熊本大学学位論文
EGF 受容体および TGF- β 受容体を介した NTHi による TLR2 発現調節機構の解明
2006
熊本大学大学院薬学教育部 分子機能薬学専攻 遺伝子機能応用学分野
三上 史
Analysis of the signaling mechanism of NTHi-induced TLR2 expression via EGFR and TGF- β signaling
Fumi Mikami
目次
第一章 緒論‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥1 第二章 NTHi による TLR2 発現誘導に対する EGFR signaling の関与‥‥‥‥‥‥‥•7
第一節 序‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥7 第二節 実験結果‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥8 第一項 NTHi による TLR2 発現誘導に対する EGFR の影響
第二項 NTHi による TLR2 発現誘導を負に制御する EGFR 下流 signaling の検討 第三項 NTHi による TLR2 発現誘導に対する EGFR signaling 関連分子Src の影響 第四項 NTHi による TLR2 発現誘導を負に制御する EGFR-p38 MAPK signaling
に対する Src の作用点の検討
第五項 NTHi による EGFR 活性化機構の検討
第六項 in vivo での NTHi による TLR2 発現誘導に対する EGFR signaling の影響 第三節 考察‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥•25 第三章 NTHi による TLR2 発現誘導に対する TβR signaling の関与‥‥‥‥‥‥‥•28 第一節 序‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥•28 第二節 実験結果‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥•29 第一項 NTHi による TLR2 発現誘導に対する TβR の影響
第二項 NTHi による TLR2 発現誘導を正に制御する TβR 下流 signaling の検討 第三項 NTHi による TβR 活性化機構の検討
第四項 NTHi による TLR2 発現誘導を正に制御する TβR-Smad3/4 signaling に
対するNF-κBの関与の検討
第五項 NTHi による TLR2 発現誘導を正に制御する TβR-Smad3/4 signaling に 対する p38 MAPK signaling の関与の検討
第六項 NTHi による TLR2 発現誘導を正に制御する TβR-Smad3/4 signaling に 対する p38 MAPK 抑制因子,MKP-1 の関与の検討
第五章 実験方法の部‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥•53 第一節 細胞培養
第二節 NTHiの培養と抽出液の調整 第三節 Transfection
第四節 Promoter assay
第五節 RNA の抽出及び Quantitative Real-Time PCR 第一項 RNA の抽出
第二項 Quantitative Real-Time PCR 第六節 RNA-mediated Interference 第七節 Immunoprecipitation
第八節 Western Blotting 第九節 Neutralization Assay
第一項 ligand neutralization assay 第二項 receptor neutralization assay 第十節 Immunodepletion
第十一節 ELISA Assay
第十二章 Immunofluorescent Staining 第十三章 In Vivo Study
謝辞‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥•59 参考文献‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥‥•60
第一章 緒論
病原菌感染に対する宿主の免疫応答機構は,自然免疫と獲得免疫に大別される.獲 得免疫は T 細胞や B 細胞に担われ,細胞性免疫,液性免疫機構により非自己である多 様な病原菌に特異的に対応している.これに対し,自然免疫は非自己である病原菌を 上皮細胞,マクロファージが非特異的に認識し,サイトカインの産生,病原菌の貪食 などを誘起すると考えられてきた.しかし,近年,細菌やウイルス,真菌などの病原 菌が宿主の体内に侵入した際に Toll-like receptor(TLR)とよばれる一群の receptor が その構成成分を認識し,炎症性サイトカインの産生といった宿主自然免疫反応を惹起 することが明らかとなった1 – 6) .TLR member は一回膜貫通型の receptor で細胞外に leucine-rich repeats,細胞質内に Toll/IL-1 receptor homology(TIR)ドメインを持つ.現 在までに哺乳類において,11 の TLR member が同定されており,それら TLR member のリガンドを Figure 1 および Table 1 に示す7) .
Figure 1 TLRs recognize molecular pattern associater with bacterial.triacylated lipoprotein for TLR1; peptidoglycan for TLR2; double-stranded RNA for TLR3; lipopolysaccharide (LPS) for TLR4; flagellin for TLR5; diacylated lipoprotein for TLR6; imidazoquinoline and its derivative R-848 for TLR7; and bacterial unmethylated CpG DNA for TLR9.
Table 1 Toll-like receptors and their ligands7)
*It is possible that these ligand preparations, particularly those of endogenous origin, were contaminated with lipopolysaccharide and/or other potent microbial components, so more-precise analysis is required to conclude that TLRs recognize these endogenous ligands. N.D., not determined; TLR, Toll-like receptor.
TLR signaling の活性化は,TIR ドメインと MyD88 や TRIF などの細胞内アダプター分 子群との会合を引き起こす.その会合を通して,NF-κB や IRF などの転写因子が活性 化され,サイトカイン,ケモカインの産生や,抗原提示といった免疫応答を誘導する.
このような TLR signaling による免疫応答の活性化は,宿主獲得免疫反応の活性化にも 関与することが明らかとなっており,TLR signaling は宿主の感染防御に中心的な役割 を果たす.代表的な TLR signaling pathway を Figure 2 に示す8).
Figure 2 TLR signaling pathway8) .TLR signaling pathways originate from the cytoplasmic TIR domain. A TIR domain-containing adaptor, MyD88, associates with the cytoplasmic TIR domain of TLRs, and recruit IRAK to the receptor upon ligand binding. IRAK then activates TRAF6, leading to the activation of the IκB kinase (IKK) complex consisting of IKKα, IKKβ and NEMO/IKKγ. The IKK complex phosphorylates IκB, resulting in nuclear translocation of NF-κB, which induces expression of inflammatory cytokines. TIRAP, a second TIR domain- containing adaptor, is involved in the MyD88-dependent signaling pathway via TLR2 and
TLRs の中でも TLR2 に関しては研究が進み,Figure 1 および Table 1 に示したように,
細菌,真菌,寄生虫といった多様な構成成分を認識することが明らかとなっている.
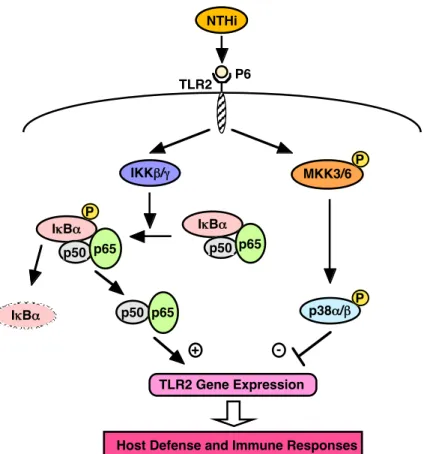
本研究室においても,TLR2 がグラム陰性バクテリアである Nontypeable Haemophilus influenzae(NTHi)の構成成分であるP6 を認識し,TLR2 signaling を活性化することを 示した. さらに,TLR2 の発現は NF-κB signaling pathway および p38 MAPK signaling pathway によって制御されており,TLR2 発現の増加は TNF-α,IL-1β,IL-8といった炎 症性サイトカインの発現を誘導するなどの知見を得た(Figure 3).したがって,
NTHi 感染時においても TLR2 は重要な役割を果たし,TLR2 の発現量の変化は宿主の
免疫応答と密接な関係があると考えられる 9 – 11) .
p38!/"
IKK"/#
I$B!
NTHi
I$B!
p50 p65 I$B!
p50 p65
p50 p65
TLR2 Gene Expression
Host Defense and Immune Responses MKK3/6
P
P P
TLR2
+ -
P6
Figure 3. Schematic representation of NTHi-induced signal transduction pathways involved in TLR2 expression in human epithelial cells. IKK, IκB kinase; NTHi, nontypeable Haemophilus influenzae; MKK, MAP kinase kinase; TLR, Toll-like receptor.
インフルエンザ桿菌 NTHi は,幼小児および成人における炎症性疾患に関与する病 原菌であり,幼小児期において,NTHi の感染は中耳炎を引き起こす上,慢性的な感染 は難聴の原因となる. 成人においては,世界の主な死亡原因のひとつである慢性閉塞 性呼吸器疾患を増悪することが知られている12) .主に感染に対する治療としては,抗 生物質が用いられるが,乱用による薬剤耐性菌の増加により,近年,効果的な予防薬,
つまりワクチンの開発が重要となってきた.インフルエンザ菌に対しては,莢膜の多 糖体を抗原とするワクチンが開発されているが,NTHi は無莢膜型のため,このワクチ ンは無効である.現在,ワクチン開発のターゲットとして P6 などの NTHi の共通抗原 である外膜タンパク質に注目し,ワクチンの開発が検討されているが,未だ臨床応用 には至っていない13 – 16) .また,NTHi は autolysis を起こしやすく,抗生物質処理など
により autolysis した後も炎症反応を続けて誘導することが知られている17) .興味深い
ことに,超音波処理によって得られる NTHi 抽出液および抗生物質処理により autolysis した NTHi を上皮細胞へ感染させると,Live NTHi と比較して NTHi による TLR2 発現 誘導をさらに促進する(Figure 4).
Relative Quantity of TLR2 mRNA
0 1 2
3
CON
NTHi
Mock LysateLive Live +
antibiotics
これは,autolysis 後の NTHi も炎症反応といった免疫応答を誘導するという報告と一致 するだけでなく,治療薬として使用される抗生物質は宿主の免疫応答を増強,あるい は過剰にするという可能性があることを意味する.つまり,NTHi の融解液中にも炎症 反応に関与する因子が含有されている可能性が高く,この因子による宿主の免疫応答 機構を解明することは,予防薬および治療薬の開発において重要であると考えられる.
NTHi 感染時における宿主の免疫応答機構としては, 前述したように,TLR2 signaling
を介した NTHi によるTLR2 の発現制御が関与している.しかしながら,autolysis を引
き起こした NTHi が TLR2 以外の receptor を介したシグナル伝達機構により,炎症反応 に影響を及ぼす TLR2 の発現誘導といった免疫応答に関与しているかは全く検討され ていない.そこで本研究では, NTHi 抽出液を用いて,未だ明らかとなっていない TLR2 以外の receptor が NTHi による TLR2 発現誘導に関与するかについて検討するこ
とで,NTHi 由来の因子,もしくは NTHi によって発現誘導される因子を推測し,その
因子が TLR2 の発現誘導といった宿主の免疫応答をどのように制御するのかについて
解明することを目的とした.
本研究において,第二章に示すように,EGFR の活性化は NTHi による TLR2 発現誘 導を負に制御すること,その signaling pathway には EGFR-Src-MKK3/6-p38 MAPK
signaling pathway が関与していることが明らかとなった.さらに,NTHi による EGFR
の活性化は transactivation によるものではなく,NTHi 由来の EGF-like factor が関与し ていることが示唆された.また,第三章に示すように,NTHi 由来の TGF-β-like factor によるTβRI/II-Smad3/4 signaling pathway の活性化は,NTHi による TLR2 発現誘導を正 に制御すること,また,そのメカニズムには NF-κB の活性化及び MKP-1 による p38
MAPK の不活性化という二つの signaling pathway が関与していることが明らかとなっ
た.このような TLR2 発現の厳密な制御は,宿主の免疫応答における TLR2 発現の制 御の重要性を示唆しており,この機構の一端を明らかとした本研究は,NTHi 感染時の 予防薬および治療薬の開発を検討する際の重要な知見のひとつであると考えられる.
第二章 NTHi による TLR2 発現誘導に対する EGFR signaling の関与
第一節 序
ErbB receptor family に属する EGFR は 170 kDa の細胞膜貫通型の receptor tyrosine
kinase であり,リガンド結合ドメイン,膜通過ドメイン,細胞内ドメインをもつ .
EGFR は上皮細胞を中心に,肝細胞,線維芽細胞,グリア細胞など広範な細胞に発現
しており,EGF family に属する EGF,TGF-α,amphiregulin 等がリガンドとして知られ ている.これらのリガンドの結合は EGFR の二量体化を促進し,tyrosine 残基の自己リ ン酸化を引き起こす.この自己リン酸化により,MAPK signaling pathway 等の signaling
pathway が活性化され,細胞増殖や分化,接着といった多様な細胞応答を誘導する18 –
21) .前述したように,元来 EGFR は EGF family の receptor として機能するが,興味深 いことに, EGFR が cytomegalovirus のようなウイルスの receptor としても機能するこ とが近年報告された22 – 25) .さらに,EGFR は Helicobacter pylori といったバクテリアに
よって transactivation されることも報告されている26) .しかしながら,このように
EGFR は EGF family だけでなく,様々なバクテリアやウイルスによっても活性化され
るにも関わらず,NTHi による TLR2 の発現誘導といった NTHi 感染時における宿主の 免疫反応への関与については明らかとなっていない.そこで本章では,EGFR signaling が NTHi による TLR2 発現誘導に関与しているかについて検討することを目的とした.
その結果,EGFR の活性化は NTHi による TLR2 発現誘導を負に制御すること,また,
その signaling pathway には EGFR-Src-MKK3/6-p38 MAPK signaling pathway が関与して いることが明らかとなった.さらに,NTHi による EGFR の活性化は transactivation に よるものではなく,NTHi 由来の EGF-like factor が関与していることが示唆された.
第二節 実験結果
第一項 NTHi による TLR2 発現誘導に対する EGFR の影響
EGFR は EGF family だけでなくバクテリアやウイルスによっても活性化されるにも
関わらず,バクテリアやウイルス感染時の炎症反応に影響を与える TLR2 の発現制御 といった宿主の免疫反応への関与については明らかとなっていない.そこで,EGFR
が NTHi による TLR2 発現誘導に関与しているかを明らかとするために,EGFR
signaling の阻害により NTHi による TLR2 発現誘導が変化するかについて,Quantitative real-time PCR(Q-PCR)を用いて検討した.その結果,NTHi による TLR2 発現誘導は
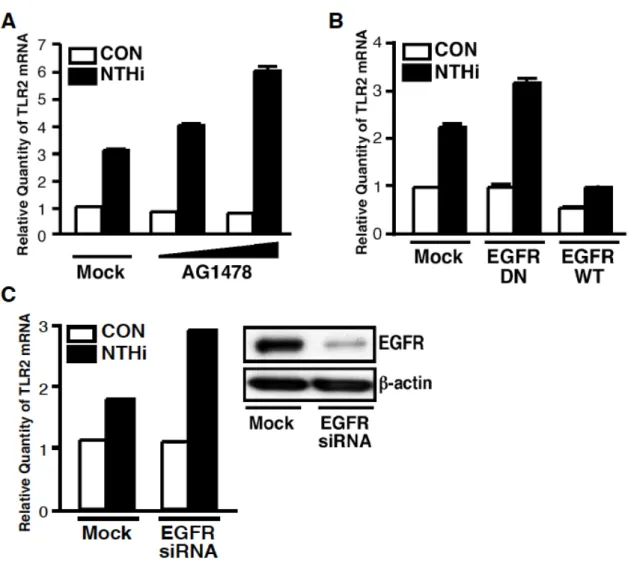
EGFR の特異的阻害剤である AG1478 の前処理により促進された(Figure 5A).また,
EGFR の dominant negative 変異体の強制発現は NTHi による TLR2 発現誘導を促進させ,
逆に,EGFR の強制発現はそれを阻害させた(Figure 5B).さらに,EGFR siRNA を 用いた EGFR の knockdown においても NTHi による TLR2 発現誘導は促進されるとい う同様の結果が得られた(Figure 5C).以上のことより,EGFR signaling は NTHi によ
る TLR2 発現誘導を負に制御することが示唆された.これらの結果をさらに確認する
ために,EGFR の発現が検出されない細胞株である MDA-MB453 (MB453)細胞と EGFR の発現が検出される 細胞株である MDA-MB468(MB468)細胞23, 27) を用いて,
NTHi による TLR2 発現誘導の相違を検討した.Figure 6A に示したように,NTHi によ
る TLR2 発現誘導は MB468 細胞と比較して MB453 細胞で促進されることが確認され
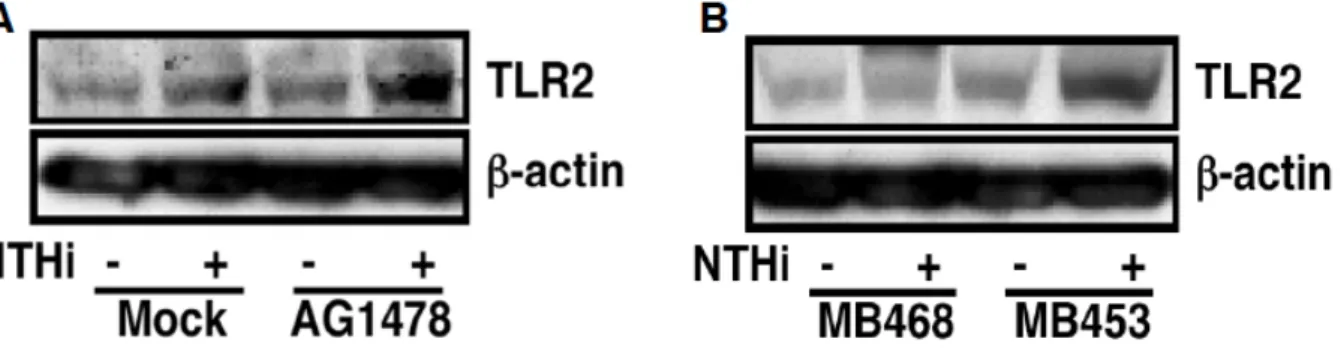
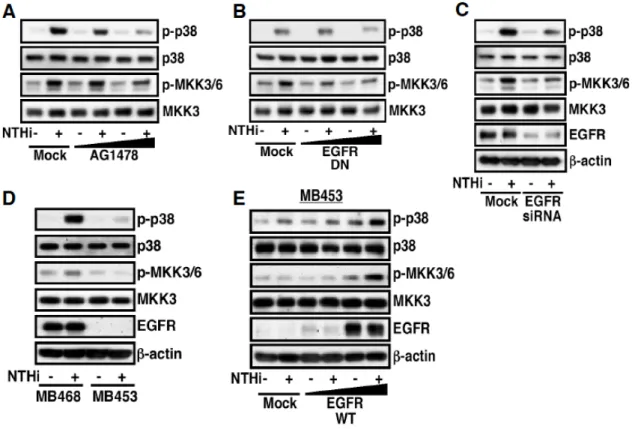
た.また,この細胞間での NTHi による TLR2 発現誘導の差異は MB453 細胞に EGFR を強制発現することで減少された(Figure 6B).次に,mRNA だけでなくタンパク質 レベルにおいても EGFR signaling が NTHi による TLR2 発現誘導を負に制御するかに ついて,Western Blotting を用いて検討した.その結果,NTHi による TLR2 タンパク質 発現誘導は AG1478 の前処理により促進した(Figure 7A).また,NTHi による TLR2 タンパク質発現誘導は MB468 細胞と比較して MB453 細胞で促進されることが確認さ れた(Figure 7B).以上のことより,EGFR signaling は NTHi による TLR2 発現誘導を 負に制御することが示された.
Figure 5 EGFR is a negative regulator for NTHi-induced TLR2 expression in HeLa cells.
A, pretreatment with AG1478 (1, 2 µM) for 1 h, a specific inhibitor for EGFR, enhanced NTHi- induced TLR2 expression at an mRNA level in HeLa cells. B, overexpression of a dominant- negative mutant of EGFR enhanced NTHi-induced TLR2 expression at an mRNA level, whereas overexpression of a wild type (WT) EGFR attenuated NTHi-induced TLR2 expression at an mRNA level in HeLa cells. C, transfection with EGFR-siRNA (100 nM) enhanced NTHi- induced TLR2 expression at an mRNA level. Quantitative real-time PCR were performed to assess TLR2 mRNA expression level after infection with NTHi for 3 h. Values are the mean +S.D.; n=3.
Figure 6 EGFR is a negative regulator for NTHi-induced TLR2 expression in MDA-MB cells. A, NTHi-induced TLR2 expression at mRNA level was markedly enhanced in EGFR- negative MDA-MB453 cells, as compared with that in EGFR-positive MDA-MB 468 cells. B, overexpression of a wild-type EGFR inhibited NTHi-induced TLR2 expression at mRNA level in MDA-MB453 cells. Quantitative real-time PCR were performed to assess TLR2 mRNA expression level after infection with NTHi for 3 h. Values are the mean + S.D.; n = 3.
Figure 7 EGFR is a negative regulator for NTHi-induced TLR2 protein expression. A, pretreatment with AG1478 (2 µM) for 1 h enhanced NTHi-induced TLR2 at the protein level in HeLa cells. B, NTHi-induced TLR2 expression at protein level was markedly enhanced in MDA-MB453 cells, as compared with that in MDA-MB 468 cells. In experiments, cells were treated with or without NTHi for 7 h. Protein lysates were then assessed by Western blotting using anti-hTLR2 antibody (H-175).
第二項 NTHi による TLR2 発現誘導を負に制御する EGFR 下流 signaling の検討
Figure 5~7 で示したように,EGFR signaling は NTHi による TLR2 発現誘導を負に制 御したことから,次にどの EGFR 下流 signaling pathway が負の制御に関与しているか について検討した.これまでに,p38 MAPK signaling が EGFR の下流 signaling pathway のひとつであること28 – 30) ,NTHi による p38 MAPK signaling の活性化は TLR2 の発現 を阻害することが報告されている10, 11).そこで,EGFR signaling は p38 MAPK signaling
を介して NTHi による TLR2 発現誘導を負に制御しているのではないかと仮説を立て
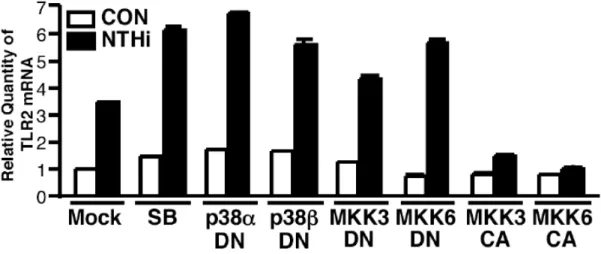
た.この仮説を証明するために,最初に p38 MAPK signaling が EGFR の下流 signaling pathway であること,及び NTHi による p38 MAPK signaling の活性化が TLR2 発現を阻 害することを確認した .EGFR のリガンドである EGF を細胞に処理し,p38 の活性化 を検討した結果,p38 のリン酸化が誘導された(Figure 8).また,Figure 9に示したよ うに,NTHi による TLR2 発現誘導は,p38 の特異的阻害剤である SB203580 の前処理 または p38α,p38β,MKK3,MKK6 の dominant negative 変異体の強制発現により促進 され,MKK3,MKK6 の constitutive active form の強制発現により阻害された.したが って,以前の報告通り確かに p38 MAPK signaling は EGFR の下流 signaling pathway で あること,NTHi による p38 MAPK signaling の活性化は TLR2 の発現を阻害することが 示された.
Figure 8 p38 MAPK signaling pathway is activated by exogenous EGF. Exogenous EGF (10 ng/ml) induced the phosphorylation of p38 in HeLa cells. In experiments, cells were treated with or without EGF for 15 min. Protein lysates were then assessed by Western blotting using anti-phosphorylated p38 antibody.
Figure 9 p38 MAPK signaling pathway is negatively involved in NTHi-induced TLR2 expression. SB203580 (1 µM) and overexpression of dominant-negative mutants of p38α, p38β2, MKK3 and MKK6 enhanced NTHi-induced TLR2 expression at mRNA level, whereas overexpression of constitutively active forms (CA) of MKK3 and MKK6 inhibited NTHi- induced TLR2 expression at mRNA level in HeLa cells. Quantitative real-time PCR was performed to assess TLR2 mRNA expression level after infection with NTHi for 3 h. Values are the mean + S.D.; n = 3.
次に,EGFR signaling が p38 MAPK signaling を介して NTHi による TLR2 発現誘導を負 に制御しているかについて,Q-PCR を用いて検討した.その結果,NTHi による TLR2 発現誘導は AG1478 または SB203580 単独の前処理においては促進されたものの,
AG1478 及び SB203580 の前処理では,SB203580 単独の前処理と比較して,それ以上
の増加は確認されなかった(Figure 10).したがって,EGFR 下流 signaling は p38 MAPK signaling であることが示唆された.
Figure 10 p38 MAPK signaling pathway acts downstream of EGFR in mediating the negative regulation of NTHi-induced TLR2 expression. Effect of AG1478 (2 µM) on NTHi- induced TLR2 expression at mRNA level in HeLa cells that were already pretreated with SB203580 (1 µM). Quantitative real-time PCR was performed to assess TLR2 mRNA expression level after infection with NTHi for 3 h. Values are the mean + S.D.; n = 3.
この可能性をさらに検討するために,EGFR signaling を阻害することで,NTHi によっ て誘導される p38 及び MKK3/6 のリン酸化が影響を受けるか,Western Blotting を用い て確認した.その結果,AG1478 の前処理,EGFR の dominant negative 変異体の強制発 現,siRNA による EGFR の knockdown による EGFR signaling の阻害は,NTHi によっ て誘導される p38 及び MKK3/6 のリン酸化を阻害した(Figure 11A~C).さらに,
NTHi によって誘導される p38 及び MKK3/6 のリン酸化は,EGFR の発現が検出されな
い MB453 細胞では確認されなかったが(Figure 11D),EGFR の強制発現により確認
されるようになった(Figure 11E).以上のことより,EGFR signaling は p38 MAPK
signaling を介して NTHi による TLR2 発現誘導を負に制御していることが示された.
Figure 11 EGFR is negatively involved in NTHi-induced TLR2 expression via a MKK3/6- p38 α/β MAP Kinase-dependent mechanism. A, pretreatment with AG1478 (1, 2 µM) inhibited NTHi-induced phosphorylation of both p38 and MKK3/6 in HeLa cells, as assessed by Western blotting using anti-phosphorylated p38 and MKK3/6 antibodies. B, overexpression of a dominant-negative mutant of EGFR attenuated NTHi-induced phosphorylation of both p38 and
また,TLR2 signaling は p38 MAPK を活性化すること,TLR2 は NTHi の receptor であることが報告されているため,EGFR だけではなくTLR2 を介した p38 MAPK
signaling の活性化も TLR2 発現の制御に関与しているかについて,TLR2 のリガンドで
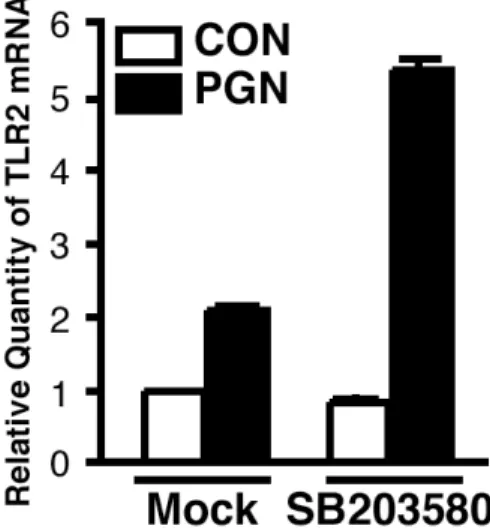
ある peptidoglycan(PGN)3, 7) を用いて検討した.SB203580 の前処理によりPGN によ って誘導された p38 MAPK signaling の活性化を阻害した結果,TLR2 発現誘導の増加 が確認された(Figure 12).したがって,TLR2 を介した p38 MAPK signaling の活性化
も TLR2 発現の負の制御に関与していることが示唆された.
Figure 12 Activation of p38 MAPK signaling pathway via TLR2 is negatively involved in TLR2 expression. Pretreatment with SB203580 (1 µM) for 1 h enhanced PGN (5 µg/ml)-induced TLR2 expression at mRNA level in HeLa cells.
Quantitative real-time PCR was performed to assess TLR2 mRNA expression level after infection with NTHi for 3 h. Values are the mean + S.D.; n = 3.
第三項 NTHi による TLR2 発現誘導に対する EGFR signaling 関連分子 Src の影響
EGFR signaling は p38 MAPK signaling を介して NTHi による TLR2 発現誘導を負に 制御していることが明らかとなったので,次に EGFR と MKK3/6-p38 MAPK を繋ぐ中 間分子を明らかとすることを目的とした.これまでに nonreceptor tyrosine kinase である c-Src が EGFR を介した MKK3/6-p38 MAPK signaling の活性化に関与していることが報 告されている31 – 33) .そこで,c-Src も NTHi による TLR2 発現誘導の制御に関与してい るのではないかという仮説をたてた.この仮説を証明するために,まず c-Src が NTHi によって活性化されるか Western Blotting を用いて検討した.Figure 13A に示したよう
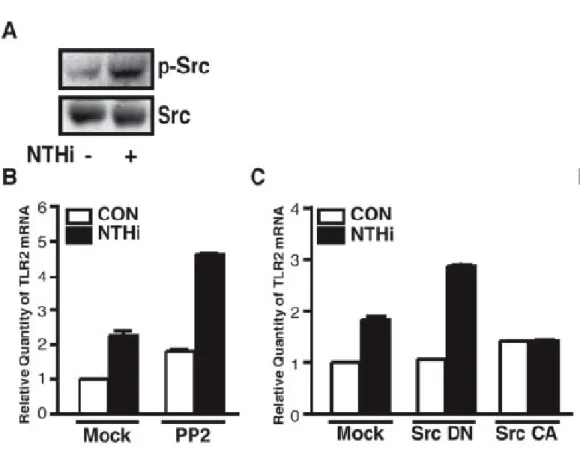
に, c-Src は NTHi 処理によってリン酸化され,活性化されることが示された.続いて,
この NTHi による c-Src の活性化が TLR2 の発現制御に関与しているか Q-PCR を用い て検討した.その結果,NTHi による TLR2 発現誘導は c-Src の特異的阻害剤である PP2 の前処理により促進された(Figure 13B).また,c-Src の dominant negative 変異 体の強制発現は NTHi による TLR2 発現誘導を促進させ,逆に,c-Src の強制発現はそ れを阻害させた(Figure 13C).以上のことより,NTHi による c-Src の活性化は TLR2 の発現誘導を負に制御することが示された.前述したように,c-Src は EGFR を介した MKK3/6-p38 MAPK signaling の活性化に関与していることから,c-Src が EGFR と
MKK3/6-p38 MAPK を繋ぐ中間分子である可能性が示唆される.
Figure 13 Activation of Src by NTHi is negatively involved in NTHi-induced TLR2 expression. A, NTHi induced the phosphorylation of Src, as assessed by Western blotting using anti-phosphorylated Src antibody. In experiments, cells were treated with or without NTHi for 5 min. B, pretreatment with PP2 (1 µM), a specific inhibitor for Src, enhanced NTHi-induced TLR2 expression at mRNA level in HeLa cells. C, overexpression of a dominant-negative mutant of Src enhanced NTHi-induced TLR2 expression at mRNA level, whereas overexpression of a constitutively active form of Src inhibited NTHi-induced TLR2 expression at mRNA level in HeLa cells. Quantitative real-time PCR were performed to assess TLR2 mRNA expression level after infection with NTHi for 3 h. Values are the mean + S.D.; n = 3.
第四項 NTHi による TLR2 発現誘導を負に制御する EGFR-p38 MAPK signaling に
対する Src の作用点の検討
c-Src が EGFR と MKK3/6-p38 MAPK を繋ぐ中間分子である可能性が示唆されたので,
次に c-Src が EGFR signaling の下流に位置するかについて,Western Blotting を用いて 検討した.まず最初に,EGFR の活性化により c-Src がリン酸化されるか,EGFR のリ ガンドである EGF 処理により確認した結果,c-Src のリン酸化が誘導された(Figure 14).
Figure 14 c-Src is activated by exogenous EGF. Exogenous EGF (10 ng/ml) induced the phosphorylation of c-Src in HeLa cells. In experiments, cells were treated with or without EGF for 5 min. Protein lysates were then assessed by Western blotting using anti- phosphorylated c-Src antibody.
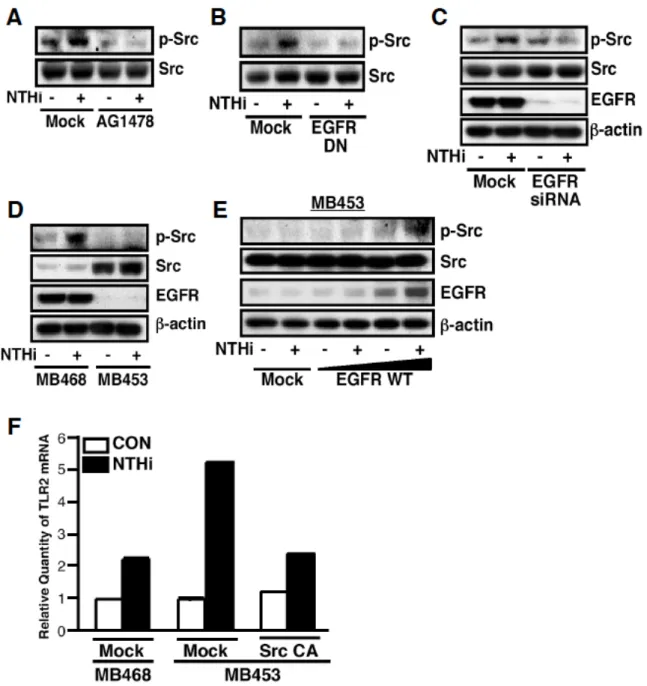
続いて EGFR signaling を阻害することで,NTHi によって誘導される c-Src のリン酸化 に影響を与えるか,Western Blotting を用いて確認した.その結果,AG1478 の前処理,
EGFR の dominant negative 変異体の強制発現,siRNA による EGFR の knockdown によ る EGFR signaling の阻害は,NTHi によって誘導される c-Src のリン酸化を阻害した
(Figure 15A~C).さらに,NTHi によって誘導される c-Src のリン酸化は,EGFR の 発現が検出されない MB453 細胞では確認されなかったが(Figure 15D),EGFR の強 制発現により確認されるようになった(Figure 15E).また,c-Src constitutive active
form を MB453 細胞に強制発現した結果,NTHi による TLR2 発現誘導を負に制御した
(Figure 15F).以上のことより, c-Src が EGFR signaling の下流に位置し,NTHi によ
る TLR2 発現誘導を負に制御していることが示された.
Figure 15 Src acts downstream of EGFR in mediating the negative regulation of NTHi- induced TLR2 expression. A, pretreatment with AG1478 (1 µM) for 1 h inhibited NTHi- induced phosphorylation of Src in HeLa cells, as assessed by Western blotting using anti- phosphorylated Src antibody. B, overexpression of a dominant-negative mutant of EGFR blocked NTHi-induced phosphorylation of Src in HeLa cells. C, transfection with EGFR-siRNA (100 nM) inhibited NTHi-induced phosphorylation of Src in HeLa cells. D, NTHi markedly induced the phosphorylation of Src in MDA-MB468 cells as compared with that in MDA-MB 453 cells. E, overexpression of a wild-type EGFR enhanced NTHi-induced phosphorylation of Src in MDA-MB453 cells. In experiments, cells were treated with or without NTHi for 5 min. F, overexpression of a constitutively active form of Src inhibited NTHi-induced TLR2 expression at mRNA level in MDA-MB453 cells. Quantitative real-time PCR was performed to assess TLR2 mRNA expression level after infection with NTHi for 3 h. Values are the mean + S.D.; n
= 3.
次に c-Src が MKK3/6-p38 MAPK signaling の上流に位置するかについて,Western Blotting を用いて検討した.その結果,PP2 の前処理,c-Src の dominant negative 変異 体の強制発現による EGFR-Src signaling の阻害は,NTHi によって誘導される p38 及び MKK3/6 のリン酸化を阻害した(Figure 16).以上のことより,EGFR-Src signaling は MKK3/6-p38 MAPK signaling の上流に位置し, NTHi による TLR2 発現誘導を負に制御 していることが示された.
Figure 16 Src acts upstream of MKK3/6-p38 pathway in mediating the negative regulation of NTHi-induced TLR2 expression. A, pretreatment with PP2 (1 µM) for 1 h inhibited NTHi- induced phosphorylation of both p38 and MKK3/6 in HeLa cells, as assessed by Western blotting using anti-phosphorylated p38 and MKK3/6 antibodies. B, overexpression of a dominant-negative mutant of Src inhibited NTHi-induced phosphorylation of both p38 and MKK3/6 in HeLa cells. In experiments, cells were treated with or without NTHi for 15 min.
第五項 NTHi による EGFR 活性化機構の検討
これまで EGFR-Src-MKK3/6-p38 MAPK signaling pathway が NTHi による TLR2 発現 誘導を負に制御することが明らかとされた.しかしながら,NTHi が EGFR を直接活性 化する,あるいは間接的に transactivation することによって TLR2 発現誘導が負に制御 されるのかは未だ明らかとなっていない.そこで,NTHi がどのように EGFR-Src-
MKK3/6-p38 MAPK signaling を活性化するかについて検討した.まず, NTHi によって
EGFR が実際に活性化されているかについて,immunoprecipitation を用いて検討した.
その結果,EGFR のリン酸化は NTHi 処理後 1 min から確認され,EGF 処理後と同様の バンドを得た(Figure 17).
Figure 17 NTHi phosphorylates EGFR like EGF does. EGFR was immunoprecipitated from HeLa cells after 1 min exposure to EGF (10 ng/ml) or NTHi using anti-EGFR or control IgG.
Immunoprecipitates were assessed by Western blotting using phosphotyrosine (p-Tyr) antibody.
EGFR のリン酸化が 1 min という非常に短い NTHi 処理時間によって得られたことから,
NTHi による EGFR の活性化は transactivation によって誘導されるのではなく,NTHi 由
来の EGF-like factor によって直接誘導されることが推測される.この仮説は,NTHi 処
理により EGF の発現が誘導されないこと,また,NTHi による TLR2 発現誘導は,
transmembrane EGF family precursors のshedding を引き起こす metalloproteinase の阻害剤
GM600134, 35) の前処理により変化しないこと,NTHi 処理後の培養溶液中で EGF タンパ
ク質発現量は変化しないことからも考えられる(Figure 18A~C).
Figure 18 NTHi does not induce EGF expression and transactivation of EGFR. A, NTHi did not induce EGF expression at mRNA level in both HeLa cells and MDA-MB 468 cells. B, pretreatment with GM6001 (1 µM) for 1 h, a specific inhibitor for MMP, did not affect NTHi- induced TLR2 expression at mRNA level in HeLa cells. Quantitative real-time PCR were performed to assess TLR2 mRNA expression level after infection with NTHi for 3 h. C, NTHi did not induce any detectable increase in EGF in the conditioned media of HeLa cells as assessed by EGF ELISA assay. Values are the mean + S.D.; n = 3.
次に NTHi 抽出液中に EGF-like factor 存在し,その NTHi 由来の EGF-like factor が NTHi による TLR2 発現誘導を負に制御するか,EGF neutralization assay を用いて検討 した.最初に EGF 中和抗体の特異性を確認するために,この EGF 中和抗体が EGF に
よる TLR2 発現抑制を中和するか検討した.その結果,EGF による TLR2 発現抑制は,
EGF を EGF 中和抗体で前処理することにより中和された(Figure 19A).次に,同じ EGF 中和抗体で NTHi を前処理した結果,NTHi による TLR2 発現誘導は HeLa 細胞及 び MB468 細胞において促進され(Figure 19B),EGFR の下流 signaling である c-Src,
MKK3/6,p38 の活性化は阻害されていることが確認された(Figure 19C).また,
NTHi 抽出液を EGF 抗体により immunodepletion した溶液で処理した細胞においても,
NTHi による TLR2 発現誘導は促進されていた(Figure 19D).したがって,NTHi 抽出
液中には EGF-like factor が存在すること,その NTHi 由来の EGF-like factor が NTHi に
よる TLR2 発現誘導を負に制御していることが示唆された.さらに,この NTHi 由来
の EGF-liike factor が EGFR を活性化し,NTHi による TLR2 発現誘導を負に制御するこ とを確認するために,EGFR neutralization assay を行った.その結果,EGFR 中和抗体 を用いて EGFR を中和したHeLa 細胞及び MB468 細胞においては,NTHi による TLR2 発現誘導は促進されていた(Figure 19E).以上のことより, NTHi 由来の EGF-like
factor が EGFR を活性化し,NTHi による TLR2 発現誘導を負に制御していることが示
された.
Figure 19 NTHi activates EGFR-Src-MKK3/6-p38 signaling pathway likely via an autocrine-independent mechanism. A, treatment with EGF inhibited TLR2 expression at mRNA level, whereas pretreatment with the EGF neutralizing antibody (25 µg/ml) counteracted the effect of EGF on TLR2 expression at mRNA level. B, pretreatment of NTHi lysates with the EGF neutralizing antibody (25 µg/ml) enhanced NTHi-induced TLR2 expression at mRNA level in both HeLa cells and MDA-MB468 cells. C, pretreatment of NTHi lysates with the EGF neutralizing antibody (25 µg/ml) also inhibited NTHi-induced phosphorylation of p38, MKK3/6 and Src in HeLa cells. D, TLR2 induction was enhanced in HeLa cells by the NTHi lysates, in which the EGF-like factors were immunodepleted using anti-EGF antibody. E, pretreatment of cells with the EGFR neutralizing antibody (1 µg/ml) enhanced NTHi-induced TLR2 expression at mRNA level in both HeLa cells and MDA-MB468 cells. Quantitative real-time PCR were performed to assess TLR2 mRNA expression level after infection with NTHi for 3 h. Values are
第六項 in vivo での NTHi による TLR2 発現誘導に対する EGFR signaling の影響
NTHi 由来の EGF-like factor が EGFR-Src-MKK3/6-p38 MAPK signaling を活性化し,
NTHi による TLR2 発現誘導を負に制御していることが in vitro の実験において明らか
となった.そこで,このシグナル伝達機構が in vivo においても NTHi による TLR2 発 現誘導の制御に関与しているか検討した.その結果,NTHi による TLR2 発現誘導は AG1478 及び PP2 の前処理により促進された(Figure 20A & B).したがって,in vivo においても EGFR-Src-MKK3/6-p38 MAPK signaling pathway が NTHi による TLR2 発現 誘導を負に制御することが確認された.
Figure 20 EGFR is a negative regulator for NTHi-induced TLR2 expression in vivo. A, pretreatment with AG1478 for 30 min greatly enhanced TLR2 induction by NTHi at mRNA level in the lung of BALB/c mice. B, pretreatment with PP2 for 30 min also greatly enhanced TLR2 induction by NTHi at mRNA level in the lung of BALB/c mice. Quantitative real-time PCR was performed to assess TLR2 mRNA expression level after infection with NTHi for 5 h.
Values are the mean + S.D.; n = 3.
第三節 考察
本章において,EGFR の活性化は NTHi による TLR2 発現誘導を負に制御すること,
また,その signaling pathway には EGFR-Src-MKK3/6-p38 MAPK signaling pathway が関 与 し て い る こ と が 明 ら か と な っ た . さ ら に ,NTHi に よ る EGFR の 活 性 化 は transactivation によるものではなく,NTHi 由来の EGF-like factor が関与していることが 示唆された.本章において得られた知見を Figure 21 に示す.
p38!/"
IKK"/#
MKK3/6
I$B!
NTHi
I$B!
p50 p65 I$B!
p50 p65
p50 p65
TLR2 Gene Expression
Host Defense and Immune Responses Src P
P
P P
TLR EGFR
+ -
Figure 21 Schematic representation of the signaling pathway involved in the negative
EGFR は細胞増殖や分化,接着といった多様な細胞応答に関与する receptor であるこ とはことはよく知られている.また,近年,EGFR がウイルスの receptor として機能す ることやウイルスにより transactivation されることが報告され,EGFR がウイルス感染 時において重要な役割を果たしていることが示唆されている22 - 25) .しかしながら,バ クテリア感染時における EGFR の役割についてはほとんど明らかとなっていなかった.
本研究は,バクテリアである NTHi が直接 EGFR を活性化し,炎症反応や免疫反応に
寄与する TLR2 発現を抑制することを明らかとした最初の知見であり,NTHi 由来の
EGF-like factor が EGFR を介して宿主の免疫反応を抑制している可能性を示唆している.
この機構の意義としては,現在のところ二つの可能性が推測される.最初の可能性と しては,NTHi が EGF-like factor を有し EGFR を利用することで,宿主の免疫反応を抑 制し,感染しやすい環境に変化させることが考えられる.二番目の可能性としては,
EGFR が NTHi の構成成分を認識することで,TLR2 の発現誘導により引き起こされる
過剰な炎症反応や免疫反応を抑制することが挙げられる.どちらにおいても,TLR2 の 発現の厳密な制御が,バクテリア感染時における宿主免疫反応の調節に不可欠である ことは間違いなく,感染における炎症反応及び免疫反応の分子機序を解明する上で有 用となると考えられる.
Figure 12 において,EGFR だけでなくTLR2 を介した p38 MAPK signaling の活性化も
TLR2 発現の負の制御に寄与していることが示唆された.では,NTHi による TLR2 発
現誘導の負の制御は,EGFR,TLR2 のどちらの receptor を介した機構によって主に制 御されているのだろうか? まず,ひとつの可能性として,上皮細胞における各々の発 現量が関わっていると推測される.したがって,EGFR は上皮細胞に広く発現してい るが,非感染時においては上皮細胞での TLR2 の発現は非常に低いため,感染初期に
おけるTLR2 発現制御への TLR2 自身の関与の可能性は低いと考えられる.つまり,
TLR2 の発現誘導は TLR2 自身によっても制御されているが,この機構は TLR2 の発現 が誘導された後である感染後期に寄与すると考えられる.次に,別の可能性として,
各々のリガンドとなる NTHi の分子の存在場所も関わっていると推測される.TLR2 の リガンドとしては,P6 という outer membrane protein が知られており9) ,Live NTHi 及 び 抗生物質などの要因により autolysis された NTHi どちらにおいても,TLR2 signaling
は活性化されると考えられる.EGFR のリガンドは,本研究で示唆したように NTHi 由 来のEGF-like factor であるが,Figure 4 で示したように, NTHi による TLR2 発現誘導 は NTHi 抽出液及び抗生物質により autolysis された NTHi 処理でのみで観察され,Live
NTHi 処理では観察されないため,TLR2 の発現抑制作用をもつ EGF-like factor は
cytosolic fraction に存在すると考えられる.つまり,EGFR が NTHi による TLR2 発現 誘導を負に制御するためには,NTHi が autolysis されることが必要であると考えられる.
Figure 19 において,NTHi 中にEGF-like factor が存在し,TLR2 の発現を抑制してい ることが確認されたが,どの NTHi の構成成分が EGF-like factor であるかは不明のまま ある.今回使用した NTHi strain 12 は未だ全 genome sequence が明らかとなっていない ため,EGF の conserved sequence を用いた homology 検索による NTHi 由来のEGF-like
factor の同定は不可能であったが,immunodepletion によりNTHi 抽出液から免疫沈降で
得られる EGF-like factor の protein sequence analysis を行うなどして, NTHi 由来のEGF-
like factor の同定を今後検討する必要があろう.
第三章 NTHi による TLR2 発現誘導に対する TβR signaling の関与
第一節 序
TGF-β family の receptor である TβR は serine / threonine kinase 活性をもつ一回膜貫通 型の receptor で,I 型と II 型に分類される. TβRI は53 kDa, TβRII は 70 kDa であり,
共に細胞外ドメイン,膜通過ドメイン,細胞内ドメインをもつ.TβRII にリガンドが 結合すると TβRI,TβRII はヘテロ四量体を形成し,TβRII は TβRI の細胞内傍細胞膜領 域の glycine,serine,threonine に富んだ配列(GC 領域)をリン酸化して活性化する.
活性化された TβRI はその下流の因子である Smad を認識してリン酸化し,細胞増殖,
分化,アポトーシス といった多様な細胞応答を制御する37 – 42) .TβR signaling は炎症反 応や免疫反応の制御にも関与しており,カビの一種である Cryptococcus neoformans に 感染したラットにおいて, TβR signaling の活性化は lysozyme の産生を誘導し,抗菌効 果を示すことが報告されている43) .一方, Trypanosoma cruzi,Leishmania species とい った病原菌感染による TGF-β の過剰産生は マクロファージの活性化を抑制するという 報告もされている44, 45) .すなわち,TβR signaling の活性化はマクロファージにおける 抗菌効果を促進,あるいは抑制するという二面的な機能を有していると考えられる.
また,近年,グラム陰性バクテリアである NTHi が TGF-β-like factor を有し,上皮細胞 における感染時の初期免疫反応であるムチンの産生を制御していることが明らかとな
った46, 47) .しかしながら,バクテリアやウイルス感染時の宿主の防御反応のひとつで
ある TLR の発現制御といった免疫応答への関与については明らかとなっていない.そ
こで本章では,TβR signaling が NTHi による TLR2 発現誘導に関与しているかを検討 することを目的とした.その結果,NTHi 由来の TGF-β-like factor によるTβRI/II- Smad3/4 signlaing pathway の活性化は,NTHi による TLR2 発現誘導を正に制御するこ と,また,そのメカニズムには NF-κB の活性化とMKP-1 による p38 MAPK の不活性 化という二つの signaling pathway が関与していることが明らかとなった.
第二節 実験結果
第一項 NTHi による TLR2 発現誘導に対する TβR の影響
TβR signaling は多様な細胞応答に関与しているにもかかわらず,バクテリアやウイ
ルス感染時の宿主の防御反応のひとつである TLR2 の発現制御といった免疫応答への 関与については明らかとなっていない.そこで TβR が NTHi による TLR2 発現誘導に 関与しているかを明らかとするために, TβR signaling の阻害により NTHi による TLR2 発現誘導が変化するかについて,Quantitative real-time PCR(Q-PCR)を用いて検 討した.その結果,NTHi による TLR2 発現誘導は TβRI の特異的阻害剤である SB431542 48 – 51) の前処理または TβRI,TβRII の dominant negative 変異体の強制発現に より阻害され, TβRI,TβRII の強制発現により促進された(Figure 22A & B).また,
TβRI,TβRII の siRNA を用いた TβRI,TβRII の knockdown においても NTHi による TLR2 発現誘導は阻害されるという同様の結果が得られた(Figure 22C & D ).以上の ことより,TβR signaling は NTHi による TLR2 発現誘導を正に制御することが示唆さ れた.これらの結果をさらに確認するために,Mv1Lu 細胞株由来で,TβRI が検出さ れない R1B 細胞と, TβRII が検出されない DR26 細胞を用いた.まず,R1B 細胞及び
DR26 細胞では,TβR signaling が伝達されないことを確認するために,Smad-binding
elements(SBE)-luciferase52) を用いて promoter assay を行った.その結果,TGF-β1 処 理による promoter 活性の増加は R1B 細胞,DR26 細胞においては検出されなかった.
次に,TLR2-luciferase53) を用いた promoter assay により これらの細胞におけるNTHi に よる TLR2 発現誘導の相違を検討した.Figure 23A に示したように,NTHi による TLR2 発現誘導は,R1B 細胞および DR26 細胞では確認されなかった.また,この細胞 間での NTHi による TLR2 発現誘導の差異は,R1B 細胞及び DR26 細胞 に TβRI,
TβRII をそれぞれに強制発現することで回復した(Figure 23B).以上のことより,
Figure 22 TGF-β receptor signaling is required for NTHi-induced TLR2 expression. A, SB431542 (1µM), a specific inhibitor for TβRI, and overexpression of a dominant-negative mutant of TβRI attenuated NTHi-induced TLR2 expression at mRNA level, whereas overexpression of a wild-type TβRI enhanced NTHi-induced TLR2 expression at mRNA level in HeLa cells. B, overexpression of a dominant-negative mutant of TβRII inhibited NTHi- induced TLR2 expression at mRNA level, whereas overexpression of a wild-type TβRII enhanced NTHi-induced TLR2 expression at mRNA level in HeLa cells. C, transfection with TβRI-siRNA (100nM) attenuated NTHi-induced TLR2 expression at mRNA level in HeLa cells.
D, transfection with TβRII-siRNA (100nM) attenuated NTHi-induced TLR2 expression at mRNA level in HeLa cells. Quantitative real-time PCR were performed to assess TLR2 mRNA expression level after infection with NTHi for 3 h. Values are the mean + S.D.; n = 3.
Figure 23 TβR signaling is a positive regulator for NTHi-induced TLR2 expression. A, NTHi induced TLR2 transcription in wild type Mv1Lu cells expressing TβRI and TβRII but not in mutant R1B and DR26 cells lacking functional TβRI and TβRII, respectively. A TLR2-luc or SBE-luc reporter vector was transfected into Mv1lu, R1B or DR26 cells. NTHi or recombinant human TGF-β1 was then added to the transfected cells 42 h after transfection. After 5 h, the cells were harvested for luciferase assay. B, overexpression of a dominant-negative mutant of TβRI or TβRII inhibited NTHi-induced TLR2 transcription in wild type Mv1Lu cells, whereas
第二項 NTHi による TLR2 発現誘導を正に制御する TβR 下流 signaling の検討
TβRI/II signaling の下流因子として Smad が重要であることが知られているため,次
に receptor-regulated Smad である Smad3 及び common-partner Smad である Smad4 が
NTHi による TLR2 発現誘導の制御に関与しているか検討した.Figure 24A に示したよ
うに,NTHi による TLR2 発現誘導は Smad3,Smad4 の dominant negative 変異体の強制 発現により阻害され, Smad3,Smad4 の強制発現により促進された.したがって,
Smad3/4 は NTHi による TLR2 発現誘導を正に制御していると考えられる.Smad3/4 の 関与をさらに確認するために,Smad4 の発現が検出されない MDA-MB468(MB468)
細胞54 – 56) を用いて,Q-PCR 及び promoter assay を行った.その結果,mRNA レベルだ
けでなく転写レベルにおいても,NTHi による TLR2 の発現誘導は Smad4 の強制発現 により促進された(Figure 24B&C ).以上より, TβRI/II-Smad3/4 signaling は NTHi に よる TLR2 の発現誘導を正に制御することが示された.
Figure 24 TβR signaling is positively regulates NTHi-induced TLR2 expression via a Smad3/4-dependent mechanism. A, overexpression of dominant-negative mutants of Smad3 and Smad4 attenuated NTHi-induced TLR2 expression at mRNA level, whereas overexpression of wild-type Smad3 and Smad4 enhanced NTHi-induced TLR2 expression at mRNA level in HeLa cells. B, overexpression of a wild-type Smad4 enhanced NTHI-induced TLR2 expression at mRNA level in MDA-MB468 cells. Quantitative real-time PCR were performed to assess TLR2 mRNA expression level after infection with NTHi for 3h. C, overexpression of a wild- type Smad4 enhanced NTHi-induced TLR2 transcription in MDA-MB468 cells. Values are the mean + S.D.; n=3.
第三項 NTHi による TβR 活性化機構の検討
これまで TβRI/II-Smad3/4 signaling pathway が NTHi による TLR2 発現誘導を正に制 御することが明らかとされた.しかしながら, NTHi が TβRI/II-Smad3/4 signaling pathway をどのような機序で活性化するかについては未だ明らかとなっていない.そこ で,そのメカニズムを解明するために,NTHi が TGF-β1 と同様に TβRI/II-Smad3/4 signaling を活性化するか,Western Blotting と immunofluorescent staining を用いて検討 した.その結果,TGF-β1 と同様に,NTHi の処理により Smad3 のリン酸化が検出され,
リン酸化された Smad3,Smad4 の核移行が誘導された(Figure 25A).次に,NTHi に よって誘導された Smad3/4 の核移行が TGF-β1-Smad 依存的な転写活性に関与している か,promoter assay により検討した.SBE 依存的な転写活性については SBE-luciferase を,TGF-β1 依存的な転写活性については plasminogen activator inhibitor-1(PAI-1)- luciferase57) を用いた結果,TGF-β1 と同様に,NTHi 処理は各々の reporter 遺伝子の転 写活性を増加させた(Figure 25B).したがって,NTHi は TβRI/II-Smad3/4 signaling を 活性化し,その転写活性に関与していることが示された.しかしながら,どのように して NTHi が TβRI/II-Smad3/4 signaling を活性化し,NTHi による TLR2 の発現誘導を 制御しているかは明らかとなっていない.そこで,まず NTHi 処理によって TGF-βs の 発現が誘導された可能性について検討した.TGF-βs の Q-PCR 及び ELISA assay を行っ た結果,NTHi 処理により TGF-βs の発現が誘導されないこと,NTHi 処理による培養 溶液中の TGF-βs タンパク質発現量は変化しないことが示された(Figure 26A & B).
そのため,TβRI/II-Smad3/4 signaling を介した NTHi による TLR2 の発現誘導の制御は,
過去の報告通り NTHi 由来の TGF-β-like factor によるものではないかと仮説をたてた.
この仮説を証明するために,TGF-β neutralization assay を用いて NTHi による TLR2 の 発現誘導の変化について検討した. Figure 27A に示したように,TGF-β 中和抗体で TGF-β 及び NTHi を前処理した結果,TGF-β による PAI-1 発現誘導及び NTHi による TLR2 発 現 誘 導 は 阻 害 さ れ た . ま た ,NTHi 抽 出 液 を TGF-β 抗 体 に よ り
immunodepletion した溶液で処理した細胞においても,NTHi による TLR2 の転写活性
の増加は阻害されていた(Figure 27B).したがって,NTHi 由来の TGF-β-like factor
Figure 25 NTHi activates TβRI/II-Smad3/4 signaling like TGF-β1 does. A, treatment with NTHi or TGF-β1 (1 ng/ml) for 5 min induced phosphorylation of Smad3 in HeLa cells, as assessed by Western blotting using anti-phosphorylated Smad3 antibody (upper panel). In addition, NTHi and TGF-β1 also induced nuclear translocation of phospho-Smad3, Smad3 and Smad4 (lower panel). Representative fields of phospho-Smad3, Smad3 and Smad4 fluorescence (lower panel) were shown in HeLa cells that were treated with NTHi or TGF-β1 (1 ng/ml) for 60 min, respectively. B, NTHi and TGF-β1 (1 ng/ml) induced Smad-regulated promoter activity of SBE-luc and TGF-β-responsive promoter activity of PAI-1-luc in HeLa cells. Values are the mean + S.D.; n=3.
Figure 26 NTHi does not induce TGF-βs mRNA and protein. A, NTHi did not induce TGF- β1, 2 and 3 expression at mRNA level in HeLa cells, as assessed by performing quantitative real-time PCR analysis. B, NTHi did not induce any detectable increase in TGF-β1, 2 and 3 in the conditioned media of HeLa cells as assessed by TGF-β ELISA assay. Values are the mean + S.D.; n=3.
Figure 27 NTHi activates TβR-Smad signaling likely via an autocrine-independent mechanism. A, preteatment of NTHi or TGF-β1 (1 ng/ml) with the TGF-β neutralizing antibody (1 µg/ml) reduced its ability to induce TLR2 or PAI-1 expression in HeLa cells at mRNA level, respectively. B, TLR2 transcription was inhibited in HeLa cells by the NTHi lysates, in which the TGFβ1-like factors were immunodepleted using anti-TGF-β1 antibody.
Values are the mean + S.D.; n=3.
第四項 NTHi による TLR2 発現誘導を正に制御する TβR-Smad3/4 signaling に対
するNF-κBの関与の検討
これまでに, NF-κB の活性化は NTHi による TLR2 の発現誘導を正に制御すること,
TβRI/II-Smad3/4 signaling は NF-κB の活性化に関与していることが報告されている58) . そこで, TβRI/II-Smad3/4 signaling を介した NTHi による TLR2 発現誘導の増加には NF-κB の活性化が関与しているのではないかという仮説を立てた.この仮説を証明す るために,まず TβRI/II-Smad3/4 signaling が NF-κB の活性化に関与していることを確 認した. NF-κB-luciferase の reporter 遺伝子を用いた promoter assay の結果,NTHi によ る NF-κB の活性化は,TβRII,Smad3,Smad4 の dominant negative 変異体の強制発現に より阻害された(Figure 28A).また,MB468 細胞への Smad4 の強制発現は,NTHi による NF-κB の活性化を促進した(Figure 28B).したがって, TβRI/II-Smad3/4
signaling は NTHi による NF-κB の活性化に関与していることが示された.そこで,次
に TβRI/II-Smad3/4 signaling の活性化が NTHi に よるTLR2 promoter 及び NF-κB
promoter の転写活性の増強に寄与するか,promoter assay により検討した.その結果,
TGF-β1 処理,または Smad3,Smad4 の強制発現により活性化された TβRI/II-Smad3/4 signaling は,NTHi による NF-κB 及び TLR2 の転写の活性化をより増強した(Figure 28C).以上のことより, TβRI/II-Smad3/4 signaling pathway は NF-κB と協調して TLR2 の転写に関与していることが示された.
Figure 28 Functional cooperation of Smad3/4 with NF-κB appears to mediate NF-κB- dependent TLR2 transcription. A, overexpression of dominant-negative mutants of TβRII, Smad3 and Smad4 inhibited NTHi-induced NF-κB activation. B, overexpression of a wild-type Smad4 enhanced NTHi-induced NF-κB activation in MDA-MB468 cells. C, activation of TGF- β signaling by TGF-β1 (1 ng/ml) or overexpression of wild-type Smad3 and Smad4 markedly
第五項 NTHi による TLR2 発現誘導を正に制御する TβR-Smad3/4 signaling に対す る p38 MAPK signaling の関与の検討
多くの細胞ストレスが NF-κB だけでなく p38 MAPK も活性化する59 - 61) .NTHi も例 外ではなく,p38 MAPK signaling pathway を活性化し,NTHi による TLR2 の発現誘導 を負に制御することが報告されている.興味深いことに,TβRI/II-Smad3/4 signaling は このNTHi による p38 MAPK の活性化を抑制することが報告されている47) ことから,
TβRI/II-Smad3/4 signaling は NTHi による p38 MAPK signaling pathway の活性化を抑制 することにより,TLR2 の発現を誘導する可能性がある.そこでこの可能性を検討する ため,TβRI/II-Smad3/4 signaling の阻害が,NTHi による p38 MAPK signaling pathway 活 性化に影響を与えるか,Western Blotting を用いて確認した.Figure 29 A~E に示したよ うに,SB431542 の前処理,TβRI,TβRII,Smad3,Smad4 の dominant negative 変異体 の強制発現,siRNA による TβRI 及び TβRII の knockdown による TβR signaling の阻害
は,NTHi によって誘導される p38 のリン酸化を著しく促進したが,その上流分子であ
る MKK3/6 のリン酸化は促進しなかった.これらの結果と一致して,NTHi と TGF-β1
の同時処理は NTHi による p38 のリン酸化を促進したが,MKK3/6 のリン酸化に変化 はみられなかった(Figure 29F).以上のことより,TβRI/II-Smad3/4 signaling は NTHi による p38 MAPK の活性化を抑制することにより,TLR2 の発現誘導を正に制御する ことが示唆された.
Figure 29 TβR-Smad3/4 signaling pathway positively mediates NTHi-induced TLR2 up- regulation via a negative cross-talk with p38 MAPK. A, pretreatment with SB431542 (1 µM) enhanced NTHi-induced phosphorylation of p38, but not MKK3/6, in HeLa cells, as assessed by Western blotting using anti-phosphorylated p38 and MKK3/6 antibodies. B, overexpression of a dominant-negative mutant of TβRI enhanced NTHi-induced phosphorylation of p38, but not MKK3/6, in HeLa cells. C, overexpression of a dominant-negative mutant of TβRII enhanced NTHi-induced phosphorylation of p38, but not MKK3/6, in HeLa cells. D, transfection with TβRI-siRNA (100nM) and TβRII-siRNA (100nM) enhanced NTHi-induced
第六項 NTHi による TLR2 発現誘導を正に制御する TβR-Smad3/4 signaling に対す る p38 MAPK 抑制因子,MKP-1 の関与の検討
Figure 29 において,TβRI/II-Smad3/4 signaling は NTHi による p38 MAPK の活性化を 抑制することにより,TLR2 の発現誘導を正に制御することが示唆された.しかしなが ら,TβRI/II-Smad3/4 signaling がどのようにして NTHi による p38 MAPK の活性化を抑 制するかについては明らかとなっていない.TβRI/II-Smad3/4 signaling の活性化は様々 な転写因子,転写調節因子とともに,標的遺伝子の転写を調節することから,NTHi に よる TβRI/II-Smad3/4 signaling の活性化が p38 MAPK を阻害するような分子の発現を誘 導するのではないかと考えた.この可能性を検討するために,タンパク質合成阻害剤 である cycloheximide 処理を行った結果,NTHi による TLR2 発現誘導は阻害された
(Figure 30).したがって,TβRI/II-Smad3/4 signaling を介した NTHi による TLR2 発 現誘導の制御には,新たなタンパク質の発現が関与していることが示唆された.
Figure 30 TLR2 expression enhancement by inhibiting NTHi-induced p38 MAPK activation via TβRI/II signaling involves de novo protein synthesis. Pretreatment with cycloheximide (20 µM), a protein synthesis inhibitor, inhibited NTHi-induced TLR2 expression at mRNA level in HeLa cells. Quantitative real-time PCR was performed to assess TLR2 mRNA expression level after infection with NTHi for 3 h. Values are the mean + S.D.; n=3.
そこで,p38 MAPK の活性化を脱リン酸化により阻害する dual specificity phosphatase である MAPK phosphatase-1(MKP-1)に着目した62 - 64) .MKP-1 の発現が,NTHi 及び TGF-β1 処 理 によっ て同様 に 誘導 され たた め (Figure 31A) ,こ の 発現 誘 導 に は TβRI/II-Smad3/4 signaling が関与しているか次に検討した.Figure 31B~F に示したよう に,SB431542 の前処理,TβRI,TβRII,Smad3,Smad4 の dominant negative 変異体の 強制発現による TβRI/II-Smad3/4 signaling の阻害は,NTHi による MKP-1 発現誘導を阻
害した.したがって,NTHi は p38 MAPK の活性化を制御する MKP-1 の発現を TβRI/II-Smad3/4 signaling を介して誘導していることが示された.
Figure 31 TβR-Smad3/4 signaling pathway is involved in NTHi-induced MKP-1 expression. A, NTHi and TGF-β1 (1 ng/ml) induced MKP-1 expression at the protein level in HeLa cells, as assessed by Western blotting using anti-MKP-1 antibody. B, pretreatment with SB431542 (1, 10 µM) and overexpression of dominant-negative mutants of TβRI, TβRII, Smad3 and Smad4 inhibited NTHi-induced MKP-1 expression at the protein level in HeLa cells.
In experiments, cells were treated with or without NTHi for 60 min.
NTHi により MKP-1 の発現が誘導されたため,Figure 29 で示された p38 MAPK の活
性化抑制によるTLR2 の発現増加は,MKP-1 に起因すると考えられる.そこで,NTHi によって誘導される MKP-1 が p38 MAPK の脱リン酸化により NTHi による TLR2 発現 誘導を正に制御しているか,MKP-1 を阻害することにより検討した.Q-PCR の結果,
MKP-1 の発現阻害剤である Ro31-8220 の前処理は NTHi による TLR2 の発現誘導を抑
た結果と一致して,Ro31-8220 の前処理により,NTHi による TLR2 タンパク質発現誘 導の阻害が,Western Blotting によっても確認された(Figure 32D).以上より,NTHi によって誘導される MKP-1 が p38 MAPK の脱リン酸化により NTHi による TLR2 発現 誘導を正に制御していることが示された.
Figure 32 TβR-Smad3/4 signaling pathway positively regulates NTHi-induced TLR2 expression via MKP-1-dependent inhibition of p38 MAPK. A, pretreatment with Ro-31- 8220 (2 µM), an inhibitor for MKP-1 expression, inhibited NTHi-induced TLR2 expression at mRNA level in HeLa cells. B, overexpression of an antisense MKP-1 construct attenuated NTHi-induced TLR2 expression at mRNA level, whereas overexpression of a wild-type MKP-1 enhanced NTHi-induced TLR2 expression at mRNA level in HeLa cells. C, transfection with MKP-1-siRNA (100nM) attenuated NTHi-induced TLR2 expression at mRNA level and ennanced NTHi-induced p38 phosphorylation in HeLa cells. D, pretreatment with Ro-31-8220 (2 µM) inhibited NTHi-induced TLR2 expression at the protein level in HeLa cells. Values are the mean + S.D.; n=3.
第七項 in vivo での NTHi による TLR2 発現誘導に対する TβR signaling の影響
NTHi 由来の TGF-β-like factor によるTβR の活性化は,NTHi による TLR2 発現誘導 を正に制御すること,また,そのメカニズムには NF-κB の活性化とMKP-1 による p38
MAPK の不活性化という二つの signaling pathway が関与していることが in vitro におい
て明らかとなった.そこで,このシグナル伝達機構が in vivo においても NTHi による
TLR2 発現誘導の制御に関与しているか検討した.その結果,NTHi による TLR2 発現
誘導は SB431542 及び Ro31-8220 の前処理により阻害された(Figure 33A & B).した がって,in vivo においても TβRI/II-Smad3/4 signaling pathway が NTHi による TLR2 発 現誘導を正に制御することが確認された.
Figure 33 TβR-Smad3/4 signaling pathway positively regulates NTHi-induced TLR2 expression in vivo. A, pretreatment with SB431542 for 30 min reduced TLR2 induction by NTHi at mRNA level in the lung of BALB/c mice. B, pretreatment with Ro-31-8220 for 30 min reduced TLR2 induction by NTHi at mRNA level in the lung of BALB/c mice. Quantitative real-time PCR was performed to assess TLR2 mRNA expression level. Values are the mean + S.D.; n = 3.
第三節 考察
本章において,NTHi 由来の TGF-β-like factor による TβRI/II-Smad3/4 signaling
pathway の活性化は,NTHi による TLR2 発現誘導を正に制御すること,また,そのメ
カニズムには NF-κB の活性化及び MKP-1 による p38 MAPK の不活性化という二つの
signaling pathway が関与していることが明らかとなった.本章で得られた知見を Figure
34 に示す.
NTHi
TLR2 Gene Expression p38!/"P
T"R TLR
-
Smad4 Smad3 Smad3P
Smad3
P Smad4
MKP-1 Expression -
IKK"/#
I$B!
I$B!
p50 p65 I$B!
p50 P
+ p65
Host Defense and Immune Responses
Figure 34 Schematic representation of the signaling pathway involved in the positive regulation of NTHi-induced TLR2 expression. As indicated, activation of TβRI/II-smad3/4 signaling acts as a positive regulator of NTHi-induced TLR2 expression via NF-κB pathway and MKP-1-dependent inhibition of p38 MAPK in vitro and in vivo. TβR, Transforming growth factor-β receptor; NTHi, Nontypeable Haemophilus influenzae; MAP kinase, mitogen-activated protein kinase; MKK, MAP kinase kinase; TLR, Toll-like receptor.