Vol. 14, No. 2, 81–86, 2015
総 説(特集)
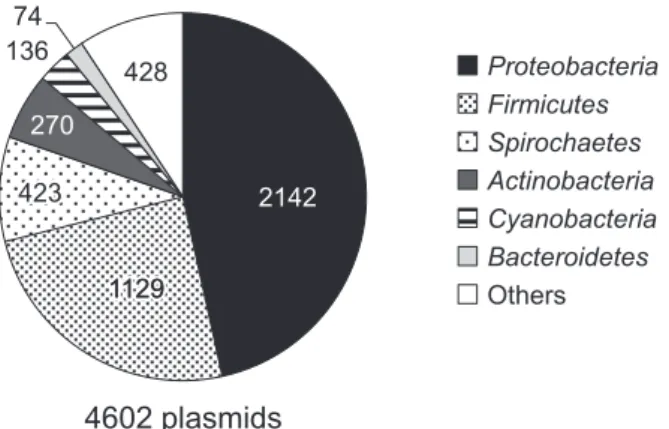
1. は じ め に プラスミドは微生物細胞内で,染色体 DNA とは物理 的に別個に存在する環状または線状の複製単位(レプリ コン)である。また,プラスミドは種々の微生物間を接 合伝達によって移動可能な遺伝因子である。プラスミド は,自身の複製・維持・伝達のために必要な遺伝子の他 に,プラスミドを有する宿主細胞に病原性,抗生物質耐 性能,物質代謝能等を付与できる。従って微生物は,プ ラスミドを介した遺伝子の水平伝播によって,新たな能 力を獲得し,様々な環境に対する急速な進化・適応能を もつと考えられている 1,7,8,11,20)。以上のことから,プラス ミドがどのような遺伝子をもつのか,どのような種類の 微生物を宿主にできるのか,という情報が,環境バイオ テクノロジー・医療分野において重要であることは,過 去の本学会誌でも述べてきた 25)。 近年の塩基配列解読技術の革新によって,微生物のゲ ノム配列の解読が急速に進むのに伴い,新たなプラスミ ドが見出され,その塩基配列が登録されてきた。2014 年 8 月の時点で米国国立生物工学情報センター(NCBI) のデータベースには,様々な種類のバクテリア,アーキ ア,真核微生物から見出された 4602 のプラスミドの全 塩基配列が登録されている。これらには,基本機能(複 製・維持・接合伝達)について詳細に研究されてきたプ ラスミドと,そうでないものが混在している。我々は, 今後ますます増加すると予想される全塩基配列の解読さ れたプラスミドについて,単純な塩基配列のみならず, その宿主や性質ごとに分類し,研究者が新たに見出した プラスミドや,プラスミド上の遺伝子が,どのようなプ ラスミドグループに属し,また水平伝播されうるのかに ついて,簡便に推定できるよう,データベースの整備を 進めている。本論文では,まだ途上ではあるがその一端 について紹介したい。なお,本データベースと,その解 析データの一部については,筆者らの最近の総説にも発 表している 18)。 2. 塩基配列が登録されたプラスミドとその宿主 プラスミドの特徴と,宿主の系統分類学上の関係は, プラスミドが微生物間をどのように伝播しているのか, を理解する上で重要である。これまでに塩基配列が解読 されたプラスミドの大半(4418)はバクテリアのもので あり,続いてアーキア(137),真核微生物(47)と続 く。バクテリアの分類については,Proteobacteria 門,Firmicutes 門,Spirochaetes 門,Actinobacteria 門,
Cyanobacteria門および Bacteroidetes 門が上位の宿主を 占め,これら 6 門で,これまでに全塩基配列を得られ て い る プ ラ ス ミ ド の 宿 主 の 90% 以 上 を 占 め て い た (Fig. 1)。ただし,この結果は,必ずしもこれらの門が プラスミドの宿主として主要であることを意味しない。 例 え ば, 多 く の 動 植 物 に 感 染 す る 病 原 菌 を 含 む Enterobacteriales科(927 プラスミド)や Pseudomona dales科(183 プ ラ ス ミ ド )( 以 上 Proteobacteira 門 ), あるいは Staphylococcus 属(220 プラスミド,Firmicutes 門),Borrelia 属(416 プラスミド,Spirochaetes 門)由 来のプラスミドが圧倒的に多かった。同様に植物との共 生細菌由来のプラスミド(Proteobacteria 門 Rhizobiales 科)も多く(143 プラスミド),登録されたプラスミド については,研究分野によって偏りが大きいことが示唆 された。従って,あるプラスミドが,どのような微生物
プラスミドゲノミクス~全塩基配列解読済のプラスミドデータベースの整備~
Genomics of Plasmids
新谷 政己
1,2*,金原 和秀
1 Masaki Shintani1,2* and Kazuhide Kimbara11 静岡大学大学院総合科学技術研究科工学専攻 〒 432–8561 静岡県浜松市中区城北 3–5–1 2 静岡大学創造科学技術大学院バイオサイエンス専攻 〒 432–8561 静岡県浜松市中区城北 3–5–1
* TEL & FAX: 053–478–1181 * E-mail: [email protected]
1 Department of Engineering, Graduate School of Integrated Science and Technology 2 Department of Bioscience, Graduate School of Science and Technology, Shizuoka University,
Hamamatsu, Shizuoka 432–8561, Japan
キーワード:プラスミド,宿主,データベース
Key words: plasmid, host, database
を宿主としうるのかを推定する際にも,偏りが生じる可 能性が高い(各門におけるプラスミドの詳細について は,前述の筆者らの総説 17) を参照して頂ければ幸いで ある)。 プラスミドの DNA としての性質(塩基長と GC 含量) には,宿主の種類や染色体 DNA との関係があると推定 される。まず,代表的な 6 門のプラスミドの塩基長につ いて,中央値とそのばらつきを比較した。その結果, Firmicutes門と Bacteroidetes 門における中央値が,他 の 4 門 と 比 べ て や や 小 さ か っ た(Fig. 2A)。 ま た, Spirochaetes門については,そのばらつきが小さかった (Fig. 2A)が,これは,先述したように本門の 423 プラ スミドのうち,416 が Borrelia 属細菌由来の非常によく 似たプラスミドが多く登録されているためと推定され た。一方,GC 含量については,各門のプラスミドごと のばらつきが大きく,中央値も異なることが示され,特 に Sphirochaetes 門 が 低 く(27.7%),Actinobacteria 門 が高かった(64.5%)(Fig. 2B)。既に西田が報告してい るように,大半のプラスミドの GC 含量は,その宿主 の染色体の GC 含量よりも低く,かつその違いは 10% 以内に留まる 16)。従って,プラスミドの GC 含量と,染 色体の GC 含量に,ある程度の比例関係があるために, Actinobacteria門(染色体の平均 GC 含量 64.1%)に由 来するプラスミドの GC 含量が高く,Sphirochaetes 門 (染色体の GC 含量 39.7%)に由来するプラスミドの
Fig. 1. The phylogenetic distribution of full-sequenced plasmids.
Fig. 2. Boxplots of plasmids’ sizes (A) and GC contents in major phyla (B). Comparisons of GC contents of host chromosome “C” and plas-mid “P” in host genera (C). Data are presented as median (horizontal solid line), 25th to 75th percentiles (box), maximum and minimum (error bars).
GC 含量が低いことが示唆された。プラスミドの GC 含 量が染色体に比べて低いのは,A や T よりも,G や C を維持するのにかかるエネルギーが高いので,生存に必 須でないプラスミドを維持するのには,少しでもコスト を下げる必要があるためではないかと推定されてい る 17)。一方,プラスミド自体が宿主内で複製・維持され るには,宿主の機能にある程度依存するため,プラスミ ド・染色体ともに GC 含量が同程度であるほうが望ま しいと予想される 17)。従って,プラスミドの GC 含量 は,宿主染色体の GC 含量へと近づく方向へと進化す ると考えられている 14,17)。これらのことから,プラスミ ドの GC 含量は,その宿主域を推定する際に重要であ るばかりでなく,宿主(あるいは類縁の種)の進化の過 程で,いつプラスミドを獲得したのかを推定するにも重 要と言える。Fig. 2C に各プラスミドの宿主に類縁の菌 株の染色体とプラスミド自体の GC 含量との違いを比 較 し た 結 果 を 示 す。 例 え ば,Proteobacteria 門 内 の Escherichia属や Salmonella 属細菌については,2014 年 8 月現在,それぞれ 63 株,46 株の染色体の完全長 DNA 配列が解読されており,また,各属に由来するプラス ミドは 310,160 の塩基配列が登録されている(ただし プラスミドの宿主全ての染色体 DNA 配列が解読され ているわけではない)。それらの染色体・プラスミドそ れぞれの GC 含量を比較したところ,双方の中央値は プラスミドの方が低く,またばらつきが大きかった (Fig. 2C)。これは,他の Pseudomonas 属,Burkholderia 属,Bacillus 属,および Staphylococcus 属細菌に関して も同様であった(Fig. 2C)。これらのうち,宿主染色体 と大きく異なる GC 含量を示すプラスミドは,比較的 最近,これらの属を宿主とした可能性がある。実験的な 裏づけが必要ではあるが,より多くのプラスミドと宿主 染色体について,データベースの整備が進めば,染色体 とプラスミドの GC 含量の違いから,その宿主や進化 の過程がある程度推測が可能になるかもしれない。 3. 塩基配列が登録されたプラスミドの分類 プラスミドは,その不和合性(incompatibility, Inc) や,接合伝達性(self-transmissible, mobilizable)を指標 として分類されてきた 3,9,10,13,19,20,22–24)。特に,同一の細胞 内におけるプラスミドの不和合性は,プラスミドを分類 する上で重要な性質であり,これまでに大腸菌を宿主と した場合(IncA~IncZ),主に Pseudomonas 属細菌を宿 主とした場合,(IncP-1~IncP-14),およびグラム陽性球 菌を宿主とした(Inc1~18)が提唱されている(昨年の 本学会誌に掲載された筆者らの総説 25) も参照)。最近 は,あるプラスミドがどの Inc 群に属するかを決める 際,不和合性試験よりも,各群の複製開始タンパク質 (Rep)をコードする遺伝子の塩基配列・アミノ酸配列 を基に分類する方法が一般的である。実際,物質代謝遺 伝子を搭載する主要なプラスミドを含む Inc 群,IncP-1,P-7,P-9 群を検出するために,Rep 遺伝子の配列に 基づいて縮重プライマーを作製し,検出に成功したとい う報告もなされている 6)。筆者らは,既存の Inc 群の Rep のアミノ酸配列クエリとし,全塩基配列が明らかになっ た 4602 のプラスミドに対して Local BLAST(TBLASTN) 解析を行った。その結果,1037 が上記 3 つの Inc 群の いずれかに属すると推定された。各宿主についてさらに 詳しく調べると,IncA-IncZ(IncA, B, F, G/U, H, I, K, L/M, N, P, Q, R, S, T, W, X, Z)に属すると推定される プラスミドの大半(402/423)が Enterobacteriales 科由 来 で あ っ た(Fig. 3)。 一 方,IncP-1~P-14(P-1, P-2, P-3, P-4, P-6, P-7, P-9)に属すると推定されるプラスミ ドは,Pseudomonadales 科の他に,Burkholderiales 科や
Enterobacteriales科由来であった(Fig. 3)。また,Inc1~ 18(Inc1, 4, 7, 8, 9, 10, 11, 13, 14, 18)に属すると推定さ れたプラスミドは,Lactobacilliales 科および Bacillales
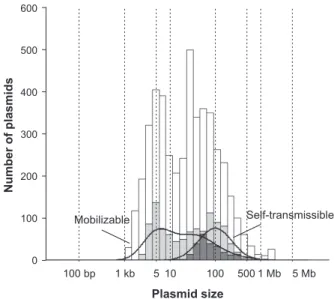
科に由来するプラスミドが主要であった(Fig. 3) 25)。
ところで,2014 年 4 月の世界保健機関(WHO)の報 告書にもあるとおり,これまで有効とされた抗生物質が
世界各地で効かなくなってきている。こうした多剤耐性 菌の蔓延にもプラスミドが寄与していると考えらえてお り,各菌株がどのようなプラスミドを保持するのかを迅 速に知る必要がある。そのため,Acinetobacter 属由来 のプラスミドは,Inc 群とは別に,Rep をコードする遺 伝子配列に基づき,GR1 から GR19 までに分類され, 各々のプラスミドを区別して検出するためのプライマー 配列も提案されている 2)。同様の理由で,病原菌として 知られる Staphylococcus や Enterococcus 属細菌由来の プラスミドについても,Rep の塩基配列を基に,26 のファ ミリーに分類され,PCR で簡便に判別可能なプライマー 配列も提案されている 4,5,12,15)。 以上の分類手法を含めると,塩基配列が解読されたプ ラスミドのうち,40%にあたる 1845 のプラスミドにつ いて分類がなされているが,次のような問題も生じる。 多くのプラスミドは複数の複製機構をもつため,どの Inc 群,あるいは Rep ファミリーに属するのかを一意に 決められない。また,こうした Inc や Rep については, 腸内細菌由来のプラスミドに情報が偏っているため,由 来の異なるプラスミドの分類は難しい。一方,近年,ス ペインの de la Cruz 博士のグループらによって,プラス ミドの接合伝達性に基づく分類手法が提唱され,上記の 問題をある程度緩和している 9,10,19)。プラスミドは,微 生物間の移動に必要な機能遺伝子が全て備わっている自 己伝達性のプラスミドと,自己伝達性プラスミドの接合 伝達に伴って移動可能な可動性プラスミドとに分けられ るが,双方のプラスミドとも,DNA の複製・移動を担 うタンパク質(MOB: mobilization)をコードする遺伝 子および接合伝達の開始点(oriT)を有する。この MOB に関しては,プラスミドの宿主を問わず,接合伝 達能を有するプラスミドが共通して保持するため,多く の種類のプラスミドについての分類が可能である。ま た,MOB 遺伝子を 1 つ以上もつプラスミドはほとんど ない。ただし,MOB に基づく分類は,接合伝達能をも たないプラスミドに対しては適用できない。自己伝達性 プラスミドは,MOB 遺伝子の他に,細胞間の接触を促 進するタンパク質(MPF: mating pair formation),IV 型 のカップリングタンパク質(T4CP),ATPase(VirB4) をもつ 19)。これらのタンパク質をコードする遺伝子群の 有無によって,そのプラスミドが自己伝達性(MOB, MPF,T4CP,VirB4 全て有する)か,可動性(上記の うち少なくとも MOB を有する)かどうかを推定できる。 既に報告されていることであるが,これまでに塩基配列 が解読されたプラスミドのサイズ分布は,4–8 kb と 32 kb 付近にピークをもつ二峰性を示す(Fig. 4)。可動 性プラスミドは,前半のピーク,後半のピーク双方に分 布しているが,自己伝達性プラスミドは,自己伝達に必 要な遺伝子数が多く,サイズが大きくなるため後半の ピークにのみ分布する(Fig. 4)。全 4602 のプラスミド のうち,自己伝達性と推定されるプラスミドが 485 (10.5%), 可 動 性 と 推 定 さ れ る プ ラ ス ミ ド が 824 (17.9%)と,全体の 28.4%が「動き得る」プラスミド と予想された(Fig. 5A)。こうしたプラスミドの宿主ご との分布は,自己伝達性プラスミドは Proteobacteria 門 にほとんどが分布し,可動性プラスミドについては,上 位 6 門いずれからも見出された(Fig. 5B)。グラム陽性 のバクテリアなど一部の微生物間のプラスミドの授受に ついては,必要な因子についてよくわかっていないた め,今後,自己伝達性プラスミドの分布についても変化 する可能性がある。 4. プラスミドデータベースの利用と今後の課題 宿主の種類およびプラスミドの性質に基づく分類を 行ったプラスミドのデータベースは,様々な利用が可能 である。例えば,注目しているタンパク質がどのような プラスミドにコードされているのかを調べる際に有効と 考えられる。筆者らは,これまでにプラスミドの異なる 宿主内における複製・維持・接合伝達能に深く寄与する と考えられる核様態タンパク質について研究を行ってき た。こうした核様態タンパク質をコードする遺伝子が, どのようなプラスミドにどれくらい分布しているのかに ついて,こうしたプラスミドデータベースを活用して調 べてきた 21)。その結果,接合伝達性のプラスミドの多く が,こうした核様態タンパク質をコードする遺伝子をも つことが明らかになった 21)。また,IncA/C(=IncP-3) 群に分類されるプラスミドは,全て核様態タンパク質 HU を共通して有することが示された(Shintani et al., 投稿中)。この他にも,例えば分解遺伝子群や抗生物質 耐性遺伝子がどのようなプラスミドによって「運ばれる のか」ということも,整備されたプラスミドのデータ ベースを用いることで容易に推定できる可能性がある。 今後は,日々増大する登録数と,また新たに明らかにさ れたプラスミドの複製・伝達に関する情報に対して迅速 に対応していくことが必要である。また,本データベー スは,プラスミドのみの情報に留まっており,宿主の染 色体の全塩基配列情報が登録された菌株も多いが,それ らの情報は反映していない。今後は,染色体の塩基配列 情報を利用できる菌株については,別個データベースを 作製することで,プラスミドと宿主の関係を考慮した解 析が可能であろう。
Fig. 4. Distribution of self-transmissible (dark gray) and mobiliz-able (light gray) plasmids according to Plasmid size.
謝 辞
本総説におけるプラスミドデータベースの整備は JSPS 科研費 2480087 の助成を受けて行った。
文 献
1) Aminov, R.I. 2011. Horizontal gene exchange in environmental microbiota. Front. Microbiol. 2: 158.
2) Bertini, A., L. Poirel, P.D. Mugnier, L. Villa, P. Nordmann, and A. Carattoli. 2010. Characterization and PCR-based repli-con typing of resistance plasmids in Acinetobacter baumannii. Antimicrob. Agents Chemother. 54: 4168–4177.
3) Carattoli, A. 2009. Resistance plasmid families in Entero
bacteriaceae. Antimicrob. Agents Chemother. 53: 2227–2238.
4) Carattoli, A., A. Bertini, L. Villa, V. Falbo, K.L. Hopkins, and E.J. Threlfall. 2005. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods. 63: 219–228.
5) Clewell, D.B. 2007. Properties of Enterococcus faecalis plas-mid pAD1, a member of a widely disseminated family of pheromone-responding, conjugative, virulence elements encod-ing cytolysin. Plasmid. 58: 205–227.
6) Dealtry, S., P.N. Holmsgaard, V. Dunon, S. Jechalke, G.C. Ding, E. Krogerrecklenfort, H. Heuer, L.H. Hansen, D. Springael, S. Zuhlke, S.J. Sorensen, and K. Smalla. 2014. Shifts in abundance and diversity of mobile genetic elements after the introduction of diverse pesticides into an on-farm biopurification system over the course of a year. Appl. Environ. Microbiol. 80: 4012–4020.
7) Frost, L.S., R. Leplae, A. Summers, and A. Toussaint. 2005. Mobile genetic elements: the agents of open source evolution. Nat. Rev. Microbiol. 3: 722–732.
8) Frost, L.S. and G. Koraimann. 2010. Regulation of bacterial conjugation: balancing opportunity with adversity. Future Microbiol. 5: 1057–1071.
9) Garcillán-Barcia, M.P., A. Alvarado, and F. de la Cruz. 2011. Identification of bacterial plasmids based on mobility and plasmid population biology. FEMS Microbiol. Rev. 35: 936– 956.
10) Garcillán-Barcia, M.P., M.V. Francia, and F. de la Cruz. 2009. The diversity of conjugative relaxases and its application in plasmid classification. FEMS Microbiol. Rev. 33: 657–687. 11) Guglielmini, J., L. Quintais, M.P. Garcillán-Barcia, F. De La
Cruz, and E.P. Rocha. 2011. The repertoire of ICE in pro-karyotes underscores the unity, diversity, and ubiquity of con-jugation. PLoS Genet. 7: e1002222.
12) Jensen, L.B., L. Garcia-Migura, A.J. Valenzuela, M. Løhr, H. Hasman, and F.M. Aarestrup. 2010. A classification system for plasmids from enterococci and other Gram-positive bacteria. J. Microbiol. Methods. 80: 25–43.
13) Lawley, T., B.M. Wilkins, and L.S. Frost. 2004. Bacterial con-jugation in Gram-negative bacteria, pp. 203–226. In G. Phillips and B. Funnell (eds.), Plasmid Biology, Washington, DC: ASM Press.
14) Lawrence, J.G. and H. Ochman. 1998. Molecular archaeology of the Escherichia coli genome. Proc. Natl. Acad. Sci. USA. 95: 9413–9417.
15) Lozano, C., L. García-Migura, C. Aspiroz, M. Zarazaga, C. Torres, and F.M. Aarestrup. 2012. Expansion of a plasmid classification system for Gram-positive bacteria and determi-nation of the diversity of plasmids in Staphylococcus aureus strains of human, animal, and food origins. Appl. Environ. Microbiol. 78: 5948–5955.
16) Nishida, H. 2012. Comparative analyses of base compositions, DNA sizes, and dinucleotide frequency profiles in archaeal and bacterial chromosomes and plasmids. Int. J. Evol. Biol. 2012: 342482.
17) Rocha, E.P. and A. Danchin. 2002. Base composition bias might result from competition for metabolic resources. Trends Genet. 18: 291–294.
18) Shintani, M., Z. Sanchez, and K. Kimbara. 2015. Genomics of microbial plasmids: classification and identification based on replication and transfer systems and host taxonomy. Front. Microbiol., doi: 10.3389/fmicb.2015.00242.
19) Smillie, C., M.P. Garcillán-Barcia, M.V. Francia, E.P. Rocha, and F. de la Cruz. 2010. Mobility of plasmids. Microbiol. Mol. Biol. Rev. 74: 434–452.
20) Sota, M. and E. Top. 2008. Horizontal gene transfer mediated by plasmids, pp. 111–181. In G. Lipps (ed.), Plasmids: Current Research and Future Trends Caister Academic Press: Horizon Fig. 5. Ratios of putative self-transmissible plasmids (black), putative mobilizable plasmids (gray), and others (white) (A) and those in each
Scientific Press.
21) Takeda, T., C.S. Yun, M. Shintani, H. Yamane, and H. Nojiri. 2011. Distribution of genes encoding nucleoid-associated pro-tein homologs in plasmids. Int. J. Evol. Biol. 2011: 685015. 22) Taylor, D.E., A. Gibreel, T.D. Lawley, and D.M. Tracz. 2004.
Anitibiotic resistance plasmids, pp. 473–491. In G. Phillips and B. Funnell (eds.), Plasmid Biology, Washington, DC: ASM Press.
23) Thomas, C.M. and A.S. Haines. 2004. Plasmids of the genus
Pseudomonas, pp. 197–231. In J.L. Ramos (ed.), Pseudomonas.
New York, USA: Plenum Publishing Corporation.
24) Udo, E.E. and W.B. Grubb. 1991. A new incompatibility group plasmid in Staphylococcus aureus. FEMS Microbiol. Lett. 62: 33–36.
25) 新谷政己,松井一泰,金原和秀,野尻秀昭.2013.環境中 におけるプラスミドの挙動解析.環境バイオテクノロジー 学会誌.13: 125–134.