB)を含むサブユニット Aα鎖、Bβ鎖と、さらにγ鎖の 異なる3 本のペプチド鎖2 組が、 N 末端側でジスルフィド結合によって二量体を形成した、 分子量 34 万の剛直な棒状タンパク質である。 各サブユニットの分子量は、Aα鎖 6.7×104 Da、Bβ鎖 5.6×104 Da、γ鎖 4.75×104 Da とされ、Bβ鎖およびγ鎖には糖鎖が付加されている。また、これら 3 種鎖のサブユニッ トポリペプチドの一次構造が明らかにされ、Aα鎖は 610 残基、Bβ鎖は 461 残基、γ鎖は 411 残基からなることが明らかになっている。Aα鎖および Bβ鎖の N 末端にはそれぞれ アミノ酸 16 残基および 14 残基からなる FPA、FPB が配置されている。これらのペプチド はいずれも負電荷を豊富に有し、フィブリノーゲンの N 末端に負電荷を与えるため、フィ ブリノーゲン分子中央部の E ドメインは負に荷電している。E ドメインは、負電荷を豊富 に含むフィブリノペプチドの切断によりフィブリンに転換すると、正に荷電する。一方、 フィブリノーゲン分子の両端部に位置する D ドメインは、負に荷電しておりフィブリンに 転換しても変化はない。フィブリノーゲンはどの部位も負に荷電しているため、分子同士 の凝集は起きにくい。しかし、フィブリンになると、中央部 E ドメインと他分子の両端部 D ドメインが引き合うため、特異的会合(フィブリンゲル形成)にプラスに作用する。 フィブリノーゲンは、中央部の E ドメイン、両端部の D ドメインが 3 重へリックス部を 介して D-E-D と配置され、さらに Aα鎖のC末端側に位置するαC ドメイン(約 420 残基) が、D ドメイン部から中央部 E ドメインに折り返した構造をしている(図 1.3 [1])。. 図 1.3 フィブリノーゲン分子の構造模式図 [1]. 5.
(9) 1-3. フィブリンゲル形成過程 血液凝固機構の最終段階においては、まず、 フィブリノーゲンにセリンプロテアーゼであ. フィブリノーゲン FPA. るトロンビンが作用することにより、E ドメ インに位置する FPA、FPB が切除され、フィ プロトフィブリル. ブリンに転換する。フィブリンは、FPA の切 断部分で互いに凝集して分子軸方向へ成長し. FPB 遊離したαCドメイン. ていき、長く伸びたプロトフィブリルを形成 する。それに引き続くプロトフィブリルどう しのラテラルな凝集により、フィブリンファ イバーを形成し、ネットワークが構築され最 終的にフィブリンゲルが形成される(図 1.4)。. フィブリンファイバー (フィブリンゲル・ネットワーク構築). 図 1.4 フィブリンゲル形成過程. 図 1.4 では、FPA 遊離後、FPB が遊離するよ うに示しているが、実際にはほぼ同時にトロンビンにより切除される。フィブリノーゲン において、立体構造的に FPB よりも FPA の方が外側に露出していることから、FPA の切 除が幾分はやくなり、プロトフィブリルの形成段階が優先的に進行する[2]。 図 1.4 に示したように、ラテラルな凝集過程においては、FPB の切除に伴い E ドメイン から遊離したαC ドメインを介した結合が他分子(プロトフィブリル)間に形成されること により促進されると報告されている[3]。. 1-4. これまでの研究動向 このようなフィブリノーゲンのゲル化は、光散乱[4-7]、濁度[8-10]、電子顕微鏡[11]など の手法を用いて研究されていて、形成されるゲルの構造や性質がフィブリノーゲンとトロ ンビンの濃度に依存することが報告されている。また、イオン強度[12]や pH の変化[13]に 対する影響を調べた研究や、2 価の陽イオン[14,15]やデキストラン[8,9,16,17]などの添加効 果が研究されている。. 6.
(10) フィブリノーゲンの構造や機能、またはフィブリンゲル形成に関する研究は、古くは電 子顕微鏡による全体像の観察[18]など、血漿由来のフィブリノーゲンを用いたマクロな観 測により行われてきた。フィブリノーゲン分子の結晶化が困難であることから、フィブリ ノーゲンの構造研究には長年を費やした。その後、構造と機能の相関関係について、フィ ブリノーゲンの構造異常により機能障害が生じている異常フィブリノーゲン血症の研究を 通して進められるようになった。科学技術の進歩により、この異常フィブリノーゲンの解 析には分子生物学的手法が用いられ、遺伝子レベルでの解析およびリコンビナントフィブ リノーゲンの作製が行われた[19,20]。さらに近年においては、このリコンビナントフィブ リノーゲン作製技術を利用し、より詳細な分子構造の解析や分子間の相互作用解析など、 フィブリノーゲンの構造や機能に関するミクロな解析が盛んに行われている[21-25]。. 1-5. 我々の研究成果 近年、当研究室においてはシクロデキストリンをはじめとする種々の糖類がフィブリン ゲル形成に与える影響を報告してきた[26-30]。博士前期課程における研究では、フィブリ ンゲル形成過程への糖添加による影響を単糖から多糖に至るまで系統的に調べ、単糖やオ リゴ糖の添加がフィブリンゲル形成を 遅延、一方、多糖の添加はフィブリン 160000. ゲル形成を促進し、糖類添加による影. ことを見出した(図 1.5 [29])。また、こ れらの糖類の添加はフィブリンファイ バーの凝集構造にも大きな影響を与え、. Intensity [ arbit. units ]. 響が糖の分子量(分子鎖長)に依存する. 120000 100000 80000 60000 40000 20000. 糖類添加による影響がフィブリンゲル. 0. 形成過程のみでなく、形成されたフィ. 0. ブリンゲルにも及ぶことを報告した [29,30]。. Control Dextran (Mw 77000) Dextran (Mw 9500) Dextran (Mw 4300) Glucose Maltose Maltotetraose Maltoheptaose. 140000. 10. 20. 30. 40. 50. 60. 70. 80. 90. 100. elapsed time [ min ]. 図 1.5 フィブリノーゲンのゲル化過程における散 乱光強度の経時変化(糖類の添加効果)[29]. 7.
(11) これまでの研究から得られた結果は、フィブリノーゲンは糖と直接相互作用し、フィブ リンゲル形成が糖の存在により制御されることを示唆した。糖の中には、タンパク質のリ フォールディングや凝集の抑制効果を持つものも知られている。このような糖は、タンパ ク質の周りに結合する水分子の数を増やすことにより、タンパク質を安定化させる。この ようなタンパク質のリフォールディングや凝集抑制効果を示す、糖以外の物質の添加によ るフィブリンゲル形成への影響について、NDSB(non-detergent sulphobetaine)を用いて検討 した[31]。その結果、NDSB 存在下においてもゲル化はするが、形成されたフィブリンゲ ルは透明なゲルで、さらにフィブリン繊維またはネットワーク構造は共焦点レーザー顕微 鏡によって観察できない程であった。NDSB 存在下においては、グルコース存在下よりも 均一かつ細かいネットワーク構造を持つフィブリンゲルが形成されていると推測され、非 常に興味深い結果であった。 また、フィブリンゲル形成過程と糖との相関関係を調べる 1 つのアプローチとして、当 研究室ではフィブリノーゲンの結合糖鎖とフィブリンゲル形成の関係を検討した。フィブ リノーゲンの糖鎖は、フィブリンゲル形成とフィブリン繊維の凝集(特にラテラル凝集)に 関与していると報告されている[32]。糖鎖除去フィブリノーゲンによる実験で、グルコー スおよびデキストランの添加は糖鎖の有無に関わらず、フィブリンゲル形成過程に影響を 与えるという結果が得られた[33]。. 1-6. 社会的意義 日本では高齢化社会を迎え、生活習慣病がますます重要な社会問題となってきた。生活 習慣病に起因する症状には血液関連の疾病が多く(例えば高血圧症、 心筋梗塞、 脳梗塞など)、 これらの疾病を治療、または予防することはこれからの社会において重要な課題である。 近年、生活習慣病予防の観点からも、血液や血流に対する関心が非常に高まっている。 血流を悪化させる原因としては、血液中の過剰な脂質や糖質が挙げられている。血栓形 成のきっかけとなる血小板凝集(一次止血)における糖または脂質の影響については、精力 的に研究されており、糖による血小板凝集の促進[34]などの報告がある。一方、フィブリ ノーゲンのゲル化がメインとなる血液凝固(二次止血)においては、脂質や糖質の直接的な. 8.
(12) 影響に関する先行研究はほとんど存在しなかった。 これまでの研究において、フィブリノーゲンのゲル化に対する添加効果を検討した糖類 の中でも、グルコースやマルトースはエネルギー源として生体内に存在し、その溶液は輸 液として臨床でも繁用されている。とりわけグルコースは、血液中に存在する最も一般的 な糖であり、血糖値は血中グルコース濃度で示される。血糖値は高すぎても、また低すぎ ても身体にダメージを与え、高血糖の状態が続く症状を示す疾病には生活習慣病の 1 つで ある糖尿病がある。 臨床的には、 糖尿病患者は血栓症を発症しやすいことがわかっている。 この原因の 1 つには、高血糖により血小板凝集能が高まることが考えられているが、これ までの研究に基づけば、グルコースの存在下においては、細い繊維により分岐の多い緻密 なネットワーク構造のフィブリンゲルが形成され、ゲルの透過能が低下したために、生体 内で血栓を溶解する酵素プラスミンによる線溶機構が効果的に進行しなくなり、形成され た血栓が溶解することなく残った結果だと考察された。 糖類と血液凝固の相関関係またはフィブリノーゲン‐糖相互作用メカニズムの早急な 解明、そしてフィブリン分子がレクチン様の糖認識・相互作用部位を持つことの分子論的 検証は、血液凝固機能のコントロールへ向けた大きな一歩として、強く期待されている。. 1-7. 本論文の構成 本研究の目的は、これまでの研究により見出されたグルコース、デキストラン、シクロ デキストリンによるフィブリノーゲンとの特異的な相互作用のメカニズムを明らかにする ことである。この目的のために、まず、単糖・二糖の立体異性体の添加効果およびレプチ ラーゼ系における糖類添加効果を検討し、フィブリンゲル形成過程と糖との相関関係につ いて、より詳細な知見を収集した。さらに、変異を導入したリコンビナントフィブリノー ゲンを作製し、ゲル化における糖類添加効果を検討することで、部位特異的な糖-フィブ リノーゲン相互作用について議論した。 本論文は、1, 2 章において序論と実験手法、3~5 章では実際の実験とその結果、6, 7 章 は本研究のまとめと今後の展望という構成である。. 9.
(13) まず 2 章では、本研究において行った実験手法(光散乱、濁度測定、共焦点レーザー顕 微鏡観察)について述べる。試料作製には分子生物学的手法を用いたが、実験測定には主と して物理学的手法を用いた。 3 章では、フィブリンゲル形成に及ぼす糖の異性体効果の検討を目的として、単糖(グル コース)および二糖(マルトース)の立体異性体を用いて行った添加実験の結果を示す。 添加 フィブリンゲル形成過程を散乱光強度測定により解析し、形成されるフィブリン繊維の凝 集構造およびネットワーク構造は濁度測定と共焦点レーザー顕微鏡観察により行った。 次の 4 章では、 フィブリノペプチド切断とフィブリンゲル形成との関係を調べるために、 FPA、FPB の両方を切断してフィブリンゲルを形成させるトロンビン系と、FPA のみを切 断するレプチラーゼ系を比較検討した結果について述べる。さらに、それぞれの系におけ る糖類の添加効果を調べ、フィブリンゲル形成過程と糖類添加効果の相関関係を検討した 結果も述べる。グルコースおよびデキストランの添加効果を、動的光散乱測定および共焦 点レーザー顕微鏡観察により検討した。また、4 章では、これまでのすべての研究結果か ら考案した、グルコースおよびデキストランの作用モデルを示す。 5 章では、部位特異的な糖-フィブリノーゲン相互作用について検討するために、変異 を導入したリコンビナントフィブリノーゲンを作製し、糖の添加効果を調べた結果を述べ る。濁度測定と共焦点レーザー顕微鏡観察により変異と糖の添加効果の関係を調べた。 6 章においては、これら 3~5 章における実験結果を総括し、本研究結果を踏まえた今後 の展望について 7 章で述べる。. 10.
(14) 2 章 理論的背景. 2-1. 光散乱[1-4] 光散乱法は、高分子の形状や分子量、分子間相互作用の大きさなどを求める実験法 である。本研究では、この光散乱法(動的光散乱法)を用いてフィブリノーゲンのゲル 化過程の挙動を検討した。本節では光散乱法について説明する。. 光散乱 月の世界では、太陽が見えていても空が真っ暗であるという。しかし、地球上では空が 青い。これは地球には空気があるからであり、空気の中に小さなゴミなどが浮かんでいる ためでもある。太陽光は空気の密度のゆらぎや浮遊するゴミによって散乱される。これは レイリー(Rayleigh)散乱と呼ばれる散乱である。散乱光の強度は波長の4乗に反比例し、 波長の短い光は強く散乱される。青色や紫色の光は黄色や赤色の光よりも強く散乱される ことになる。そのため空は青く見える。一方、日の出や日の入りの太陽光は地面近くの空 気中の長い距離を通ってくるので、青色や紫色の光は途中で大半が散乱されてしまい、赤 色の光だけが届く。そのため日の出や日の入りの太陽は赤い。このように地球で生活を送 る私たちにとって、光散乱という現象は非常に身近である。 光散乱法は、高分子や多数の粒子から構成される系に光を照射し、ゆらぎの観測から、 ゆらぎの振幅、振動数、緩和時間などを調べ、散乱をひきおこす基になっている粒子,分 子の特性を決定するという実験法である。主に濃度、密度分子配向のゆらぎが光散乱に起 因する。したがって、光散乱法は光の波長程度のゆらぎをもつ粒子に関する研究に適して いる。光散乱法は一般にレイリー散乱のことを指すが、そのうちでも手法の違いにより静 的光散乱(Static Light Scattering ; SLS)法と動的光散乱(Dynamic Light Scattering ; DLS)法に 分けられる。. 11.
(15) 静的光散乱 静的光散乱(SLS)法では、重量平均分子量 Mw、2乗平均回転半径<Rg2>、第2ビリアル 係数 A2 などが、散乱光強度の時間平均から求めることができる。 散乱強度は入射光強度 I0 と散乱体積 V に比例する。しかし、観測点との距離 L の2乗に 反比例して減少する。したがって、異なる装置、光学系で測定した結果を比較する時、規 格化された散乱強度は次のように定義される。(入射光が非偏光の場合). Rθ =. I s (θ ) V 1 + cos θ I 0. (. L2. 2. ). (1-1). (1-1)式に示した Rθ をレイリー比と呼ぶ。また、(1-1)式において、IS は散乱光強度、θ は 散乱角を示す。光散乱測定装置で得られる散乱光強度に比例した信号から、このレイリー 比を計算するには、レイリー比が既知の標準物質について強度測定を行い、装置定数を求 める。 高分子系においてΔRθは次式のように書ける。. Kc 1 = + 2 A2 c + 3 A3 c 2 + L ∆Rθ M w ⋅ P (θ ). (1-2). ここで、P(θ)は粒子散乱関数、K は光学定数、c は散乱体の質量濃度を示す。また、A3 は 第3ビリアル係数である。A2 は高分子と溶媒の親和性を表す重要な量であり、良溶媒系で 貧溶媒系では 0 に近い値をとる。 特にθ点では A2 = 0 となる。 は 10-4 cm3mol/g2 以上の値を、. (1-2)式は、θ→0、c→0、θ,c→0 の極限において次のように変形できる。 Kc 1 = + 2 A2 c + 3 A3 c 2 + L ∆Rθ θ →0 M w. (1-3). Kc 1 1 2 2 = 1 + Rg q + ⋅ ⋅ ⋅ 3 ∆Rθ c →0 M w . (1-4). Kc 1 = ∆Rθ θ →0,c →0 M w. (1-5). 通常、(1-1)式を sin2(θ/2)に対してプロットする Zimm プロット[5]を行い、(1-3)、(1-4)、. (1-5)式から A2、<Rg2>1/2、Mw を求める。. 12.
(16) 動的光散乱 動的光散乱(DLS)法では、ゆらぎの経時変化、すなわち散乱光強度の時間相関を求め、 流体力学的半径や並進拡散定数などを求めることができる。これらは散乱光電場の時間相 関関数から求まるが、電場の測定が困難であることから、比較的容易に得ることができる 散乱光強度の時間相関関数を用いて議論する。. 電場の時間相関関数および散乱強度の時間相関関数はそれぞれ(1-6)、(1-7)式のように定 義される。. G (1) (t ) ≡ E (q,0)E * (q, t ). (1-6). G (2 ) (t ) ≡ I s (q,0)I s (q, t ) = E (q,0)E * (q,0)E (q, t )E * (q, t ). (1-7). (1-7)式は次式のように変形できる。 G (2 ) (t ) = E (q,0 )E * (q,0 )E (q, t )E * (q, t ) = E (q,0)E * (q,0) E (q, t )E * (q, t ) + E (q,0 )E * (q, t ) E * (q,0)E (q, t ) (1-8) 2. = G (1) (0) + G (1) (t ). 2. ここで、. g (1) (t ) ≡ G (1) (t ) / G (1) (0 ) g (2 ) (t ) ≡ I s (q,0)I s (q, t ) / I s (q,0). 2. = G (2 ) (t ) / G (1) (0). 2. (1-9). と定義すると、(1-8)式から Siegert の関係式[6]が得られる。. g (2 ) (t ) = 1 + g (1) (t ). 2. (1-10). ここまでは、散乱体上での入射光の広がりは点光源(無限小)であり、試料−検出器間距 離に比して検出器前のピンホールは無限小であるという2つの条件を前提としている。. 13.
(17) しかし、実際には有限サイズのピンホール径での測定が行われるので(1-10)式は、. g (2 ) (t ) = 1 + β g (1) (t ). 2. (1-11). となる。(1-11)式において、β(0<β<1)は干渉性因子と呼ばれる、光学系に依存した定 数を示す。. (1-11)式をふまえて、単分散希薄系について議論する。互いに相互作用を持たない粒子の 集団運動(ブラウン運動)について時間相関関数を求める。この時、粒子間に相互作用がな いため、ブラウン運動はランダムな並進拡散運動となる。 G (1) (t ) ≡ E (q, t ′)E * (q, t ′ + t ) N. [. ]. N. * exp[− iq ⋅ rm (t ′ + t )] = ∑ a j exp iq ⋅ r j (t ′) ∑ a m j. m. [. N N. (1-12). ]. = ∑ ∑ G j G m* exp iq rm (t ′ + t ) − r j (t ′) j m. [. N. ]. = ∑ a j a *j exp iq ⋅ δr j (t ) j. この系において粒子は(1-13)式の拡散方程式を満足していることから、平均<exp[iq・ δr(t)]>は(1-14)式のようにあらわされる。. δr 2 exp− 4 Dt . 1. W (δr, t ) =. (4πDt )3 2 exp[iq ⋅ δr (t )] = ∫ exp[iq ⋅ δr j (t )]W (δr, t )dδr = exp[- Dq 2 t ]. (1-13) (1-14). よって、. [. ]. [. G (1) (t ) = N a exp − Dq 2 t = I exp − Dq 2 t 2. [. g (1) (t ) = exp − Dq 2 t. ]. ]. (1-15). が得られ、これより(1-16)式が得られる。 g (1) (t ) ≡ g (2 ) (t ) ≡. E (0 )E * (t ) I (0 ) I (0 )I (t ) I (0 ). 2. [. = exp − Dq 2 t. [. ]. = 1 + β exp − 2 Dq 2 t. ]. 14. (1-16).
(18) 散乱体が単分散粒子の希薄溶液である場合、拡散係数 D から粒子の流体力学的半径 Rh が次式の Stokes-Einstein の関係式を用いることで推定可能である。. Rh =. kT 6πηD. (1-17). (1-17)式において、k はボルツマン定数、T は絶対温度、η は溶媒の粘度を示す。. 15.
(19) 2-2. 濁度測定[7,8]. フィブリノーゲン‐トロンビン系では、通常、白濁したゲルが形成される。濁度測 定では、このゲル化に伴う濁度の経時変化だけでなく、濁度を解析することにより繊 維(フィブリンファイバー)に関する3つのパラメーターを求めることができる。本節 では、濁度の解析方法について簡単に説明する。. 溶液の濁度τは、透過光の散乱による強度減少の尺度として定義される。吸収の起こら ない試料においては、散乱光強度から算出することが可能である。散乱光強度は入射光と 散乱光の方向との間の角度θに依存する。. τ = ∫0π 2πd 2 (iθ I 0 )sin θdθ. (. (2-1). ). iθ I 0 = Rθ 1 + cos 2 θ d 2. (2-2). (2-1)、(2-2)式において、iθは単位体積あたりの散乱強度、I0 は入射光の強度、d は散乱 体積と検出器の距離を示す。散乱因子 Rθはレイリー比であり、散乱粒子の大きさと形に依 存する。長く細い棒状の粒子に対する散乱因子は次式で与えられる。[9-11]. Rθ = cKλµ 4n sin (θ 2 ). (2-3). (2-3)式において、c は濃度、λは入射光の真空における波長、μは棒状粒子の質量と棒 の軸方向の長さの比(Dalton/cm)、そして n は溶液の屈折率を示す。また、定数 K は次式で 定義され、dn/dc は溶質の示差屈折率、N はアボガドロ数を示す。. K = 2π 2 n 2 (dn dc ) Nλ4 2. (2-4). 16.
(20) (2-1)式に iθ/I0 と Rθを代入すると、. (. ). τ = (πcKλµ 2n )∫0π 1 + cos 2 θ (sin θ ) sin(θ 2 )dθ. (2-5). となる。また、(2-5)式の積分を実行すると、. τ = (44 15)πKcλµ n. (2-6). となり、さらに K を置き換えると、. [. ]. τ = (88 15)π 3 n(dn dc )2 c N µλ− 3. (2-7). となる。(2-7)式は、τ vs. λ-3 のプロットが原点を通る直線になり、その傾きが質量と長 さの比(μ)に比例していることを意味している。 フィブリノーゲンのゲル(フィブリンゲル)の濁度は、かなりの波長範囲にわたり 1/λ3 に比例する。しかし、ゲル化に伴ったμ値の増加につれて、その比例関係は成立しなくな る。これは大きなμ値を持ったフィブリン繊維の直径が波長に比べて大きくなったためで ある。その場合には、直径を含んだ次の関係式から求められる。. (44 15)πKcλ. (. ). nτ = µ −1 1 + 184π 2 ρ 2 n 2 77λ2 + K. (2-8). この(2-8)式は、(2-4)式を用いて K を置き換えることで次式のように書き換えることができ る。. [(88 / 15)π. 3. n(dn / dc ) / N 2. ] (c / τλ ) 3. (. )(. ). = µ −1 + 184π 2 n 2 77 ρ 2 µ λ−2. (2-9). (2-9)式において、ρは棒状粒子の断面の回転半径であり、円柱形の粒子の半径 r は、次式 の関係にある。. ρ2 = r2 / 2. (2-10). さらに、定数 A と B を用いて簡略化した形に書き換えると次式であらわされる。. (c τλ ) = Aµ 3. −1. (. ). + B r 2 µ λ−2 17. (2-11).
(21) このとき、定数 A と B は次のようになる。. [. A = (88 15 )π 3 n(dn dc ) N. (. 2. ]. −1. ). B = 92π 2 n 2 77 A. (2-12). (2-13). (2-11)式は、c/τλ3 vs. λ-2 のプロットが直線関係にあり、切片の値がμ値に反比例するこ とを示す。また、切片に対する傾きの比は棒状繊維の半径(r)の二乗に比例する。. フィブリンゲルにおいては、(2-7)式と(2-11)式を用いて濁度を解析し、繊維の質量と軸方 向の長さの比(μ)と繊維の半径(r)、そしてμ/πr2(繊維の密度)の 3 つのパラメーター を求めて検討した。このとき定数 A と B は以下の値を使用した。. A = 6.76 × 10 22 B = 1.41 × 10 24 このとき、 屈折率n と示差屈折率(dn/dc)は、 それぞれ水とフィブリノーゲンの波長 633 nm における値である。. 18.
(22) 2-4. 共焦点レーザー顕微鏡[12-15]. 共焦点レーザー顕微鏡の開発により、電子顕微鏡では不可能であった生きた細胞の 観察、さらには細胞内の立体微細構造や分子の動きまでも可視化できるようになった。 本節では、共焦点レーザー顕微鏡の原理について簡単に説明する。. 共焦点レーザー顕微鏡(Confocal Laser Scanning Microscope ; CLSM)は、 試料の一部分に焦 点を絞りレーザーを照射し、発生した蛍光を検出することにより試料の3次元構造を解像 度よく構築することができる顕微鏡である。十数年前に生物試料用の機種が開発されて以 来、顕微鏡の一般的機種の一つとして普及、定着している。 一般的な光学顕微鏡の結像光学系と共焦点光学系を図 2-3.1 に示す。. 結像光学系. 共焦点光学系 光検出器. 画像センサ. ピンホール. 像位置. ビームスプリッタ 点光源. ランプ. ピンホール. 対物レンズ. 試料(合焦点). 試料. 試料(焦点外れ). 図 2-3.1 光学顕微鏡の結像光学系と共焦点光学系. 19.
(23) 共焦点光学系では、対物レンズの焦点位置と共役な位置(像位置)に円形の開口をもつピ ンホールを配置することで、焦点のあった位置のみの光を検出することが可能となってい る。通常の光学顕微鏡では、決められた領域を均一に照射することが重要になるが、共焦 点光学系においては点光源から出射した光は、対物レンズにより1点に集光するように照 射する。点光源として、一般的には特定の波長をもち、直線性に優れているレーザー光を 使用し、強い光を1点に集光させるので、試料を均一に照明する通常の光学顕微鏡に比べ、 周辺からの不要な散乱光がなく、コントラストが向上する。試料の表面で反射された光は 同じ光路を戻り、ビームスプリッタにより分離されて、ピンホール上に集光される。この 光学系では、焦点以外からの反射光のほとんどがピンホールでカットされる為、焦点位置 のみの情報が得られる。通常の光学顕微鏡では、焦点位置以外からの画像が重畳している のに対し、共焦点顕微鏡では焦点の完全に合った、コントラストが鮮明でクリアな画像を 得ることが可能である。したがって、測定点を 走査して焦点位置を変えることにより、厚みの ある試料においても薄い切断面の像を得ること が可能であり、その像から3次元像を構築する ことも可能である。これは光学セクショニング 機能と呼ばれ、共焦点光学系の特長の1つであ る(図 2-3.2)。 通常の光学顕微鏡では『解像度を高くする』. 図 2-3.2 光学セクショニング. ことと『焦点深度を深くする』ことは二律背反 する条件であり、高倍率では特に問題であった. 明るさ 通常の顕微鏡. が、共焦点顕微鏡はこの問題を解決している。 共焦点光学系では、試料が焦点位置に正しく置 かれる時に明るさが最大となり、その前後で明 るさは激減する(図 2-3.3 実線)。これに対し、. 共焦点顕微鏡. 通常の光学顕微鏡では、焦点位置前後において 明るさに変化はない(図 2-3.3 点線)。焦点面の. −. 合焦点. +. 鋭い選択性が、共焦点顕微鏡による高さ方向測 図 2-3.3 焦点位置と明るさの関係. 定や、焦点深度拡張の原理である。. 20.
(24) 3章 フィブリンゲル形成に及ぼす糖の異性体効果. これまでの研究により、フィブリンゲル形成におけるグルコース、デキストラン、 シクロデキストリンによる添加効果から、糖の種類や分子量に依存した、糖とフィブ リノーゲンとの特異的な相互作用が見出された。本章では、フィブリノーゲンとの相 互作用における特異性についてさらに詳しく検討するために、単糖と二糖の立体異性 体を用いて、フィブリンゲル形成に及ぼす影響を、散乱光強度の経時変化、濁度測定、 共焦点レーザー顕微鏡観察により検討した。また、フィブリノペプチド切断の有無に ついては HPLC により確認した。. 3-1. 緒言 これまで、グルコース、デキストラン、シクロデキストリンなどの様々な糖を用いて、 フィブリンゲル形成過程に及ぼす糖類の影響を検討してきた。 デキストランは 50 年以上も前から代用血漿として繁用されてきた多糖であり、フィブリ ノーゲンのゲル化を促進することが知られていたが[1-4]、それに加え、近年、糖添加によ る影響をグルコースからデキストランに至るまで系統的に調べた結果、添加による効果は 糖の分子量に依存し、分子量の増加と共に遅延から促進へと推移することが明らかになっ た。その効果は、デキストランの分子量にも依存していた[5-7]。また、環状のオリゴ糖で あり、その空洞内に様々な分子を包接することができる特徴をもつシクロデキストリン (CD)では、α-CD とγ-CD の添加はフィブリノーゲンのゲル化を遅延し、β-CD の添加は ゲル化を阻害することが明らかになっている[8-10]。 これらの研究から、糖とフィブリノーゲンとの相互作用は、糖の種類や分子量に依存し た特異的な作用であることが見出された。そこで、フィブリノーゲンと糖の相互作用にお 21.
(25) ける特異性についてさらに詳しく検討する為に、単糖と二糖の立体異性体を用いて、フィ ブリンゲル形成に及ぼす影響を調べた。単糖はグルコースを基準にアルドヘキソースの立 体異性体(マンノース・ガラクトース・アロース)を選び、一方、二糖はグルコース 2 分子 からなるグルコシル-グルコース型の二糖(マルトース・イソマルトース・セロビオース・ トレハロース)を選んだ。これら糖の立体異性体の添加がフィブリンゲル形成に及ぼす影響 について、散乱光強度の経時変化、濁度測定、共焦点レーザー顕微鏡観察により検討した。 また、トロンビンの酵素活性への糖の影響を調べるために、フィブリノペプチド切断の有 無を HPLC により確認した。. 22.
(26) 3-2. 試料調製と測定法 〈試料調製〉 フィブリノーゲン 撥水処理(シリコンコート)したバイアルビンにウシフィブリノーゲン(Sigma-Aldrich) を秤量し、PBS を加える。この時の濃度は約 20 mg/ml とした。室温(20℃)で 2~3 時間、 時々ゆっくりと転倒混和しながら完全に溶解させる。このようにして調製したフィブリ ノーゲン溶液を、PBS を用いて一晩透析した。透析後、その一部(300 µl 程度)を 10 倍希 釈して波長 280 nm での吸光度測定を行い、吸光係数 1.51 ml/(mg∙cm)を用いて濃度を決 定した。目的濃度に調製した後 PET チューブに小分けし、液体窒素で凍結してディープ フリーザー中(-80℃)に保存した。実験測定時の最終濃度は 2.0 mg/ml で行った。. トロンビン ウシトロンビン(Wako Pure Chemical Industries;1000 U)は試薬瓶に直接 PBS を加え、 室温でゆっくり混和する。その溶液を 50 U/ml に希釈して、200 μl ずつマイクロチュー ブに小分けし、液体窒素で凍結した後、ディープフリーザー(-80℃)で保存した。実験 測定時の最終濃度は 0.01 NIH units/ml で行った。 本研究に用いたフィブリノーゲン濃度は、生体濃度の下限濃度近辺に設定してあり、 トロンビン濃度は実験系に適当である濃度を選択した。トロンビンは、生体内では前駆 体(プロトロンビン)として存在しているため、必ずしも生体濃度に対応させたものでは ない。 フィブリノーゲンおよびトロンビンの濃度条件の設定には、ゲル化時間を主として考 慮した。いくつかの濃度の組み合わせで動的光散乱測定を行い、ゲル化時間を見積もっ た。その結果を表 3-2.1 に示す。ゲル化時間はフィブリノーゲン濃度およびトロンビン 濃度の両方に依存し、フィブリノーゲン濃度が高くなるにつれてゲル化時間は遅くなり、 一方、トロンビン濃度が高くなるにつれてゲル化時間は早くなった。また、ゲル化時間. 23.
(27) だけでなく形成されるフィブリンファイバーの凝集構造にも、フィブリノーゲン、トロ ンビンの各濃度に依存した変化が現れ、フィブリノーゲンおよびトロンビンの濃度が高 いほど、繊維の長さと質量の比を示すμ値は小さくなることがわかっている[11]。. 表 3-2.1 ゲル化時間のフィブリノーゲン濃度・トロンビン濃度依存性 Thrombin. 0.005 NIH units/ml. 0.01 NIH units/ml. 0.05 NIH units/ml. 1.0 mg/ml. 35 min. 20 min. 5 min. 2.0 mg/ml. 60 min. 30 min. 10 min. 3.0 mg/ml. 90 min. 40 min. 20 min. Fibrinogen. さらに、フィブリンゲルのネットワーク構 造のトロンビン濃度依存性も報告されている [12,13]。図 3-2.1 はトロンビン濃度変化にとも なうフィブリンゲルのネットワーク構造の変 化を共焦点レーザー顕微鏡で観察した報告例 である[13]。図 3-2.1 で観察したフィブリンゲ ルは、フィブリノーゲン濃度は 2.7 mg/ml で 一定とし、トロンビン濃度を(a)0.6, (b)0.1, (c)0.05, (d)0.03 NIH units/ml と変化させたもの である。トロンビン濃度が高いほど、観察さ れる繊維の数が増加することがわかる。 これらの結果から、トロンビン濃度が高い 場合には、プロトフィブリルの形成が瞬時に 進行しゲル化(ネットワーク形成)が早くなる ため、その後の繊維凝集(成長)は抑制される と考えられる。. 24. 図 3-2.1 フィブリンゲルのネットワーク 構造の共焦点レーザー顕微鏡観 察結果[13]。トロンビン濃度: (a)0.6, (b)0.1, (c)0.05, (d)0.03 NIH units/ml, フィブリノーゲン濃 度: 2.7 mg/ml 一定。.
(28) 糖(単糖・二糖) グルコース、マンノース、ガラクトース、マルトース(Wako Pure Chemical Industries)、 アロース、イソマルトース、セロビオース、トレハロース(Sigma-Aldrich)は、すべて濃 度が測定時に 10 mg/ml になるように PBS に溶解した。 単糖は、グルコース(図 3-2.2(A))を基準にアルドヘキソースの立体異性体を選んだ。 マンノース(図 3-2.2(B))はグルコースと C2 の立体配置が 1 つだけ異なるアルドヘキソー スの立体異性体の1つである。また、同様にグルコースと C4 の立体配置が異なるガラ クトース(図 3-2.2(C))、C3 の立体配置が異なるアロース(図 3-2.2(D))を用いた。 また、二糖はグルコース2分子からなるグルコシル-グルコース型の二糖を選んだ。 デンプンの酵素による加水分解で生じるマルトース(α-D-グルコピラノシル-(1→4)-Dグルコース, 図 3-2.3 (a))、その α(1→6)型のイソマルトース(図 3-2.3 (b))、セルロース中 の繰り返し単位であるβ(1→4)型のセロビオース(図 3-2.3 (c))、植物、昆虫、微生物など 天然に幅広く存在し、タンパク質を乾燥や凍結から守る生体保護機能を持つα,α(1→1) 型のトレハロース(図 3-2.3 (d))を用いた。. 溶媒 : Phosphate buffer saline(PBS) PBS(リン酸緩衝液;pH = 7.4)は組成、136.89 mM NaCl, 8.10 mM Na2H PO4, 2.68 mM KCl, 1.47 mM KH2PO4 のものを調製して使用した。. 25.
(29) (A)α-D-Glucose. (B)α-D-Mannose. CH2OH O H H OH. CH2OH. H OH. H. H. H. OH. OH. OH. H. OH H. CH2OH. O. H. H. OH. OH. H. H. H. (D)α-D-Allose. CH2OH H OH. O. OH H. (C)α-D-Galactose OH. H OH. OH. O. H H. H. H OH. OH. OH. 図 3-2.2 単糖の構造(Haworth 式). (a) Maltose Glc(α1-4)Glc H. CH2OH O H H OH. CH2OH O H H OH. O. OH. H. H. H. (b) Isomaltose Glc(α1-6)Glc. H. OH. OH. H. H. OH. OH. H. (c) Cellobiose Glc(β1-4)Glc H. CH2OH O H OH H. OH H. OH. O. H. CH2OH O H OH H. H H. CH2OH O H OH H. CH2 H O H. H OH. O H. OH. OH H. OH. H. OH. (d) Trehalose Glc(α1-1α)Glc OH. H. H. OH. OH. CH2OH O H H OH. H H. H O. H. OH. 図 3-2.3 グルコシル-グルコース型二糖の構造. 26. OH. H OH HOH2C O H. H OH.
(30) 〈測定法〉 まず、フィブリノーゲンとトロンビンのストック溶液をディープフリーザーから取り 出し、水の中に浸けて融解した。トロンビンはストック溶液を PBS で 1 NIH units/ml に 希釈した。フィブリノーゲンはストック溶液を PBS または糖溶液を加えて、2.0 mg/ml の濃度に調製し試料とした。加えた糖溶液は試料中の最終濃度が 10 mg/ml になるように PBS にて調製した。試料の 1/100 の量のトロンビン溶液を加え、最終濃度を 0.01 NIH units/ml とした。トロンビンを加えた時刻を t = 0 とし、経時変化を観測した。. [散乱光強度測定] 光散乱測定では光学的コンタミネーションを避けるため、トロンビンを加えるところ からクリーンボックス内で行った。トロンビンを混ぜた後、ポアサイズ 0.2 µm のシリン ジフィルターを用いてすばやく濾過し、セルに注入した。トロンビンを加えた後は、全 ての操作を可能な限り素早く行った。測定は、光源に Ar イオンレーザー(波長 488.0 nm) を用い、散乱角 30°、測定温度 37±0.01℃で行った。 動的光散乱(DLS)測定では高分子のゲル化過程において、 図 3-2.4 に示すような特徴的 な3 つの挙動が観測される[14,15]。 散乱強度はゲル化に伴い急激に増加し(図3-2.4 (A))、 自己相関関数における初期振幅(β値)の低下と長い緩和モードの出現が見られ(図 3-2.4 (B))、緩和時間分布関数の広幅化と長時間側へのピークのシフト(図 3-2.4 (C))が起こる。 フィブリノーゲンのゲル化もこの高分子のゲル化挙動に沿っており、ゲル化過程は散 乱強度変化と照らし合わせると次の様に解釈できる(図 3-2.5)。フィブリノーゲンがトロ ンビンによりフィブリンに変換し、互い違いに軸方向に結合してプロトフィブリルを形 成する過程:図 3-2.5(A)では、散乱強度は少しずつ増加する。次にプロトフィブリルが 結合していき 3 次元のネットワークを構築(ゲル化)していく過程:図 3-2.5(B)では、散 乱強度は急激に増加する。このことから、グラフの立ち上がり直後付近をゲル化点と定 義し、その時間をゲル化時間とする。例えば、図 3-2.5 のグラフにおいてゲル化時間は 約 40 分となる。 その後、 ゲル化がほぼ終了し、 ゲル全体のゆらぎが観測される:図 3-2.5(C)。 散乱強度は分子量に依存して増加する為、ゲル化に伴い散乱光強度が顕著に増加する。 本章では、図 3-2.5 に示した様に段階的に起こるフィブリノーゲンのゲル化過程への糖 添加の影響を、単糖と二糖の立体異性体について散乱光強度の経時変化から検討した。 27.
(31) (B) (A). (C). 図 3-2.4 動的光散乱測定によるゲル化過程(ゲル化点)の解析[1]。(A)散乱強度の経時変化、(B) 自己相関関数の経時変化、(C)緩和時間分布関数の経時変化。. (Ⅱ). フィブリノーゲン. プロトフィブリル FPB. 40000. (A). FPA. (B). 遊離したαCドメイン. Intensity [arbit. units]. (Ⅰ). 30000. (C) 20000. (B). 10000. (A) 0. (C) フィブリンファイバー (フィブリンゲル・ネットワーク構築). 0. 20. 40. 60. 80. elapsed time [min]. 図 3-2.5 フィブリノーゲンのゲル化模式図(Ⅰ)とゲル化過程で観測される散乱光強度変化 (Ⅱ)。 (A)フィブリンが互い違いに結合していく過程、(B)プロトフィブリルから 3 次元ネットワークの構築(ゲル化)、(C)ゲル全体のゆらぎ。. 28.
(32) [フィブリノペプチドの遊離測定(HPLC)] フィブリノーゲンはトロンビンによってフィブリノペプチド A 鎖(FPA)、B 鎖(FPB) が切断され、フィブリンへと変換することによりゲル化する。この FPA と FPB の切断 はゲル化過程において重要である。本章では、糖の添加時における FPA と FPB の遊離 の有無を HPLC により検出した。 HPLC 測定用の試料の調製には、散乱光強度測定に用いたフィブリノーゲン溶液とト ロンビン溶液と同様のものを用い、最終濃度はそれぞれ 2.0 mg/ml、0.01 NIH units/ml で ある。添加した糖の最終濃度も同様で、10 mg/ml とした。フィブリノーゲン溶液にトロ ンビンを添加し、37℃で6時間反応させ、完全にゲル化させた。そのゲルを約 3 分間煮 沸し、その後超遠心分離(20000×g, 1 hr)を行って上清を採取し、ポアサイズ 0.2 µm のシ リンジフィルターで精製したものを試料とし、HPLC 測定を行った。 HPLC は、ポンプ(LC-10Ai)、コミュニケーションバスモジュール(CBM-10A)、オン ラインデガッサー(DGU-4A)を備え付けた、島津 HPLC システムを用いて行った。解析 は、UV-VIS 検出器(SPD-A10i)によって行い、カラムは CAPCELL PACK C1 SG300(4.6 mm i.d. ×150 mm、Shiseido, Japan)を使用した。 フィブリノペプチドは、Kehl ら[16]が行った方法を改良して分離を行った。酢酸アン モニウム緩衝液の代わりに、0.1 % TFA(トリフルオロ酢酸)水溶液とアセトニトリル中 TFA 0.1 %の溶液を溶媒として使用し、このアセトニトリル中 TFA 0.1 %の溶液を 5 ~17.5 %に濃度変化させて、流速 1.0 ml/min で 25 分間分離を行った。検出は波長 215 nm において行った。 HPLC の結果から FPA と FPB のピークを特定するために、各ピークのアミノ酸シーケ ンスを、報告されている FPA と FPB のアミノ酸シーケンス[17]と比較した。今回比較に 用いたアミノ酸シーケンスを示す。. FPA EDGSDPPSGDFLTEGGGVR FPB PGA-FPTDYDEGQDDRPKVGLGAR (PGA ; pyro-glutamic acid). 29.
(33) [濁度測定] 本章ではさらに、ネットワークを構築するフィブリンファイバー(繊維)の太さや密度 への影響を、濁度の波長依存性より解析した。 濁度測定は、日立 U-2000A 分光光度計を用いて波長 400~800 nm の濁度を一定時間ご と(3 min または 5 min ごと)にスキャンした。濁度の解析法は、2-2 節に示した通りであ る。波長スキャンの 25 nm ごとの値を使用し、(2-7)式と(2-11)式を用いて、ゲルを構成す る繊維(フィブリンファイバー)の質量と軸方向の長さの比(μ)と繊維の半径(r)、そして 繊維の密度(μ/πr2)を算出した。 濁度解析におけるフィッティングの代表的な例を図 3-2.6 に示す。この図は、デキス トラン(分子量 77000)を添加した場合の解析例である。経過時間が短く成長の程度が低 いうちは(2-7)式で良く近似できるが、しだいに繊維が成長し、太くなってくると原点を 通る直線で近似できなくなり、(2-7)式での解析が不可能になる。図 3-2.6 (A)では、トロ ンビンを加えてから 15 分以上経過したときのプロットは、明らかに原点を通る直線に フィッティング出来ない。その場合には(2-11)式を用いて解析を行う。その解析結果が、 図 3-2.6 (B)である。19 分以上経過後は(2-11)式で良く近似されることがわかる。このよ うに、繊維の成長とともに(2-7)式から(2-11)式へと移行し、解析を行った。. 7. 3 min. τ. 5 4 3 2. 20. (B). 15 10 5. 1 0. 13 min 19 min 25 min 31 min 37 min 60 min. 25. 10-10C/τλ3. 6. 30. (A). 9 min 15 min 21 min 25 min 31 min 37 min 55 min. 0. 4. 8. 12. 16. 0. 20. 10-12/λ3. 図 3-2.6. 0. 1. 2. 3. 4. 5. 6. 7. 10-8/λ2. 濁度解析におけるフィッティングの例。プロットは各経過時間ごとの波長スキャ ンにより求めた濁度を用いて解析した値を示す。(A)は(2-7)式、(B)は(2-11)式によ る解析。. 30.
(34) [共焦点レーザー顕微鏡によるフィブリンゲル観察] 濁度解析の結果からは、ゲル化時間だけでなく形成されるフィブリンファイバーに対 する影響を検討することができる。本章ではさらに、形成されたフィブリンゲルを共焦 点レーザー顕微鏡(CLSM)により、ゲルを構成するネットワーク構造を観察した。 FITC (fluorescein isothiocyanate) FITC(図 3-2.7)は、蛍光試薬 Fluorescein に親タンパ. HO. O. O. ク基である NCS 基を結合させたもので、水に溶けて 強い黄緑色蛍光(λex=495 nm, λem=520 nm)を発する。ア. COOH. ミノ基と容易に反応する為、アミノ酸、ペプチド、タ ンパク質、酵素などの蛍光ラベルに適している。本研 NCS. 究ではフィブリンゲルをこの FITC で染色し、ネット 図 3-2.7. ワーク構造を CLSM で観察した。. FITC の分子構造. 測定は、北海道大学電子科学研究所ニコンイメージングセンターにて行い、電動倒立 顕微鏡 TE2000-E(Nikon)とリアルスペクトルイメージング共焦点レーザー顕微鏡システ ム C1si(Nikon)を用いて観察した。 観察するゲルは 35mm ガラスボトムディッシュに直接作成した。他の測定と同様に、 フィブリノーゲン溶液にトロンビン溶液に加えゲル化させる。糖を添加する場合も同様 で、フィブリノーゲン溶液と糖溶液を混合しておき、トロンビンを加えてゲル化させる。 完全にゲル化させる為に、数時間(4~6 時間程度)、37℃のインキュベータ内で静置する。 この間、試料が乾燥、蒸発しないように注意をしながら、時々ゲル化のチェックも行っ た。完全にゲル化させた後、5 μM の FITC/PBS 溶液を 5~6 時間かけて浸透させる。そ の後、PBS でゲルを洗浄し FITC 溶液を取り除き、さらに PBS に浸して数時間静置し、 余分な FITC を取り除いた。染色と脱色の操作もすべて 37℃のインキュベータ内で行い、 使用する FITC 溶液や洗浄・脱色用の PBS もすべて同インキュベータ内で 37℃に温め、 ゲルに与えるショックを最小限に止めるよう努めた。ゲルの調製、染色、脱色の操作は すべて北海道大学低温科学研究所 落合正則助教にご協力いただき、低温科学研究所に て行った。試料によってはシネリシスを起こしやすいものも多いため、電子科学研究所 への移動の際も保温し物理的な衝撃を与えないように細心の注意を払った。. 31.
(35) 3-3. 単糖の添加効果 単糖(グルコース、マンノース、ガラクトース、アロース;10 mg/ml)を添加したときの 散乱光強度の経時変化を、 糖を添加しないフィブリノーゲン-トロンビンのみの系(コント ロールと表記する)における結果と共に図 3-3.1 に示す。ゲル化点(ゲル化時間)は、散乱光 強度が急激に増加した直後の時間にほぼ対応している。コントロールは、約 35 分でゲル化 した。これに対して糖を添加した場合は、いずれもゲル化が観測され、グルコースは約 60 分、マンノースは約 70 分、ガラクトースは約 60 分、アロースは約 40 分で、ゲル化時間に 顕著な差が見られた。単糖(グルコース、マンノース、ガラクトース、アロース)添加によ る効果は、いずれもフィブリンのゲル化を遅延し、その遅延効果はグルコース添加と比較して マンノース添加の方が大きく、アロース添加では小さかった。以前、単糖(グルコース)・ オリゴ糖・多糖を用いてフィブリンゲル形成過程への糖添加による影響を調べた結果、グ ルコース添加によるゲル化の遅延効果が最も大きかったが[5-7]、単糖の中でもマンノース. intensity [ arbit. units ]. 60000 Control Glucose Mannose Galactose Allose. 50000 40000 30000 20000 10000 0 0. 50. 100. 150. 200. elapsed time [ min ] 図 3-3.1. フィブリノーゲンのゲル化過程における散乱光強度の経時変化。 糖はすべて最終濃度 10 mg/ml で添加。. 32.
(36) 添加による遅延効果がさらに大きいことが新たに明らかとなった。また、ゲル化後の散乱 光強度は、単糖やオリゴ糖の添加効果を調べた結果と同じ傾向を示し、ゲル化の遅延効果 が大きいほどゲル化後の散乱光強度は小さくなった。散乱光強度測定によるフィブリンゲ ル形成過程の解析により、添加した 4 種類の単糖の立体構造の違いがゲル化時間だけでな くゲル状態にも影響を与えることが明らかになった。. これまでの研究においてフィブリンゲル形成過程への影響を報告した単糖(グルコー ス)・オリゴ糖(マルトース、マルトテトラオース、マルトヘプタオース)・多糖(デキスト ラン)は、いずれもトロンビンとの相互作用は認められず、トロンビンの酵素活性に影響を 及ぼさないことが明らかになっている。このことから、これらの糖はフィブリノーゲンに 直接作用し、フィブリンゲル形成に影響を及ぼすことが示唆された[5-7]。本研究で新たに 検討している単糖の立体異性体添加による影響についても、これまでと同様にしてフィブ リノーゲンとトロンビンのどちらとの相互作用に起因するのかを確認する必要がある。そ こで、糖とトロンビンの相互作用の有無を確認するために、HPLC によりフィブリノペプ チドの遊離を検出した。 フィブリノーゲンはトロンビンによってフィブリノペプチド A(FPA)、B(FPB)が切断さ れ、フィブリンへと変換することによりゲル化する。この FPA と FPB の切断はゲル化過 程において非常に重要である。特に、フィブリノーゲンのゲル化を阻害する添加物に関し ては、トロンビンによるフィブリノペプチド遊離の有無を慎重に検討すべきである。当研 究室におけるβ-シクロデキストリン(β-CD)あるいはβ-シクロデキストリン硫酸塩(β-CD 硫酸塩)添加による影響の報告[8-10]を参考にすると、添加物が FPA と FPB の切断に影響し ている可能性があることがわかる。特に、今回のようなゲル化を遅延する添加物において は FPA と FPB 切断への影響も考えられ、FPA と FPB の遊離を確認することは、添加した 糖がゲル化の遅延という影響を及ぼす機構を検討する上で重要な情報となり得る。 フィブリノペプチド(FPA、FPB)の遊離を、HPLC を用いて検出した結果を図 3-3.2 に示 す。図 3-3.2 は上から順番にコントロール、グルコース、マンノース、ガラクトース、ア ロースを添加したときの結果である。コントロールでは、4 つのピークが観測された。4 つのうち、明確なアミノ酸シーケンスが確認できたのは、左から 1 番目と 3 番目のピーク である。これを 3-2 節に示したシーケンスと比較した結果、1 番目のピークが FPA のシー 33.
(37) ケンスと完全に一致した。しかし、3 番目のピークは前項の FPB のシーケンスとは若干異 なり、N 末端の 2 つのアミノ酸残基が欠落していた。この原因は現段階では不明である。. 250. Control. 200 150 100 50 0 250. Glucose. 200 150. absorbance at 215nm x103. 100 50 0 250. Mannose. 200 150 100 50 0 250. Galactose. 200 150 100 50 0 250. Allose. 200 150 100 50 0 0. 5. 10. 15. 20. 25. elapsed time [ min ]. 図 3-3.2 フィブリノーゲンのゲル化に伴うフィブリノペプチドの遊離検出(HPLC) の結果。上からコントロール、グルコース添加、マンノース添加、ガラク トース添加、アロース添加における結果。 糖の最終濃度は 10 mg/ml。. 34.
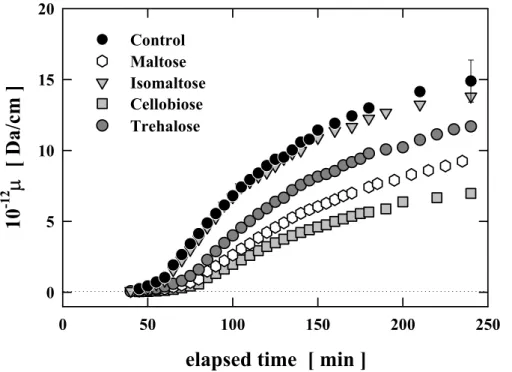
(38) グルコース、マンノース、ガラクトース、アロース添加のいずれの場合も、FPA および FPB の切断に関してはコントロールとの有意な差は見られなかった。この結果から、グル コースをはじめ、本研究で添加した単糖はすべてフィブリノペプチドの切断に全く影響を 及ぼしていないことが明らかとなった。 さらに、 これらの単糖はフィブリノーゲンと直接、 相互作用していると考えられるが、フィブリノーゲンへの作用部位は、FPA および FPB の 切断に関与していない部分であることが明らかとなった。. HPLC によるフィブリノペプチド遊離検出の結果は、添加した単糖はフィブリノーゲン に直接作用して、フィブリンゲルの形成に影響を及ぼしていることを強く示唆するもので あった。本研究ではさらに、ネットワークを構築するフィブリンファイバー(繊維)の太さ や密度への影響を、濁度の波長依存性より解析した。 散乱光強度測定と同様の条件で濁度測定を行い、ネットワークを形成する繊維について 調べた。(2-7)式と(2-11)式を用いて、単位長さ(L)あたりの質量(M)の値μ(M/L)の経時変 化を求め、 コントロールと糖を添加した場合とを比較した。 単糖を添加した場合の結果を、 図 3-3.3 に示す。. 10-12µ [ Da/cm ]. 20 Control Glucose Mannose Galactose Allose. 15. 10. 5. 0 0. 50. 100. 150. 200. 250. elapsed time [ min ] 図 3-3.3 フィブリノーゲンのゲル化に伴うμ値(M/L 比)の経時変化。添加した糖の最 終濃度はすべて 10 mg/ml。. 35.
(39) 図 3-3.3 から、コントロールのゲル状態における値が約 15×1012 Da/cm とわかる。フィ ブリノーゲン1分子が約 7.6×103 Da/nm であるから、フィブリノーゲンが約 200 倍(1500 / 7.6 ≅ 200)にまで凝集していることになる。すなわち、プロトフィブリルが約 100 本凝集 し繊維を形成していることになる。図 3-3.3 におけるμ値の経時変化は、図 3-3.1 に示した 光散乱の散乱光強度の経時変化の挙動に一致している。また、単糖を添加すると最終段階 の値がコントロールに比べ大きく減少するという結果も、光散乱測定での散乱光強度の変 化と一致していた。特に、ゲル化の遅延効果が最も大きかったマンノースではコントロー ルとの差が顕著に現れた。ゲル状態における値は約 2.5×1012 Da/cm であり、フィブリノー ゲンの約 30 倍まで凝集(250 / 7.6 ≅ 30)、すなわちプロトフィブリルが約 15 本凝集した繊 維を形成しているとわかる。この結果から、単糖の添加により繊維の成長(凝集)が顕著に 抑えられ、また、添加した 4 種類の単糖の立体構造の違いが形成されるフィブリンファイ バーの凝集構造に影響を与えていると推測できる。 図 3-3.4 と図 3-3.5 に、(2-11)式を用いて求めた繊維の密度(μ/πr2)と、繊維の半径 r の 経時変化を示す。コントロールでは、ゲル化に伴い繊維の密度、半径ともに増加する。単 糖を添加した場合にも、繊維の密度と半径の増加が見られるが、コントロールに比べてや や低い値を示し、ゲル化の遅延効果が最も大きいマンノースにおいては、その差は顕著で ある。最終段階における繊維の密度は、コントロールで 7×1022 Da/cm3、ゲル化の遅延効 果が小さかったアロースでは 5×1022 Da/cm3、それ以外のグルコース、マンノース、ガラ クトースはほぼ同じ程度で 3×1022 Da/cm3 と、いずれも単糖の存在下では形成された繊維 の密度が小さいことがわかった。また、繊維の半径 r の値を比較すると、コントロールは 8×10-6 cm (80 nm)、マンノースは 5×10-6 cm (50 nm)となり、マンノース存在下で形成され るフィブリンファイバーが細いことがわかる。 これらの結果から、グルコースを始めとするゲル化への遅延効果を示した単糖の存在下 では、コントロールに比べ、細くルーズな繊維が形成されることが明らかになった。単糖 を添加した場合は、ゲル化が遅延され、ゆっくりと時間をかけてゲル化が終了する。完全 にゲル化した後のゲルをコントロールのゲルと比較すると、単糖を添加したものの方が白 濁の度合いが弱い。これは目視でもはっきりと観察できるほど顕著である。この事実と濁 度の解析結果を合わせると、凝集が抑制されることにより細くルーズな繊維が形成される と同時に、繊維形成の不均一の度合いが小さいことが推測できる。 36.
(40) 10-22µ/πr2 [ Da/cm3 ]. 12 Control Glucose Mannose Galactose Allose. 10 8 6 4 2 0. 0. 50. 100. 150. 200. 250. elapsed time [ min ] 図 3-3.4 フィブリノーゲンのゲル化に伴うμ/πr2 値(繊維の密度)の経時変化。添加し た糖の最終濃度はすべて 10 mg/ml。. 14 Control Glucose Mannose Galactose Allose. 12. 106r [ cm ]. 10 8 6 4 2 0. 0. 50. 100. 150. 200. 250. elapsed time [ min ] 図 3-3.5 フィブリノーゲンのゲル化に伴う r 値(繊維の半径)の経時変化。添加した糖 の最終濃度はすべて 10 mg/ml。. 37.
(41) 濁度解析の結果から、単糖の添加がゲル化時間だけでなく、形成されるフィブリンファ イバーにも影響を与えていることが明らかになった。そこで、形成されたフィブリンゲル を FITC で染色し、共焦点レーザー顕微鏡を用いてゲルのネットワーク構造の観察を行っ た。この観察は、コントロールのゲルと、単糖(グルコース、マンノース、ガラクトース、 アロース)を添加して形成されたゲルについて行った。 フィブリノーゲン濃度とトロンビン 濃度、そして添加した単糖の濃度も散乱光強度測定や濁度測定と同じ条件でゲルの作成を 行った。ゲルの観察結果として、図 3-3.6 に顕微鏡写真を示す。図 3-3.6 A はコントロール、 B はグルコース添加、C はマンノース添加、D はガラクトース添加、E はアロース添加の フィブリンゲルの写真である。実際には FITC 染色により黄緑色の蛍光像として観察して いるが、ここでは白色で蛍光を示す。また CLSM の画像は蛍光像であり、ゲルネットワー クの実像を表すものではない。したがって、この画像から、ネットワークを形成する繊維 の太さなどを議論することはできない。しかし、この写真から各単糖の添加が形成される フィブリンゲルのネットワーク構造にも影響を与えていることが明らかになった。 図 3-3.6 の A と C を比較すると、C の方が繊維の数が圧倒的に多いことがわかる。マン ノースの添加で繊維の数が増加することは、濁度解析の結果(μ値)とも良く一致する。つ まり、マンノース存在下ではμ値が小さくなることから、フィブリノーゲンは凝集が抑制 されてコントロールに比べて少ない数のプロトフィブリルで繊維を形成するため、その結 果形成される繊維の数が増加するものと考えられる。また B、E、D でも C と類似した観 察結果となり、コントロールに比べ繊維数が多い。さらに、単糖を添加した B、C、D、E において観察されるネットワークは非常に細かく均一である。この結果も、濁度解析の結 果一致している。FITC の染色によるフィブリンゲルの CLSM 観察で、単糖の添加がネッ トワーク構造にも影響を与えることが明らかになった。. 本節では、フィブリンゲル形成に及ぼす単糖の添加効果を議論した。単糖(グルコース、 マンノース、ガラクトース、アロース)添加によるゲル化の遅延効果と、フィブリンファイ バーの凝集構造変化、さらにフィブリンゲルのネットワーク構造に対する影響は、単糖の 立体構造の違いに依存した影響(単糖の異性体効果)を及ぼすことが明らかになった。. 38.
(42) B. A. C. E. D. 図 3-3.6 フィブリンゲルの CLSM 画像。A コントロール、B グルコース添加、C マンノース 添加、D ガラクトース添加、E アロース添加。糖の最終濃度は 10 mg/ml。図中の白 いバーは 50μm を示す。. 39.
図


+4

関連したドキュメント
ガラス繊維補強セメント(GRC)は主として建築用
本研究は、tightjunctionの存在によって物質の透過が主として経細胞ルー
この小論の目的は,戦間期イギリスにおける経済政策形成に及ぼしたケイ
カウンセラーの相互作用のビデオ分析から,「マ
たRCTにおいても,コントロールと比較してク
哺乳類のヘモグロビンはアロステリック蛋白質の典
実際, クラス C の多様体については, ここでは 詳細には述べないが, 代数 reduction をはじめ類似のいくつかの方法を 組み合わせてその構造を組織的に研究することができる
および皮膚性状の変化がみられる患者においては,コ.. 動性クリーゼ補助診断に利用できると述べている。本 症 例 に お け る ChE/Alb 比 は 入 院 時 に 2.4 と 低 値
