Ⅱ . サンショウウオからみた丹沢
石原龍雄
1)・ 林 義雄
2)・ 草野 保
3)・ 山崎 泰
4)・ 北垣憲仁
5)Salamanders in the Tanzawa Mountains
Tatsuo Ishihara, Yoshio Hayashi, Tamotsu Kusano, Yasushi Yamazaki & Kenji Kitagaki
要 約 2004 年 3 月から 2006 年 1 月にかけて, 丹沢山地の 24 地点でハコネサンショウウオとヒダサンショウウオの幼生を中心に分 布状況を調べた. その結果, 両種とも 1993 年から 1995 年にかけて行った丹沢大山自然環境総合調査 (山崎ほか , 1997) 等に比べて出現個体数の減少が認められた.2004 年 2005 年共に豪雨による著しい出水に見舞われて沢が荒れ, シカによ る森林の荒廃と出水による減少とが区別し難い状況であるが, 沢の両岸の下層植生が失われ, 土壌の流失や落葉層の消失 など変態後の生活環境の悪化が認められた. また, 丹沢と同じく出水の被害を受けた箱根山地のハコネサンショウウオがあま り減少していないことを考慮すると, 森林の荒廃による影響がサンショウウオにも及んでいる可能性が高い. ヒダサンショウウオについては, 周辺の分布地と隔離され, 遺伝的にも異なる集団の可能性が考えられていたが, 今回の DNA の分析で東京都産の個体群と区別することができ, 遺伝的に異なる集団であることが確認された. 山梨県産の個体群と の比較も必要であるが, 同地域の生息地も荒廃しており, 延べ 13 回の採集にもかかわらず検体を入手できなかった. 1. はじめに 丹 沢 山 地 に は, ハ コ ネ サ ン シ ョ ウ ウ オ Onychodactylus japonicus (Houttuyn) とヒダサンショウウオ Hinobius kimurae Dunn の 2 種類のサンショウウオが分布している. 両種とも 勾配が急な源流域 (沢) で産卵し, 幼生期間を水中で過 ごした後, 変態して上陸後は森林で土壌動物の一員として 生活する. 成体は繁殖期の前に産卵場所に移動するが, 非繁殖期に発見されることは稀で, 陸上での生活はよくわ かっていない. このため, 生息の確認は通常幼生で行わ れる. 生息地は斜面の崩落や出水による被害が出やすい急峻 な地形で, 砂防堰堤などが作られやすい場所にあたる. 丹沢山地では, 関東大震災による地滑り等もあり, 砂防堰 堤工事等による影響も大きかったのではないかと考えられ るが, 分布に関する文献は, 斉藤ほか (1964), 酒匂川 文化財調査委員会 (1973), 柴田 (1979), 田辺 (1985) など部分的な分布資料しかなく, 変化を比較することができ ない.1993 年から 1995 年に行われた前回の調査 (山崎 ほか, 1997) により初めて丹沢全体の分布傾向を知ること ができた (図 1, 2). 170 地点延べ 212 回の調査によって 確認できたサンショウウオの生息地は 81 地点で, ハコネサ ンショウウオの生息地が 75 地点, ヒダサンショウウオの生息 地が 27 地点であった. 幼生の垂直分布は,ハコネサンショ ウウオが標高 420 ~ 1160m, ヒダサンショウウオが標高 420 ~1100m であった. 丹沢山地ではその後も自然環境の劣化が進み, 特にシ カによる荒廃が問題となった. 今回の調査では, 荒廃の 著しい東丹沢と, シカの密度がまだ低く比較的環境が保た れている西丹沢の沢を 3 本ずつ選び東西で比較するととも に, 前回の調査と比較することにした. また, 前回の調査 でヒダサンショウウオが出現した沢や生息している可能性の ある沢を踏査した. ほかに,1989 年にシカが出現して以来 目撃例が増えているものの下層植生への影響が見られない 箱根山地の生息地も比較のために調査した. 調査地点は 丹沢山地の 24 地点で延べ 43 回行った. 丹沢山地のヒダサンショウウオは, 分布の東限の個体群 の一つでもあることから (環境庁編, 1982), 保全する意 味の高い個体群であると考えられる. アロザイムによる生 物地理学的研究では, ヒダサンショウウオは主に 3 つのグ ループから成り,その一つである東部グループの分布域は, 全分布域に対して比較的狭く, 遺伝的に最も離れている (Matsui et al., 2000). この研究には丹沢山地の標本は含 まれていないが, 地理的な関係からは, 東部グループに 含まれると考えられる. ただし, 丹沢山地の個体群は, 東 京都や山梨県の産地に生息する個体群 (湯本, 2000; 草 野ほか, 2001) と分布が連続していない可能性もあることか ら (山崎ほか, 1997), 丹沢山地と周辺地域の個体群の遺 伝的な関係について改めて調査することは, 本種の保全を 考える上でも重要である. そこで, 丹沢山地と周辺地域に 生息するヒダサンショウウオについて, 保全遺伝学的な視 点からも調査を行った. なお, 魚類担当の勝呂尚之氏からは両生類の分布情報 の提供をいただいた. 八並俊聡氏には卒業論文のほか, 調査時の写真を多数提供いただいた. また, 下山良平氏 には, 木曽福島産ヒダサンショウウオの生育地をご教示い ただいた. 合わせて厚く御礼申し上げる. 2. 調査方法 サンショウウオの分布域である上流から源流を沢沿いに踏 査し, 幼生を中心に生息確認を行うとともに, 両生爬虫類 や環境の記録に努めた. また, 時間当たりの出現数からお よその生息個体数の多寡を知るため, サンショウウオが出 現した時点で時刻と個体数を記録し, 出現した場所に戻し た. ヒダサンショウウオについては,DNA 分析用の検体と して,幼生の尾端 3mm ほどを新しいカッターの刃で切除し, エタノール (> 99.5 %) 入りバイアルに保存した. その後, 幼生は発見した場所に戻した. このほかヒダサンショウウオ の繁殖生態を知るため, 成体の観察を行った. DNA の個体群間の比較のため, 東京 1) 箱根丹沢陸水研究会 2) 大阪府立大学大学院理学系研究科 客員研究員 3) 首都大学東京大学院理工学研究科 4) 箱根丹 沢陸水研究会 5) 都留文科大学 丹沢大山総合調査学術報告書 (2007)
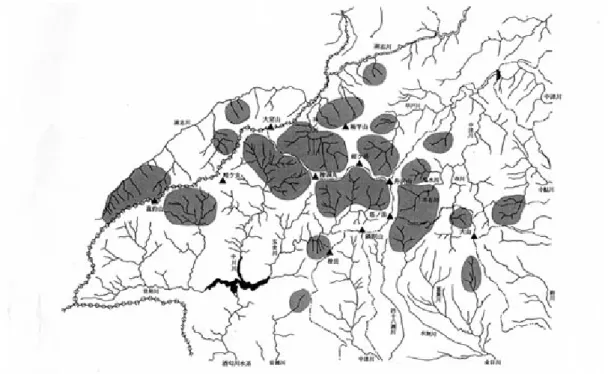
図 1. ハコネサンショウウオ分布図 ( 山崎ほか , 1997 を改変 ) 図 2. ヒダサンショウウオ分布図 (山崎ほか, 1997) 都の T1 沢 (日原川-多摩川水系), T2 沢 (成木川-荒 川水系),T3 沢 (多摩川-多摩川水系), T4 沢 (養沢川 -秋川-多摩川水系),T5 沢 (逆川-北秋川-多摩川 水系),T6 沢 (南秋川-多摩川水系),長野県の N1 沢 (木 曽川水系) からの検体も用いた.
DNA の抽出には DNeasy Tissue Kit (QIAGEN, 69504) を用い,付属のマニュアルに従い作業した.PCR およびシー ケンスは, タカラバイオ株式会社 ・ ドラゴンジェノミクスセン ターに委託した. 対象領域としては,mtDNA D-loop 領域
の 3' 端にある比較的変異に富む領域を選んだ (林 ・ 草 野, 2006). PCR には, 5'-CCT AAA TCT GAA CTA ACG GTG AG-3' (P1), 5'-CCA AAC CTT TAT GTT TCT GAG AT-3' (P2) のプライマーセットを用いた. これをテンプレー トとして,P1, P2 および 5'-GTG CTG TGC TTT AAT TAA GCT AC-3' (P3), 5'-TTT TTT TAC CTA TTG AAT TAT C-3' (P4) の各プライマーでシーケンスを行い, P3-P4 間 の分析を行った.波形データの処理および,系統解析には, MEGA3 (Kumar et al., 2004) を用いた.
3. 調査結果および考察 (1) 出現種類 調査期間中に, 両生類 9 種類, 爬虫類 8 種類, 魚類 2 種類等が出現した. 両生類 : ハコネサンショウウオ ・ ヒダサンショウウオ ・ アズ マヒキガエル ・ ヤマアカガエル ・ タゴガエル ・ ナガレタゴガ エル ・ シュレーゲルアオガエル ・ モリアオガエル ・ カジカ ガエル 爬虫類 : ニホンカナヘビ ・ ニホントカゲ ・ シマヘビ ・ アオ ダイショウ ・ ジムグリ ・ ヤマカガシ ・ シロマダラ ・ マムシ 魚類 : イワナ ・ ヤマメ その他 : カワネズミ ・ ムカシトンボが出現. (2) サンショウウオの出現状況 調査を行った 24 地点でハコネサンショウウオが 13 地点, ヒダサンショウウオが 4 地点 (うち 1 地点は魚類調査による 新生息地)で確認された.ハコネサンショウウオとヒダサンショ ウウオの混棲地は 3 地点であった. 前回の調査で出現した のに今回の調査で出現しなかった調査地点は, ハコネサン ショウウオで 4 地点, ヒダサンショウウオで 7 地点, 両種と も確認できなかった沢が 3 地点あった. また, 出現した地 点でも調査 1 時間当りの出現個体数も減少していた. 分布 の下限が上流に後退している生息地も 2 地点あった. 逆 に, 前回は工事中のため調査しなかった塩水沢でハコネサ ンショウウオが, 大洞沢ではヒダサンショウウオが出現した. 今回の調査と前回の調査の出現状況を表 1 に示す. 沢の 名称は保護のために記号で表す. 表中の出現個体数は, 調査 1 人 1 時間当りの出現数で, 採集時間が特定できな い場合は総出現数を ( ) で示す. 下層植生は, サンショ ウウオ成体の生息環境に関係する下層植生の多寡につい て,A : 多い ・ B : 少ない ・ C : ほとんど無い, で表記した. 底石の状態は, サンショウウオの幼生が潜む浮き石に注目 し,A : 浮き石が多い ・ B : 浮き石がある ・ C : 浮き石がほ とんど無い, で表記した. 斜面の石については, 上陸後の 幼体や成体が潜む場所として沢の斜面の石の存在に注目 し,A : 多い ・ B : 普通 ・ ほとんど無い, で表記した. なお, 丹沢の東西で比較することについては, サンショウ ウオの主要な分布地である中心部もすでにシカによる影響 を強く受けていたことや, シカの密度の低い西側の地域で は大又沢水系を除いてサンショウウオが分布する沢がない ことから比較できなかった. 今回の調査でヒダサンショウウオが出現した沢とハコネサ ンショウウオが比較的多い沢に共通しているのは, 河床に 石の堆積が多く, 沢の両岸にも石の堆積が多いことである. 繁殖移動期のハコネサンショウウオの成体や上陸直後の幼 体は, 沢沿いの石の下に潜んでいるが, 荒廃により林床植 生や落葉層が失われても堆積した石の下が生活場所となり うると考えられる. 岩の斜面では, シカが侵入できないため か, スズタケ等の植生が保たれていることもあった. このよう な沢は, サンショウウオの幼生が潜む場所や増水時に避難 する場所が多く,幼体や成体が潜む地中の隙間も多く,元々 サンショウウオにとって好適な環境であったとも考えられる. また, これらの沢も一様にこのような環境があるのではなく, 石の少ない場所や, 岩盤を樋のように流れる場所と滝が連 続する場所ではサンショウウオは出現しなかった.このほか, 1 支流では, 減 少が目立った. (3) 箱根山地との比較 箱根山地のハコネサンショウウオの生息地は, 丹沢のよ うに岩山で荒々しい沢ではなく, 土壌が多く石が比較的少 ない沢が多い. 沢沿いの小規模な工事でも淀みにシルト が厚く堆積したり, 流下した砂によって河床が埋まってハ コネサンショウウオが激減したり, 掘潜型を除く水生昆虫が 激減する現象が見られている (箱根生物研究会, 1981). 箱根山地でも 2005 年 8 月 25 日 1 時から 26 日 3 時の間 に 567.5mm の降水があり, 2002 年に続いて土石流が発 生するなど大きな被害を受けている. それにもかかわらず, 2006 年 1 月 4 日の T 沢では, 1 時間当り 18 個体出現し, 大型で肥満度の高い個体が出現した. 酒匂川水系の支流 である S 沢では, 2005 年 12 月 31 日に 1 時間当り 22.3 個 体,2006 年 7 月 27 日に 30 個体出現した. さらに須雲川 の流程が短い 1 支流では, 2005 年に八並俊聡氏によっ て高密度の生息が確認されている (図 5-12). さらに標識 放流による個体数推定では, 伏流区間を含む 230m から, 2006 年 5 月には 2363 個体, 2006 年 6 月には 2386 個体 という結果が得られている. 標識個体数 324 個体, 496 個 体に対し再捕獲個体は 68 個体, 121 個体と高く, 箱根生 物研究会 (1981) による 400m の区間で約 5000 個体とい う値に比べると,はるかに信頼度が高いと考えられる (八並, 2006). 酒匂川をはさんで丹沢山地に隣接する箱根山地では, 1989 年に仙石原でシカが目撃され, その後現在では広範 囲で記録されているものの下層植生や落葉層への影響は 認められていない. 箱根山地の生息地では, スズタケや草 本, 低木, 落枝落葉に覆われており, 丹沢のような異常 な状態ではない. 林床の乾燥化や落葉層の消失は上陸後 の幼体や成体にも影響を与えると考えられ (石原, 1990), 同様に出水の被害を受けた箱根山地に比べて丹沢山地の 減少が著しいのは, シカによる森林の荒廃による影響を示 すものと思われる. (4) ヒダサンショウウオの繁殖について 2005 年 1 月 5 日に, 中川川水系の L 沢においてヒダ サンショウウオの繁殖情況を調査したところ, 雄 2 個体 ・ 雌 1 個体の成体が出現し, 年内に産卵場に移動している ことが確認できた. この 1 ペアを水温 10℃で飼育したとこ ろ,2005 年 2 月 15 日に産卵した. 卵数は 15 卵 + 25 卵 = 40 卵であった. 前回の調査 (山崎ほか , 1997) では, 1995 年 5 月 17 日に, 発生の進んだ 10 卵 + 15 卵 = 25 卵から 20 卵 + 35 卵 = 55 卵が確認されている. 現地の 3 月の水温は 2~4℃のため, 実際の産卵は今回の人工産 卵より遅いと思われる.2005 年 6 月 8 日にはふ化幼生を 確認した. 標高の低い E 沢では, 1994 年 5 月 29 日にふ 化幼生を確認している. なお, ふ化時の全長は, 全長約 30mm であった. (5) ヒダサンショウウオ DNA の個体群間の比較 丹沢山地 3 地点から 9 個体, 奥多摩山地 6 地点から 6 個体, 木曽福島 1 地点から 2 個体のヒダサンショウウオ について,mtDNA の塩基配列が得られた. 対象領域の P3-P4 間を 317 bp にアライメントしたとき, トランジッション
������������� ���� ����� ����� ���� ����� ���� �� ���������� ���������� ��� � ���� ���������� ������������� ���������� ������������� �� ���������� � ��� � � ���������� ������������ �� ���������� ��������� ��� � � ���������� ������� �� ���������� � � ������������� ���������������� ������������ ���� ����� ����� ���� ����� ���� �� ���������� � � � � ���������� ������� �� ���������� �������� � � � ���������� ���������������� � � � ���������� ������� � � � ���������� ����������������� �� ���������� �������� � � � � ���������� ������ ����� ���� ����� ����� ���� ����� ���� �� ���������� ������� � � � ���������� ���������� ���������� ������� ���������� ������� ��������� ���� ����� ����� ���� ����� ���� �� ���������� ����������������� � � � ���������� ���������������� � � � ���������� �������� � � � ���������� ������ ���������� �������������� ��� ���������� �������� � � � ���������� ������� �� ���������� ����� �� ���������� �������� � � � ���������� ��������������� �� ���������� ��������� � � � ���������� ��������� ���������� ����������������� �� ���������� ���������������� � � � ���������� ���������������� ���������� �������������� ���������� �������������� ���������� ������������� ���������� ����� ���������� ������������ �� ���������� ������������ � � � ���������� ������� ���������� ����������� ��������� ���� ����� ����� ���� ����� ���� �� ���������� ������ � � � ���������� ��������� �� ���������� ������� � � � ���������� ����������������� ���������� ��������������� �� ���������� � � � � ���������� ���������������� ������������� ���� ����� ����� ���� ����� ���� �� ���������� ������ � � � ���������� ��������� ���� ���� ����� ����� ���� ����� ���� �� ���������� ��������� � � � ���������� ������� � � � �� ���������� ������� � � � �� ���������� ������� � � � 表 1. サンショウウオの出現状況 ハコネ : ハコネサンショウウオ ヒダ : ヒダサンショウウオ
7 個所, トランスバージョン 6 個所, 合わせて 13 個所の置 換, そして 4 個所のギャップがあり, 計 17 個所の変異サイ トが認められた (図 3). 丹沢山地内の 9 個体の遺伝子配 列には, 変異が認められなかった (ハプロタイプ E1). そ れに対し, 奥多摩山地からは,3 種類のハプロタイプが認 められた (E2, E3, E4). 木曽福島の 2 個体は, 同じ配列 であった (C1). 各地域のハプロタイプは異なり, 本調査 では, 計 5 種類のハプロタイプが確認された. 丹沢山地の ハプロタイプは, 奥多摩山地のものと比較し, 数塩基異なっ ていた. 同属であるトウキョウサンショウウオのデータ (林 ・ 草野, 2006) も加えて解析した UPGMA 法 (Unweighted Pair-Group Method with Arithmatic mean: 非加重結合法) による系統樹を図 4 に示した. 各枝には 1,000 回試行の ブートストラップ値 (%) を記した. 丹沢山地と奥多摩山地 の個体群間の遺伝的距離は近いが,224-G/A の 1 塩基の 置換により区別された (図 3). 両地域とは異なるグループ に属する木曽福島産 (Matsui et al., 2000) との遺伝的距 離は離れており, トウキョウサンショウウオで知られている南 北間の差異 (林 ・ 草野, 2006) 以上であった (図 4). 以 上の結果より, 丹沢山地の個体群は, 同じ東部グループ に属するものの, 近隣の奥多摩山地の個体群とは, 遺伝 的に区別可能であることが示唆された. 4. サンショウウオ類の保護について 今回の調査では陸上環境の荒廃が目立ち, サンショウウ オ類だけでなく, カエル類や陸産貝類の保護も含めて, 下 層植生を回復させる必要がある. 山崎ほか (1997) が指 摘したように砂防堰堤や護岸についても, サンショウウオ類 やカエル類に影響を及ぼしていると考えられ, 生息地であ る源流にはできるだけ設けず, やむなく作る場合はスリット 型など影響の少ない構造にする必要がある. 既存の構造 るなどの改善を図る必要がある. イワナについては,1988 年に酒匂川水系で移入個体が繁殖しているのが著者の 1 人山崎によって確認されて以来急激に分布を広げており, 源流部のサンショウウオ分布域にも放流されている. 栃本 (1993; 1996) によれば, イワナによるハコネサンショウウオ 成体の捕食, アマゴによるヒダサンショウウオ成体の捕食が 報告されており, 食害を防止するためにも生息地へのイワ ナ ・ ヤマメの密放流を禁止すべきである. サンショウウオの 減少が著しい場所については放流魚の駆除も必要である. 今回の調査では大きな出水が重なり, 前回の調査からの 変化を捉えるのが困難であった. 沢という変化の激しい環 境では 10 年という間隔は長すぎ, 調査範囲は狭くてももっ と短い間隔でモニタリングを続ける必要がある. ヒダサンショウウオの DNA 分析の結果からは, 丹沢山地 の個体群は近隣の個体群と区別され, 本個体群の持つ重 要性が改めて強調された. また, このような遺伝子レベル での差は, 生物地理学的な考察を行う上でも重要である. 今回調査された領域が, わずか 315 塩基程にすぎないこと から, 全ゲノム中には, より膨大な生物地理学的情報が記 録されており, それらは本種の起源や, 丹沢山地の個体 群の生い立ちについて知る手がかりになると予想される. し たがって, 丹沢山地の個体群の遺伝的固有性を守るため, 何らかの保全対策を講じる価値は高いと考えられた. また, 遺伝的多様性は, 個体群の健全さを計る尺度とし ても重要であると考えられている. 丹沢山地では, 奥多摩 山地よりも標本数が多かったにもかかわらず, 多型が認め られなかった. このことについては標本数よりも地点数が少 なかったことが関係している可能性がある. 一般的に, サ ンショウウオでは繁殖成功の年変動が大きく, 運良く生き 残った特定の雌が, 複数年産卵することにより個体群が維 持されていると考えられる上に (草野 ・ 川上, 1999), 移動 性も極めて乏しいため (草野 ・ 川上, 1999), 同じ地点で は, 特定のハプロタイプが優占する場合が多いのではない かと考えられる. 本調査では, 調査した地点数や標本数が 少なかったため, 丹沢大山地域においても, 複数のハプロ タイプが存在する可能性は大いにある. したがって, 遺伝 的多様性の現状を把握するためにも, 地点数を増やしての 調査が望まれる. L��n=5) H� (1) D� (3) T5� (1) T6� (1) T1� (1) T2� (1) T3� (1) T4� (1) N1� (2) �� �� �� �� ����� ������� ����� ��������� ������ ��������� ����� ��������� E1 E2 E3 E4 C1 Kimura 2-parameter ����� ����� ����� ������ 49 100 67 65 54 99 95 83 100 0.00 0.01 0.02 0.03 0.04 0.05 0.06 図 4. 丹沢山地と周辺地域のヒダサンショウウオの系統関係 ����������������������������������������������������������������������� ����������������������������������������������������������������������� ����������������������������������������������������������������������� ����������������������������������������������������������������������� ����������������������������������������������������������������������� ����������������������������������������������������������������������� ����������������������������������������������������������������������� ����������������������������������������������������������������������� ����������������������������������������������������������������������� ����������������������������������������������������������������������� ���������� ������� ����������������������������������� ����������������� ������������������������������������������������������������������������ ������������������������������������������������������������������������ ������������������������������������������������������������������������ ������������������������������������������������������������������������ ������������������������������������������������������������������������ ������������������������������������������������������������������������ ������������������������������������������������������������������������ ������������������������������������������������������������������������ ������������������������������������������������������������������������ �������������� ������� ������������������������������������������� ����� ������������������������������������������������������������������������ ������������������������������������������������������������������������ ������������������������������������������������������������������������ ������������������������������������������������������������������������ ������������������������������������������ ����������������������������� ������������������������������������������������������������������������ ������������������������������������������������������������������������� ������������������������������������������������������������������������ ������������������������������������������������������������������������ ����������������������������������������������� ����������������������������������������������� ������������������������������������������������ ����������������������������������������������� ���������� ��� ��� ��� ��� ��� ��� ��� �������� 図 3. ヒ ダ サ ン シ ョ ウ ウ オ の 各 ハ プ ロ タ イ プ の 遺 伝 子 配 列 (317bp にアライメント)
図 5-1. E 沢. 下層植生が失われ, 土砂が流下している. ヒダサン ショウウオは出現しなかった 図 5-2. 移入されたイワナ. 釣り人により, 滝や堰堤の上のサンショ ウウオの生息域にも放流されている 図 5-3. サンショウウオが出現しなくなった沢の斜面. 下層植生が失 われ, 上陸後のサンショウウオが潜む環境がほとんど無い 図 5-4. H 沢. ハコネとヒダがあまり減少していない沢. 下層植生も 残り, 河床や斜面の石の量も多い 図 5-5. L 沢. ハコネとヒダがあまり減少していない沢. 下層植生が 残り, 河床や斜面の石の量も多い 図 5-6. L 沢の斜面 下層植生が残り, 石も多く上陸後のサンショウ ウオが潜む場所が多い 図 5-7. D 沢. 減少が著しいもののハコネとヒダが出現した. 下層植生 はほとんど無いが,河床に石が多く,斜面にも石が多い場所がある. 図 5-8. ヒダサンショウウオ幼生 流水性サンショウウオであるが, 急 流への適応はハコネサンショウウオより劣る
図 5-9. D沢のヒダサンショウウオの生息場所. 土石流によって形成 されたように思われる. 涸れ沢からの石の供給も多い 図 5-10. 箱根山地の S 沢. 大きな出水が続いたが,ハコネサンショ ウウオはあまり減少していない. 石は多くないが下層植生は 豊富 図 5-11. 箱根山地の T 沢. 出水による河床の侵食が著しいが, 下層 植生や落葉層が豊富. サンショウウオはそれほど減少していな い. 図 5-12. 箱根山地の U 沢の 1 調査区で出現したハコネサンショウウ オ. 大きな出水にもかかわらず減少していなかった. (八並俊聡 氏撮影) 文 献 箱根生物研究会, 1981. ハコネサンショウウオ調査報告 . 箱根 町文化財研究紀要第 12 号 . 70pp. 箱根町教育委員会 . 林 義雄 ・ 草野 保 , 2006. ミトコンドリア遺伝子 D-loop HV2 領域に基づくトウキョウサンショウウオの地域間変異. 日本爬虫両棲類学報, 2006: 1-7. 石原龍雄, 1990. 箱根町の天然記念物 . ハコネサンショウウ オ. 33pp.,1pl. 大涌谷自然科学館 . 環境庁編, 1982. 日本の重要な両生類・は虫類の分布 (全 国版) . 第 2 回自然環境保全基礎調査 (緑の国勢 調査) 動物分布調査 (両生類 ・ は虫類) 報告書, 263pp. 環境庁 . 小池裕子 ・ 松井正文, 2003. 保全遺伝学 . 328pp. 東京大 学出版会, 東京 .
Kumar, S., K. Tamura & M. Nei, 2004. MEGA3: Integrated software for molecular evolutionary genetics analysis and sequence alignment. Briefings in Bioinformatics, 5: 150-163. 草野 保 ・ 川上洋一 , 1999. トウキョウサンショウウオは生き残 れるか?-東京都多摩地区における生息状況調査報 告書-. 69pp. トウキョウサンショウウオ研究会 , 東京 . 草野 保 ・ 植田健仁 ・ 初芝伸吾 , 2001. 東京におけるヒダサ ンショウウオとハコネサンショウウオの生息分布. 爬虫 両生類学会報, 2001: 1-7.
Matsui, M., Y. Misawa, K. Nishikawa & S. Tanabe, 2000. Allozymic variation of Hynobius kimurae Dunn
(Amphibia, Caudata). Comparative Biochemistry and Physiology Part B, 125: 115-125. 斉藤 晋 ・ 関根和伯 ・ 土屋清喜 ・ 北沢右三 , 1964. 丹沢山 水系の動物生態学的研究. 国立公園協会編 , 丹沢大 山学術調査報告書, pp.302-334. 神奈川県 . 酒匂川文化財調査委員会, 1973. 酒匂川流域動物調査報 告. 酒匂川文化財総合調査報告書 , pp.172-185. 神奈 川県教育委員会. 柴田敏隆, 1979. 動物分布調査報告書 (両生類 ・ は虫類) 神奈川県. 第 2 回自然環境保全基礎調査 . 16pp. 環 境庁. 田辺真吾, 1985. 丹沢の両生類 . 両生爬虫類研究会誌 , (32): 5-12. 栃本武良, 1993. アマゴに丸呑みされていたヒダサンショウ ウオ. 兵庫陸水生物 , (43): 42. 栃本武良, 1996. イワナに丸呑みされていたハコネサンショ ウウオ. 兵庫陸水生物 , (47): 36. 山崎 泰 ・ 石原龍雄 ・ 梶野 稔 ・ 北垣憲仁 , 1997. 丹沢のサ ンショウウオ類. 神奈川県公園協会 ・ 丹沢大山自然環 境総合調査団企画委員会編, 丹沢大山自然環境総合 調査報告書, pp480-492. 神奈川県環境部 . 八並俊聡, 2006. 箱根におけるハコネサンショウウオの生息 状況. 玉川大学農学部卒業研究論文 , 108pp. 湯本光子, 2000. 山梨県の両生類の分布について . 千葉中 央博自然誌研究報告特別号, (3): 23-32.